|
|
||||||||||||||||||||||||
|
生境简化重塑榕树-榕小蜂互作网络: 挥发物组成与群落结构的适应性响应
生物多样性
2026, 34 (2):
25328-.
DOI: 10.17520/biods.2025328
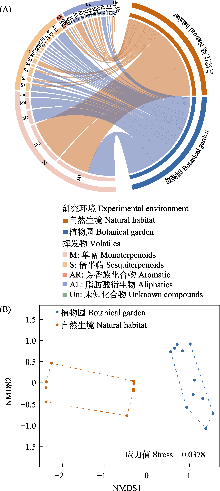
植物园(botanical garden)作为相对均质化的人工环境, 为研究植物-传粉者互作的环境适应机制提供了理想模型。榕树(Ficus spp.)与榕小蜂构成的专性互惠系统高度依赖雌花期榕果挥发物的化学通讯。榕树作为重要的观赏和生态树种, 常被引种至植物园、公园等人工环境进行异地保存。然而, 引种后榕树挥发物组成及榕小蜂群落结构是否会发生适应性变化尚不明确, 这种生境转换是否影响其互作稳定性仍待探究。本研究以自然生境和植物园中的大果榕(Ficus auriculata)为研究对象, 采用动态顶空采样结合气相色谱-质谱联用仪(GC-MS)分析雌花期榕果挥发物, 并通过雄果榕小蜂群落调查, 比较两种生境下榕蜂互作的适应性差异。研究结果表明: (1)挥发物组成显著分化, 自然生境以防御相关的脂肪酸衍生物为主, 而植物园中以吸引传粉蜂的萜烯类化合物为主; (2)榕小蜂群落结构差异显著, 自然生境多样性更高, 非传粉蜂数量显著高于植物园; 而植物园中传粉蜂占比99.07%, 远高于自然生境(76.55%); (3)自然生境榕-蜂互作网络具有更高的连接度、模块性和稳健性, 表明自然生境榕蜂关系更复杂紧密, 同时模块间具有高独立性, 使自然生境榕小蜂群落更加适应外界干扰, 对环境变化有较强的抵抗力。而植物园则表现出更强的专化性和嵌套性, 由于环境条件均一化使榕小蜂群落对环境变化敏感性增加。本研究首次系统揭示了榕树迁地保护至植物园后挥发物-榕小蜂互作网络的适应性调整机制, 证实生境简化会通过改变化学信号和生态位分配重塑共生关系。成果不仅为理解专性共生系统的环境适应性提供了理论基础, 也为榕树迁地保护及生物多样性维持策略提供科学依据。
表2
自然生境和植物园榕果-榕小蜂互作网络的网络参数观测值(重抽样均值 ± 标准差)
正文中引用本图/表的段落
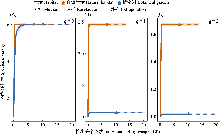
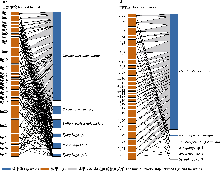
使用bipartite包绘制榕果-榕小蜂二分网络以分析榕小蜂群落分布状况。计算下列网络指标(刘灿然, 2002), 具体的计算方法可参考冯志荣等(2023): (1)网络连接度(connectance, C), 表示榕小蜂在榕果中的存在情况, 值越大表示榕小蜂在榕果中的存在越普遍, 表征榕小蜂物种与榕果资源的连接复杂度; (2)互作专化度(specialization, H2′), 表示一种榕树上所有榕小蜂物种对榕果资源利用的整体特异性, 值越大表示榕小蜂利用的榕果越多, 代表一个群落中的榕小蜂物种对榕果资源利用的整体专一化程度; (3)加权嵌套性(weighted nestedness metric based on overlap and decreasing fill, WNODF), 表示对榕小蜂物种丰度的加权, 代表了榕小蜂物种对榕果资源利用的嵌套程度; (4)模块性(modularity, Q)和稳健性(robustness, R), 表示去掉榕果资源后, 榕小蜂群落的稳健程度, 表征榕小蜂群落的稳定性。为比较不同生境中互作网络的差异, 采用自展法(bootstrapping), 对每个网络进行20次重抽样以构建新的网络, 计算网络参数, 对各参数进行999次置换检验(permutation t-test), 评估两种生境下榕果-榕小蜂二分网网络指标的差异(Wang et al., 2022)。将榕果看成一个近似椭球体, 计算榕果内部体积, 用Hmisc包对榕果内部体积与榕小蜂丰富度进行Pearson相关性分析。两种生境下榕小蜂群落组成差异的分析方法与1.4.1挥发物的分析一致。
通过构建榕果-榕小蜂互作网络显示, 自然生境的互作网络复杂度显著高于植物园(图5), 具体表现为: 传粉蜂在两种生境的榕果中均占绝对优势, 但自然生境中非传粉蜂数量及其与榕果的互作关系更丰富。网络拓扑特征存在显著差异(P < 0.05) (表2): 自然生境具有更高的连接度(0.87 vs. 0.31)、模块性(0.14 vs. 0.01)和稳健性(0.75 vs. 0.49), 而植物园则表现出更强的互作专化度(0.36 vs. 0.21)和加权嵌套性(46.81 vs. 35.92)。这些结果共同表明, 生境简化不仅降低了榕小蜂群落的物种多样性, 更显著削弱了榕果-榕小蜂互作网络的结构复杂性和生态稳定性。
本文的其它图/表
|
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()