
|
|
||
|
青藏高原盆栽一年生和多年生豆科牧草对土壤线虫群落的影响
生物多样性
2024, 32 (7):
24040-.
DOI: 10.17520/biods.2024040
查明一年生及多年生豆科牧草对土壤线虫群落的影响, 可为青藏高原地区科学种植豆科牧草提供技术支撑。作者于2020-2022年在川西北红原县盆栽种植一年生光叶紫花苕(Vicia villosa, VV)和多年生紫花苜蓿(Medicago sativa, MS), 以天然草地的植物作对照(control, CK)。每年9月下旬对土壤线虫组成、密度、多样性和营养类群, 植物的高度、盖度、生物量及土壤理化性质进行调查。结果表明: (1) 3种处理间的线虫群落组成结构存在差异, 并随年际变化而增大; (2)光叶紫花苕的土壤线虫群落密度显著高于紫花苜蓿和对照, 紫花苜蓿的线虫类群数和Shannon-Wiener指数显著高于光叶紫花苕和对照; 随着年际变化, 光叶紫花苕的线虫类群数及Shannon-Wiener指数、紫花苜蓿的线虫密度及Shannon-Wiener指数均显著下降; (3)光叶紫花苕的食真菌和植物寄生线虫密度显著高于紫花苜蓿和对照, 捕食杂食线虫相对密度显著低于紫花苜蓿和对照; 紫花苜蓿的植物寄生线虫相对密度显著低于光叶紫花苕和对照; 随年际变化, 光叶紫花苕的食细菌和植物寄生线虫相对密度分别显著下降和上升, 紫花苜蓿和对照的食细菌线虫和食真菌线虫密度及食细菌线虫相对密度显著下降, 捕食杂食线虫相对密度显著上升; (4)光叶紫花苕线虫群落的基础指数、结构指数和通道指数显著高于紫花苜蓿; (5)影响土壤线虫群落的主要环境因子主要为土壤pH、有效钾和植物群落高度、盖度和地上生物量。研究结果表明, 一年生和多年生豆科牧草分别有利于维持较高的线虫群落密度和多样性, 二者间作可能有利于提高线虫群落密度、多样性及生态功能。 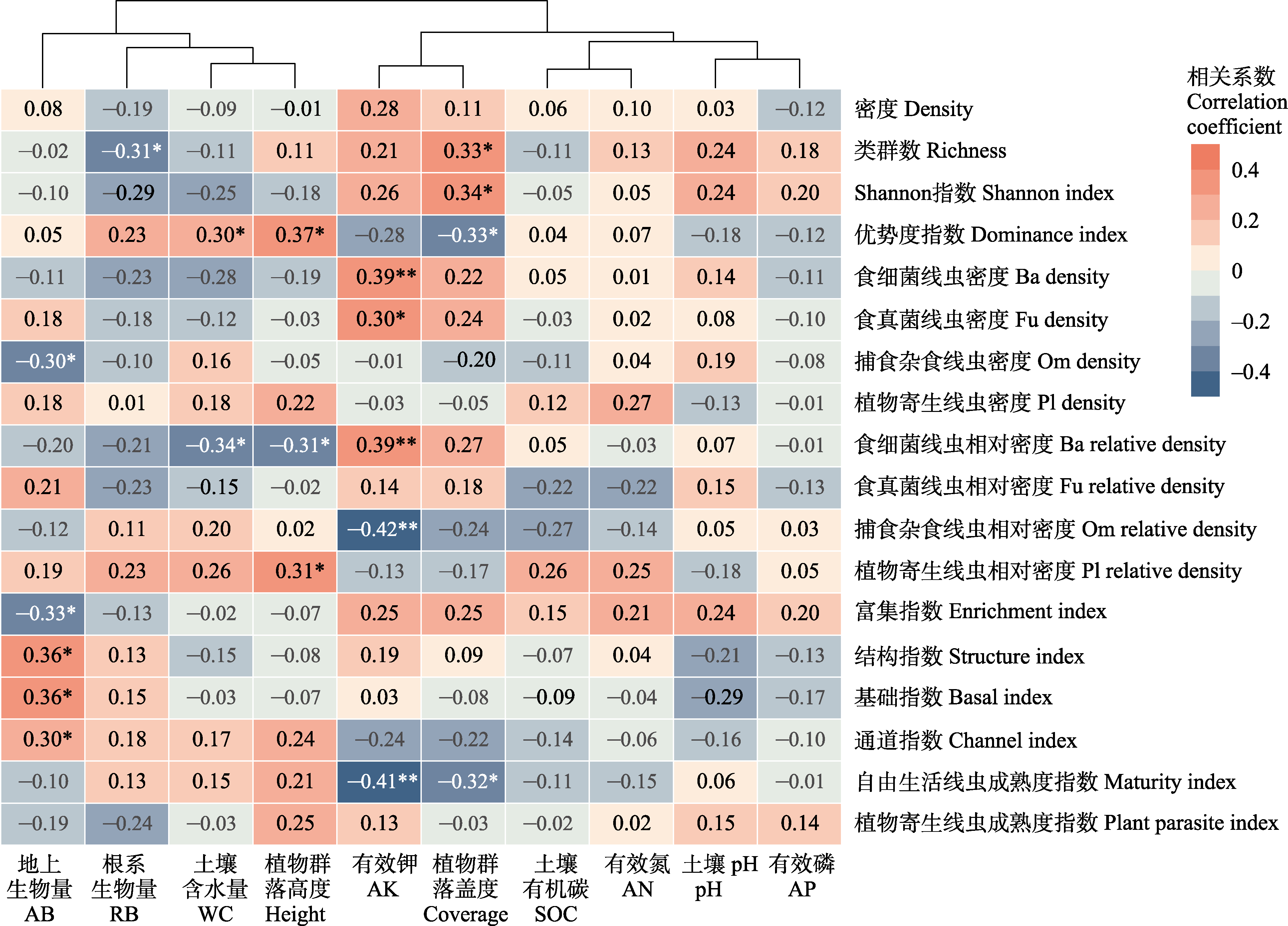 View image in article
图6
土壤线虫群落各指数与环境因子相关分析热图。Ba: 食细菌线虫; Fu: 食真菌线虫; Om: 捕食杂食线虫; Pl: 植物寄生线虫。* P < 0.05; ** P < 0.01。
正文中引用本图/表的段落
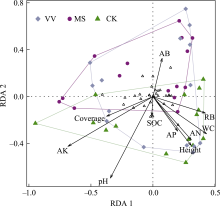
土壤线虫群落各指数与环境因子相关分析如图6所示。类群数与植物群落盖度显著正相关, 与根系生物量显著负相关(P < 0.05); Shannon-Wiener指数与植物群落盖度显著正相关(P < 0.05); 优势度指数与盖度显著负相关(P < 0.01), 与植物群落高度和土壤含水量显著正相关(P < 0.05)。食细菌线虫与食真菌线虫密度与有效钾显著正相关(P < 0.01), 捕食杂食线虫与植物地上生物量显著负相关(P < 0.05)。食细菌线虫相对密度与土壤含水量和植物群落高度显著负相关(P < 0.05), 与有效钾显著正相关(P < 0.01), 捕食杂食线虫相对密度与有效钾显著负相关(P < 0.01), 植物寄生线虫相对密度与植物群落高度显著正相关(P < 0.05)。结构指数、基础指数、通道指数都与植物地上生物量显著正相关(P < 0.05), 富集指数与植物地上生物量显著负相关(P < 0.05), 成熟度指数与有效钾(P < 0.01)、植物群落盖度(P < 0.05)呈显著负相关。总体上, 有效钾、植物地上生物量和植物群落盖度是影响土壤线虫群落各指数的最重要环境因子。
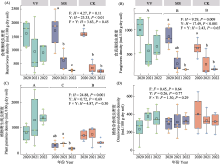
本研究发现, 光叶紫花苕、紫花苜蓿、对照间土壤线虫群落组成结构存在明显差异, 并随建植年限增加而增大。天然高寒草地转变为人工草地后, 植物群落组成改变、多样性降低。植物群落变化又可以影响土壤微生物群落结构(于皓等, 2023)、湿度、温度及养分等(薛会英等, 2013), 这些因素变化均可以直接或间接地影响土壤线虫群落。杨锐等(2020)研究发现, 天然草地转变为燕麦(Avena sativa)地后, 土壤线虫群落结构发生明显改变, 密度显著增加。此外, 植物种类不同也会对土壤线虫群落产生较大影响(De Deyn et al, 2004; Bengtsson et al, 2009)。魏雪等(2022)研究发现, 土壤线虫群落结构在6种单播人工草地间存在差异, 苜蓿人工草地线虫群落密度低于其他人工草地。已有研究表明不同植物的凋落物和根系的营养物质含量及根系分泌物的差异均可影响土壤动物群落(Zhao et al, 2021; 马金豪等, 2023)。土壤理化性质对线虫群落也有重要影响(Zhao et al, 2014)。本研究中, 有效钾与食细菌、食真菌线虫密度以及食细菌线虫相对密度显著正相关(图6)。在本区域的前期相关研究也发现这一现象(杨锐等, 2020; 魏雪等, 2022)。土壤钾能提高食微线虫密度, 可能是由于土壤养分变化可以影响植物生长及营养物质含量, 从而间接影响土壤线虫群落结构组成(Zhao et al, 2014)。已有研究表明, 在青藏高原地区施钾肥可以提高植物生物量及蛋白质含量(刘文辉等, 2010), 有利于微生物生长, 间接促进食微线虫密度。此外, 食细菌线虫相对密度与土壤含水量和植物高度呈显著负相关, 有研究表明土壤干燥时, 相对易于形成较大的土壤团聚体, 可为微生物提供更丰富的资源(Jiang et al, 2015), 从而间接影响食细菌线虫密度。有效钾与捕食杂食线虫相对密度呈显著负相关, 可能是因为有效钾能够提高食微线虫密度导致。另外, 相对于其他营养类群, 捕食杂食线虫密度在不同处理和年际间均无显著变化, 说明捕食杂食线虫是最不敏感的类群。其他研究也发现低营养级线虫密度比高营养级线虫密度更易受植物种类变化的影响(Wardle et al, 2003; De Deyn et al, 2004), 可能是因为捕食杂食线虫的营养级较高, 不直接取食植物。
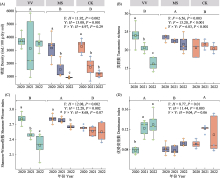
光叶紫花苕的类群数和Shannon-Wiener指数显著低于紫花苜蓿, 主要是光叶紫花苕处理的线虫类群数随种植年限增加而显著降低, 而紫花苜蓿处理的线虫类群数相对稳定。线虫组成类群的变化可能与不同植物种类输入地下的资源的数量与质量不同有关(Wardle et al, 2004)。本研究中, 线虫类群数与根系生物量呈显著负相关(图6), 而在3种处理中紫花苜蓿的根系生物量相对最低(附录5)。此外, 相关分析表明, 植物群落盖度与类群数、Shannon- Wiener指数呈显著正相关, 与优势度指数呈显著负相关, 而植物高度则与优势度指数呈显著正相关。本研究中紫花苜蓿为直立型上繁草, 光叶紫花苕为匍匐型上繁草, 不同株丛形状可影响植物群落盖度(薛会英等, 2013), 从而影响线虫群落的光、湿度等微环境。已有研究也表明植株性状特征与土壤生物之间存在密切关系(Mariotte et al, 2018)。因此, 不同牧草的生理及植株形态特征差异可能是造成线虫群落多样性差异的原因。
本研究发现一年生和多年生牧草的生长年限对线虫群落影响不同。随着年际变化, 光叶紫花苕的类群数和Shannon-Wiener指数显著下降, 密度无显著变化(图2)。主要是因为光叶紫花苕处理中一些线虫稀有类群逐渐消失, 而矮化属(Pl3)、拟盘旋属(Pl3)等植物寄生线虫密度显著增加。紫花苜蓿和对照的线虫密度随着种植年限增加显著下降, 而类群数和Shannon-Wiener指数相对稳定(图2), 主要是因小杆属(Ba1)、广杆属(Ba1)、巴氏属(Fu3)和原细齿属(Fu4)等食细菌、食真菌线虫密度普遍下降。造成这种差异的原因可能是因为一年生光叶紫花苕对土壤的物质归还较多, 为植物寄生线虫提供了丰富的食物资源, 有利于植物寄生线虫的生存。而多年生紫花苜蓿和对照的食真菌和食细菌线虫密度显著下降, 可能是因为花盆土壤有效钾等养分含量随着种植年限增加下降, 而有效钾含量与食细菌和食真菌线虫密度呈显著正相关(图6)。
本文的其它图/表
|


{kind=link}