中国中亚热带常绿阔叶林群落木本植物多样性比较
王晓凤, 米湘成, 王希华, 江明喜, 杨涛, 张健, 沈泽昊
生物多样性
2023, 31 ( 11):
23296-.
DOI: 10.17520/biods.2023296
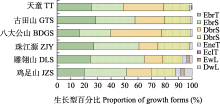
常绿阔叶林是我国亚热带的地带性植被类型, 也是我国最重要的植被类型之一。常绿阔叶林植被型下各种群落类型的物种组成、结构、动态等特征已有大量研究, 本研究基于浙江的天童和古田山、湖南的八大公山以及云南的珠江源、雕翎山和鸡足山6个大型常绿阔叶林动态监测样地的调查数据, 对其木本植物的物种组成、区系成分、生长型类型百分比、种-面积关系进行了综合比较, 以期进一步了解中国中亚热带常绿阔叶林在东、中、西部地区的物种多样性差异。结果显示: (1)中国中亚热带中部的八大公山样地物种的科、属、种丰富度最高, 其次为东部的天童和古田山样地, 而西部的珠江源、雕翎山和鸡足山样地的丰富度最低, 相同植被类型样地的物种组成相似性最高, 东、中、西部的常绿阔叶林植被的物种组成存在显著的差异; (2)东部样地木本植物的科、属均以热带区系为主, 尤其泛热带分布的科、属最多, 而中、西部样地虽然以热带科为主, 但以温带属居多, 北温带分布属最多; (3)东、中部3个样地木本植物以乔木成分为主, 天童、古田山和八大公山样地乔木种比例分别是57.4%、51.6%和55.9%, 西部样地灌木(珠江源、雕翎山和鸡足山样地分别是56.9%、54.2%、48.5%)成分均高于乔木(43.1%、41.7%、42.7%), 中部样地木本植物落叶种(59.2%)占比远高于常绿种(40.8%), 东、西部样地则是常绿种比例高于落叶种, 天童、古田山、珠江源、雕翎山、鸡足山样地常绿种比例分别是51.3%、59.1%、62.7%、69.8%、58.3%; (4)八大公山和雕翎山样地的常绿种(分别有97种和67种)和落叶种(141种, 29种)在数量上差距最大, 雕翎山和珠江源样地的常绿种(69.8%, 62.7%)和落叶种(30.2%, 37.3%)在比例上差距最大。上述研究结果反映了中国中亚热带典型常绿阔叶林、山地常绿落叶阔叶混交林、半湿润常绿阔叶林的群落物种组成、物种多样性和群落结构的差异, 为指示中亚热带常绿阔叶林植被从东到西的物种周转特征和植被变化规律提供了科学依据。
自然保护区(风景区) Nature reserve
(scenic spot) | 天童国家森林公园 Tiantong National Forest Park | 古田山国家级自然保护区 Gutian Mountains National
Nature Reserve | 八大公山国家级自然保护区 Badagongshan Mountains National Nature Reserve | 珠江源省级自然保护区 Zhujiangyuan Provincial Nature Reserve | 雕翎山省级自然保护区 Diaoling Mountains Provincial Nature Reserve | 鸡足山风景名胜区
Jizu Mountains Scenic Spot | | 面积 Area (ha) | 349 | 8,107 | 20,000 | 117,937 | 613 | 6,165 | | 海拔 Altitude (m) | 300-653.3 | 500-1,258 | 1,000-1,890.4 | 2,158-2,444 | 2,040-2,593.6 | 1,780-3,248 | | 气候类型 Climate type | 亚热带季风气候
Subtropical monsoon climate | 亚热带季风气候
Subtropical monsoon climate | 山地湿润季风气候
Mountain humid monsoon climate | 中亚热带高原季风气候 Mid-subtropical plateau monsoon climate | 中亚热带高原季风气候 Mid-subtropical plateau monsoon climate | 中亚热带高原季风气候 Mid-subtropical plateau monsoon climate | | 年均温 Mean annual temperature, MAT (℃) | 16.2 | 15.3 | 11.5 | 18.1 | 19.2 | 18.5 | | 最热月均温 Mean temperature of hottest month, MTHM (℃) | 28.1 | 28.9 | 22.8 | 22.3 | 23.0 | 22.5 | | 最冷月均温 Mean temperature of coldest month, MTCM (℃) | 4.1 | 4.1 | 0.1 | 11.7 | 13.7 | 12.8 | 年均降水量
Mean annual precipitation, MAP (mm) | 1,551 | 1,963.7 | 2,105.4 | 963.5 | 845 | 825.2 | | 年均雨日 Mean annual rainy days, MARD (d) | - | 142.5 | 176 | - | - | - | | 年均无霜期 Mean annual frost free days, MAFD (d) | 237.8 | 250 | 190 | - | 250 | 220 | 年均日照总时数
Mean annual total sunshine hours, MATSH (h) | - | 1,334.1 | - | 2,108.2 | - | 2,063.3 | | 平均相对湿度 Mean relative humidity, MRH (%) | 85 | 92.4 | 90 | - | 75 | - | | 种子植物科数 Families number of seed plants | 148 | 149 | 165 | 150 | 112 | 163 | | 种子植物属数 Genera number of seed plants | 506 | 648 | 709 | 461 | 355 | 447 | | 种子植物种数 Species number of seed plants | 968 | 1,426 | 1,775 | 1,210 | 655 | 841 | 参考文献
Reference | 张光富, 2000 | 宫贵权等, 2007; 祝燕等, 2008; 张田田等, 2020 | 李乾玺, 2017; 秦运芝等, 2018 | 温庆忠, 2002; 应红涛和张文芳, 2018; 杨涛等, 2023 | 吴娅萍等, 2016; 杨涛等, 2023 | 赵本忠, 2005; 杨涛等, 2023 |
View table in article
表4
中国中亚热带6个常绿阔叶林动态监测样地所属自然保护区或风景名胜区的生物气候条件
正文中引用本图/表的段落
6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地。自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003)。从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异。局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制。本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一。在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4)。天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候。在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一。此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994)。在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因。
6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ... 1 2021 ... 根据《中国植物志》(中国科学院中国植物志编辑委员会, 1959-2004)和中国植物物种信息数据库(http://db.kib.ac.cn/)对6个样地所有胸径(DBH) ≥ 1 cm的木本植物的中文名和拉丁名进行统一整理(宋永昌等, 2015), 并在此基础上进行科、属的统计.参照《中国被子植物科属综论》(吴征镒等, 2003)和《种子植物分布区类型及其起源和分化》(吴征镒等, 2006)对所有木本植物进行科、属和种的区系类型划分.在R语言中使用ecodist包(Goslee & Urban, 2021)计算样地间物种组成的Jaccard相似性. ... Toward a better understanding of the regional causes of local community richness 1 2008 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ... 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征 1 1998 ... 吴征镒(1980)在《中国植被》中将中国亚热带常绿阔叶林划分为典型常绿阔叶林、季风常绿阔叶林、山地常绿阔叶苔藓林和山顶苔藓矮林4个植被亚型, 并根据优势种和生境类型划分出11个群系组40个群系.而《云南植被》(吴征镒和朱彦丞, 1987)将云南省内的常绿阔叶林划分为5个植被亚型, 分别是季风常绿阔叶林、半湿润常绿阔叶林、中山湿性常绿阔叶林、山地苔藓常绿阔叶林和山顶苔藓矮林, 在植被亚型之下又划分了11个群系组20个群系, 与中国亚热带东部的典型常绿阔叶林存在明显差异.国内对常绿阔叶林的研究始于20世纪40、50年代, 几十年中, 研究人员在常绿阔叶林的区系成分与物种组成(朱华, 2021)、物种多样性及其维持机制(贺金生等, 1998)、群落演替与恢复(丁圣彦和宋永昌, 2004; Wang et al, 2007)、生态系统的结构和功能(Huang et al, 2007; Lin et al, 2012)、生态系统过程及其对全球变化的响应(Zhou et al, 2013)等方面已经开展了大量研究. ... 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征 1 1998 ... 吴征镒(1980)在《中国植被》中将中国亚热带常绿阔叶林划分为典型常绿阔叶林、季风常绿阔叶林、山地常绿阔叶苔藓林和山顶苔藓矮林4个植被亚型, 并根据优势种和生境类型划分出11个群系组40个群系.而《云南植被》(吴征镒和朱彦丞, 1987)将云南省内的常绿阔叶林划分为5个植被亚型, 分别是季风常绿阔叶林、半湿润常绿阔叶林、中山湿性常绿阔叶林、山地苔藓常绿阔叶林和山顶苔藓矮林, 在植被亚型之下又划分了11个群系组20个群系, 与中国亚热带东部的典型常绿阔叶林存在明显差异.国内对常绿阔叶林的研究始于20世纪40、50年代, 几十年中, 研究人员在常绿阔叶林的区系成分与物种组成(朱华, 2021)、物种多样性及其维持机制(贺金生等, 1998)、群落演替与恢复(丁圣彦和宋永昌, 2004; Wang et al, 2007)、生态系统的结构和功能(Huang et al, 2007; Lin et al, 2012)、生态系统过程及其对全球变化的响应(Zhou et al, 2013)等方面已经开展了大量研究. ... Functional diversity versus species diversity: Relationships with habitat heterogeneity at multiple scales in a subtropical evergreen broad-leaved forest 1 2014 ... 森林动态监测样地是森林群落生物多样性科学综合研究平台, 是揭示异质景观中物种和群落多样性空间特征和维持机制的主流研究途径之一(马克平, 2011).自2004年中国科学院生物多样性委员会启动中国森林生物多样性监测网络(CForBio)建设项目以来, 国内先后建立了25个大型森林样地和近60个辅助样地, 其中包含7个大型常绿阔叶林样地(http://www.Cncdiversitas.cn/zyxm/cforbio/js/) (马克平, 2017), 亚热带常绿阔叶林结构和组成的时空格局研究进入了新的发展阶段(马克平, 2017), 围绕物种功能性状(Hu et al, 2014)、群落谱系结构(黄建雄等, 2010)、密度制约与种间相互作用(Luo et al, 2012; Wu et al, 2017)、异质生境的空间和环境效应(周光霞等, 2016)、植物?土壤微生物关系(Gao et al, 2017)等进行了深入研究, 极大深化了对常绿阔叶林群落构建的关键过程和驱动机制的认识.为了揭示常绿阔叶林群落物种多样性的动态特征, 研究人员还在大样地尺度上进行了常绿阔叶林群落物种组成、植物谱系、生态习性等方面的比较分析(宋永昌等, 2015; 车俭等, 2020).其中, 宋永昌等(2015)首次全面比较了我国亚热带中东部8个大型常绿阔叶林样地的木本植物物种组成和生态特征.尽管前人已指出以云贵高原为主要分布区的西部亚热带常绿阔叶林在群落类型和优势物种组成上与东部地区不同(吴征镒和朱彦丞, 1987), 但关于中亚热带湿润与半湿润常绿阔叶林的比较群落学研究还十分缺乏; 在亚热带地带性气候和地形地貌东西分化格局影响下, 中亚热带东、西部常绿阔叶林的群落结构、物种组成和多样性的定量特征迄今仍缺少清晰的认识. ... Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in Eastern China 1 2007 ... 吴征镒(1980)在《中国植被》中将中国亚热带常绿阔叶林划分为典型常绿阔叶林、季风常绿阔叶林、山地常绿阔叶苔藓林和山顶苔藓矮林4个植被亚型, 并根据优势种和生境类型划分出11个群系组40个群系.而《云南植被》(吴征镒和朱彦丞, 1987)将云南省内的常绿阔叶林划分为5个植被亚型, 分别是季风常绿阔叶林、半湿润常绿阔叶林、中山湿性常绿阔叶林、山地苔藓常绿阔叶林和山顶苔藓矮林, 在植被亚型之下又划分了11个群系组20个群系, 与中国亚热带东部的典型常绿阔叶林存在明显差异.国内对常绿阔叶林的研究始于20世纪40、50年代, 几十年中, 研究人员在常绿阔叶林的区系成分与物种组成(朱华, 2021)、物种多样性及其维持机制(贺金生等, 1998)、群落演替与恢复(丁圣彦和宋永昌, 2004; Wang et al, 2007)、生态系统的结构和功能(Huang et al, 2007; Lin et al, 2012)、生态系统过程及其对全球变化的响应(Zhou et al, 2013)等方面已经开展了大量研究. ... 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响 1 2010 ... 森林动态监测样地是森林群落生物多样性科学综合研究平台, 是揭示异质景观中物种和群落多样性空间特征和维持机制的主流研究途径之一(马克平, 2011).自2004年中国科学院生物多样性委员会启动中国森林生物多样性监测网络(CForBio)建设项目以来, 国内先后建立了25个大型森林样地和近60个辅助样地, 其中包含7个大型常绿阔叶林样地(http://www.Cncdiversitas.cn/zyxm/cforbio/js/) (马克平, 2017), 亚热带常绿阔叶林结构和组成的时空格局研究进入了新的发展阶段(马克平, 2017), 围绕物种功能性状(Hu et al, 2014)、群落谱系结构(黄建雄等, 2010)、密度制约与种间相互作用(Luo et al, 2012; Wu et al, 2017)、异质生境的空间和环境效应(周光霞等, 2016)、植物?土壤微生物关系(Gao et al, 2017)等进行了深入研究, 极大深化了对常绿阔叶林群落构建的关键过程和驱动机制的认识.为了揭示常绿阔叶林群落物种多样性的动态特征, 研究人员还在大样地尺度上进行了常绿阔叶林群落物种组成、植物谱系、生态习性等方面的比较分析(宋永昌等, 2015; 车俭等, 2020).其中, 宋永昌等(2015)首次全面比较了我国亚热带中东部8个大型常绿阔叶林样地的木本植物物种组成和生态特征.尽管前人已指出以云贵高原为主要分布区的西部亚热带常绿阔叶林在群落类型和优势物种组成上与东部地区不同(吴征镒和朱彦丞, 1987), 但关于中亚热带湿润与半湿润常绿阔叶林的比较群落学研究还十分缺乏; 在亚热带地带性气候和地形地貌东西分化格局影响下, 中亚热带东、西部常绿阔叶林的群落结构、物种组成和多样性的定量特征迄今仍缺少清晰的认识. ... 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响 1 2010 ... 森林动态监测样地是森林群落生物多样性科学综合研究平台, 是揭示异质景观中物种和群落多样性空间特征和维持机制的主流研究途径之一(马克平, 2011).自2004年中国科学院生物多样性委员会启动中国森林生物多样性监测网络(CForBio)建设项目以来, 国内先后建立了25个大型森林样地和近60个辅助样地, 其中包含7个大型常绿阔叶林样地(http://www.Cncdiversitas.cn/zyxm/cforbio/js/) (马克平, 2017), 亚热带常绿阔叶林结构和组成的时空格局研究进入了新的发展阶段(马克平, 2017), 围绕物种功能性状(Hu et al, 2014)、群落谱系结构(黄建雄等, 2010)、密度制约与种间相互作用(Luo et al, 2012; Wu et al, 2017)、异质生境的空间和环境效应(周光霞等, 2016)、植物?土壤微生物关系(Gao et al, 2017)等进行了深入研究, 极大深化了对常绿阔叶林群落构建的关键过程和驱动机制的认识.为了揭示常绿阔叶林群落物种多样性的动态特征, 研究人员还在大样地尺度上进行了常绿阔叶林群落物种组成、植物谱系、生态习性等方面的比较分析(宋永昌等, 2015; 车俭等, 2020).其中, 宋永昌等(2015)首次全面比较了我国亚热带中东部8个大型常绿阔叶林样地的木本植物物种组成和生态特征.尽管前人已指出以云贵高原为主要分布区的西部亚热带常绿阔叶林在群落类型和优势物种组成上与东部地区不同(吴征镒和朱彦丞, 1987), 但关于中亚热带湿润与半湿润常绿阔叶林的比较群落学研究还十分缺乏; 在亚热带地带性气候和地形地貌东西分化格局影响下, 中亚热带东、西部常绿阔叶林的群落结构、物种组成和多样性的定量特征迄今仍缺少清晰的认识. ... 2 2001 ... 6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ...
6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ... Communities and Ecosystems 1 1970 ... 生长型的百分比结构能反映群落环境特征和植物对环境的适应, 以及不同种间的趋同适应性(Rowe & Speck, 2005).本文依据Whittaker生长型系统来划分统计到的所有木本植物的类型(Whittaker, 1970), 具体仿照宋永昌等(2015)简化的三级分类, 即木本植物生长型I级划分为乔木(T)、灌木(S)和半灌木(Ss), II级划分为常绿(E)和落叶(D), III级划分为阔叶(br)、针叶(ne)、簇叶(cl)和竹叶(ba), 藤本植物生长型II级划分为木质藤本(wL)和草质藤本(hL).划分每个植物的生长型后, 统计每个样地各类生长型的百分占比. ... Community composition and nested-subset analyses: Basic descriptors for community ecology 1 1996 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ... Relative importance of density dependence and topography on tree mortality in a subtropical mountain forest 1 2017 ... 森林动态监测样地是森林群落生物多样性科学综合研究平台, 是揭示异质景观中物种和群落多样性空间特征和维持机制的主流研究途径之一(马克平, 2011).自2004年中国科学院生物多样性委员会启动中国森林生物多样性监测网络(CForBio)建设项目以来, 国内先后建立了25个大型森林样地和近60个辅助样地, 其中包含7个大型常绿阔叶林样地(http://www.Cncdiversitas.cn/zyxm/cforbio/js/) (马克平, 2017), 亚热带常绿阔叶林结构和组成的时空格局研究进入了新的发展阶段(马克平, 2017), 围绕物种功能性状(Hu et al, 2014)、群落谱系结构(黄建雄等, 2010)、密度制约与种间相互作用(Luo et al, 2012; Wu et al, 2017)、异质生境的空间和环境效应(周光霞等, 2016)、植物?土壤微生物关系(Gao et al, 2017)等进行了深入研究, 极大深化了对常绿阔叶林群落构建的关键过程和驱动机制的认识.为了揭示常绿阔叶林群落物种多样性的动态特征, 研究人员还在大样地尺度上进行了常绿阔叶林群落物种组成、植物谱系、生态习性等方面的比较分析(宋永昌等, 2015; 车俭等, 2020).其中, 宋永昌等(2015)首次全面比较了我国亚热带中东部8个大型常绿阔叶林样地的木本植物物种组成和生态特征.尽管前人已指出以云贵高原为主要分布区的西部亚热带常绿阔叶林在群落类型和优势物种组成上与东部地区不同(吴征镒和朱彦丞, 1987), 但关于中亚热带湿润与半湿润常绿阔叶林的比较群落学研究还十分缺乏; 在亚热带地带性气候和地形地貌东西分化格局影响下, 中亚热带东、西部常绿阔叶林的群落结构、物种组成和多样性的定量特征迄今仍缺少清晰的认识. ... 云南雕翎山自然保护区种子植物区系研究 2 2016 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ...
6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ... 云南雕翎山自然保护区种子植物区系研究 2 2016 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ...
6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ... 浙江天童国家森林公园微地形与植被结构的关系 1 2005 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ... 浙江天童国家森林公园微地形与植被结构的关系 1 2005 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ... 曲靖市珠江源自然保护区现状与发展对策探讨 2 2018 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ...
6个样地间的物种组成相似性显示中国亚热带常绿阔叶林物种组成存在明显的东、西部差异.生境异质性、生态位分化和扩散限制是解释群落物种组成差异的主要因素和生态过程(Hubbell, 2001; Leibold & McPeek, 2006).在种水平上, 西部3个样地中, 珠江源与鸡足山样地的物种组成相似性较低, 雕翎山与鸡足山样地的物种组成相似性略高于其与珠江源的相似性, 这与3个样地的距离关系相反.中性理论认为, 物种扩散能力受到空间距离限制, 距离越远的地点间物种组成的相似性越低(Hubbell, 2001), 这可能是导致西部3个样地物种组成差异的原因.八大公山样地与东部样地的物种组成相似性明显高于其与西部样地的相似性, Tuomisto等(2003)对亚马逊热带雨林beta多样性的研究显示, 扩散限制在大范围(研究地点间的距离超过80 km)上对群落物种组成差异的解释度不及环境因子, 即东、中、西部样地间的物种组成差异可能是环境因子主导的.在科水平上, 滇中高原地区东、中、西部3个样地的科组成有较大差异, 这可能与该地区的地史过程(唐志尧等, 2009)有关, 滇中高原位于云南高原面上, 与横断山区关系密切, 又受到金沙江及其支流切割, 地质上还受喜马拉雅造山运动和古地中海向西撤退的影响, 生境复杂, 物种形成较强烈(李锡文, 1995), 植物区系上有热带和温带过渡和交叉的特点, 地理位置上又是云南南北植物交流交汇的通道, 具有一定的复杂性(Li et al, 2015). ... 曲靖市珠江源自然保护区现状与发展对策探讨 2 2018 ... 6个样地中, 八大公山样地物种丰富度最高, 东部样地物种丰富度普遍高于西部样地.自然群落中, 物种数往往随取样面积的增加而增加(Worthen, 1996), 物种多样性总是受到局域气候、区域物种多样性的控制(Harrison & Cornell, 2003).从取样面积来看, 八大公山样地面积在6个样地中最大, 雕翎山和珠江源样地面积最小, 取样面积的差异可能影响了样地间的物种丰富度差异.局域物种库是区域物种库物种经环境筛选过滤、种间竞争等过程而稳定存留下来的物种组合(Diamond, 1975), 受到区域物种库的限制.本研究中, 八大公山国家级自然保护区种子植物总计有165科709属1,775种(卢志军等, 2013), 天童国家森林公园共有种子植物148科506属968种(杨永川等, 2005), 古田山国家级自然保护区种子植物总数为149科648属1,426种(祝燕等, 2008), 珠江源省级自然保护区种子植物共150科461属1,210种(应红涛和张文芳, 2018), 雕翎山省级自然保护区种子植物共112科355属655种(吴娅萍等, 2016), 鸡足山风景名胜区种子植物共163科447属841种(表4), 八大公山国家级自然保护区种子植物科、属、种丰富度均最高, 而雕翎山省级自然保护区科、属、种丰富度都最低, 珠江源、鸡足山在属水平上丰富度低于天童和古田山, 区域物种库大小可能是造成样地物种多样性差异的原因之一.在气候方面, 6个样地都处于亚热带季风气候区, 但东、中、西部样地气候特征迥异(表4).天童和古田山样地属于海洋性季风气候, 八大公山样地属于山地湿润季风气候, 珠江源、雕翎山、鸡足山样地则属于高原季风气候.在6个样地中, 相较于其他样地, 八大公山样地的降水最丰富, 气温最低, 霜期最长, 气候条件可能是造成其物种多样性最高的原因之一.此外, 八大公山样地保留了第三纪乃至晚白垩纪的孑遗物种, 其物种区系具有古老和邃古性, 这也可能是导致该样地木本植物物种丰富的原因(祁承经等, 1994).在西部3个样地中, 珠江源样地植被处于火烧后恢复中期阶段, 雕翎山样地属于择伐后恢复植被, 鸡足山样地只有局部强度火烧干扰痕迹, 这些可能是导致这3个样地物种丰富度差异的原因. ...
本文的其它图/表
|