|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
昆虫病原真菌感染昆虫宿主的机制和宿主昆虫的防御策略研究进展
生物多样性
2023, 31 (11):
23273-.
DOI: 10.17520/biods.2023273
昆虫病原真菌是寄生于昆虫而引起昆虫死亡的一类真菌, 具有高杀虫效力且对环境的影响较小, 常被用作生物杀虫剂来控制害虫, 具有重要的应用价值。本文对昆虫病原真菌的分类信息进行了归纳总结, 系统阐述了真菌侵染宿主的过程以及在感染过程中对宿主行为的改变及孢子传播的机制。随后梳理了昆虫在面对感染时采取相应抵抗感染的策略, 主要包括体壁防御和自身免疫系统的防御以及行为上避免真菌病原体。在此基础上讨论了真菌对昆虫的致病机制以及与昆虫的防御机制二者联系紧密的协同进化关系。最后我们对未来的研究方向进行了展望, 强调应该依据传染病传播动力学的基本模型探究昆虫虫口密度与环境中的孢子浓度对传染病暴发的影响, 研究真菌传播规律以指导田间应用。
表1
昆虫病原真菌及其分类
正文中引用本图/表的段落
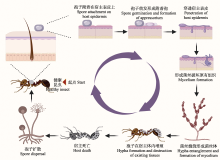
昆虫病原真菌是一类影响宿主发育和生殖过程并最终导致宿主死亡的真菌, 目前已经发现有750-1,000种真菌对昆虫具有致病性(Rajula et al, 2020), 分属于真菌界的12个纲。这些致病真菌常见于子囊菌门、接合菌门、卵菌门、壶菌门、担子菌门和虫霉门(Hibbett et al, 2007; Litwin et al, 2020)。其中, 虫霉门曾属于接合菌门, Humber (2012)年将其提升为独立的一个门。昆虫病原真菌的宿主几乎分布在所有的目中, 最常见的是半翅目、双翅目、鞘翅目、鳞翅目、直翅目和膜翅目。本文提到的昆虫病原真菌及其分类见表1。需要指出的是, 一些真菌的宿主范围有限, 例如粉虱座壳孢(Aschersonia aleyrodis)只感染粉虱(Ingle et al, 2022); 莱氏绿僵菌(Metarhizium rileyi, 旧称莱氏野村菌)只感染鳞翅目和半翅目昆虫(Fronza et al, 2017; Umaru & Simarani, 2022); 而球孢白僵菌和金龟子绿僵菌能够感染多个昆虫目中的700多个物种(Ebani & Mancianti, 2021)。
昆虫病原真菌能够以孢子的形式存在于植物表面、土壤或者空气中。与细菌或病毒不同, 昆虫病原真菌不需要被昆虫摄取, 它们能够直接穿透宿主完整的表皮到达血腔(Sinha et al, 2016; Bilgo et al, 2018)。真菌对宿主表皮的粘附主要涉及两个过程: 由非特异性疏水力和静电力组合介导的被动附着以及通过特异性配体和粘附素相互作用的附着 (Holder & Keyhani, 2005; Wang & St. Leger, 2007)。球孢白僵菌和金龟子绿僵菌都会产生疏水性分生孢子, 这些分生孢子含有由疏水蛋白组成的表面杆状层, 有助于孢子立即紧密结合到疏水表面 (Holder & Keyhani, 2005; Holder et al, 2007)。第二步涉及粘附素产生, 例如已被表征的粘附素MAD1和MAD2, 能够直接介导真菌对宿主细胞的粘附, 对真菌定殖起着重要的作用(Wang & St. Leger, 2007; Barelli et al, 2011)。
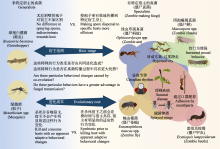
了解病原体的传播过程, 即它如何进入易感宿主引起感染并传播是传染病研究的基础(van Seventer & Hochberg, 2017), 可以基于昆虫病原真菌的传播机制和传染病传播动力学的模型开展以下研究: (1)建立关于种群中易感者、感染者以及环境中孢子浓度随时间变化的微分方程模型。其中, 易感者表示目前没有患病, 但无免疫能力, 可以被感染而患病的宿主昆虫, 感染者表示已经被感染, 并且可以把疾病传染给易感者的昆虫。每只感染昆虫死亡后都会释放大量孢子, 所以通常来说, 环境中被感染昆虫的虫口密度越高, 环境中的孢子浓度也越高, 又由于真菌对昆虫的感染是通过孢子传播实现的, 因此可将孢子浓度引入模型。(2)将造成环境中孢子浓度减少的原因概括为孢子死亡以及粘附感染其他昆虫造成的孢子转移。环境中孢子浓度的减少可以符号化表示为环境中孢子的移除率, 其倒数即为真菌传染病的感染期。(3)计算模型的基本再生数。基本再生数可以表示每只被感染的昆虫死亡后产生的孢子感染的健康昆虫数量与感染期的乘积。如果基本再生数大于1, 表示一只被感染的昆虫在感染期内产生的孢子平均能感染的健康昆虫数量超过1只, 此时由最初少量孢子引起的传染病可以在昆虫种群内传播, 否则疾病将自然消亡。这通常要求最初环境中的孢子浓度达到一个阈值(Inglis et al, 2012; Boomsma et al, 2014)。(4)计算真菌传染病存在时, 宿主种群密度达到稳定的平衡点。若真菌与其宿主长期共存, 那么由真菌引起的疾病可以看作宿主种群固有的一种区域性疾病, 即地方病。地方病是否存在也是由昆虫的种群密度以及环境中孢子浓度决定的。今后建议从昆虫的虫口密度与环境中的孢子浓度入手开展研究, 通过控制模型参数探究不同种群密度和孢子浓度与病原真菌的流行关系。基于传染病传播动力学模型进行真菌传播规律的研究可对未来利用真菌防治害虫和控制有益昆虫的病害提出科学的指导建议。
本文的其它图/表
|