
|
|
||
|
基于中国科学院西北高原生物研究所馆藏标本分析青藏高原雀形目鸟类物种和功能多样性
生物多样性
2023, 31 (5):
22638-.
DOI: 10.17520/biods.2022638
物种多样性和功能多样性是生物多样性的两个主要研究内容, 研究不同区域物种多样性和功能多样性可以为生物多样性保护提供重要的理论支撑。本研究以中国科学院西北高原生物研究所馆藏的青藏高原11个生态地理分区雀形目鸟类为研究对象, 结合其性状数据, 计算3种物种α多样性指数(物种丰富度、Shannon熵和Pielou均匀度)、3种功能α多样性指数(功能丰富度、功能均匀度和功能离散度)、物种和功能β多样性及其对应的周转和嵌套成分, 并分析了物种多样性与功能多样性之间的关系。研究结果表明: (1)青藏高原鸟类物种α多样性呈东南高、西北低的趋势。功能丰富度和功能均匀度显示出明显的空间差异, 全区物种丰富度和功能丰富度平均值较低, 功能离散度在全区均较高; (2)物种β多样性高于功能β多样性, 但二者整体均具有较高的数值。物种β多样性主要由周转成分构成, 而嵌套和周转成分在功能β多样性中占比接近; (3)功能丰富度与物种丰富度及Shannon熵呈显著正相关关系。物种和功能β多样性呈显著相关关系, 物种和功能周转成分贡献率呈显著相关关系。以上结果表明功能丰富度较高的分区中鸟类占据较大的功能性状空间, 而分区中功能均匀度和功能离散度较高则体现出鸟类可以更均匀、更有效地利用食物资源。结合物种和功能α、β多样性可以很好地表征鸟类多样性与生态系统的关系。 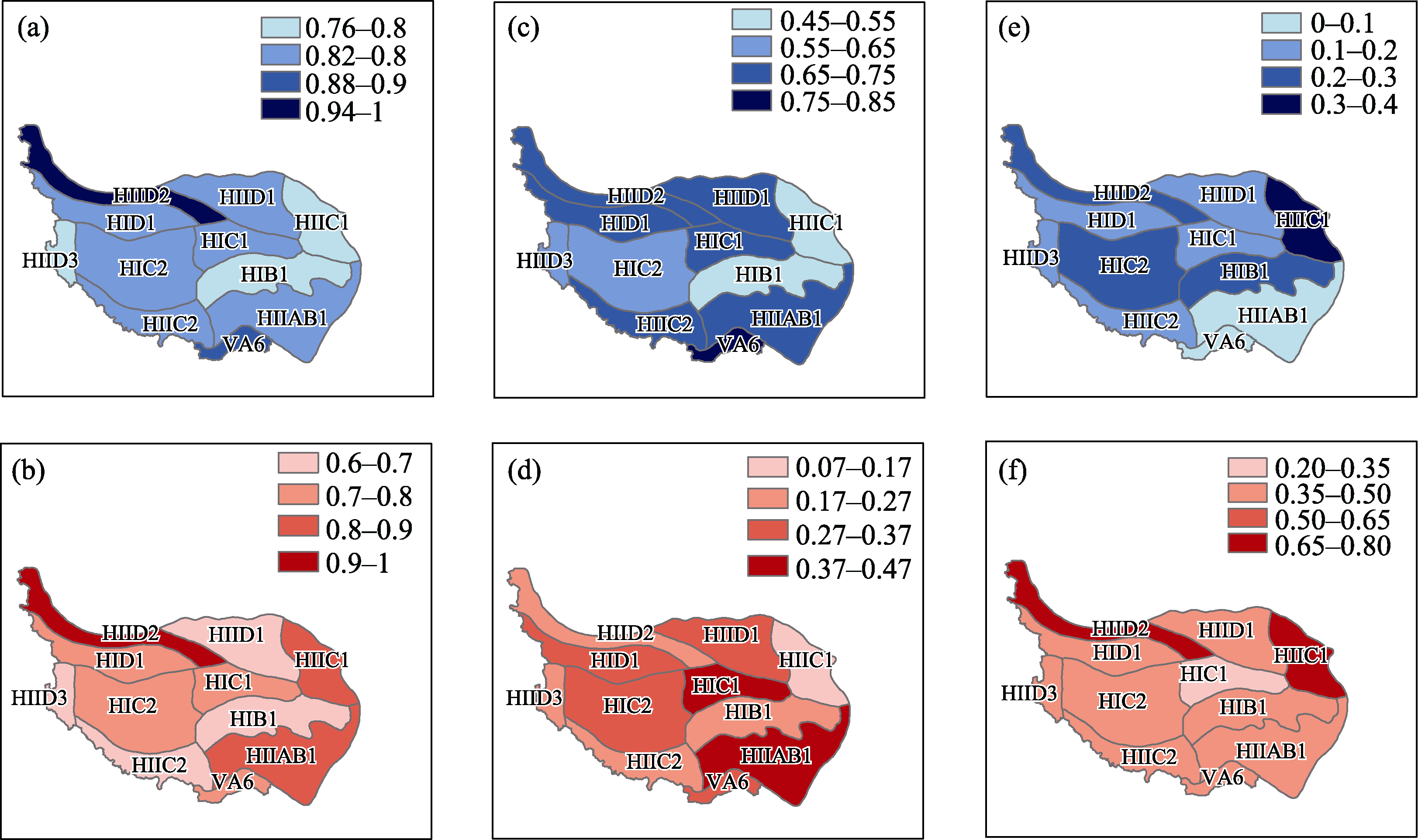 View image in article
图2
不同生态地理分区物种β多样性(a)、周转成分(b)、嵌套成分(c)和功能β多样性(d)、周转成分(e)、嵌套成分(f)空间模式。
各分区指标值计算为该分区与其余分区配对比较的平均值。
正文中引用本图/表的段落
青藏高原全区域鸟类物种β多样性变化范围为0.42-0.98 (平均值为0.85 ± 0.11, 表1), HIID2分区物种β多样性最高, HIID3、HIIC1和HIB1分区β多样性最低(图2a)。不同分区共有物种数仅为非共有物种数的18.8%。周转成分在物种β多样性中占主要部分(表1, 图2b, c), 占β多样性总值的78.8% ± 20.0%)。
青藏高原全区域鸟类功能β多样性变化范围为0.13-1.00 (0.75 ± 0.20, 表1), HIID2分区功能β多样最高, HIID3、HIID1、HIB1和HIIC2分区β多样性最低 (图2d)。不同分区功能性状空间重合面积仅为非重合面积的26.7%。与物种β多样性不同, 嵌套成分在功能β多样性中占主要部分(表1, 图2e, f), 占β多样性总值的60.7% ± 35.9%。
从表1和图2可以看出, 青藏高原全区域鸟类物种和功能β多样性均较高, 说明不同分区之间物种组成和性状特征差异较大, 此结果可由以下两方面来解释。一方面, 各分区平均物种丰富度为61, 仅占区域总物种丰富度的21.8%; Fric呈现相似的结果, 平均Fric占总Fric之比仅为22.9% (图1)。其次, 与研究区域低物种丰富度和低Fric类似, 各分区共有物种数和功能性状空间重合面积也较低(分别为18.8%和26.7%)。此结果也被多项研究所证实(Villéger et al, 2013; Xu et al, 2021)。如青藏高原东部区域较高的平均丰富度和共有物种数导致了较低的物种β多样性, 平均值仅为0.48左右(Xu et al, 2021)。低共有物种数和低重合面积却导致了不同的 β多样性组成。周转成分是物种β多样性的主要部分, 在物种β多样性中占比为78.8%; 而嵌套成分构成功能β多样性的主要部分, 占功能β多样性的比例为60.7% (表1)。物种周转描述了不同群落间物种发生替换, 它反映了环境过滤或空间、历史因素的限制; 而嵌套的产生是由于物种的丧失使得拥有较少物种的群落成为拥有较多物种群落的子集, 它反映的是一种非随机的物种丧失过程(Baselga, 2010)。本文结果说明青藏高原全区域较高的鸟类物种β多样性主要由物种替换造成, 而功能β多样性是区域内物种替换和物种丧失共同作用的结果。
本文的其它图/表
|



{kind=link}