|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
八大公山亚热带森林木质残体中大型无脊椎动物群落特征
生物多样性
2022, 30 (12):
21476-.
DOI: 10.17520/biods.2021476
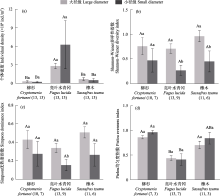
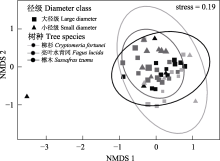
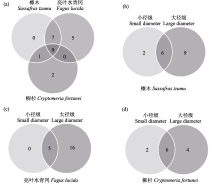
木质残体可为大型无脊椎动物提供重要栖息地、食物等资源, 并影响其生物多样性。目前针对不同树种、径级及分解阶段的木质残体如何调控土壤大型无脊椎动物群落结构尚不清楚, 相关研究在亚热带森林地区尤为稀缺。为此, 本文选取湖南省八大公山国家级自然保护区柳杉(Cryptomeria fortunei)、亮叶水青冈(Fagus lucida)及檫木(Sassafras tzumu) 3种树种为研究对象, 每种树种分别选取两类径级(直径分别为10 ± 2 cm、4 ± 2 cm)不同分解阶段的木质残体, 对其中的大型无脊椎动物进行调查。调查于2020年10-11月完成。结果显示: 共捕获大型无脊椎动物2,558只, 隶属4门10纲23目, 不同树种的优势类群、常见类群及稀有类群均存在差异。亮叶水青冈木质残体中大型无脊椎动物个体密度显著高于柳杉和檫木。亮叶水青冈和檫木大径级木质残体中大型无脊椎动物Shannon-Wiener多样性指数显著高于小径级, 3个树种大径级木质残体中大型无脊椎动物的类群数、特有类群数均大于小径级。木质残体中大型无脊椎动物的Shannon-Wiener多样性指数、Simpson优势度指数及Pielou均匀度指数与木材密度显著负相关, 表明随着分解的进行木质残体中大型无脊椎动物群落呈明显变化趋势。木质残体的理化性质(相对含水率、全氮、全碳及碳氮比)和土壤温度、湿度与木质残体中大型无脊椎动物群落特征具有相关性。研究初步表明, 大型无脊椎动物群落特征在所选树种、径级及分解阶段木质残体中具有差异, 在亚热带森林中同时保留不同树种、不同大小径级的木质残体或有利于增加大型无脊椎动物多样性。
表1
柳杉、亮叶水青冈及檫木木质残体大型无脊椎动物群落组成
正文中引用本图/表的段落
用于测量理化性质的木质残体带回到室内, 立即测量每个5 cm圆盘的重量, 记为湿重G湿 (g)。圆盘自然风干后, 用电锯将圆盘沿与土壤接触面的垂直线分为两半, 记为圆盘1和圆盘2, 分别称其风干重G风1 (g)、G风2 (g)。圆盘1用于测量木质残体理化性质, 圆盘2作为备用样品储存。圆盘1用纯净水完全浸泡使其饱和, 擦拭表面水分后称重, 记为饱和重G饱1 (g)。采用排水法(Zuo et al, 2014)测定圆盘的体积, 即在量筒中倒入一定量的水, 读出水面刻度体积V1, 然后将吸饱水的圆盘放入, 并用薄刀片按压使其完全浸入水中, 观察此时的体积V2。最后将圆盘沥干水分, 放入信封, 置于65℃烘箱中烘至恒重, 称重, 记为烘干重G (g)。根据圆盘1的烘干重和体积, 可以计算出整个圆盘的密度(Zuo et al, 2014)。根据圆盘1风干重G风1、饱和重G饱1、烘干重G及圆盘2风干重G风2, 可以算出整个圆盘的烘干重G烘 (g)和饱和湿重G饱 (g), 再结合圆盘湿重G湿, 计算圆盘相对含水率(%) (张瑜和金光泽, 2016)和饱和含水率(%) (杨霞等, 2019)。计算公式如下:
本次调查共收集到木质残体中大型无脊椎动物2,558只, 隶属于4门10纲23目(表1)。优势类群1个, 即双翅目幼虫, 个体总数为1,818只, 占比为71.07%; 常见类群5个, 包括膜翅目、等翅目、鞘翅目、鞘翅目幼虫和姬马陆目, 个体总数614只, 占比24.00%; 稀有类群包括石蜈蚣目、蜘蛛目、蜈蚣目、综合目等18个类群, 个体总数126只, 占比4.93%。
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||