|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
拉萨河中下游纤毛虫群落时空分布模式及其驱动机制
生物多样性
2022, 30 (6):
22012-.
DOI: 10.17520/biods.2022012
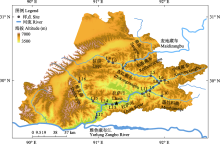
为了探究拉萨河中下游纤毛虫群落的组成模式、时空多样性格局及其维持机制, 本文于2015年5月和8月以及2016年10月在拉萨河中下游17个样点进行采样, 采用活体观察、鲁哥氏碘液固定染色以及Wilbert蛋白银法相结合的物种鉴定方法, 对纤毛虫群落结构进行了研究。通过Shannon多样性指数、Margalef丰富度指数、物种数分析群落结构时空上的差异性; 通过共现网络分析纤毛虫类群之间的相互作用; 通过冗余分析(redundancy analysis, RDA)探讨水体理化因子对纤毛虫群落结构的影响。结果表明, Shannon多样性指数在季节和河段间没有显著性差异; Margalef丰富度指数、物种数在河段间存在极显著性差异; 中游和下游河段共现网络节点间的相关关系均以正相关为主; 溶解氧(DO)、总氮(TN)、总磷(TP)、总溶解盐(TDS)是影响纤毛虫群落结构的关键因子。综上所述, 拉萨河中下游纤毛虫群落结构在季节间没有显著差异, 在空间上具有显著差异; 纤毛虫在纲级水平上类群间的相互作用以协同作用为主导, 不同类群间存在复杂的相互作用, 整体上互作关系在春季较为复杂、夏季较为简单; 影响拉萨河中下游纤毛虫群落结构是多个环境因子共同作用的结果。
表2
拉萨河中下游纤毛虫共现网络的关键拓扑结构特征
正文中引用本图/表的段落
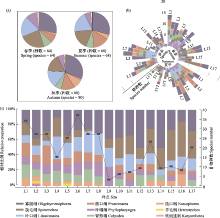
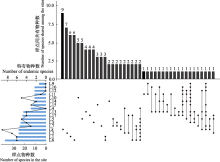
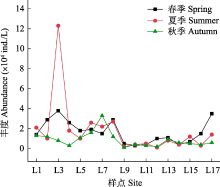
从季节上看, 物种数排序为秋季 > 夏季 > 春季。春季鉴定纤毛虫64种, 隶属8纲18目31科33属, 以寡膜纲种类最多, 旋毛纲次之; 夏季有68种, 隶属于8纲20目35科39属, 以寡膜纲种类最多, 旋毛纲次之; 秋季有80种, 隶属于9纲19目29科32属, 以寡膜纲种类最多, 叶口纲次之。从空间上来看, 物种数为中游 > 下游。中游纤毛虫类群以寡膜纲和叶口纲为主, 支流雪绒藏布样点L6物种数最多(37种), 支流拉曲样点L4最少(16种); 下游以寡膜纲和旋毛纲为主, 干流样点L13物种数最多(24种), 支流墨竹玛曲样点L9最少(7种)。从拉萨河中下游季节上和空间上来看, 纤毛虫群落组成不尽相同, 表现出时空差异性(图2)。
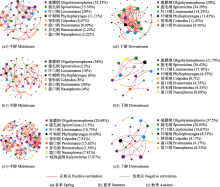
不同河段间的群落共现网络分析结果显示(图6, 表2), 中游河段纤毛虫群落的网络中边和点的数量明显高于下游河段。中游和下游河段节点间的相关关系均以正相关为主, 说明纤毛虫类群间协同作用强于竞争作用。此外, 除春季下游外, 其余5个网络的模块化系数为0.714-0.881; 6个网络的平均路径长度为1.00-1.42。在河段上, 中、下游河段图密度、聚类系数差异不大; 在季节上, 春季 > 秋季 > 夏季, 说明纤毛虫类群间互作关系在春季较为复杂、夏季较为简单。
Lupatini等(2014)认为不同物种之间的相互作用是影响群落分布格局和生态系统功能的重要因素之一, 尤其是在生态系统循环过程中, 相比较于物种丰富度和丰度以及多样性, 微生物类群之间的相互作用似乎对生态系统过程和功能的贡献更大。在共现网络中, 两个节点间边的正、负相关性分别代表相连物种之间的互惠和竞争关系(Zhang et al, 2018)。研究发现, 整个中下游河段, 网络节点之间正相关所占比例较大, 表明纲级水平上纤毛虫类群协作关系强, 而竞争关系弱。虽然同类群生物往往由于对资源的竞争普遍存在拮抗作用(Selbmann et al, 2013), 但是在极端生境中它们之间的协同作用往往占据主导地位。这说明在生态网络中不同微生物类群之间的互作关系表现为协同作用还是拮抗作用, 会受具体生境的影响。网络模块化指标表明, 除春季下游外, 其余5个网络均具有模块化结构; 6个网络的平均路径长度与已报道的显示“小世界” (small-world)特性的网络平均路径长度在类似范围, 说明所构建的网络均具有“小世界”属性(Deng et al, 2012)。网络的连接数可反映纤毛虫类群间的连通性(Deng et al, 2012), 具有较高连通性的网络对于环境干扰反应更为迅速(Zhou et al, 2010; Zhang et al, 2018)。拉萨河中下游春季纤毛虫群落的网络连通性比夏、秋季更高且更为复杂, 中游河段的网络连通性比下游更高且更为复杂(表2)。由此可推断, 在季节上, 春季纤毛虫群落可能对拉萨河的环境变化反应更加敏感; 在空间上, 中游纤毛虫群落可能对拉萨河的环境变化反应更加敏感。
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||