
|
|
||
|
适合度的相对性与路径依赖的自然选择
生物多样性
2022, 30 (1):
21323-.
DOI: 10.17520/biods.2021323
自然选择理论认为生物个体或者种群在进化的过程中, 其基因或者性状、行为策略的选择一定是能够提高其适合度或者达到某个可期的“目标”。然而, 随着某个突变基因或者性状特征、行为策略在种群中扩散, 其期望收益将随着其在种群中分布的密度变化或环境改变而发生改变, 这就是适合度景观的悖论, 即静态的、固定可期望的收益可能因此而不存在。基于动态而非静态适合度景观的概念, 我们提出路径依赖的自然选择概念。路径依赖的自然选择过程中, 一个突变的基因或表型在某种环境下随机产生, 但是该基因或表型在某些特定环境下会产生正反馈。尤其是在正反馈与随机漂变的共同作用下, 多条路径的演化就可能发生, 并且其路径的形成将同时受到其种群进化历史过程和空间特征分布等因素的强烈影响。而在不同路径下, 由于观测维度、角度和尺度的不同, 适合度意义将因此而存在不同。在此意义下, 自然选择更可能选择路径频率而不是适合度大小。基于上述概念, 我们借鉴现代物理学中复函数的方法, 来描述多重动力对物种形成或者生物特征、种群进化等路径依赖的演化过程, 以期为同域物种、隐存种形成以及生物多样性演化提供解释机制。 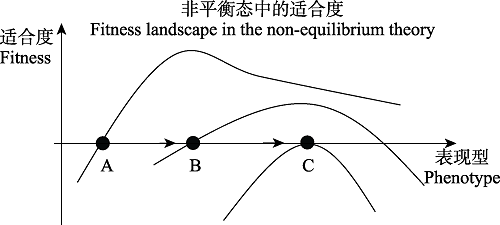 View image in article
图2
适合度演化的非平衡态理论。这时适合度景观是动态的。在A点时, 右侧性状适合度比较大, 物种向适合度较大的B性状演化。但演化到B点时, 适合度景观发生变化, 适合度并没有如预期增长。
正文中引用本图/表的段落
达尔文在《物种起源》(Darwin, 1859)中指出, 适合度是衡量一个个体存活和繁殖成功机会的参量。一个基因的突变或者表型特征能否在自然选择的过程中保留下来, 其核心的动力就是该基因突变或者表型特征能否提高个体的适合度。上述自然选择理论思想作了一个基本的前提假设, 即该突变基因或者性状特征的期望收益是个固定常数。在图1展示的静态适合度景观中, 表型演化至生态位1和生态位2时适合度达到局部最优, 表型向着这两个对应的表型值演化以提高自己的适合度(图1)。然而, 自然选择理论这个基本的前提假设却面临着一个巨大的理论挑战: 随着某个突变或者表型特征在种群中扩散, 其在种群中的密度分布将因此而改变,基因突变或者表型特征的期望收益也将必然发生改变。例如在图2所示的动态适合度景观中,在A点时, 右侧性状适合度比较大, 物种向适合度较大的B性状演化; 但演化到B点时, 在物种之间的相互作用和环境反馈的共同作用下, 适合度景观发生变化, 适合度并没有如预期增长。
基于静态的适合度景观思想, Traulsen等(2007)讨论了进化最优路径的问题。他们认为表型特征是沿着时间或者速度最快的路径进化的, 因而就存在所谓的最优路径。在此情况下, 自然选择过程也许存在多条路径, 在选择强度较大时, 所需时间最短的路径被自然选择下来。然而, 正如前文所述, 适合度景观并非是静态的, 而是随着其性状特征空间分布的频率变化而变化, 存在动态变化(李敏岚等, 2021) (图2)。也就是说, 当进化目标不固定时, 路径最短或者速度最快的最优路径将失去其逻辑的合理性。
在图2所展示的动态适合度景观的A点时, 右侧表型的适合度较大, 物种向适合度较大的B点演化。但演化至B点时, 种群结构的改变使得适合度景观发生变化, 适合度并没有如预期增长。这是因为, 有限种群中的表型在演化的过程中同时受到随机漂变和选择的力量(Ewens, 2004), 其适合度会发生改变。以合作策略在种群中的演化为例, 其适合度表示为:
本文的其它图/表
|


{kind=link}