|
|
|||||||||||||||||||||||||||||||||||
|
观光木片断化居群的遗传多样性和交配系统
生物多样性
2012, 20 (6):
676-684.
DOI: 10.3724/SP.J.1003.2012.09083
观光木(Tsoongiodendron odorum)是我国特有的濒危植物, 有居群较小、间断分布的特点。为探讨生境片断化对观光木居群的遗传多样性和交配系统的影响, 我们采用8对微卫星引物对采自广东南昆山的观光木片断化居群的61个成株和15个家系共780个种子进行了基因分型, 调查了5个空间隔离的斑块中观光木的两代遗传多样性以及各个层次(居群、斑块及个体)的交配系统参数。结果显示: 片断化生境中观光木成株遗传多样性水平适中(HE = 0.522), 种子遗传多样性比成株稍低(HE = 0.499), 但没有显著的差异, 近交系数在种子中也没有显著的升高, 暗示生境片断化并没有侵蚀观光木的遗传多样性; 多位点交配系统分析(MLTR)结果表明, 该片断化生境中观光木为高度异交树种(tm = 1.000), 只有较少的双亲近交(biparental inbreeding)和相关性交配(correlated mating)事件发生, 但有效花粉供体(effective pollen donor)数目较少(Nep为3.7-5.4); 5个斑块间异交率差异不明显, 但小的斑块有效花粉供体相对较多; 另外, 观光木个体之间的异交率存在明显差异, 少数个体存在自交现象。这些结果为濒危物种观光木的长期保护提供了重要的遗传学信息。
表1
观光木片断化居群中每个斑块的成株、采种成株、果实和种子数
正文中引用本图/表的段落
观光木(Tsoongiodendron odorum)是我国特有的濒危植物, 有居群较小、间断分布的特点。为探讨生境片断化对观光木居群的遗传多样性和交配系统的影响, 我们采用8对微卫星引物对采自广东南昆山的观光木片断化居群的61个成株和15个家系共780个种子进行了基因分型, 调查了5个空间隔离的斑块中观光木的两代遗传多样性以及各个层次(居群、斑块及个体)的交配系统参数。结果显示: 片断化生境中观光木成株遗传多样性水平适中(HE = 0.522), 种子遗传多样性比成株稍低(HE = 0.499), 但没有显著的差异, 近交系数在种子中也没有显著的升高, 暗示生境片断化并没有侵蚀观光木的遗传多样性; 多位点交配系统分析(MLTR)结果表明, 该片断化生境中观光木为高度异交树种(tm = 1.000), 只有较少的双亲近交(biparental inbreeding)和相关性交配(correlated mating)事件发生, 但有效花粉供体(effective pollen donor)数目较少(Nep为3.7-5.4); 5个斑块间异交率差异不明显, 但小的斑块有效花粉供体相对较多; 另外, 观光木个体之间的异交率存在明显差异, 少数个体存在自交现象。这些结果为濒危物种观光木的长期保护提供了重要的遗传学信息。
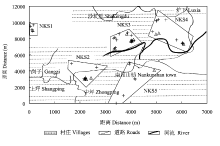
南昆山(23°27′N, 114°38′E)地跨广东龙门、从化、增城1县2市, 位于北回归线内, 该地区观光木居群分布在海拔400-500 m的常绿阔叶林群落中。本研究所选取的观光木典型片断化居群位于南昆山自然保护区之外, 成株零散分布于南昆山镇周围, 人类活动频繁, 人为干扰较为严重。在观光木自然分布居群内采用高精度GPS对成株进行精确定位, 共有成株61株。每株采集新鲜叶片, 置于装有硅胶的密封塑料袋快速干燥, 以备DNA提取。根据空间分布, 将成株划分为片断化隔离的5个小斑块(图1)。在当年结果的成株中选取15株进行果实的采集, 由于观光木果实较少, 每株只能采集果实2-4个, 平均3.8个。每个果实中随机选取种子14-17粒, 共有780粒(表1), 置于4℃冰箱中保存备用。
采用改良的3%CTAB法(Doyle & Doyle, 1987) 提取成株干燥叶子以及子代种子的基因组DNA, 用1.0%的琼脂糖凝胶电泳(λ/Hind III marker)检测其质量, 并用紫外分光光度计(GeneQuant, Eppendorf, Germany)检测模板DNA浓度。本研究选用了8对扩增条带清晰的SSR引物, 其中7对是已发表的观光木SSR引物(TOD1, TOD2, TOD5, TOD6, TOD8, TOD11, TOD12; Jiang et al., 2011), 另外1对是新开发的SSR引物TOD52(引物序列为F: 5'-CTCACC- AACACAAGCGAGAA-3'; R: 5'-CCTC TTCTTCCT- CTTCTTCTTCTTC-3', 退火温度为55℃), 引物由上海生工生物工程有限公司合成。采用FAM、HEX、TAMRA和ROX4种荧光染料对SSR正向引物的5'端分别进行标记, 用这8对引物对61个成株以及780个子代的DNA样本进行PCR扩增。PCR扩增反应在PTC-200的PCR仪上进行。PCR扩增反应体系和程序参照Jiang等(2011)。将扩增产物送至北京基诺博实生物科技有限公司, 在ABI3730自动测序仪上进行电泳分离检测, 得到SSR的荧光峰图。
由表4中可见, 南昆山片断化观光木居群总的多位点异交率(tm)为1.000, 单位点异交率(ts)为0.997(0.995-0.999)。南昆山片断化居群的双亲近交水平很低, tm-ts仅为0.003。观光木rp(m)为0.229(0.186-0.272), 表明观光木存在相关性交配。根据Nep = 1/ rp(m) (Sun & Ritland, 1998) , 观光木居群的有效花粉供体数目(Nep)较少, 为4.4(3.7-5.4)。rp(s)-rp(m)大于零, 但不显著, 表明只有小部分花粉供体是近亲关系。
南昆山片断化生境中的观光木种子遗传多样性比成株稍低, 等位基因丰富度也相对较低, 但模拟检测发现这些参数在两个世代之间并没有显著的差别, 表明观光木居群并没有因为生境片断化而导致种子的遗传侵蚀。种子的近交系数没有显著的增加, 相反, 所有斑块内的种子以及斑块NKS1、NKS3、NKS5内的成株表现出杂合子过剩的现象。环境压力和近交衰退是导致对杂合子选择的两个因素。很多研究表明居群杂合子过量是由于自然选择更有利于杂合体, 使杂合个体具有更大的适合度, 从而在居群中占据优势地位, 如山金车(Arinica montana) (Luijten, 2000)和毛柄小勾儿茶(Berchemiella wilsonii var. pubipetiolata) (Kang et al., 2005)。一般而言, 造成居群中杂合子过剩的原因主要有繁殖居群数量少、超显性、对纯合子的逐步选择、负选型交配及无性繁殖等(Stoeckel et al., 2006; Rasmussen & Kollmann, 2008)。观光木成株数目少, 且花多果少导致种子产量低, 致使繁殖居群过小, 这些可能是南昆山片断化观光木居群杂合子过剩的原因。本研究中, 种子的近交系数比亲本低, 可能是因为早期表达的致死效应或亚致死效应使幸存的种子中存在更多的杂合个体, 从而导致杂合子过剩。
本文的其它图/表
|
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()