
|
|
||
|
西藏横断山区溪流细菌beta多样性组分对气候和水体环境的响应
生物多样性
2020, 28 (12):
1570-1580.
DOI: 10.17520/biods.2019390
理解沿环境或空间梯度的群落组成变化(即beta多样性)一直是生态学和保护生物学的中心问题, 且beta多样性的形成机制及其对环境的响应已成为当前生物多样性研究的热点问题。本文以西藏横断山区怒江和澜沧江两个流域入江溪流中的细菌为研究对象, 使用Baselga的beta多样性分解方法, 基于Sørensen相异性指数将细菌的beta多样性分解为周转(turnover)和嵌套(nestedness)两个组分, 探究了细菌beta多样性及其分解组分随海拔距离的分布模式, 并且衡量了环境、气候和空间因子的相对重要性。结果表明, 两个流域中细菌的群落结构显著不同。两个流域的细菌总beta多样性和周转组分随海拔距离的增加而增加, 周转组分占总beta多样性的比例较大。气候和环境因子是两个流域中细菌总beta多样性及周转过程的重要预测因子, 并且所有的显著因子均为正相关, 其中环境因子中相关性最高的为海拔距离(R 2= 0.408, P < 0.001), 而气候因子中相关性最高的为年均温差(R 2= 0.417, P < 0.001)。方差分解结果暗示嵌套组分主要受空间扩散的影响; 总beta多样性和周转组分在环境较恶劣的澜沧江主要受环境过滤的影响, 而在环境较温和的怒江主要受空间扩散和环境过滤的共同影响。此外, 较为恶劣的环境条件会增加细菌的总beta多样性和周转率, 并且会形成更强的环境筛选作用去影响细菌群落的物种组成。我们的研究表明对西藏横断山区水体细菌多样性的保护需要从整个流域入手, 而非少量的生物多样性热点地区。 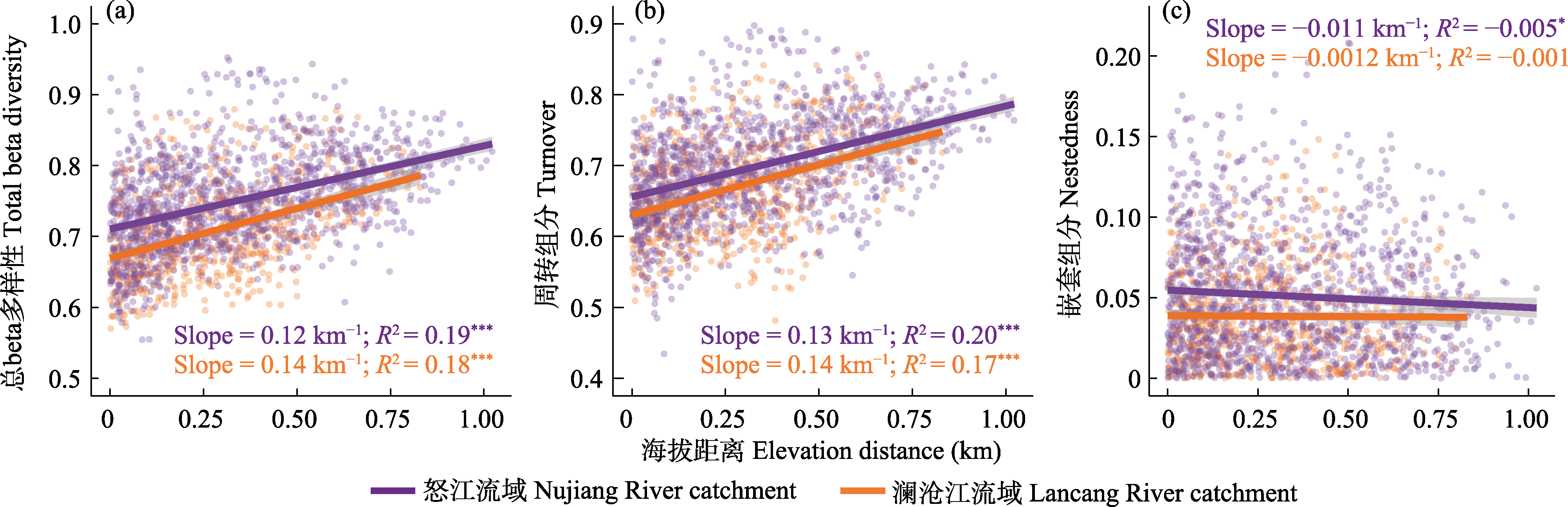 View image in article
图2
澜沧江和怒江流域细菌总beta多样性(a)、周转组分(b)和嵌套组分(c)与海拔距离的关系。其显著性水平分别为: * P ≤ 0.05, ** P ≤ 0.01, *** P ≤ 0.001。
正文中引用本图/表的段落
对于细菌beta多样性及其分解组分, 总beta多样性和周转组分均在两个流域中表现出了随海拔距离增加而增加的趋势(图2a, b)。怒江流域的细菌beta多样性大于澜沧江, 总beta多样性的均值分别为0.735和0.689, 而周转组分的均值分别为0.685和0.652。澜沧江流域沿海拔梯度的总beta多样性变化最快, 斜率为0.14/km, 而怒江流域为0.12/km; 对于周转组分, 澜沧江流域的斜率为0.14/km, 而怒江流域为0.13/km。但嵌套组分仅在怒江流域有显著的下降趋势, 斜率为-0.011/km (图2c)。
研究发现, 随着海拔距离的增加, 细菌beta多样性分解组分的差异增大(图2)。我们计算的细菌beta多样性分解组分随海拔的变化模式与典型的海拔距离衰减模式相同。出现这种模式可能是因为特定的细菌群落会出现在适宜生存的海拔高度以及栖息地中, 导致其在整个海拔梯度上分布不均匀。同时, 由于我们的采样地点为入江溪流, 水体相对比较独立, 物种的扩散具有很强的限制性, 距离衰减关系会更强(Barberan & Casamayor, 2010)。目前已经有类似的海拔梯度分布模式在一些物种中被研究(Swenson et al, 2010; Wang et al, 2012; Bishop et al, 2015)。与Bishop等(2015)提出的结果相似, 我们发现周转组分占总beta多样性的成分比较大。在特定的海拔高度下, 细菌群落会通过演替出新的细菌来应对环境选择。细菌对于环境的适应性分布是造成细菌总beta多样性和周转过程沿海拔梯度出现显著海拔距离衰减模式的重要原因。我们发现的物种周转过程为细菌总beta多样性形成的主要驱动力, 凸显了将整个山区作为生物多样性库的重要性, 生物保护要针对整个海拔梯度内的物种。对于嵌套组分, 我们没有观察到显著的海拔距离衰减模式(P > 0.01)。如果观察到嵌套组分占总beta多样性的成分比较大, 那么生物保护只需要针对物种丰富度最高的海拔高度即可(Socolar et al, 2016)。
我们还发现, 细菌在相对恶劣的环境中具有更高的beta多样性以及周转组分。通常认为海拔对于细菌群落beta多样性分布的影响为间接影响, 即通过对气候和环境等因素的作用对群落产生影响。随着海拔的升高, 气温会逐渐下降。与低海拔相比, 高海拔地区的环境相对比较恶劣, 从图2可以发现, 在澜沧江和怒江流域, 细菌总beta多样性随着海拔距离的增加而增加。此外, 相对于气候温暖湿润的怒江流域, 比较干燥寒冷的澜沧江流域中细菌beta多样性更高, 其均值为0.735, 随海拔距离变化的斜率也更大, 为0.14/km (> 0.12/km)。对于周转组分, 也表现出同样的规律, 澜沧江流域细菌周转组分随海拔变化的斜率为0.14/km (> 0.13/km)。在距离衰减模型中, 更高的斜率代表着群落的物种组成是更强烈的环境选择的结果(Hanson et al, 2012)。这表明, 尽管距离衰减是一个普遍现象, 但其强度可能因气候而异。
我们使用MRM探究了各种因子对beta多样性及其周转和嵌套组分的影响。结果表明, 海拔与细菌总beta多样性和周转组分具有最高的相关性(表2, R2 = 0.408, P < 0.001)。表明海拔是影响细菌群落物种组成的主要因素。此外, 其他环境因子和气候因子对于细菌beta多样性和其周转过程也有较强的相关性。环境因子中, 总氮、金属离子含量, 水深, pH等对beta多样性有较强的作用, 这一结果表明溪流生态系统受环境因子的调控(Heino et al, 2014)。我们的发现与之前的研究结果相似, 例如在水体化学因子中, pH值对细菌的影响最大, 而栖息地因素的重要性则较小(Lear et al, 2009); 营养盐(例如总氮)会限制细菌的生物量, 因此总氮与细菌多样性多呈显著的相关性(Carr et al, 2005)。气候因子中, 大部分因子都与细菌beta多样性和其周转过程有较强的相关性。但是从图2我们可以看出, 不同的气候条件下, 细菌的beta多样性随海拔梯度的变化模式并没有发生改变。我们认为气候变化会影响到当地细菌的物种组成, 但并不能影响细菌在两个流域中的空间分布格局。这也表明细菌对于气候变化的适应能力比较强, 所以气候发生变化时, 细菌可以很好地适应环境(Wang et al, 2012)。
本文的其它图/表
|

{kind=link}