



生物多样性下降已成为全球现象(Ripple et al, 2017; Ceballos et al, 2020), 尤其是陆地大型食肉动物(31种)由于栖息地丧失和破碎化、猎物缺乏和偷猎等, 超过75%的物种种群持续下降, 面临局部或完全灭绝的危险(Dirzo et al, 2014; Ripple et al, 2014b)。随着大型食肉动物的广泛衰退, 在众多生态系统产生了营养级联效应, 导致其所处生态系统的结构和功能发生变化(Estes et al, 2011; Ripple et al, 2014a; Winnie & Creel, 2017)。虎(Panthera tigris)是陆地生态系统体型最大的食肉动物之一和最濒危的大型猫科动物, 作为生物多样性保护的旗舰物种, 在维护生态系统功能和服务中发挥着关键作用。虎起源于距今300-200万年前, 但成为一个物种之后, 其有效种群数量持续下降; 在距今100-11万年间一直维持在5-8万只。从距今11万年前的末次冰期开始, 由于气候的往复波动以及7.5万年前印度尼西亚苏门答腊岛多巴火山的超级爆发, 亚洲部分地区的虎灭绝, 有效种群数量持续下降至今, 但同时也促成了虎从中国西南和东南亚一带向外扩散, 最终形成9个亚种(Liu et al, 2018)。虎位于食物链顶端, 必须有充足的大型猎物和大面积完整的栖息地才能满足其生存需要, 因此, 对栖息地丧失和破碎化等干扰非常敏感(Ripple et al, 2014b; Wolf & Ripple, 2017)。近1个世纪以来, 野生虎种群数量下降了96%, 仅存的种群分布在76个虎保护景观(Tiger Conservation Landscapes, TCLs, 面积118万km2)中, 其面积不到历史范围的7%, 4个亚种野外灭绝, 幸存的5个亚种均面临不同程度的威胁或灭绝风险(Dinerstein et al, 2007; Seidensticker, 2010; Goodrich et al, 2015)。从20世纪70年代开始, 包括中国在内的多个国家开展了各种保护行动, 包括建立保护区、恢复栖息地、颁布捕猎禁令和反盗猎等, 进入21世纪, 各国的保护初见成效, 部分亚种数量有所回升, 但是大部分种群仍分布在较小的、隔离的栖息地内, 已显示出明显的种群地理遗传结构(Kenney et al, 2014), 野生虎保护迫在眉睫。
虎具有很强的适应力, 能够适应广泛的气候条件、生态系统和猎物种类(Schaller, 1967; Sunquist, 2010), 曾经广布于亚洲大部分地区, 活动区域海拔可高达4,500 m (Tempa et al, 2019)。研究显示在减少人类干扰的前提下, 野生虎种群存在自行恢复的潜力(Harihar et al, 2009; Walston et al, 2010; Wikramanayake et al, 2011; Lamichhane et al, 2018)。正是在这个前提下, 2010年亚洲13个虎分布国在俄罗斯圣彼得堡召开世界虎峰会(Tiger Summit 2010), 制定了全球虎恢复“TX2计划”, 提出保护栖息地、打击盗猎和非法贸易、跨境合作执法、与当地社区合作、提升野生虎种群和栖息地管理有效性、历史分布地重引入6项主要保护措施, 并正式提出下一个虎年即2022年使全球野生虎数量翻倍的目标, 这一目标表明了改变野生虎命运所需要的最大的雄心和承诺(Global Tiger Initiative Secretariat, 2011)。
为了实现这一目标, Walston等(2010)提出了“6%解决方案” (the six percent solution), 认为恢复野生种群的关键在于景观中的“源点” (source sites), 这里的源点指的是能生存至少25只繁殖雌虎并且嵌入周边景观后能够支持至少50只繁殖虎种群的栖息地, 识别的42个源点仅占虎现存栖息地的6% (约90,000 km2), 却拥有近70%的野生虎(2,154只)。与Walston等(2010)的源点保护策略不同, Wikramanayake等(2011)提出实现野生虎数量翻倍的“景观保护策略”, 认为保护工作和资金应分配到所有的TCLs, 加强栖息地连通性和保护集合种群至关重要。由于所有TCLs中没有任何种群可繁殖的个体超过250只(Dinerstein et al, 2007), 当前保护工作的重点已从单一的保护区转向由质量和连通水平不同的栖息地组成的大尺度异质景观(Vasudev et al, 2017)。总之, 野生虎的恢复需要确保核心繁殖种群或源点的安全, 同时保持源点和周围恢复区之间景观尺度上的连接(Walston et al, 2010; Wikramanayake et al, 2011; Harihar et al, 2018), 所以二者保护的理念是一致的。
自2010年世界虎峰会召开已12年, 不同方案的保护成效如何? 虎还面临哪些新威胁? 在2022年中国农历虎年之际, 也是野生虎保护的重要时间节点上, 本文通过梳理近几十年的文献, 总结了野生虎每个亚种的研究进展、种群状态和主要致危因素, 评价了已有研究重点与空缺, 最后为未来亚洲虎种群的研究和保护提出了建议。
1 野生虎种群研究进展
我们使用“tigers (or Panthera tigris)”作为检索词在Web of Science数据库和谷歌学术搜索引擎进行检索, 共检索到1946年1月至2021年10月的各类文献2,246篇, 逐条删除与虎无关和无法找到原文或摘要的文献后, 筛选出1,584篇。按照研究对象分类, 去除未指明具体亚种的260篇和已灭绝亚种的10篇后, 以孟加拉虎(P. t. tigris)为研究对象的有576篇, 东北虎(P. t. altaica) 345篇, 苏门答腊虎(P. t. sumatrae) 90篇, 马来虎(P. t. jacksoni) 20篇, 印支虎(P. t. corbetti) 60篇, 华南虎(P. t. amoyensis) 41篇, 同时182篇研究对象多于1个亚种。以研究领域分类,生理生化方向的119篇, 行为的52篇, 生态与保护的728篇, 圈养繁育的64篇, 遗传的170篇, 疾病的297篇, 其他154篇(图1)。
图1
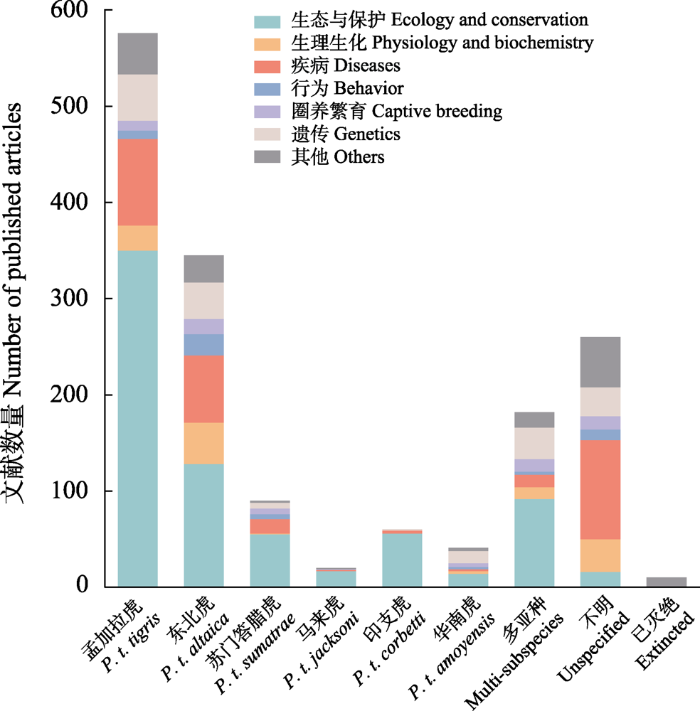
图1
20世纪40年代以来按亚种和类别发表的虎文献统计
Fig. 1
Number of published tiger (Panthera tigris) articles by subspecies and category since 1940s
孟加拉虎具有较长的研究历史, 占文献总数的43.8%, 其次为东北虎, 而受制于分布国经济水平和战争等原因, 苏门答腊虎、马来虎和印支虎的相关研究都较少, 尤其是马来虎, 仅有20篇, 华南虎的研究数量少可能是因为这个亚种已野外灭绝, 仅有少数野外灭绝前的早期文献记录和针对圈养个体的研究。作为野生动物保护的明星物种之一, 虎的生态与保护方向的研究最多, 而其余的研究也多为保护和恢复服务, 例如遗传方向的许多文章关注亚种的基因多样性、种群间的基因流等。数量仅次于生态与保护的疾病方向和生理生化的研究大多数集中于动物园的圈养个体, 与圈养繁育方向有一定程度的重叠, 例如圈养动物之间的传染疾病、圈养动物的生理指标水平等。行为方向研究数量的稀少可能是因为野生虎密度低、行踪隐蔽, 同时作为独居的大型动物倾向于主动远离人类, 使野外种群的行为研究必须借助于项圈等设备, 存在一定难度和局限。
2 野生虎种群状态和分布
虎属于脊椎动物亚门哺乳纲食肉目猫科豹属(Panthera), 最近共同祖先可追溯至大约11.2万年前的晚更新世, 自晚更新世以来存在两次主要扩散事件, 第一次是距今11-7万年前苏门答腊虎与大陆上的虎分离, 从中南半岛进入苏门答腊岛, 同时少数中国西南部避难种群进入华南地区; 第二次发生在距今5-1万年前, 是其他8个虎亚种主要的形成时期, 亚种间基因流程度低, 均具有独特的进化史(Liu et al, 2018)。目前, 巴厘虎(P. t. balica)、爪哇虎(P. t. sondaica)、里海虎(P. t. virgata)和华南虎4个亚种于20世纪40-80年代相继野外灭绝。根据世界自然保护联盟(International Union for Conservation of Nature, IUCN)的估计, 2010年亚洲野生虎总数不足3,200只(Goodrich et al, 2015), 但在俄罗斯圣彼得堡虎保护峰会上13个虎分布国将种群数量基线调整至3,633只, 并承诺到2022年种群数量达到5,845只(附录1)。近期文献和各国评估报告显示目前全球5个虎亚种的数量已增加到大约4,694只(图1, 附录1), 只有印度、不丹和俄罗斯完成了预期目标。2022年IUCN发布的红色名录显示全世界仅有4,485只虎(不包括幼崽)生活在野外(其中, 成年虎3,140只) (Goodrich et al, 2022), 虎在缅甸的大部分森林已经消失(Lynam et al, 2006), 在马来西亚种群数量也迅速减少(Darmaraj, 2019), 在越南、柬埔寨和老挝3个国家已经野外灭绝(O’Kelly et al, 2012; Rasphone et al, 2019)。虽然近年来没有确切的证据, 但朝鲜也可能有虎分布。尽管过去的12年中虎种群在部分保护区中逐步恢复, 但是由于非法盗猎、猎物减少和栖息地退化等威胁的持续存在, 保护区外种群下降趋势仍很明显, 大部分TCLs进一步破碎和退化, 特别是在东南亚地区(见后文)。Walston等(2010)识别的源点中, 有2个已经丧失, 其中一个来自泰国, 另一个来自老挝, 但是最初未被列入源点的不丹已经记录了> 50只的雌虎种群(Tempa et al, 2019)。Harihar等(2018)评估了研究和保护基础较好的18个源点的野生虎种群及其栖息地现状, 结果显示这18个点的野生虎数量仍未饱和(现共有165只, 有潜力容纳585只), 特别是其中15个点仍然处于虎数量恢复的初级阶段, 因为这些地区猎物的恢复需要30-50年。这是一个缓慢的过程, 这也表明其他国家将很难完成2022年预期目标。由于各亚种所处的地理环境不同, 猎物种类和密度差异极大, 以及人类的干扰导致虎的密度在不同亚种和国家间异质性非常高(表1, 附录2), 如猎物丰富的南亚超过10只/100 km2, 而猎物密度低的东北亚不到1只/100 km2 (附录2)。
表1 不同国家和不同亚种虎密度(只/100 km2)估计
Table1
| 平均值 Average | 最大值 Max. | 最小值 Min. | 中位值 Median | 标准误 Standard error | |
|---|---|---|---|---|---|
| 国家 Country | |||||
| 孟加拉国 Bangladesh | 4.14 | 7.00 | 2.40 | 4.20 | 0.59 |
| 不丹 Bhutan | 1.70 | 3.70 | 0.24 | 1.42 | 0.82 |
| 中国 China | 0.28 | 0.32 | 0.24 | 0.28 | 0.04 |
| 印度 India | 3.45 | 14.00 | 0.19 | 2.20 | 0.39 |
| 印度尼西亚 Indonesia | 1.67 | 2.80 | 0.74 | 1.56 | 0.44 |
| 马来西亚 Malaysia | 1.28 | 1.95 | 0.61 | 1.28 | 0.67 |
| 缅甸 Myanmar | 1.20 | - | - | - | - |
| 尼泊尔 Nepal | 2.43 | 5.40 | 0.92 | 0.97 | 0.94 |
| 俄罗斯 Russia | 0.42 | 0.60 | 0.25 | 0.42 | 0.10 |
| 泰国 Thailand | 1.20 | 2.10 | 0.36 | 1.25 | 0.27 |
| 亚种 Subspecies | |||||
| 孟加拉虎 P. t. tigris | 3.37 | 14.00 | 0.03 | 2.39 | 0.33 |
| 东北虎 P. t. altaica | 0.37 | 0.60 | 0.24 | 0.32 | 0.07 |
| 苏门答腊虎 P. t. sumatrae | 1.67 | 2.80 | 0.74 | 1.56 | 0.44 |
| 印支虎 P. t. corbetti | 1.20 | 2.10 | 0.36 | 1.20 | 0.23 |
| 马来虎 P. t. jacksoni | 1.28 | 1.95 | 0.61 | 1.28 | 0.67 |
2.1 孟加拉虎
现存5个亚种中孟加拉虎的数量最多, 大约占野生虎总数的70%, 主要分布在印度、尼泊尔、不丹和孟加拉国, 在缅甸和中国西南也有少量跨境分布。1758年, 孟加拉虎被瑞典自然学家卡尔·林奈定为虎的模式种, 20世纪由于栖息地丧失和盗猎等原因数量剧减至1,800只左右, 目前经过保护总数恢复至近3,500只, 最大的种群分布在印度, 约为2,967只, 其次为尼泊尔235只、不丹103只、孟加拉国114只(Dhakal et al, 2014; DoFPS, 2015; DNPWC & DFSC, 2018; Jhala et al, 2021)。印度的野生孟加拉虎种群平均密度在3.45 (0.19-14)只/100 km2 (表1), 其中不同分布区的密度存在极大差异, 每100 km2的孟加拉虎数量可能从不足1只到大于10只(Wildlife Institute of India, 2021) (附录2)。尼泊尔的野生孟加拉虎种群平均密度为2.43 (0.92-5.40)只/100 km2, 其中最高密度曾出现在巴迪亚国家公园(Bardia National Park) (5.40只/100 km2), 最低的为帕萨国家公园(Parsa National Park) (0.92只/100 km2) (DNPWC & DFSC, 2018)。2014-2015年的调查显示不丹的种群密度在0.23只/100 km2左右(Tempa et al, 2019)。孟加拉国仅有的种群分布于和印度接壤的孙德尔本斯三角洲红树林, 孟加拉国在2015和2018年分别组织了两次调查, 显示当地孟加拉虎数量在这3年中上升了约8%, 目前总数在114只左右, 种群密度为2.55只/100 km2 (Aziz et al, 2017b, 2019)。我国与印度接壤的西藏地区一直有孟加拉虎活动记录(张恩迪等, 2002; 王渊等, 2019)。王渊等(2019)于2013-2018年的调查显示, 墨脱县仅存1-3只孟加拉虎非定居个体, 但缺乏影像证据。直到2019年, Li等(2021)在西藏墨脱县第一次拍摄到孟加拉虎的影像, 之后其他研究者在藏南地区发现当地至少有11只虎, 这些信息表明中印边境可能存在一个小种群。
2.2 东北虎
东北虎是分布于最北部高纬度地区的一个亚种, 基因组中与脂肪代谢和能量生成相关的基因显示出正选择的迹象, 表明此亚种对寒冷环境的适应(Armstrong et al, 2021)。历史上, 东北虎广泛分布于我国东北、俄罗斯远东和朝鲜半岛, 仅100多年种群数量就从20世纪初的2,500多只骤降至2000年初的500只左右(田瑜等, 2009)。目前东北虎主要形成了3个相对独立的种群, 最大的种群位于俄罗斯远东的锡霍特-阿林山脉, 这里的种群在20世纪40年代仅剩20-30只(Henry et al, 2009), 截至2015年已恢复到至少500只(包括100只幼崽), 相比于2005年增长了15% (WWF-Russia & Vladivostok, 2016), 近十几年来多次监测到有个体从这个大种群扩散到我国的完达山(Qi et al, 2021)。东北虎的第2个种群位于犹太自治州(Jewish Autonomous Region)和阿穆尔州(Amur Region), 由俄罗斯自2013年起救助放归的野生个体组成, 目前这个成功重建的种群数量已超过20只(Rozhnov et al, 2021), 并已扩散到小兴安岭和大兴安岭(Zhou et al, 2022), 为该区域东北虎种群恢复提供了希望。我国境内的东北虎种群在2000年左右仅剩12-16只个体, 受益于黑龙江和吉林东部(老爷岭)中俄跨境种群的保护, 自2010年以来表现出明显的恢复趋势, 2012年8月至2014年7月期间, 北京师范大学在现在的东北虎豹国家公园东部共监测到至少26只东北虎(15只成体, 5只亚成体和6只幼崽) (Wang et al, 2016)。经与毗邻的俄罗斯豹地国家公园(Land of the Leopard National Park)联合监测显示, 中俄跨境小种群个体数量至少38只, 是东北虎的第3个主要种群(Feng et al, 2017); 尽管该种群正以每3年大约13 km的速度沿中俄边境向中国内部扩散(Ning et al, 2019), 但长期与俄罗斯的大种群隔离(Sorokin et al, 2016)。
Qi等(2021)的研究显示2013-2018年中国境内红外相机记录到野生东北虎个体数量至少54只(包括20只幼崽), 平均雌雄性比0.73, 低于大多数虎种群的1.5或2 (Sugimoto et al, 2012; Miquelle et al, 2015), 表明此种群以雄性个体为主。所有出现点的总占用生境面积约为4.8万km2, 主要分布在老爷岭、张广才岭、完达山、小兴安岭4个景观斑块内, 但约90%的个体和全部的繁殖个体出现在老爷岭, 也就是现在的东北虎豹国家公园。2014年以来, 我们记录到2只个体分别是从老爷岭和完达山迁移到张广才岭, 表明景观之间的连通性对未来种群恢复的重要性(Wang et al, 2015)。东北虎豹国家公园东部沿中俄边境区虎密度为0.20-0.27只/100 km2 (Xiao et al, 2016; Wang et al, 2018), 远低于俄罗斯锡霍特-阿林保护区稳定种群的1只/100 km2 (Miquelle et al, 2015)。因此, 尽管在中国境内有广泛的栖息地, 并经过了10多年的集约化管理(清除猎套、猎物恢复、巡护和系统监测), 但虎密度仍然比俄罗斯稳定种群低, 制约东北虎种群在中国恢复和扩散的一个主要因素是大型有蹄类猎物(如梅花鹿Cervus nippon和马鹿C. elaphus)的空间分布不均, 猎物物种多样性较低(Wang et al, 2018; Qi et al, 2021)。
2.3 苏门答腊虎
苏门答腊虎仅分布于印度尼西亚苏门答腊岛的热带雨林中, 是苏门答腊岛3个虎亚种中仅存的1种, 另外2个亚种爪哇虎和巴厘虎均已灭绝。苏门答腊虎是所有虎亚种中体型最小的, 这有可能与苏门答腊岛上缺乏大型有蹄类猎物有关, 基因组数据显示与体型发育相关的ADH7基因已在该亚种中显示出强烈的正选择信号(Liu et al, 2018)。随着天然林的消失, 苏门答腊虎的种群数量不断减少, 20世纪70年代总数在1,000只左右(Borner, 1978), 到了2000年成年个体总数已经下降至742只, 2012年的调查显示成年个体总数约为618只, 分布于苏门答腊岛15个破碎的景观中(Luskin et al, 2017), 种群平均密度为1.67 (0.74-2.80)只/100 km2 (表1)。
2.4 马来虎
马来虎是2004年从印支虎中独立出来的新亚种, 为马来半岛的特有物种, 主要分布在马来半岛中部和南部(Luo et al, 2004; Shevade et al, 2017)。20世纪中叶马来虎数量高达3,000只, 盗猎和栖息地丧失让这一亚种的数量剧减, 2014年前后调查显示马来虎的总数在250-340只, 其中可繁殖的成年个体数量约为80-120只(Kawanishi, 2015)。马来西亚早在2009年开始实施马来西亚国家虎保护行动计划(NTCAP), 其预期目标是到2020年使该国的虎数量增加到1,000只, 然而, 2016-2020年的全国种群调查显示当前马来虎总数不到150只(WWF-Malaysia, 2022)。极度濒危的马来虎正处于种群崩溃的中间阶段, 未来10年存在高度的野外灭绝风险。
2.5 印支虎
印支虎曾经广泛分布于中南半岛, 但是目前已从东南亚大部分国家消失, 仅分布于泰国、缅甸和中缅边境。缅甸的印支虎种群从2010年的80只左右下降到目前仅约20只, 只分布在少数保护区内。近期在Htamanthi保护区的调查显示种群密度仅为0.81只/100 km2 (Moo et al, 2018; Naing et al, 2019)。泰国是印支虎在东南亚最后的据点, 2010年泰国的印支虎数量为190-250只(Pisdamkham et al, 2010), 而2016年泰国政府调查报告显示数量仅为101-128只(DNP, 2016), 共2个存活种群分布于泰国与缅甸交界的西部森林景观综合体(25,000 km2) (western forest complex, WEFCOM)和东部的东巴耶延山-考爱山森林景观综合体(6,155 km2) (Dong Phayayen- Khao Yai Forest Complex, DPKYCOM) (Ash et al, 2021b)。WEFCOM的核心区会卡肯野生生物保护区(Huai Kha Khaeng Wildlife Sanctuary, HKK) (19,000 km2)是印支虎保护的核心区(> 55只), 2005-2012年这里的虎种群密度变化范围为1.27-2.09只/100 km2 (Duangchantrasiri et al, 2016)。在过去的10年里, 已有虎开始从HKK向周边的保护区扩散, 表明这里已成为野生虎扩散的种源区。Phumanee等(2021)对WEFCOM北部的嵋翁国家公园(Mae Wong National Park)和隆兰国家公园(Khlong Lan National Park)的调查共监测到10只成年虎(4雄和6雌)和6只幼崽, 至少有2只成体来自HKK, 密度约为0.36只/100 km2。Ash等(2020)对DPKYCOM的调查显示印支虎总数为20 (14-33)只, 密度为0.63 (0.32-1.21)只/100 km2。可见, HKK之外的保护区印支虎密度仍然非常低, 远低于东南亚常见的2只/100 km2, 主要是因为大型猎物稀缺, 恢复缓慢。我国学者于2007年在西双版纳国家级自然保护区用红外监测相机拍下了野生印支虎的图像(冯利民等, 2013), 但是目前云南西南部和缅甸接壤的保护区内尚未有明确的调查统计。
3 野生虎种群致危因素
随着亚洲人口的快速增长和社会经济的发展, 人类对于自然资源的需求与日俱增, 森林采伐、毁林垦荒等活动对原始森林植被造成了严重的破坏, 直接导致了虎的栖息地的丧失和退化, 加上盗猎的影响, 自1997和2005年以来虎占域范围已分别下降了53%和20% (Goodrich et al, 2022)。破碎化的栖息地将野生虎分割为多个孤立的小种群, 缺乏基因交流使小种群面临遗传多样性下降和近交衰退的风险(Mondol et al, 2013; Kenney et al, 2014; Sorokin et al, 2016)。森林丧失和与家畜的竞争也使大型野生有蹄类猎物数量和空间分布快速下降, 造成了虎的猎物的缺乏(Feng et al, 2021a; Jornburom et al, 2020)。虽然各国均颁布了相关的法律和保护措施, 但是盗猎和非法贸易对虎及其猎物种群的威胁仍然存在。亚洲人口数量和道路密度的不断上升使人类及家畜与虎接触的机会显著增加, 使保护区管理、廊道建设和人虎冲突管理难度上升, 人虎的频繁接触亦使疾病(尤其是犬瘟热)在野生动物与家畜间蔓延(Inskip & Zimmermann, 2009; Goodrich, 2010; Kawanishi et al, 2014; Gilbert et al, 2020)。除此之外, 生态旅游的兴起也对野生虎种群产生新的干扰, 为保护工作带来新的挑战(Tyagi et al, 2019)。
3.1 栖息地丧失和退化
虎是典型的山地林栖物种, 栖息的生境从东北亚的温带落叶阔叶林和针阔混交林, 到南亚和东南亚的热带雨林、常绿阔叶林和红树林。栖息地的质量对于野生虎种群至关重要, 作为伏击型的捕猎者, 虎需要茂密的林下植被作为捕猎的掩护(Sunarto et al, 2012)。例如, 孙德尔本斯三角洲地区的孟加拉虎绝大多数的捕猎地点位于红树林而不是开阔的海岸(Khan & Chivers, 2007), 因此, 虎常对茂密的植被表现出明显的偏好, 避开开阔和植被受到破坏(如林下放牧导致的开阔生境)的区域(Wang et al, 2016; Bhattarai & Kindlmann, 2018; Sarkar et al, 2018)。在过去的100年里, 虎已经从亚洲西南部和中部、两个印尼岛屿(爪哇和巴厘岛)以及东南亚和东亚的大部分地区消失。亚洲是一个人口密集、发展迅速的地区, 这给虎生存所需的大片完整栖息地带来了巨大的压力, 而森林的持续破坏和转为农牧业用地、商业采伐、居民点和道路建设一直是虎栖息地丧失和破碎的主要原因, 目前破坏仍在继续, 甚至出现在过去很少受影响的亚洲高地(Yang et al, 2022)。2001-2014年间76个TCLs中约有8万km2的森林消失, 相比于2000年减少了7.7%, 其中位于保护区内的森林减少2.2万km2 (5.7%); 29个优先TCLs中约有5.8万km2森林消失, 相比于2000年减少6.9%; Walston等(2010)提出的6%解决方案中的源点已丧失1,676 km2的森林, 单个源点的森林丧失面积最大为435 km2, 森林大面积的丧失导致至少400只虎失去潜在栖息地(Joshi et al, 2016)。受世界林产品市场和国家财政政策变化以及农业集约化和扩张的影响, 南亚和东南亚的虎栖息地丧失尤为严重(尤其是柬埔寨) (Dasgupta et al, 2014; Hu et al, 2021; Namkhan et al, 2021), 早在1996-2006年间已减少41% (Sanderson, 2006), 但在2001-2014年里, TCLs的森林又减少超过7万km2, 远高于东北亚森林的丧失(约9,000 km2) (Joshi et al, 2016)。Forrest等(2011)关于TCLs内土地管理成效的研究显示, 只有21%的区域受到法律保护, 同时受到监管、预算和执行方面的限制, 保护区的管理效率普遍较差, TCLs的森林很大一部分有资源开采(木材、石油和天然气、矿产等)的特许经营权。
东南亚地区的毁林一直被认为是印支虎、马来虎和苏门答腊虎数量下降和灭绝的重要原因之一, 尤其是近期虎在越南、柬埔寨和老挝的灭绝(Dasgupta et al, 2014; Namkhan et al, 2021); 该地区是油棕(Elaeis guineensis)、橡胶(Hevea brasiliensis)等热带植物产品和木材的出口地区, 采伐和经济作物种植是导致原始森林植被减少和生物多样性丧失的主要原因(Wilcove et al, 2013)。2001-2019年间东南亚地区平均每年丧失3.22万km2森林, 且2010年后森林加速消失, 并从低地森林向更高海拔的陡坡地区转移, 有可能将热带森林转变为重要的净碳源(Feng et al, 2021b)。Luskin等(2017)对苏门答腊虎的研究发现, 2000-2012年间大约21.1%的天然林转为油棕林, 虎种群数量下降了16.6%, 密度下降了31.9%, 仅有2个景观繁殖虎的数量大于25只。如果森林持续转为人工林, 未来几十年或更短的时间内将很难有虎可持续生存(Poor et al, 2019)。
亚洲是受道路影响而导致局部濒危物种数量最多的地区之一(Ceia-Hasse et al, 2017; Quintana et al, 2022)。路网的全球扩张对野生动物存在多方面影响, 包括栖息地丧失和破碎化、阻碍个体扩散和基因交流, 以及带来的路杀和盗猎的风险, 另外道路的噪音、灯光能降低栖息地的质量等(Carter et al, 2020; Ash et al, 2021a; Quintana et al, 2022)。Quintana等(2022)评估了道路对所有顶级食肉动物的灭绝风险, 虎是仅次于懒熊(Melursus ursinus)的受道路威胁最大的物种之一, 主要的威胁包括遗传多样性的下降(Sorokin et al, 2016; Thatte et al, 2018)和非法盗猎的增长(Sharma et al, 2014)。Kerley等(2002)通过GPS项圈技术研究了道路对东北虎个体生存率的影响, 在为期9年的研究中, 在无道路地区活动的2只成年雌性个体全部存活, 而生活在一级道路地区的6只成年雌性全部死亡或者消失, 道路地区的幼崽成活率也显著低于无道路地区。目前在所有76个TCLs道路总长达13.4万km, 总密度达162.4 m/km2, 大约43%的虎繁殖区和57%的TCLs处于道路影响严重的地区, 已导致虎和猎物的数量减少20%以上(Carter et al, 2020)。在中国“一带一路”倡议等重大投资项目的刺激下, 到2050年, 将有近2.4万km的新建道路位于TCLs内, 预期包括虎在内的更多物种会暴露在更高的道路密度影响下。目前, 根据IUCN的物种红色名录评估, 虎没有被列为受交通基础设施威胁的物种。Palmeirim和Gibson (2021)的研究显示南亚和东南亚现有421座水坝, 共淹没了13,750 km2虎的栖息地。随着水电基础设施在未来几十年的扩建, 在虎的分布区将有41座计划中的大坝准备开工, 大多数水坝都将建设于优先保护区内; 据估算, 这将至少影响已知全球野生虎种群的1/5 (占20.8%-22.8%, 729只虎)。
3.2 猎物缺乏
健康的大型有蹄类猎物是否充足与野生虎种群的生存最密切(Karanth et al, 2004), 尤其是大型鹿科动物在虎的食谱中具有不可替代的地位, 几乎所有野生虎种群分布区域内均有一到多种大型鹿科动物生存(Sunquist et al, 1999; Hayward et al, 2012)。Hayward等(2012)对不同亚种食性的研究显示, 虎对于野猪(Sus scrofa)、水鹿(Cervus unicolor)、马鹿和沼鹿(C. duvauceli)存在显著的偏好。这些偏好猎物对虎种群生存力至关重要, 它们在提供充足食物的同时降低了虎的捕猎风险, 且能够满足雌虎喂养幼崽的需求(Miller et al, 2014)。但在不同区域, 虎捕食的大型猎物种类和偏好不同。在印度和尼泊尔, 虎主要的猎物包括白斑鹿(Axis axis)、水鹿、印度野牛(Bos gaurus)和野猪等(Bagchi et al, 2003; Bhattarai & Kindlmann, 2018; Aziz et al, 2020; Krishnakumar et al, 2022)。中俄边境东北虎小种群捕食最多的则是野猪和梅花鹿(Kerley et al, 2015)。在泰国的会卡肯野生生物保护区, 当地两种主要猎物印度野牛和爪哇野牛(B. javanicus)被捕食个体的平均体重分别是397.9 kg和423.9 kg, 成年野牛在虎的猎物中占到了近半数(Khaewphakdee et al, 2020)。
作为大型肉食动物, 虎对猎物的需求量较大, 以东北虎为例, 一只成年雄性平均每天需要的食物最少为5.2 kg, 非繁殖育幼期的成年雌性为每日3.9 kg, 而抚育4只幼崽的雌性每日需要的食物量约为11.4 kg (Miller et al, 2014)。Zhang等(2013)的研究发现, 当地猎物生物量低于195 kg/km2时便无法支持东北虎种群的生存。Sandom等(2017)在全球范围内的研究发现, 多种大型猫科动物的数量下降与当地猎物数量的减少存在强烈的正相关关系。Barber- Meyer等(2013)对尼泊尔TAL景观地区(Terai Arc Landscape)的研究也发现孟加拉虎种群的减少与猎物的缺乏存在显著的关系。Carter等(2019)的研究发现, 猎物减少导致孟加拉虎领地面积增加, 从而导致环境能承载的孟加拉虎总数减少; 同时领地面积增加意味着捕猎消耗的能量增加, 能导致死亡率上升。猎物数量的下降也会导致捕食者需要增加捕猎距离, 增加了与人发生冲突甚至遭遇路杀等事故的风险(Wolf & Ripple, 2016)。
Mahmood等(2021)对亚洲大型动物群的研究发现, 大型有蹄类动物的分布区域均在骤减, 即使在许多保护区中也出现了大型食肉动物缺失的情况。虎猎物缺乏的主要原因是人类砍伐森林导致的栖息地丧失、盗猎以及放牧带来的竞争压力。Kawanishi等(2014)对马来虎的主要猎物水鹿的研究发现森林采伐直接导致了水鹿栖息地的丧失, 是其数量在20世纪下降的最主要因素, 而采伐修建的道路又为之后的盗猎提供了便利, 在21世纪盗猎成为水鹿的最大威胁。泰国西部森林景观复合体的研究显示, 当地的大型野生有蹄类, 包括水鹿、印度野牛和爪哇野牛的数量由于盗猎和栖息地破碎化大量减少, 仅少量分布于适宜栖息地, 即使在保护区的森林中, 由于大型有蹄类数量的下降, 虎种群数量难以增长(Jornburom et al, 2020)。Zhang等(2013)对完达山脉的调查发现, 2002-2008年间, 由于高强度的捕猎, 当地的3种有蹄类种群(马鹿、狍Capreolus pygargus和野猪)数量下降了40%-45%, 严重威胁了东北虎的长期生存。放牧活动也可能导致家畜与野生有蹄类形成竞争, 导致虎的猎物种群数量下降(Chundawat et al, 1999; Nayak et al, 2013; Wolf & Ripple, 2016)。Wang等(2016)和Feng等(2021a)对东北虎和猎物的研究发现, 当地高强度的放牧活动与大型有蹄类猎物形成了较强的竞争, 作为东北虎主要猎物的野猪、梅花鹿和狍均对牛表现出了时空上的回避或多度下降。此外, 自2018年底以来, 亚洲爆发的非洲猪瘟疫情对野猪种群的生存产生了新的威胁, 而其对虎种群生存的影响需要今后的持续关注(Luskin et al, 2021)。
3.3 人虎冲突
人虎冲突(human-tiger conflicts)是指人与虎中任何一方对另外一方的不利影响, 当虎对当地社区的生计和安全造成威胁时, 就会发生人虎冲突(Pettigrew et al, 2012)。由于人类活动不断向森林深处扩张并挤压野生动物的生存空间, 虎攻击牲畜和人的事件呈现上升趋势, 并可能会导致邻近社区对虎的报复性捕杀, 给保护和恢复带来持续威胁(Inskip & Zimmermann, 2009; Goodrich, 2010)。人类活动对虎捕食的干扰导致虎需要更大的领地来满足捕猎的需求, 对孟加拉虎等多种野生动物的研究均显示, 即使猎物数量相似, 在人类干扰强度更高的地区, 动物的领地范围更大, 与人类相遇和发生冲突的几率随之增加(Carter et al, 2019)。人类活动造成的猎物缺乏可能导致虎转而捕食家畜, 促使人虎冲突进一步加剧。当野生猎物种群的生物量密度低于812.41 ± 1.26 kg/km2时, 大型猫科动物倾向于攻击体型较大的家畜, 如家牛(Bos taurus), 当野生猎物生物量密度继续降低至544.57 ± 1.19 kg/km2以下时则开始攻击体型更小的牲畜, 如绵羊(Ovis aries)和山羊(Capra aegagrus var. hircus) (Khorozyan et al, 2015)。例如, 东北虎豹国家公园长期林下放牧活动显著降低了该地区有蹄类猎物的多度, 尤其是牛排除了虎最主要的猎物梅花鹿(Feng et al, 2021a), 猎物密度过低使虎转而捕食家畜, 已导致严重的人虎冲突, 2008-2014年间公园内就发生了超过370次虎捕食家牛的事件(Wang et al, 2016)。
人对冲突的感受会影响对野生动物的包容程度, 会决定人是否采取报复性猎杀行动(Lyngdoh et al, 2014)。在人口密度较高的南亚和东南亚地区, 例如, 印度、尼泊尔和印度尼西亚等国, 伤人和伤畜事件频发。1978-1997年间, 印度尼西亚的苏门答腊虎共致死146人, 致伤30人, 杀死至少870头家畜(Nyhus & Tilson, 2004); 尼泊尔奇旺国家公园(Chitwan National Park)的孟加拉虎在2007-2014年间共致死32人, 致伤22人, 杀死了至少351只家畜(Dhungana et al, 2018; Bhattarai et al, 2019); 2005-2011年间印度中部地区的孟加拉虎袭击人事件有103起(Dhanwatey et al, 2013)。即使是人口密度低的俄罗斯锡霍特-阿林山脉, 2000-2009年间平均每年也发生1.9起伤人事件(Goodrich et al, 2011)。大型野生动物的伤人和伤畜行为会带来经济损失, 当负面影响不断累积, 就会影响公众对动物保护工作的支持态度。当个体的态度在强度上达到一定程度, 就会做出相应的行为而影响动物, 如报复性猎杀(Khorozyan et al, 2015), 尤其是在经济较不发达、缺少政府补偿机制的地区。例如, 印度尼西亚Kerinci Seblat国家公园的调查显示, 2000-2005年间约有3只成年虎和4只幼虎死于捕捉牲畜时农民的驱赶以及报复性捕杀(Nugraha & Sugardjito, 2009)。另外, 人虎冲突可能促使村民加入盗猎或者为盗猎提供帮助, 获取报酬以弥补牲畜的损失(Bhattarai et al, 2019)。综上, 频发的伤人事件使得人虎冲突恶化, 盗猎和报复性捕杀对保护工作提出巨大挑战(Goodrich, 2010)。
3.4 盗猎
以虎的皮毛、骨头、肉和身体其他部位等的非法交易为目的的偷猎是野生虎面临的主要威胁, 导致部分地区野生种群大量丧失甚至国家尺度上(如老挝)的灭绝(Goodrich et al, 2015; Rasphone et al, 2019)。亚洲大约有100万km2潜在适宜栖息地没有虎定居(Sanderson et al, 2006), 表明偷猎是对野生虎种群及猎物的最大威胁之一(Chapron et al, 2008)。Robinson等(2015)的研究发现1992-2012年间盗猎是俄罗斯境内东北虎死亡的最主要原因, 造成每年种群数量减少17%-19%。马来西亚的Belum Temengor地区, 在2008-2019年间死亡的45只野生虎个体中, 有39只死于盗猎(Ten et al, 2021)。理论模型表明当繁殖雌性的年死亡率超过15%时, 它们就会灭绝, 即使有充足的猎物, 种群仍难以恢复(Chapron et al, 2008)。
目前, 非法盗猎对虎及其猎物和其他大型兽类的威胁仍然存在并且呈上升趋势, 在东南亚地区尤其严重(Lynam, 2010; Johnson et al, 2016; Gray et al, 2018; Benítez-López et al, 2019), 甚至保护区都无法杜绝(Risdianto et al, 2016; Aziz et al, 2017a)。盗猎的动机包括炫耀、彰显身份地位和有罪不罚、腐败、贫穷、野生肉类和医药产品的需求, 以及人兽冲突等(Harrison et al, 2016; Skidmore, 2022)。Campbell等(2019)对印度尼西亚Kerinci Seblat国家公园的研究发现, 2012-2015年间猎套数量和分布范围显著增加, 表明当地的盗猎活动日趋猖獗, 并由于经费的限制使得执法力度有限, 即使设立了保护区仍然不能将盗猎的威胁彻底清除。WWF 2021年发布的《无声的陷阱: 东南亚猎套危机》(Silence of the Snares: Southeast Asia’s Crisis)显示(Belecky & Gray, 2020), 2005-2019年间在5个东南亚国家(柬埔寨、印度尼西亚、老挝、马来西亚、越南)的11个保护区共清除猎套371,856个(每年清除约53,000个), 据估计, 在柬埔寨、老挝、越南的保护区中猎套约有1,230万个, 密度超过110个/km2。盗猎行为不但导致大型兽类快速消失, 推动空林(empty forest)形成(Benítez-López et al, 2019), 同时增加人兽冲突和人畜共患疾病的机会(Johnson et al, 2020)。
3.5 传染病
野生虎可能感染的病原体包括猫冠状病毒(feline coronavirus, FCoV)、猫细小病毒(feline parvovirus, FPV)、犬细小病毒(canine parvovirus, CPV)、犬瘟热病毒(canine distemper virus, CDV)、弓形虫(Toxoplasma gondii)和脑囊尾蚴(Cerebral cysticercosis)等(Goodrich et al, 2012; McCauley et al, 2021; Nur-Farahiyah et al, 2021; Phuentshok et al, 2021)。以往认为传染病并不是常见的威胁虎种群长期生存的主要因素, 但是近年来随着多个亚种感染CDV并致死案例的逐渐增加, 该疾病成为威胁虎的又一重要因素, 甚至危及到亚种的存亡。俄罗斯境内2000-2004年间在6只东北虎个体中检测出CDV, 截止到2013年, CDV所造成的东北虎死亡数量已占总数量(大约450只)的1% (Seimon et al, 2013; Gilbert et al, 2014, 2020)。2019年以来马来虎已有2起CDV确诊病例被报道(Ten et al, 2021); Mulia等(2021)对1988-2016年间野外捕捉的21只苏门答腊虎进行了CDV检测, 发现其中43%的个体为病毒中和抗体阳性, 表明该亚种CDV暴露的时间很早。
CDV是一种近乎全球分布的多宿主单链RNA病毒, 属于副黏病毒科麻疹病毒属(Morbillivirus), 拥有较高的致死率, 可在猫科、犬科、熊科、鼬科等动物之间传播, 被认为是对包括虎在内的食肉动物威胁最大的一种高流行性病毒(Martinez- Gutierrez & Ruiz-Saenz, 2016; 吴桥兴等, 2021)。犬瘟热疫情爆发已经被证实与多种大型食肉动物的衰退或濒临灭绝有关, 包括塞伦盖蒂狮(Panthera leo) (Roelke-Parker et al, 1996)和埃塞俄比亚狼(Canis simensis) (Gordon et al, 2015)。Robinson等(2015)研究发现1992-2012年间CDV是仅次于盗猎而造成东北虎死亡的重要原因。该病毒最初会引起呼吸道和胃肠道疾病, 但可以进一步感染中枢神经系统, 导致行为异常, 例如失去对人的恐惧和方向感, 并丧失捕猎的能力, 野外已观测到虎感染CDV后被车辆撞死以及雌虎抛下幼崽离开的行为(Miquelle et al, 2015)。
CDV传播的主要路径是通过含有病毒粒子的呼吸道分泌物、其他体液、排泄物和分泌物(如尿液、粪便)等形成气溶胶, 并通过呼吸道、眼结膜等途径进入易感动物体内, 潜伏期约7 d, 传染期可达60-90 d, 病毒可在25℃存活48 h, 5℃存活14 d (Pollock, 1982), 在家犬中较为盛行, 例如, 东北虎分布区中狗的CDV血清阳性率高达16%-41% (Gilbert et al, 2020)。强烈的人类干扰使家犬广泛侵入虎的栖息地, 包括保护地, 极大地促进了CDV在家犬与食肉动物和有蹄类动物之间跨种传播并形成传染池, 如狗獾(Meles meles)、貉(Nyctereutes procyonoides)、赤狐(Vulpes vulpes)、紫貂(Martes zibellina)、梅花鹿和野猪等这些易感物种通过被捕食和直接接触等途径感染虎, 病毒的多寄主特性和高变异性使得疾病爆发后往往难以得到控制, 对小种群的危害最大(Gilbert et al, 2020)。种群生存力分析显示, 在种群数量不大于25只的虎种群中, CDV造成其50年内灭绝的概率高达56% (Gilbert et al, 2014)。即使是稍大的种群也会面临风险, 俄罗斯锡霍特-阿林保护区在1966年仅有虎3-4只, 2005年恢复到38只, 但截止到2012年数量又降至9只, 其中2009-2011年间12只死亡的东北虎中, 7只与CDV有关(Miquelle et al, 2015)。
3.6 旅游
虎的观光和旅游业是发展中国家重要的产业和经济来源, 同时也是宣传野生虎保护和筹集资金的重要渠道(Thapa et al, 2017), 但是以野生动物为主题的旅游产业一直充满争议, 需要更充分的研究和谨慎的管理。首先, 道路和其他相关设施的修建可能破坏野生动物栖息地, Bindra等(2010)和Rastogi等(2015)在印度科比特保护区(Corbett Tiger Reserve)的研究发现, 旅游开发、基础设施建设和垃圾污染等对虎栖息地植被的破坏和生态廊道的阻断产生了负面影响。其次, 人类噪音可能对动物产生干扰, Tyagi等(2019)对班达加(Bandhavgarh National Park)和坎哈(Kanha National Park)两个国家公园孟加拉虎粪便样本的糖皮质激素分析显示, 在旅游高峰期, 人声和车辆的干扰导致孟加拉虎应激水平显著上升, 可能对虎的健康和种群状况产生影响。同样, 非洲象(Loxodonta africana)的研究也表明游客和车辆导致了非洲象糖皮质激素分泌水平的上升和攻击、预警等行为的增加(Szott et al, 2019a, b)。短期的生理变化和行为改变经过长期积累可能对动物的活动如捕食和繁殖产生重要的影响, 除此之外和人类的频繁接触同样会带来疾病的风险, 以及路杀等问题(Higginbottom, 2004)。
3.7 小种群与近交衰退
近交(inbreeding)是指具有亲缘关系的个体之间交配繁殖。近交可导致个体成活率、繁殖力、种群遗传多样性和健康下降, 从而降低物种适应环境变化的能力, 是导致物种濒危的重要遗传机制之一(魏辅文等, 2021)。自然界的物种存在从小到大的连续分布变化, 并具有不同的隔离程度, 但许多濒危兽类常以孤立小种群形式存在(Haddad et al, 2015), 同时还可能经历过严重的历史瓶颈, 加重近交衰退的危害。理论(Lande, 1994; Lynch et al, 1995)和实证研究(Burrows et al, 1994)都表明, 孤立小种群灭绝风险最大, 如非洲野狗(Lycaon pictus) (Woodroffe & Sillero-Zubiri, 2020)和埃塞俄比亚狼(Marino & Sillero-Zubiri, 2011)。保护生物学的小种群范式(small population paradigm)表明, 由于近交衰退和种群随机性, 孤立小种群有害等位基因固定的概率大, 易于进入灭绝旋涡(extinction vortex)的进程。近交个体能表达有害的、部分隐性的等位基因, 这些等位基因通过血缘关系从亲本遗传而来, 导致近交衰退(Charlesworth & Charlesworth, 1987), 降低种群的平均适合度, 最终导致种群规模缩小, 并可能导致灭绝(Keller & Waller, 2002)。
亚洲所有虎种群历史上都经历过严重的种群瓶颈, 基因组多态性水平较低(Luo et al, 2004, 2019)。随着栖息地破碎化和人类影响加剧, 当前虎种群自20世纪70年代以来一直处于隔离状态(Kenney et al, 2014), 大部分种群生活在破碎化的景观中(< 100个个体); 种群之间的基因流被人类干扰景观阻断, 面临遗传多样性下降、近交衰退和遗传漂变的威胁(Armstrong et al, 2021)。印度野生孟加拉虎种群的全基因组数据显示孤立小种群存在清除有害突变的机制, 然而, 如果种群长期处在数量比较小的状况下, 有害突变最终会逐渐累积, 能出现持续的近交衰退(Khan et al, 2021)。印度西北部伦滕波尔(Ranthambhor)国家公园内, 62只虎生活的保护区周围都是居民区, 由于长期没有其他个体输入新的基因流, 缺乏避开近亲繁殖的机制, 半数个体具有一个同样的母系祖先(Sharma et al, 2013; Armstrong et al, 2021)。即使是遗传多样性相对较高的孟加拉虎近期得到了恢复, 但在印度次大陆大部分种群仍分布在周边人口密度极高的隔离保护区, 近期显示出明显的遗传分化, 说明其具有明显的系统地理分布格局(Sharma et al, 2013; Armstrong et al, 2021)。Mondol等(2013)的研究显示, 印度次大陆的孟加拉虎历史上存在大量线粒体DNA变异, 由于近代生境的持续破碎化和连通性下降, 93%的线粒体单倍型在现代种群中丢失, 导致了遗传变异的丧失和种群之间遗传分化的显著增加。
遗传变异低的小型孤立种群受遗传漂变的影响, 可快速发生表型进化。例如, 印度东部西密里波保护区(Similipal Natural Reserve)的虎祖先范围小, 并由于地理上长期与其他种群隔离, 遗传漂变和近亲繁殖导致Taqpep基因发生突变, 超过1/3个体发生伪黑化症(pseudomelanistic), 俗称黑虎(Sagar et al, 2021)。小种群的近交压力也与个体健康直接相关。东北虎从其他亚种中分出的时间最晚, 遗传多样性最低, 特别是位于俄罗斯滨海边疆区西南部以及我国老爷岭一带的跨境种群与俄罗斯锡霍特-阿林山脉的大种群长期隔离(Dou et al, 2016; Sorokin et al, 2016; Luo et al, 2019), 已呈现中等近交水平。近期对中国一侧的种群遗传分析显示种群中50%的个体为表亲或者半血亲, 23%的个体近交系数为中或高值, 近交系数和肠道寄生虫数量、肠道菌群均表现出了强烈的相关性(Ning et al, 2022)。
4 虎的研究和保护建议
虎是亚洲森林生态系统的旗舰物种, 它的未来取决于亚洲各国政府通过保护大片适宜的栖息地并保持栖息地的连通性来创造有效的TCLs。首先, 源点地区栖息地和猎物恢复以及虎源种群保护最为紧急(Walston et al, 2010), 这是因为源点地区承载了绝大多数的野生繁殖个体, 然而当前许多源点受到威胁, 极大地限制了种群的增长和扩散潜力(Harihar et al, 2018)。其次, 恢复源点地区(包括保护区)之间的连通性、扩大保护区面积、提升已有保护区的管理和执法水平、周边社区积极参与保护工作等对亚洲各国虎种群的恢复至关重要(Harihar et al, 2020; Goodrich et al, 2022)。最后, 消除虎及其猎物的偷猎和非法贸易、疾病管控、减少保护区内外栖息地的丧失和退化以确保TCLs间功能连通性和资金投入等都是实现成功保护的关键。
4.1 有蹄类猎物的恢复
亚洲有足够的森林栖息地支持野生虎数量翻倍(Joshi et al, 2016), 但这一潜力被大部分地区的猎物缺失所限制(Harihar et al, 2018; Mahmood et al, 2021)。虎平均每年吃掉其领地内10%的猎物(Sunquist, 1981; Karanth et al, 2004), 一只成年雄性每年需要4,000 kg的食物(55-60只中等大小有蹄类猎物, 下同), 而没有幼崽的雌性每年需要3,000 kg食物(40-45只) (Sunquist, 1981), 养育3只幼崽的雌性每年消耗4,500 kg食物(60-75只) (Sunquist, 1981; Karanth et al, 2004)。在缺乏大型猎物的情况下, 即使小型猎物物种相当丰富, 野生虎种群也无法长期维持和繁殖(Sunquist, 1981; Karanth & Sunquist, 1995)。亚洲部分景观已进行了多年有力的保护(如泰国东部的森林综合体和我国的东北虎豹国家公园) (Duangchantrasiri et al, 2016; Wang et al, 2016), 但保护区内外虎密度的差异表明, 野生虎种群在景观尺度上的恢复是一个缓慢的过程, 即使是在生态连通度很高的地区可能也需要几十年的时间才能实现。要将大部分景观的虎密度提高到与保护好的源点相匹配的水平(如2只/100 km2), 需要加强巡逻以限制偷猎, 以及更直接的干预措施来增加猎物, 如改善有蹄类动物的栖息地质量、减少林下大型家畜过牧的竞争和加强非洲猪瘟疫情的控制等。对于东北虎的潜在栖息地, 应继续实施现有的保护措施(如反偷猎和清山清套活动、有蹄类猎物冬季的食物补充等), 以促进东北虎种群和大型猎物(如野猪、梅花鹿和马鹿)的恢复, 特别是在我国目前唯一的东北虎繁殖区老爷岭森林景观地区。只有恢复大型有蹄类猎物的种群数量, 才有可能增加虎种群的数量, 并促进个体向其他景观扩散和定居, 在这一方面尼泊尔野生虎种群成功恢复的案例为亚洲各国提供了可借鉴的经验(Lamichhane et al, 2018)。
4.2 栖息地连通性恢复和降低人类干扰
遗传变异对物种的生存和适应环境变化至关重要, 恢复和维持种群之间的基因连通性是避免近交衰退和局部灭绝的重要手段。模拟研究表明, 促进亚种内的基因流动可能比简单的鼓励本地种群扩张能够更有效地保存物种的遗传多样性(Bay et al, 2014; Thatte et al, 2018)。尤其是对于人口密度很高的印度次大陆和东南亚地区来说, 维持森林廊道是比恢复大片完整栖息地更现实和有效的手段(Harihar et al, 2020)。印度中部生存着1,000多只孟加拉虎, 自2014年来增长了50%。这里分布的虎数量超过整个印度虎数量的1/3 (Jhala et al, 2020), 依靠森林廊道虎种群具有很高的历史和当代基因流率, 维持着很高的遗传多样性(Sharma et al, 2012; Natesh et al, 2017)。如果森林廊道遭到破坏, 将导致杂合度降低35%和保护区内虎平均灭绝概率提高56%, 在这种情况下, 增加虎数量仅会减少12%的灭绝概率(从56%减少到44%)。Walston等(2010)识别的42个潜在源点种群中, 超过70% (30)都处于小种群状态(繁殖雌虎 < 12), 另外, 76个TCLs中, 超过50%也处于小种群状态, 这些小种群可能已经处于近交灭绝阈值的早期阶段。模拟研究显示, 对于小种群即使每一代有1-4只扩散的雄虎也不能提高种群生存力, 30年内灭绝的可能性超过90% (Kenney et al, 2014)。因此, 为了减少局部灭绝的风险, 目前仍保有野生虎分布的国家需要做出长期的政策承诺, 保持森林覆盖以及保护区和周围景观之间的连通性以增加可繁殖种群的规模, 另外需要对破碎化的栖息地构建生态廊道以增加种群之间的基因交流。考虑到亚洲人口规模和未来持续的经济发展, 基础设施开发、大坝建设和其他类似的项目要进行详细的土地利用规划, 优先考虑包括虎在内的生物多样性的保护。同样, 我国东北虎种群亟需有效的管理措施以提高不同栖息地的生态连通性, 并将其纳入中俄跨境景观的发展规划, 虎才会恢复(Dou et al, 2016; Wang et al, 2016)。成功的保护依赖于人类干扰的大幅减少, 除了需要加大对盗猎的管控和执法力度之外, 需要为当地居民提供保护工作岗位以增加其从野生动物保护中获得的收益, 需要发展生态服务型经济以帮助牧民转业, 从而降低人虎冲突。
图2
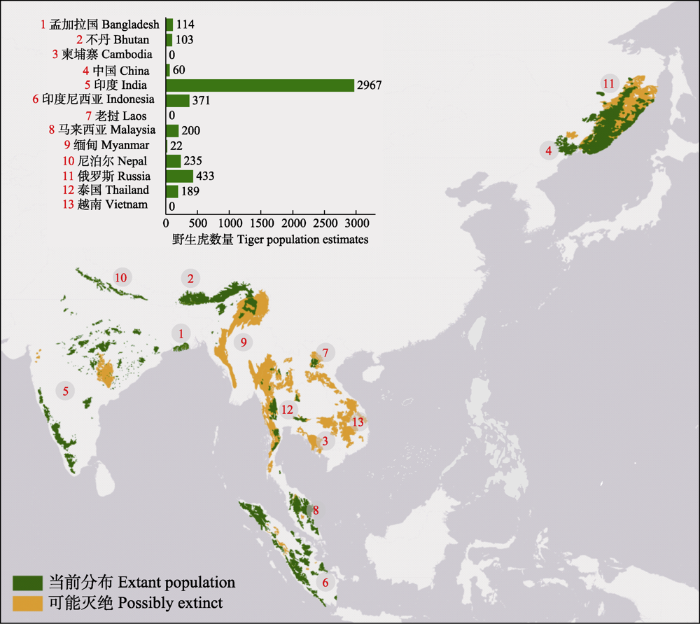
图2
亚洲野生虎的分布范围和种群数量(数据来源见附录1)。图中的数字代表不同的国家。
Fig. 2
Current geographic distribution and population estimates of wild tigers (Panthera tigris) in Asia (data sources are provided in the Appendix 1). The numbers in the figure represent various countries.
4.3 个体重引入
重引入(reintroduction)作为一种重要的保护手段, 可以有效地增加野生种群数量, 缓解近交繁殖压力, 如佛罗里达美洲狮(Puma concolor coryi)和猎豹(Acinonyx jubatus)种群恢复经验强调了重引入在种群管理中的重要作用(Johnson et al, 2010; Buk et al, 2018)。在虎局部灭绝的地区, 个体重新引入是一种可行的恢复方法, 虽然这绝非易事, 但在印度和俄罗斯已有成功的再引入案例(Anuradha et al, 2016; Sarkar et al, 2016)。类似的重引入计划可以帮助中国、越南、柬埔寨、老挝、中亚等国家或地区进行野生种群的重建和恢复(Chestin et al, 2017; Gray et al, 2017)。然而, 大型食肉动物的重新引入往往是困难和有争议的。“问题动物”的失败很容易引发当地人的负面态度, 逆转已有的保护成果。消除偷猎、恢复猎物种群和当地社区的准备工作等是重引入的必要前提, 可能需要10-20年的努力。另外, 重引入后的种群恢复通常是一个缓慢的过程(Miquelle et al, 2015; Duangchantrasiri et al, 2016), 可能需要长达数十年的持续支持才能成功。因此, 重新引入计划必须考虑成功所需的长期财政、后勤和人力资源的需求(Gray et al, 2017)。同时在实施基因拯救之前, 对当地种群和重引入个体遗传背景的调查也是必要的, 一方面能够避免不同亚种之间不必要的基因污染, 另一方面以解决遗传多样性缺失和近交衰退问题为目标时可以考虑选取遗传背景最接近的另一亚种进行重引入, 需要对不同种群根据具体情况确定最有利于保护目标的政策。东北虎种群在中国的恢复前景尚不明朗, 老爷岭以外的景观中重引入东北虎雌性个体, 对于快速建立繁殖种群, 从而维持这些斑块中种群的生存意义重大, 所以对其重新引入的相关研究和准备可能是必要的。老爷岭景观内生存的东北虎已处于中等近交衰退阶段, 可考虑通过俄罗斯大种群的雌性个体再引入来降低有害突变的频率, 并增加有害等位基因位点的杂合度, 从而提高平均适合度。
4.4 疾病控制
栖息地和种群的破碎化极大地增加了人和家犬与野生动物接触的机率, 使得野生虎感染各种传染病的可能性大大增加(Gupta et al, 2021)。未来对于各种疾病, 尤其是CDV影响的研究应成为野生虎保育的重点方向之一。目前针对虎感染CDV的研究主要在中国和俄罗斯东北虎种群中展开, 针对其他地区的研究程度不一。考虑到不同亚种都不同程度地暴露于CDV, 首先, 亟需开展流行病学研究以确定虎的CDV宿主, 这对于制定适当的缓解战略, 包括为宿主物种或虎本身接种疫苗的可能性至关重要。Gilbert等(2020)的研究表明, 利用各种捕获机会为野生虎接种疫苗能够显著降低CDV导致的种群灭绝风险。其次, 需要建立结合CDV流行病学的种群生存力模型, 以探讨不同感染情景的影响, 并确定处于高风险的种群。另外, 对收集的尿液、死前样本(全血、血清、结膜拭子)和死后样本(包括脑组织、淋巴结、肺和膀胱)进行CDV检测应成为常规(Seimon et al, 2013)。最后, 管控疾病的传播途径是必要的。以CDV为例, 尽管家犬不是虎感染CDV的唯一途径, 但是给家犬接种疫苗并避免进入保护区, 可以减少传播途径, 有效减少虎的CDV感染率(Gupta et al, 2021)。目前, 已有圈养虎(Ten et al, 2021)、野生印支豹(Panthera pardus fusca) (Mahajan et al, 2022)和北美野生白尾鹿(Odocoileus virginianus)等动物感染新冠病毒(SARS-CoV-2)的案例(Chandler et al, 2021), 考虑到新冠病毒的全球流行趋势, 该病毒是否也会成为野生虎一个潜在的威胁因素, 需要密切监测。
4.5 反盗猎
偷猎不仅会直接导致野生虎数量下降, 在未来很长的一段时间里也会影响其种群遗传多样性, 而遗传多样性的降低将会使虎在面对疾病爆发、气候变化、生境丧失等问题时变得更加脆弱, 增加其灭绝风险。禁猎法律的颁布是虎保护历史上重要的一步, 苏联1947年的捕猎禁令是将东北虎从灭绝边缘拯救的关键(Chapron et al, 2008)。所有虎分布国在禁猎上已经达成共识(Global Tiger Initiative Secretariat, 2011), 但是执法力度的缺失始终存在于保护工作中, 尤其是经济较为落后的东南亚国家, 所以亚洲各国政府必须改善国家的执法, 加强立法, 有效遏制猎套等行为, 并探索应用现代技术提升自然保护地反盗猎管理能力。另外, 组织社区居民广泛参与并共同开展反盗猎工作, 同时提升当地经济水平也是减轻执法压力的途径之一(Mohsanin et al, 2013)。近年来, 尼泊尔境内孟加拉虎种群恢复迅速, 仅巴迪亚国家公园的虎数量就从2009年的18只增长到2018年的87只, 这主要得益于十几年来尼泊尔在社区支持和反盗猎等方面的巨大努力(Rauniyar, 2021)。
4.6 跨境合作
跨境分布的个体在全球虎种群中占有重要的比例, 除了苏门答腊虎, 其他现存的4个亚种均存在跨境分布现象, 例如中俄边境的东北虎种群、印度与尼泊尔和孟加拉国边境的孟加拉虎种群等。由于社会、经济、文化和政治上的差异, 对跨境种群采取孤立的保护方法不是适当的措施, 所以促进所有跨境区域的生物多样性整体保护, 减少基础设施建设(如道路)导致的栖息地破碎化、维护基因交流, 对全球虎保护至关重要(Farhadinia et al, 2021)。许多国境线上布设了铁丝网、围墙等设施, 有时也是交通运输设施较为密集的地带, 这些人类设施可能对虎及其猎物的移动产生阻碍, 所以, 国家之间合作共同建立跨界廊道、跨境保护区、加强跨境研究与监测和资金投入等应成为综合保护和国际合作计划的关键组成部分(Liu et al, 2020)。跨境合作提高执法行动能力也能帮助打击跨国的盗猎和非法野生动物贸易。南亚和东南亚等国的边境地区如中-缅-老边境是生物多样性热点区, 同时也是盗猎和非法贸易的重灾区, 每年有数以吨计的非法野生动物制品在该地区流通。尼泊尔通过与我国和印度的合作达到了虎、亚洲象(Elephas maximus)和独角犀牛(Rhinoceros unicornis)的“零盗猎”目标, 证明不同国家的执法部门合作能够更好地打击盗猎和非法野生动物制品贸易活动(Acharya, 2016; Liu et al, 2020)。我国的虎种群恢复和保护计划同样依赖于国际合作。例如, 最有希望恢复的东北虎很大程度上依赖于俄罗斯的源种群, 2019年6月, 中俄发表联合声明, 已将东北虎跨界保护和廊道建设作为未来合作的重要组成部分。尽管国内孟加拉虎和印支虎分布数量极其稀少, 且主要为非定居个体, 但其热点分布区域均位于中国与周边国家的边界区域, 通过跨境合作机制的建立, 实施一定的生态保护措施, 在恢复边境区域栖息地质量和猎物种群的同时, 吸引境外个体向国内扩散并定居, 逐渐恢复国内种群是存在很大可能性的。
4.7 加强种群监测和研究
濒危物种的重建和管理需要基于科学和证据的适应性方法, 以识别和解决关键保护问题(Buk et al, 2018)。长期科学研究是保护虎的重要手段, 能够帮助研究者了解目标种群的变化情况, 掌握种群动态(存活率、增长率和补员等)、性比、食性、栖息地偏好等重要信息, 为保护政策的制定和调整提供科学的指导(Majumder et al, 2017)。由于虎种群恢复缓慢但下降的速度可能很快, 因此准确、实时的种群动态监测信息是有效保护的重要前提, 我国东北虎的保护和恢复受益于有效的调查方法和基于红外相机的长期监测平台的建设, 特别是实时监测系统和人工智能的应用(王天明等, 2020)。目前印度、尼泊尔、不丹、孟加拉国、印度尼西亚和我国已经建立了长期监测体系, 能够定期统计境内野生虎的种群数量(或占域), 但是对其他国家的虎种群数量及其分布处于了解不足、数据缺失或者数据多年未更新的状况, 亟需大量资金投入加强虎和同域分布的其他野生动物的监测和研究。今后, 各国需将调查范围和调查方法标准化, 以准确地评估种群的时间和空间变化趋势。在对虎及其猎物种群进行动态监测的同时, 遗传信息的监测也是必要的, 尤其是在如今野生虎种群普遍面临栖息地破碎化和近交衰退威胁的情况下, 利用采集粪便等非损伤性的取样方法对种群的遗传多样性和近交程度的监测对保护工作的开展也具有重要的指导意义(Dou et al, 2016; Ning et al, 2022)。
附录 Supplementary Material
附录1 2010年俄罗斯圣彼得堡世界虎峰会和近期虎在13个分布国的种群数量估计
Appendix 1 Tiger populations of range countries as declared at the St. Petersburg Summit in 2010 and most recent estimates
附录2 不同国家与亚种虎的密度列表
Appendix 2 List of population density estimates of wild tigers (Panthera tigris) from various countries and subspecies
参考文献
A Walk to Zero Poaching for Rhinos in Nepal
Significance of mate selection and adult sex ratio in tiger reintroduction/ reinforcement programs
DOI:10.1111/jzo.12331 URL [本文引用: 1]
Recent evolutionary history of tigers highlights contrasting roles of genetic drift and selection
DOI:10.1093/molbev/msab032
PMID:33592092
[本文引用: 4]

Species conservation can be improved by knowledge of evolutionary and genetic history. Tigers are among the most charismatic of endangered species and garner significant conservation attention. However, their evolutionary history and genomic variation remain poorly known, especially for Indian tigers. With 70% of the world's wild tigers living in India, such knowledge is critical. We re-sequenced 65 individual tiger genomes representing most extant subspecies with a specific focus on tigers from India. As suggested by earlier studies, we found strong genetic differentiation between the putative tiger subspecies. Despite high total genomic diversity in India, individual tigers host longer runs of homozygosity, potentially suggesting recent inbreeding or founding events, possibly due to small and fragmented protected areas. We suggest the impacts of ongoing connectivity loss on inbreeding and persistence of Indian tigers be closely monitored. Surprisingly, demographic models suggest recent divergence (within the last 20,000 years) between subspecies and strong population bottlenecks. Amur tiger genomes revealed the strongest signals of selection related to metabolic adaptation to cold, whereas Sumatran tigers show evidence of weak selection for genes involved in body size regulation. We recommend detailed investigation of local adaptation in Amur and Sumatran tigers prior to initiating genetic rescue.© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Estimating the density of a globally important tiger (Panthera tigris) population: Using simulations to evaluate survey design in Eastern Thailand
DOI:10.1016/j.biocon.2019.108349 URL [本文引用: 1]
Environmental factors, human presence and prey interact to explain patterns of tiger presence in Eastern Thailand
DOI:10.1111/acv.12631 URL [本文引用: 1]
Opportunity for Thailand’s forgotten tigers: Assessment of the Indochinese tiger Panthera tigris corbetti and its prey with camera-trap surveys
DOI:10.1017/S0030605319000589 URL [本文引用: 1]
Phylogeography of Panthera tigris in the mangrove forest of the Sundarbans
DOI:10.3354/esr01188 URL [本文引用: 1]
Spatial differences in prey preference by tigers across the Bangladesh Sundarbans reveal a need for customised strategies to protect prey populations
DOI:10.3354/esr01052 URL [本文引用: 1]
Status of Tiger in the Sundarban of Bangladesh 2018
Investigating patterns of tiger and prey poaching in the Bangladesh Sundarbans: Implications for improved management
DOI:10.1016/j.gecco.2016.12.001 URL [本文引用: 1]
Using non-invasively collected genetic data to estimate density and population size of tigers in the Bangladesh Sundarbans
DOI:10.1016/j.gecco.2017.09.002 URL [本文引用: 1]
Prey abundance and prey selection by tigers (Panthera tigris) in a semi-arid, dry deciduous forest in western India
DOI:10.1017/S0952836903003765 URL [本文引用: 1]
Influence of prey depletion and human disturbance on tiger occupancy in Nepal
DOI:10.1111/j.1469-7998.2012.00956.x URL [本文引用: 1]
A call for tiger management using “reserves” of genetic diversity
DOI:10.1093/jhered/est086 URL [本文引用: 1]
Intact but empty forests? Patterns of hunting-induced mammal defaunation in the tropics
Human disturbance is the major determinant of the habitat and prey preference of the Bengal tiger (Panthera tigris tigris) in the Chitwan National Park, Nepal
DOI:10.2478/eje-2018-0002 URL [本文引用: 2]
Managing human-tiger conflict: Lessons from Bardia and Chitwan National Parks, Nepal
DOI:10.1007/s10344-019-1270-x URL [本文引用: 2]
Conservation of severely fragmented populations: Lessons from the transformation of uncoordinated reintroductions of cheetahs (Acinonyx jubatus) into a managed metapopulation with self-sustained growth
DOI:10.1007/s10531-018-1606-y URL [本文引用: 2]
Demography, extinction and intervention in a small population:The case of the Serengeti wild dogs
Two species, one snare: Analysing snare usage and the impacts of tiger poaching on a non-target species, the Malayan tapir
DOI:10.1016/j.biocon.2019.01.009 URL [本文引用: 1]
Road development in Asia: Assessing the range-wide risks to tigers
Effects of human-induced prey depletion on large carnivores in protected areas: Lessons from modeling tiger populations in stylized spatial scenarios
DOI:10.1002/ece3.5632
[本文引用: 2]
Prey depletion is a major threat to the conservation of large carnivore species globally. However, at the policy-relevant scale of protected areas, we know little about how the spatial distribution of prey depletion affects carnivore space use and population persistence. We developed a spatially explicit, agent-based model to investigate the effects of different human-induced prey depletion experiments on the globally endangered tiger (Panthera tigris) in isolated protected areas-a situation that prevails throughout the tiger's range. Specifically, we generated 120 experiments that varied the spatial extent and intensity of prey depletion across a stylized (circle) landscape (1,000 km(2)) and Nepal's Chitwan National Park (1,239 km(2)). Experiments that created more spatially homogenous prey distributions (i.e., less prey removed per cell but over larger areas) resulted in larger tiger territories and smaller population sizes over time. Counterintuitively, we found that depleting prey along the edge of Chitwan National Park, while decreasing tiger numbers overall, also decreased female competition for those areas, leading to lower rates of female starvation. Overall our results suggest that subtle differences in the spatial distributions of prey densities created by various human activities, such as natural resource-use patterns, urban growth and infrastructure development, or conservation spatial zoning might have unintended, detrimental effects on carnivore populations. Our model is a useful planning tool as it incorporates information on animal behavioral ecology, resource spatial distribution, and the drivers of change to those resources, such as human activities.
Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction
Global exposure of carnivores to roads
DOI:10.1111/geb.12564 URL [本文引用: 1]
SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus)
The impact on tigers of poaching versus prey depletion
DOI:10.1111/j.1365-2664.2008.01538.x URL [本文引用: 3]
Inbreeding depression and its evolutionary consequences
DOI:10.1146/annurev.es.18.110187.001321 URL [本文引用: 1]
Tiger re-establishment potential to former Caspian tiger (Panthera tigris virgata) range in Central Asia
DOI:10.1016/j.biocon.2016.11.014 URL [本文引用: 1]
Tigers in Panna: Preliminary results from an Indian tropical dry forest
Vyāghranomics in space and time: Estimating habitat threats for Bengal, Indochinese, Malayan and Sumatran tigers
DOI:10.1016/j.jpolmod.2014.01.014 URL [本文引用: 2]
Large carnivore attacks on humans in central India: A case study from the Tadoba-Andhari Tiger Reserve
DOI:10.1017/S0030605311001803 URL [本文引用: 1]
Living with tigers Panthera tigris: Patterns, correlates, and contexts of human-tiger conflict in Chitwan National Park
The fate of wild tigers
DOI:10.1641/B570608 URL [本文引用: 2]
Defaunation in the Anthropocene
DOI:10.1126/science.1251817
PMID:25061202
[本文引用: 1]
We live amid a global wave of anthropogenically driven biodiversity loss: species and population extirpations and, critically, declines in local species abundance. Particularly, human impacts on animal biodiversity are an under-recognized form of global environmental change. Among terrestrial vertebrates, 322 species have become extinct since 1500, and populations of the remaining species show 25% average decline in abundance. Invertebrate patterns are equally dire: 67% of monitored populations show 45% mean abundance decline. Such animal declines will cascade onto ecosystem functioning and human well-being. Much remains unknown about this "Anthropocene defaunation"; these knowledge gaps hinder our capacity to predict and limit defaunation impacts. Clearly, however, defaunation is both a pervasive component of the planet's sixth mass extinction and also a major driver of global ecological change. Copyright © 2014, American Association for the Advancement of Science.
Practical Plan to Improve Tiger Population 2015-2035 (20 Years)
Counting the Tigers in Bhutan: Report on the National Tiger Survey of Bhutan 2014-2015
Dynamics of a low-density tiger population in Southeast Asia in the context of improved law enforcement
DOI:10.1111/cobi.12655
PMID:27153529
[本文引用: 3]
Recovering small populations of threatened species is an important global conservation strategy. Monitoring the anticipated recovery, however, often relies on uncertain abundance indices rather than on rigorous demographic estimates. To counter the severe threat from poaching of wild tigers (Panthera tigris), the Government of Thailand established an intensive patrolling system in 2005 to protect and recover its largest source population in Huai Kha Khaeng Wildlife Sanctuary. Concurrently, we assessed the dynamics of this tiger population over the next 8 years with rigorous photographic capture-recapture methods. From 2006 to 2012, we sampled across 624-1026 km(2) with 137-200 camera traps. Cameras deployed for 21,359 trap days yielded photographic records of 90 distinct individuals. We used closed model Bayesian spatial capture-recapture methods to estimate tiger abundances annually. Abundance estimates were integrated with likelihood-based open model analyses to estimate rates of annual and overall rates of survival, recruitment, and changes in abundance. Estimates of demographic parameters fluctuated widely: annual density ranged from 1.25 to 2.01 tigers/100 km(2), abundance from 35 to 58 tigers, survival from 79.6% to 95.5%, and annual recruitment from 0 to 25 tigers. The number of distinct individuals photographed demonstrates the value of photographic capture-recapture methods for assessments of population dynamics in rare and elusive species that are identifiable from natural markings. Possibly because of poaching pressure, overall tiger densities at Huai Kha Khaeng were 82-90% lower than in ecologically comparable sites in India. However, intensified patrolling after 2006 appeared to reduce poaching and was correlated with marginal improvement in tiger survival and recruitment. Our results suggest that population recovery of low-density tiger populations may be slower than anticipated by current global strategies aimed at doubling the number of wild tigers in a decade.© 2015 Society for Conservation Biology.
Trophic downgrading of planet earth
DOI:10.1126/science.1205106
PMID:21764740
[本文引用: 1]
Until recently, large apex consumers were ubiquitous across the globe and had been for millions of years. The loss of these animals may be humankind's most pervasive influence on nature. Although such losses are widely viewed as an ethical and aesthetic problem, recent research reveals extensive cascading effects of their disappearance in marine, terrestrial, and freshwater ecosystems worldwide. This empirical work supports long-standing theory about the role of top-down forcing in ecosystems but also highlights the unanticipated impacts of trophic cascades on processes as diverse as the dynamics of disease, wildfire, carbon sequestration, invasive species, and biogeochemical cycles. These findings emphasize the urgent need for interdisciplinary research to forecast the effects of trophic downgrading on process, function, and resilience in global ecosystems.
Big cats in borderlands: Challenges and implications for transboundary conservation of Asian leopards
DOI:10.1017/S0030605319000693 URL [本文引用: 1]
Collaboration brings hope for the last Amur leopards
Population status of the Indochinese tiger (Panthera tigris cobetti) and density of the three primary ungulate prey species in Shangyong Nature Reserve, Xishuangbanna, China
西双版纳尚勇自然保护区野生印支虎及其三种主要有蹄类猎物种群现状调查
中国野生印支虎及其猎物种群状况的野外实地研究一直处于空白。本研究使用足迹鉴别法、粪堆计数法,首次对西双版纳尚勇自然保护区野生印支虎种群数量现状及该区域内的虎猎物种群状况进行了调查研究。结果显示:2004 ~ 2009 年间,确认西双版纳保护区存在3 只成年印支虎个体(2 雌1 雄),西双版纳尚勇保护区拥有比较丰富的有蹄类种群,其中虎的主要猎物:水鹿平均密度为7.63 (7.40 ~ 9.23)只/ km<sup>2 </sup>;赤麂平均密度为17. 39 (11.33 ~24.94)只/ km<sup>2</sup>,野猪平均密度为10.26 (7.69 ~ 14.51) 只/ km<sup>2 </sup>,该区域虎猎物生物量为1 715. 74 kg/ km<sup>2</sup> 。本研究还探讨了该区域印支虎种群的保护前景以及中国境内开展虎种群调查的适用办法等。
Effects of free-ranging livestock on sympatric herbivores at fine spatiotemporal scales
DOI:10.1007/s10980-021-01226-6 URL [本文引用: 3]
Upward expansion and acceleration of forest clearance in the mountains of Southeast Asia
DOI:10.1038/s41893-021-00738-y URL [本文引用: 1]
Single-species conservation in a multiple-use landscape: Current protection of the tiger range
DOI:10.1111/j.1469-1795.2010.00428.x URL [本文引用: 1]
Estimating the potential impact of canine distemper virus on the Amur tiger population (Panthera tigris altaica) in Russia
Distemper, extinction, and vaccination of the Amur tiger
Global Tiger Recovery Program 2010-2022
Human-tiger conflict: A review and call for comprehensive plans
DOI:10.1111/j.1749-4877.2010.00218.x
PMID:21392348
[本文引用: 3]
Human-tiger (Panthera tigris Linnaeus, 1758) conflicts (HTC), manifested primarily as attacks on people and domestic animals, exacerbate at least 2 major threats to tigers: (i) conflicts often result in mortality or removal of tigers from the wild; and (ii) they result in negative attitudes towards tigers by local people, thereby reducing support for tiger conservation. Although HTC has decreased over the past century, it will likely increase if current and proposed conservation initiatives to double tiger populations are successful. Increased HTC could undermine successful conservation initiatives if proactive steps are not taken to reduce HTC. The present paper provides a review of the impacts of HTC and the measures taken to reduce it in ways that reduce negative impacts on both humans and tigers, and stresses the need for development and implementation of comprehensive plans to reduce HTC.© 2010 ISZS, Blackwell Publishing and IOZ/CAS.
Panthera tigris
Serosurvey of free-ranging Amur tigers in the Russian Far East
Wild Amur tigers (Panthera tigris altaica, n=44) from the Russian Far East were tested for antibodies to feline leukemia virus, feline corona virus (FCoV), feline immunodeficiency virus, feline parvovirus (FPV), canine distemper virus (CDV), Toxoplasma gondii, and Bartonella henselae. Antibodies to FCoV, CDV, FPV, and T. gondii were detected in 43, 15, 68, and 42% of tigers, respectively. No differences were detected in antibody prevalence estimates between tigers captured as part of a research program and those captured to mitigate human-tiger conflicts. Domestic dogs (Canis familiaris) were tested as a potential source for CDV; 16% were vaccinated against CDV and 58% of unvaccinated dogs were antibody positive for CDV. A high percentage of tigers were exposed to potential pathogens that could affect the survival of this species. We recommend continued monitoring of wild tigers throughout Asia, development of standardized sampling and postmortem examination procedures, and additional research to better understand potential domestic and wild animal sources for these pathogens.
Conflicts between Amur (Siberian) tigers and humans in the Russian Far East
DOI:10.1016/j.biocon.2010.10.016 URL [本文引用: 1]
Panthera tigris
Canine distemper in endangered Ethiopian wolves
DOI:10.3201/eid2105.141920
PMID:25898177
[本文引用: 1]
The Ethiopian wolf (Canis simensis) is the world's rarest canid; ≈500 wolves remain. The largest population is found within the Bale Mountains National Park (BMNP) in southeastern Ethiopia, where conservation efforts have demonstrated the negative effect of rabies virus on wolf populations. We describe previously unreported infections with canine distemper virus (CDV) among these wolves during 2005-2006 and 2010. Death rates ranged from 43% to 68% in affected subpopulations and were higher for subadult than adult wolves (83%-87% vs. 34%-39%). The 2010 CDV outbreak started 20 months after a rabies outbreak, before the population had fully recovered, and led to the eradication of several focal packs in BMNP's Web Valley. The combined effect of rabies and CDV increases the chance of pack extinction, exacerbating the typically slow recovery of wolf populations, and represents a key extinction threat to populations of this highly endangered carnivore.
The wildlife snaring crisis: An insidious and pervasive threat to biodiversity in Southeast Asia
DOI:10.1007/s10531-017-1450-5 URL [本文引用: 1]
A framework for assessing readiness for tiger Panthera tigris reintroduction: A case study from eastern Cambodia
DOI:10.1007/s10531-017-1365-1 URL [本文引用: 2]
Mathematical study of the influence of canine distemper virus on tigers: An eco-epidemic dynamics with incubation delay
Habitat fragmentation and its lasting impact on Earth’s ecosystems
Recovery planning towards doubling wild tiger Panthera tigris numbers: Detailing 18 recovery sites from across the range
Responses of tiger (Panthera tigris) and their prey to removal of anthropogenic influences in Rajaji National Park, India
DOI:10.1007/s10344-008-0219-2 URL [本文引用: 1]
Demographic and ecological correlates of a recovering tiger (Panthera tigris) population: Lessons learnt from 13-years of monitoring
DOI:10.1016/j.biocon.2020.108848 URL [本文引用: 2]
Impacts of hunting on tropical forests in Southeast Asia
DOI:10.1111/cobi.12785
PMID:27341537
[本文引用: 1]
Although deforestation and forest degradation have long been considered the most significant threats to tropical biodiversity, across Southeast Asia (Northeast India, Indochina, Sundaland, Philippines) substantial areas of natural habitat have few wild animals (>1 kg), bar a few hunting-tolerant species. To document hunting impacts on vertebrate populations regionally, we conducted an extensive literature review, including papers in local journals and reports of governmental and nongovernmental agencies. Evidence from multiple sites indicated animal populations declined precipitously across the region since approximately 1980, and many species are now extirpated from substantial portions of their former ranges. Hunting is by far the greatest immediate threat to the survival of most of the region's endangered vertebrates. Causes of recent overhunting include improved access to forests and markets, improved hunting technology, and escalating demand for wild meat, wildlife-derived medicinal products, and wild animals as pets. Although hunters often take common species, such as pigs or rats, for their own consumption, they take rarer species opportunistically and sell surplus meat and commercially valuable products. There is also widespread targeted hunting of high-value species. Consequently, as currently practiced, hunting cannot be considered sustainable anywhere in the region, and in most places enforcement of protected-area and protected-species legislation is weak. The international community's focus on cross-border trade fails to address overexploitation of wildlife because hunting and the sale of wild meat is largely a local issue and most of the harvest is consumed in villages, rural towns, and nearby cities. In addition to improved enforcement, efforts to engage hunters and manage wildlife populations through sustainable hunting practices are urgently needed. Unless there is a step change in efforts to reduce wildlife exploitation to sustainable levels, the region will likely lose most of its iconic species, and many others besides, within the next few years.© 2016 Society for Conservation Biology.
Prey preferences of the tiger Panthera tigris
DOI:10.1111/j.1469-7998.2011.00871.x URL [本文引用: 2]
In situ population structure and ex situ representation of the endangered Amur tiger
DOI:10.1111/j.1365-294X.2009.04266.x
PMID:19555412
[本文引用: 1]
The Amur tiger (Panthera tigris altaica) is a critically endangered felid that suffered a severe demographic contraction in the 1940s. In this study, we sampled 95 individuals collected throughout their native range to investigate questions relative to population genetic structure and demographic history. Additionally, we sampled targeted individuals from the North American ex situ population to assess the genetic representation found in captivity. Population genetic and Bayesian structure analyses clearly identified two populations separated by a development corridor in Russia. Despite their well-documented 20th century decline, we failed to find evidence of a recent population bottleneck, although genetic signatures of a historical contraction were detected. This disparity in signal may be due to several reasons, including historical paucity in population genetic variation associated with postglacial colonization and potential gene flow from a now extirpated Chinese population. Despite conflicting signatures of a bottleneck, our estimates of effective population size (N(e) = 27-35) and N(e)/N ratio (0.07-0.054) were substantially lower than the only other values reported for a wild tiger population. Lastly, the extent and distribution of genetic variation in captive and wild populations were similar, yet gene variants persisted ex situ that were lost in situ. Overall, our results indicate the need to secure ecological connectivity between the two Russian populations to minimize loss of genetic diversity and overall susceptibility to stochastic events, and support a previous study suggesting that the captive population may be a reservoir of gene variants lost in situ.
Overview of recent land-cover changes in biodiversity hotspots
DOI:10.1002/fee.2276 URL [本文引用: 1]
Human-felid conflict: A review of patterns and priorities worldwide
DOI:10.1017/S003060530899030X URL [本文引用: 2]
Recovery of tigers in India: Critical introspection and potential lessons
DOI:10.1002/pan3.10177 URL [本文引用: 1]
Status of tigers, co-predators and prey in India, 2018
To protect or neglect? Design, monitoring, and evaluation of a law enforcement strategy to recover small populations of wild tigers and their prey
DOI:10.1016/j.biocon.2016.08.018 URL [本文引用: 1]
Genetic restoration of the Florida panther
DOI:10.1126/science.1192891
PMID:20929847
[本文引用: 1]
The rediscovery of remnant Florida panthers (Puma concolor coryi) in southern Florida swamplands prompted a program to protect and stabilize the population. In 1995, conservation managers translocated eight female pumas (P. c. stanleyana) from Texas to increase depleted genetic diversity, improve population numbers, and reverse indications of inbreeding depression. We have assessed the demographic, population-genetic, and biomedical consequences of this restoration experiment and show that panther numbers increased threefold, genetic heterozygosity doubled, survival and fitness measures improved, and inbreeding correlates declined significantly. Although these results are encouraging, continued habitat loss, persistent inbreeding, infectious agents, and possible habitat saturation pose new dilemmas. This intensive management program illustrates the challenges of maintaining populations of large predators worldwide.
Habitat use by tiger prey in Thailand’s Western Forest Complex: What will it take to fill a half-full tiger landscape?
DOI:10.1016/j.jnc.2020.125896 URL [本文引用: 2]
Tracking changes and preventing loss in critical tiger habitat
Tigers and their prey: Predicting carnivore densities from prey abundance
Prey selection by tiger, leopard and dhole in tropical forests
DOI:10.2307/5647 URL [本文引用: 1]
Panthera tigris ssp. jacksoni
Extinction process of the sambar in Peninsular Malaysia
Inbreeding effects in wild populations
DOI:10.1016/S0169-5347(02)02489-8 URL [本文引用: 1]
Effects of roads and human disturbance on Amur tigers
DOI:10.1046/j.1523-1739.2002.99290.x
PMID:35701953
[本文引用: 1]
Understanding the effects of human disturbance on endangered wildlife populations is critical to their conservation. We examined the effects of roads and human disturbance on the survivorship and foraging efficiency of Amur tigers ( Panthera tigris altaica) on and near Sikhote-Alin State Biosphere Zapovednik, Primorye Krai ( province), Russia. To evaluate the effects of roads, we estimated survivorship of radiocollared tigers and their cubs living in three types of areas: (1) areas with primary roads, (2) areas containing secondary roads, and (3) areas with minimal or no road access. We classified a tiger into one of these three treatments based on which types of roads bisected their 50% minimum convex polygon home ranges. Over a 9-year period (1991-2000), adult female survival was greatest (χ = 12.2, df = 2, p = 0.002) for radiocollared tigers in roadless areas. All adult female tigers in roadless areas survived their tenure in those locations (n = 2), whereas all died or disappeared prematurely from areas with primary roads (n = 6). Cub survival was lower in areas with primary and secondary roads than in roadless areas (χ = 10.9, df = 1, p < 0.009). We evaluated the effects of human disturbance at kill sites by examining 86 kills made by 15 tigers determining whether human disturbance had occurred at the kill site, and examining prey carcasses after tigers left, to estimate the percent meat eaten and whether the tiger abandoned the kill following human disturbance. Tigers undisturbed at kills consumed more meat ( Z = 3.71, p = 0.0002) from each kill than disturbed tigers did. Undisturbed tigers also spent more time at each kill site than disturbed tigers did ( Z = 2.3; p = 0.02). Abandonment of kills occurred in 63% of 24 instances when tigers were disturbed by people. Because roads decrease the survivorship and reproductive success of tigers, we recommend that in habitats managed for tigers, construction of new roads should be prohibited wherever possible and access to secondary roads (e.g., logging roads) should be reduced or prevented wherever possible. Protected areas seem to cease functioning as source populations where road access exists, and unprotected areas-the majority of Amur tiger range-cannot sustain stable populations with the increasing threat of human access to tiger habitat.
A comparison of food habits and prey preference of Amur tiger (Panthera tigris altaica) at three sites in the Russian Far East
DOI:10.1111/1749-4877.12135
PMID:25939758
[本文引用: 1]
Prey availability is one of the principal drivers of tiger distribution and abundance. Therefore, formulating effective conservation strategies requires a clear understanding of tiger diet. We used scat analysis in combination with data on the abundance of several prey species to estimate Amur tiger diet and preference at 3 sites in the Russian Far East. We also examined the effect of pseudoreplication on estimates of tiger diet. We collected 770 scats across the 3 sites. Similar to previous studies, we found that tigers primarily preyed on medium to large ungulates, with wild boar, roe, sika and red deer collectively comprising 86.7% of total biomass consumed on average. According to Jacobs' index, tigers preferred wild boar, and avoided sika deer. Variation in preference indices derived from these scat analyses compared to indices derived from kill data appear to be due to adjustments in biomass intake when sex-age of a killed individual is known: a component missing from scat data. Pseudoreplication (multiple samples collected from a single kill site) also skewed results derived from scat analyses. Scat analysis still appears useful in providing insight into the diets of carnivores when the full spectrum of prey species needs to be identified, or when sample sizes from kill data are not sufficient. When sample sizes of kill data are large (as is now possible with GPS-collared animals), kill data adjusted by sex-age categories probably provides the most accurate estimates of prey biomass composition. Our results provide further confirmation of the centrality of medium ungulates, in particular wild boar, to Amur tiger diet, and suggest that the protection of this group of species is critical to Amur tiger conservation. © 2015 International Society of Zoological Sciences, Institute of Zoology/Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd.
Weights of gaur (Bos gaurus) and banteng (Bos javanicus) killed by tigers in Thailand
DOI:10.1002/ece3.6268
PMID:32551089
[本文引用: 1]
The primary prey of tigers across much of South-East Asia has been depleted, reducing the ability of already limited habitat to support tigers. To better understand the extent to which two of the largest prey species, gaur () and banteng (), contribute to the tiger's diet, we estimated the average size of these species killed by tigers. This information is needed to more accurately calculate biomass of these species in the tiger's diet and to devise strategies to increase tiger carrying capacity where habitat is fragmented and limited in west-central Thailand. We used temporally clumped locations of 24 satellite radio-collared tigers to identify their kill sites and obtained mandibles from 82 gaur and 79 banteng. Kills were aged by teeth eruption sequence, sectioning the M1 molar and counting cementum annuli. Of all gaur killed, 45.2% were adults; of all banteng killed, 55.7% were adults. The average weight of banteng killed was 423.9 kg, which was similar to the 397.9 kg average weight for gaur. The mean weight of both prey species is 3.5-4.5 times greater than the predicted 1:1 preferred prey to predator ratio. In the absence of medium-sized prey, killing these larger animals may be especially critical for female tigers provisioning nearly independent young when male offspring are already larger than the mother. This is the first study to present data on the average weights of gaur and banteng killed in South-East Asia, and these results suggest that these are key prey species to target in tiger prey recovery efforts.© 2020 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Genomic evidence for inbreeding depression and purging of deleterious genetic variation in Indian tigers
Habitat preferences of tigers Panthera tigris in the Sundarbans East Wildlife Sanctuary, Bangladesh, and management recommendations
DOI:10.1017/S0030605307012094 URL [本文引用: 1]
Big cats kill more livestock when wild prey reaches a minimum threshold
DOI:10.1016/j.biocon.2015.09.031 URL [本文引用: 2]
Diet composition and prey preference of tiger, leopard, and dhole in Kalakkad-Mundanthurai Tiger Reserve, Southern Western Ghats, India
Rapid recovery of tigers Panthera tigris in Parsa Wildlife Reserve, Nepal
DOI:10.1017/S0030605317000886 URL [本文引用: 2]
Risk of population extinction from fixation of new deleterious mutations
DOI:10.1111/j.1558-5646.1994.tb02188.x
PMID:28568413
[本文引用: 1]
The fixation of new deleterious mutations is analyzed for a randomly mating population of constant size with no environmental or demographic stochasticity. Mildly deleterious mutations are far more important in causing loss of fitness and eventual extinction than are lethal and semilethal mutations in populations with effective sizes, N, larger than a few individuals. If all mildly deleterious mutations have the same selection coefficient, s against heterozygotes and 2s against homozygotes, the mean time to extinction, t¯, is asymptotically proportional to e4Nes/Ne for 4N s > 1. Nearly neutral mutations pose the greatest risk of extinction for stable populations, because the magnitude of selection coefficient that minimizes t¯ is about ŝ = 0.4/N. The influence of variance in selection coefficients among mutations is analyzed assuming a gamma distribution of s, with mean s¯ and variance σs2. The mean time to extinction increases with variance in selection coefficients if s¯ is near ŝ, but can decrease greatly if s¯ is much larger than ŝ. For a given coefficient of variation of s, c=σs/s¯, the mean time to extinction is asymptotically proportional to Ne1+1/c2 for 4Nes¯>1. When s is exponentially distributed, (c = 1) t¯ is asymptotically proportional to Ne2. These results in conjunction with data on the rate and magnitude of mildly deleterious mutations in Drosophila melanogaster indicate that even moderately large populations, with effective sizes on the order of N = 10, may incur a substantial risk of extinction from the fixation of new mutations.© 1994 The Society for the Study of Evolution.
Camera-trap surveys reveal high diversity of mammals and pheasants in Medog, Tibet
DOI:10.1017/S0030605319001467 URL [本文引用: 1]
Transboundary frontiers: An emerging priority for biodiversity conservation
DOI:S0169-5347(20)30079-3
PMID:32668213
[本文引用: 2]
The world's biomes and their associated ecosystems are artificially fractured by geopolitical boundaries that define countries. Yet 'transboundary' landscapes often overlap with biodiversity hotspots, contain surprisingly important ecosystems, and provide critical habitats for threatened species. Notwithstanding, biodiversity in these landscapes is increasingly imperiled by infrastructure, including walls and fences along borders and cross-border roads that drive landscape fragmentation and the loss of ecological connectivity. Associated problems due to reduced governance (e.g., illegal wildlife trade) also undermine conservation efforts in these important regions. In this review, we distinguish positive and negative effects of transboundary frontiers on biodiversity conservation, discuss lessons from existing frameworks, and identify scenarios that can maximize opportunities for biodiversity conservation in transboundary frontiers.Copyright © 2020 Elsevier Ltd. All rights reserved.
Genome-wide evolutionary analysis of natural history and adaptation in the world’s tigers
DOI:10.1016/j.cub.2018.09.019
[本文引用: 4]
No other species attracts more international resources, public attention, and protracted controversies over its intraspecific taxonomy than the tiger (Panthera tigris) [1, 2]. Today, fewer than 4,000 free-ranging tigers survive, covering only 7% of their historical range, and debates persist over whether they comprise six, five, or two subspecies [3, 4, 5, 6]. The lack of consensus over the number of tiger subspecies has partially hindered the global effort to recover the species from the brink of extinction, as both captive breeding and landscape intervention of wild populations increasingly require an explicit delineation of the conservation management units [7]. The recent coalescence to a late Pleistocene bottleneck (circa 110 kya) [5, 8, 9] poses challenges for detecting tiger subspecific morphological traits, suggesting that elucidating intraspecific evolution in the tiger requires analyses at the genomic scale. Here, we present whole-genome sequencing analyses from 32 voucher specimens that resolve six statistically robust monophyletic clades corresponding to extant subspecies, including the recently recognized Malayan tiger (P. tigris jacksoni). The intersubspecies gene flow is very low, corroborating the recognized phylogeographic units. We identified multiple genomic regions that are candidates for identifying the adaptive divergence of subspecies. The body-size-related gene ADH7 appears to have been strongly selected in the Sumatran tiger, perhaps in association with adaptation to the tropical Sunda Islands. The identified genomic signatures provide a solid basis for recognizing appropriate conservation management units in the tiger and can benefit global conservation strategic planning for this charismatic megafauna icon.
Phylogeography and genetic ancestry of tigers (Panthera tigris)
Tigers of the world: Genomics and conservation
DOI:10.1146/annurev-animal-020518-115106 URL [本文引用: 3]
Sumatran tiger survival threatened by deforestation despite increasing densities in parks
DOI:10.1038/s41467-017-01656-4
PMID:29208916
[本文引用: 2]
The continuing development of improved capture-recapture (CR) modeling techniques used to study apex predators has also limited robust temporal and cross-site analyses due to different methods employed. We develop an approach to standardize older non-spatial CR and newer spatial CR density estimates and examine trends for critically endangered Sumatran tigers (Panthera tigris sumatrae) using a meta-regression of 17 existing densities and new estimates from our own fieldwork. We find that tiger densities were 47% higher in primary versus degraded forests and, unexpectedly, increased 4.9% per yr from 1996 to 2014, likely indicating a recovery from earlier poaching. However, while tiger numbers may have temporarily risen, the total potential island-wide population declined by 16.6% from 2000 to 2012 due to forest loss and degradation and subpopulations are significantly more fragmented. Thus, despite increasing densities in smaller parks, we conclude that there are only two robust populations left with >30 breeding females, indicating Sumatran tigers still face a high risk of extinction unless deforestation can be controlled.
African Swine Fever threatens Southeast Asia’s 11 endemic wild pig species
Securing a future for wild Indochinese tigers: Transforming tiger vacuums into tiger source sites
DOI:10.1111/j.1749-4877.2010.00220.x
PMID:21392350
[本文引用: 1]
A century ago, tigers (Panthera tigris Linnaeus, 1758) were so common in parts of Southeast Asia as to be considered pests, and governments sponsored their killing. Habitat loss and fragmentation, market-driven poaching and loss of prey have since led to the disappearance of Indochinese tigers from most their former range. Despite 15 years of dedicated tiger conservation funding, national estimates of Indochinese tiger subpopulations can at best only be roughly approximated. The future for the subspecies appears grim unless very focused efforts can be applied to stabilize and recover subpopulations. On a regional scale, the 2 proposed subspecies Panthera tigris corbetti and P. tigris jacksoni are effectively managed as separate conservation units. Evaluating where to place conservation efforts should consider the vulnerability (likelihood of extinction) and irreplaceability (likelihood that an area contributes uniquely to regional conservation) of tiger subpopulations. Only 1 site in Thailand supporting <200 individuals (Huai Kha Khaeng-Thung Yai) is considered low vulnerability, and is irreplaceable. Five sites in Lao, Thailand and Peninsular Malaysia are medium vulnerability and irreplaceable. Priorities at these 6 sites are to double tiger numbers within 10 years through protection and monitoring. Seven sites in Lao, Thailand and Myanmar are high vulnerability and irreplaceable, and might be recovered if government commitment to tigers, staff capacity and legal frameworks for tiger protection are established. Tigers are extremely vulnerable or even extinct in Cambodia's Eastern Plains and the site is irreplaceable for tigers because it represents the only large (>10,000 km(2) ) block of dry forest habitat available in the region. A reintroduction program is the only option to recover tigers there.© 2010 ISZS, Blackwell Publishing and IOZ/CAS.
Developing a national tiger action plan for the union of Myanmar
A century ago, tigers were considered pests in Myanmar. Hunters claimed thousands, yet populations persisted. In the past century, because of habitat loss and prey depletion, coupled with the recent demand for traditional medicines, tiger populations have been reduced to a few hundred individuals. As a first step toward long-term planning for tigers, and to guide efforts to increase protected area coverage, the Myanmar government in 1998 initiated a project to develop a revised National Tiger Action Plan. Extensive surveys confirmed tigers in only 4 of 17 survey locations. Significant populations are thought to persist in the far North and far South of the country, where large, intact forests offer the potential for tiger recovery. With partnerships and collaborations, tiger populations can be protected in the short term (<5 years) by expanding protected areas and corridors, mobilizing enforcement staff to reduce poaching of tigers and prey, and amending existing wildlife legislation in accordance with international laws. Over the long term (5-20 years), recovery of Myanmar's tiger populations will depend on increasing support from local people, zoning tiger areas to reduce habitat loss and disturbance, and maintaining connectivity of existing national and transboundary forests. This article reviews the development of a new National Tiger Action Plan for the Union of Myanmar and discusses a blueprint for conservation measures aimed at saving tigers from extinction.
Mutation accumulation and the extinction of small populations
DOI:10.1086/285812 URL [本文引用: 1]
Prey preferences of the snow leopard (Panthera uncia): Regional diet specificity holds global significance for conservation
Detection of SARS-CoV-2 in a free ranging leopard (Panthera pardus fusca) in India
DOI:10.1007/s10344-022-01608-4 URL [本文引用: 1]
Historical and current distribution ranges and loss of mega-herbivores and carnivores of Asia
Long-term monitoring of a Bengal tiger (Panthera tigris tigris) population in a human-dominated landscape of Central India
DOI:10.1007/s10344-016-1070-5 URL [本文引用: 1]
Canis simensis
Diversity of susceptible hosts in canine distemper virus infection: A systematic review and data synthesis
DOI:10.1186/s12917-016-0702-z
PMID:27170307
[本文引用: 1]
Background: Canine distemper virus (CDV) is the etiological agent of one of the most infectious diseases of domestic dogs, also known as a highly prevalent viral infectious disease of carnivores and posing a conservation threat to endangered species around the world. To get a better panorama of CDV infection in different Orders, a retrospective and documental systematic review of the role of CDV in different non-dog hosts was conducted. The bibliographical data were collected from MedLine/PubMed and Scopus databases. Data related to Order, Family, Genus and Species of the infected animals, the presence or absence of clinical signs, mortality, serological, molecular or antigenic confirmation of CDV infection, geographic location, were collected and summarized. Results: Two hundred seventeen scientific articles were considered eligible which includes reports of serological evaluation, and antigenic or genomic confirmation of CDV infection in non-dog hosts. CDV infects naturally and experimentally different members of the Orders Carnivora (in 12 Families), Rodentia (four Families), Primates (two Families), Artiodactyla (three Families) and Proboscidea (one Family). The Order Carnivora (excluding domestic dogs) accounts for the vast majority (87.5 %) of the records. Clinical disease associated with CDV infection was reported in 51.8 % of the records and serological evidence of CDV infection in apparently healthy animals was found in 49.5 % of the records. High mortality rate was showed in some of the recorded infections in Orders different to Carnivora. In non-dog hosts, CDV has been reported all continents with the exception of Australasia and in 43 different countries. Conclusions: The results of this systematic review demonstrate that CDV is able to infect a very wide range of host species from many different Orders and emphasizes the potential threat of infection for endangered wild species as well as raising concerns about potential zoonotic threats following the cessation of large-scale measles vaccination campaigns in the human population.
Serologic survey of selected pathogens in free-ranging Bengal tigers (Panthera tigris tigris) in Nepal
DOI:10.7589/JWD-D-20-00046
PMID:33822151
[本文引用: 1]
Serum samples of 11 Bengal tigers (Panthera tigris tigris) from Chitwan National Park in Nepal, collected between 2011-17, were evaluated for the presence of antibodies to eight diseases commonly investigated in large felids. This initial serologic survey was done to establish baseline information to understand the exposure of Nepal's free-ranging tiger population to these diseases. Tiger serum samples collected opportunistically during encounters such as translocation, human conflict, and injury were placed in cold storage for later use. Frozen serum samples were assessed for feline coronavirus (FCoV), feline immunodeficiency virus, feline leukemia virus, feline herpesvirus (FHV), canine distemper virus, canine parvovirus-2 (CPV-2), leptospirosis (LEP; seven serovars), and toxoplasmosis (TOX). Six tigers were found to be positive for LEP, eight for CPV-2, five for FHV, one for FCoV, and 10 for TOX. Tigers, like other wild felids, have been exposed to these common pathogens, but further research is needed to determine the significance of these pathogens to the Nepali population.© Wildlife Disease Association 2021.
Amur tiger (Panthera tigris altaica) energetic requirements: Implications for conserving wild tigers
DOI:10.1016/j.biocon.2013.12.012 URL [本文引用: 2]
Population dynamics of Amur tigers (Panthera tigris altaica) in Sikhote-Alin Biosphere Zapovednik: 1966-2012
DOI:10.1111/1749-4877.12141
PMID:26037543
[本文引用: 5]
In 2010, the world's tiger (Panthera tigris) range countries agreed to the goal of doubling tiger numbers over 12 years, but whether such an increase is biologically feasible has not been assessed. Long-term monitoring of tigers in Sikhote-Alin Biosphere Zapovednik (SABZ), Russia provided an opportunity to determine growth rates of a recovering population. A 41-year growth phase was followed by a rapid decline in tiger numbers. Annual growth rates during the growth phase averaged 4.6%, beginning near 10% in the earliest years but quickly dropping below 5%. Sex ratio (females per male) mirrored growth rates, declining as population size increased. The rapid decline from 2009 to 2012 appeared to be tied to multiple factors, including poaching, severe winters and disease. Reproductive indicators of this population are similar to those of Bengal tiger populations, suggesting that growth rates may be similar. These results suggest that, first, tiger populations likely in general grow slowly: 3-5% yearly increases are realistic and larger growth rates are likely only when populations are highly depressed, mortality rates are low and prey populations are high relative to numbers of adult females. Second, while more research is needed, it should not be assumed that tiger populations with high prey densities will necessarily grow more quickly than populations with low prey densities. Third, while growth is slow, decline can be rapid. Fourth, because declines can happen so quickly, there is a constant need to monitor populations and be ready to respond with appropriate and timely conservation interventions if tiger populations are to remain secure. Finally, an average annual growth rate across all tiger populations of 6%, required to reach the Global Tiger Initiative's goal of doubling tiger numbers in 12 years, is a noble but unlikely scenario. © 2015 International Society of Zoological Sciences, Institute of Zoology/Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd.
Assessing the threat of human consumption of tiger prey in the Bangladesh Sundarbans
DOI:10.1111/j.1469-1795.2012.00571.x URL [本文引用: 1]
First structured camera-trap surveys in Karen State, Myanmar, reveal high diversity of globally threatened mammals
DOI:10.1017/S0030605316001113 URL [本文引用: 1]
Combined effects of climate change and sea-level rise project dramatic habitat loss of the globally endangered Bengal tiger in the Bangladesh Sundarbans
DOI:10.1016/j.scitotenv.2019.01.383 URL [本文引用: 1]
Exposure of wild Sumatran tiger (Panthera tigris sumatrae) to canine distemper virus
DOI:10.7589/JWD-D-20-00144
PMID:33822169
[本文引用: 1]
Canine distemper virus (CDV) is recognized as a conservation threat to Amur tigers (Panthera tigris altaica) in Russia, but the risk to other subspecies remains unknown. We detected CDV neutralizing antibodies in nine of 21 wild-caught Sumatran tigers (42.9%), including one sampled on the day of capture, confirming exposure in the wild.© Wildlife Disease Association 2021.
Population density estimates and conservation concern for clouded leopards Neofelis nebulosa, marbled cats Pardofelis marmorata and tigers Panthera tigris in Htamanthi Wildlife Sanctuary, Sagaing, Myanmar
DOI:10.1017/S0030605317001260
[本文引用: 1]
The clouded leopard Neofelis nebulosa is a potent ambassador species for conservation, occurring from the Himalayan foothills eastwards to Indochina, between which Myanmar is a biogeographical land bridge. In Myanmar's Northern Forest Complex, the species co-occurs with the tiger Panthera tigris, leopard Panthera pardus, marbled cat Pardofelis marmorata, golden cat Catopuma temminckii and leopard cat Prionailurus bengalensis. We deployed cameras within the Htamanthi Wildlife Sanctuary over 2 consecutive years. In 2014-2015 we deployed 82 camera stations around the Nam Pa Gon stream (Catchment 1) for 7,365 trap days. In 2015-2016 we deployed 80 camera stations around the Nam E Zu stream (Catchment 2) for 7,192 trap days. In Catchment 1 we identified five tigers from 26 detections, five clouded leopards from 41 detections (68 photographs) and 11 marbled cats from 13 detections. Using Bayesian-based spatial capture-recapture we estimated the densities of tigers and clouded leopards to be 0.81 +/- SD 0.40 and 0.60 +/- SD 0.24 individuals per 100 km(2), respectively. In Catchment 2 we identified two tigers from three detections, nine clouded leopards from 55 detections and 12 marbled cats from 37 detections. Densities of clouded leopards and marbled cats were 3.05 +/- SD 1.03 and 8.80 +/- SD 2.06 individuals per 100 km(2), respectively. These differences suggest that human activities, in particular gold mining, are affecting felid populations, and these are a paramount concern in Htamanthi. We demonstrate the importance of Htamanthi within the Northern Forest Complex and highlight the Yawbawmee corridor as a candidate for protection.
Loss and vulnerability of lowland forests in mainland Southeast Asia
DOI:10.1111/cobi.13538 URL [本文引用: 2]
Conservation priorities for endangered Indian tigers through a genomic lens
DOI:10.1038/s41598-017-09748-3
PMID:28851952
[本文引用: 1]
Tigers have lost 93% of their historical range worldwide. India plays a vital role in the conservation of tigers since nearly 60% of all wild tigers are currently found here. However, as protected areas are small (< 300 km(2) on average), with only a few individuals in each, many of them may not be independently viable. It is thus important to identify and conserve genetically connected populations, as well as to maintain connectivity within them. We collected samples from wild tigers (Panthera tigris tigris) across India and used genome-wide SNPs to infer genetic connectivity. We genotyped 10,184 SNPs from 38 individuals across 17 protected areas and identified three genetically distinct clusters (corresponding to northwest, southern and central India). The northwest cluster was isolated with low variation and high relatedness. The geographically large central cluster included tigers from central, northeastern and northern India, and had the highest variation. Most genetic diversity (62%) was shared among clusters, while unique variation was highest in the central cluster (8.5%) and lowest in the northwestern one (2%). We did not detect signatures of differential selection or local adaptation. We highlight that the northwest population requires conservation attention to ensure persistence of these tigers.
Impact of cattle grazing on wild ungulate habitat in Kanha-Pench Corridor, Madhya Pradesh
Dispersal of Amur tiger from spatial distribution and genetics within the eastern Changbai Mountain of China
DOI:10.1002/ece3.4832
[本文引用: 1]
Population dispersal and migration often indicate an expanded habitat and reduced inbreeding probability, and to some extend reflects improvement in the condition of the population. The Amur tiger population in the northern region of the Changbai mountain in China mostly distributes along the Sino-Russian border, next to the population in southwest Primorye in Russia. The successful dispersal westward and transboundary movement are crucial for the persistence of the Amur tiger in this area. This study explored the spatial dispersal of the population, transboundary migration, and the genetic condition of the Amur tiger population within the northern Changbai mountain in China, using occurrence data and fecal samples. Our results from 2003 to 2016 showed that the Amur tiger population in this area was spreading westward at a speed of 12.83 +/- 4.41 km every three years. Genetic diversity of the Amur tiger populations in southwest Primorye was slightly different than the population in our study area, and the potential individual migration rate between these two populations was shown to be about 13.04%. Furthermore, the relationships between genetic distances and spatial distances indicated the existence of serious limitations to the dispersal of the Amur tiger in China. This study provided important information about spatial dispersal, transboundary migration, and the genetic diversity of Amur tigers in China, showed the urgent need for Amur tiger habitat restoration, and suggested some important conservation measures, such as corridor construction to eliminate dispersal barriers and joint international conservation to promote trans-boundary movement.
Inbreeding status and implications for Amur tigers
DOI:10.1111/acv.12761 URL [本文引用: 2]
Assessment and management options of human-tiger conflicts in Kerinci Seblat National Park, Sumatra, Indonesia
DOI:10.3106/041.034.0303 URL [本文引用: 1]
Isolation and genetic characterization of canine parvovirus in a Malayan tiger
DOI:10.3389/fvets.2021.660046 URL [本文引用: 1]
Characterizing human-tiger conflict in Sumatra, Indonesia: Implications for conservation
Identifying conservation successes, failures and future opportunities; assessing recovery potential of wild ungulates and tigers in Eastern Cambodia
Impacts of hydropower on the habitat of jaguars and tigers
DOI:10.1038/s42003-021-02878-5
PMID:34887507
[本文引用: 1]
The rapid expansion of hydropower across tropical landscapes has caused extensive habitat loss and degradation, triggering biodiversity loss. Despite known risks to freshwater biodiversity, the flooding of terrestrial habitats caused by dam construction, and associated impacts on terrestrial biota, have been rarely considered. To help fill this knowledge gap, we quantified the habitat loss following inundation of hydropower reservoirs across the range of two iconic species, jaguars and tigers. To do so, we compiled existing and planned dams intersecting the distribution of these apex predators. We found 164 dams intersecting the jaguar range, in total flooding 25,397 km. For tigers, we identified 421 dams, amounting to 13,750 km. As hydropower infrastructure is projected to expand in the decades ahead, these values are expected to increase greatly, particularly within the distribution of jaguars where the number of dams will nearly quadruple (429 planned dams). Despite the relatively few dams (41) planned across the range of tigers, most will intersect priority conservation areas for this species. We recommend a more cautious pursuit of hydropower in topographically flat regions, to avoid extensive habitat flooding which has occurred in the Neotropics, and avoiding dam construction in priority conservation landscapes for tigers.© 2021. The Author(s).
Human-carnivore conflict in China: A review of current approaches with recommendations for improved management
DOI:10.1111/j.1749-4877.2012.00303.x
PMID:22691204
[本文引用: 1]
Human-wildlife conflict (HWC) is a conservation concern that increasingly threatens the continued existence of some of the world's most endangered species. With an increase in human population, urban sprawl and subsequent encroachment on wild land, human and wildlife interaction has become inevitable. In the majority of cases, this interaction results in a negative outcome for humans, wildlife or both. In China, these key elements, along with a decrease in wild prey species, have resulted in the expansion of HWC encounters, and the need for alleviating this conflict has become a conservation priority. Loss of human life, livestock and/or crops is most often the catalysts that fuel HWC. Techniques to alleviate conflict around the world have included preventative measures and mitigation techniques, such as financial compensation and other incentive programs. Both types of measures have had variable success. We review the current status of human-carnivore conflict management in China, and, drawing lessons from around the globe, we make recommendations for improving conservation management in China. For example, an increase in law enforcement in nature reserves is vital to reducing human disturbance in prime carnivore habitat, thereby reducing conflict encounters. Also, modifications to current wildlife compensation programs, so that they are linked with preventative measures, will ensure that moral hazards are avoided. Furthermore, investigating the potential for a community self-financed insurance scheme to fund compensation and increasing efforts to restore wild prey populations will improve the outcome for wildlife conservation. Ultimately, HWC management in China will greatly benefit from an integrative approach.© 2012 ISZS, Blackwell Publishing and IOZ/CAS.
Cerebral cysticercosis in a wild Bengal tiger (Panthera tigris tigris) in Bhutan: A first report in non-domestic felids
DOI:10.1016/j.ijppaw.2021.02.003 URL [本文引用: 1]
Tiger density, movements, and immigration outside of a tiger source site in Thailand
Thailand Tiger Action Plan 2010-2022
Experimental canine parvovirus infection in dogs
In specific pathogen-free dogs, clinical signs of experimental canine parvovirus infection were mild, inconsistent and transient. Clinical signs were more pronounced in conventionally-raised dogs, but the severe disease reported in field cases was not reproduced in either group. A pronounced plasma viremia occurred on the 2nd to 4th day post-infection (d.p.i.) in dogs challenged oronasally. Antibody was detectable on the 5th d.p.i. Marked pyrexia was rare, but a significant temperature rise usually coincided with the appearance of antibody and the cessation of viremia. Significant lymphopenia, but not leukopenia, occurred on the 3rd to 7th d.p.i. Virus could be readily isolated from fecal matter on the 3rd to 8th d.p.i.; a few dogs continued to shed virus for up to 12 days. In dogs challenged parenterally, the onset of elevated temperatures, viral shed and antibody production occurred 24-48 hours sooner. Convalescent dogs were no longer contagious for susceptible contact animals 25 days or longer after challenge, although infectious virus persisted in feces for more than 6 months at room temperature. Active giardiasis seemed to exacerbate the clinical syndrome, although treatment with corticosteroids or anti-thymocyte serum did not.
Mapping and predicting forest loss in a Sumatran tiger landscape from 2002 to 2050
DOI:S0301-4797(18)31199-X
PMID:30368149
[本文引用: 1]
Riau Province in central Sumatra, with its peatland, lowland, and montane forest habitats, was once a stronghold for Sumatran tiger (Panthera tigris sumatrae) populations. Today, Riau may have one of the highest deforestation rates in the world and wildlife populations are dwindling, with natural forest now comprising approximately only 18% of the province, mostly contained within protected areas. Agriculture (acacia, rubber, and oil palm) makes up the majority of Riau's land cover and deforestation for the creation of new plantations is rampant. Natural forest and tigers still remain in Bukit Tigapuluh National Park and Rimbang Baling Wildlife Reserve, which remain connected to tiger populations in montane forest on the western edge of Sumatra. In this study, using freely available Landsat imagery and a maximum likelihood classification algorithm, we create land cover maps for central Sumatra from 2002 to 2016. We then use current land cover, elevation, and slope variables to predict changes from forest to plantation from 2016 to 2050 at five year intervals using a multilayer perceptron neural network. Finally, we compare connectivity based on a 100 km distance threshold (based on potential tiger dispersal) across the landscape and across years. Land cover maps had 80-90% accuracy, and we predict forest in Tesso Nilo and the western edge of the study area to be lost by 2050 given current rates of deforestation. Our connectivity analysis shows that Tesso Nilo and the area between Rimbang Baling and Bukit Tigapuluh are important components for maintaining connectivity throughout the study area. Focusing conservation and rehabilitation efforts on forests close to plantations in flat areas, including Tesso Nilo, is necessary to maintain forests and increase connectivity in Riau to ensure future habitat connectivity for survival of tigers and Sumatra's other diverse endemic species.Copyright © 2018 Elsevier Ltd. All rights reserved.
Integrated assessments call for establishing a sustainable meta-population of Amur tigers in northeast Asia
DOI:10.1016/j.biocon.2021.109250 URL [本文引用: 3]
Severe conservation risks of roads on apex predators
DOI:10.1038/s41598-022-05294-9
PMID:35190573
[本文引用: 3]
The global expansion of road networks threatens apex predator conservation and ecosystem functioning. This occurs through wildlife-vehicle collisions, habitat loss and fragmentation, reduced genetic connectivity and increased poaching. We reviewed road impacts on 36 apex predator species and assessed their risk from current roads based on road exposure and species vulnerability. Our findings reveal all apex predators are exposed to road impacts. Eight of the ten species with the highest risk occur in Asia, although other high-risk species are present in the Americas, Africa and Europe. The sloth bear suffers the highest risk of all apex predators, followed by the tiger and dhole. Based on species risk from roads, we propose a widely applicable method to assess the potential impact of future roads on apex predators. We applied this method to proposed road developments in three areas: the Brazilian Amazon, Africa, and Nepal, to locate high-impact road segments. Roughly 500 protected areas will be intersected by these roads, threatening core apex predator habitats. We advocate the need for rigorous road development planning to apply effective mitigation measures as an urgent priority and to avoid construction in wilderness areas and predator strongholds.© 2022. The Author(s).
Documenting the demise of tiger and leopard, and the status of other carnivores and prey, in Lao PDR’s most prized protected area: Nam Et-Phou Louey
Wildlife-tourism, local communities and tiger conservation: A village-level study in Corbett Tiger Reserve, India
DOI:10.1016/j.forpol.2015.04.007 URL [本文引用: 1]
Trophic cascades from wolves to grizzly bears in Yellowstone
DOI:10.1111/1365-2656.12123 URL [本文引用: 1]
Conserving the world’s megafauna and biodiversity: The fierce ugency of now
Status and ecological effects of the world’s largest carnivores
Examining the shifting patterns of poaching from a long-term law enforcement intervention in Sumatra
DOI:10.1016/j.biocon.2016.10.029 URL [本文引用: 1]
Mortality of Amur tigers: The more things change, the more they stay the same
DOI:10.1111/1749-4877.12147
PMID:26096683
[本文引用: 2]
Poaching as well as loss of habitat and prey are identified as causes of tiger population declines. Although some studies have examined habitat requirements and prey availability, few studies have quantified cause-specific mortality of tigers. We used cumulative incidence functions (CIFs) to quantify cause-specific mortality rates of tigers, expanding and refining earlier studies to assess the potential impact of a newly emerging disease. To quantify changes in tiger mortality over time, we re-examined data first collected by Goodrich et al. (; study period 1: 1992-2004) as well as new telemetry data collected since January 2005 (study period 2: 2005-2012) using a total of 57 tigers (27 males and 30 females) monitored for an average of 747 days (range 26-4718 days). Across the entire study period (1992 to 2012) we found an estimated average annual survival rate of 0.75 for all tigers combined. Poaching was the primary cause of mortality during both study periods, followed by suspected poaching, distemper and natural/unknown causes. Since 2005, poaching mortality has remained relatively constant and, if combined with suspected poaching, may account for a loss of 17-19% of the population each year. Canine distemper virus (CDV) may be an additive form of mortality to the population, currently accounting for an additional 5%. Despite this relatively new source of mortality, poaching remains the main threat to Amur tiger survival and, therefore, population growth. © 2015 International Society of Zoological Sciences, Institute of Zoology/Chinese Academy of Sciences and Wiley Publishing Asia Pty Ltd.
A canine distemper virus epidemic in Serengeti lions
(Panthera leo).DOI:10.1038/379441a0 URL [本文引用: 1]
Restoration of the Amur tiger (Panthera tigris altaica) population in the northwest of its distribution area
DOI:10.1134/S1062359021080239 URL [本文引用: 1]
High frequency of an otherwise rare phenotype in a small and isolated tiger population
Setting Priorities for the Conservation and Recovery of Wild Tigers: 2005-2015
Learning from the past to prepare for the future: Felids face continued threat from declining prey
DOI:10.1111/ecog.03303 URL [本文引用: 1]
Movement and home range characteristics of reintroduced tiger (Panthera tigris) population in Panna Tiger Reserve, central India
DOI:10.1007/s10344-016-1026-9 URL [本文引用: 1]
Ecological preferences of large carnivores in remote, high-altitude protected areas: Insights from Buxa Tiger Reserve, India
DOI:10.1017/S0030605317000060 URL [本文引用: 1]
Saving wild tigers: A case study in biodiversity loss and challenges to be met for recovery beyond 2010
DOI:10.1111/j.1749-4877.2010.00214.x
PMID:21392347
[本文引用: 1]
Wild tigers are being annihilated. Tiger range countries and their partners met at the 1st Asian Ministerial Conference on Tiger Conservation in January 2010 to mandate the creation of the Global Tiger Recovery Program to double the number of tigers by 2022. Only 3200-3600 wild adult tigers remain, approximately half of the population estimated a decade ago. Tigers now live in only 13 countries, all of which are experiencing severe environmental challenges and degradation from the effects of human population growth, brisk economic expansion, rapid urbanization, massive infrastructure development and climate change. The overarching challenge of tiger conservation, and the conservation of biodiversity generally, is that there is insufficient demand for the survival of wild tigers living in natural landscapes. This allows the criminal activities of poaching wild tigers and their prey and trafficking in tiger derivatives to flourish and tiger landscapes to be diminished. The Global Tiger Recovery Program will support scaling up of practices already proven effective in one or more tiger range countries that need wider policy support, usually resources, and new transnational actions that enhance the effectiveness of individual country actions. The program is built on robust National Tiger Recovery Priorities that are grouped into themes: (i) strengthening policies that protect tigers; (ii) protecting tiger conservation landscapes; (iii) scientific management and monitoring; (iv) engaging communities; (v) cooperative management of international tiger landscapes; (vi) eliminating transnational illegal wildlife trade; (vii) persuading people to stop consuming tiger; (viii) enhancing professional capacity of policy-makers and practitioners; and (ix) developing sustainable, long-term financing mechanisms for tiger and biodiversity conservation.© 2010 ISZS, Blackwell Publishing and IOZ/CAS.
Canine distemper virus: An emerging disease in wild endangered Amur tigers (Panthera tigris altaica)
Tiger poaching and trafficking in India: Estimating rates of occurrence and detection over four decades
DOI:10.1016/j.biocon.2014.08.016 URL [本文引用: 1]
Spatial genetic analysis reveals high connectivity of tiger (Panthera tigris) populations in the Satpura-Maikal landscape of Central India
DOI:10.1002/ece3.432
PMID:23403813
[本文引用: 1]
We investigated the spatial genetic structure of the tiger meta-population in the Satpura-Maikal landscape of central India using population- and individual-based genetic clustering methods on multilocus genotypic data from 273 individuals. The Satpura-Maikal landscape is classified as a global-priority Tiger Conservation Landscape (TCL) due to its potential for providing sufficient habitat that will allow the long-term persistence of tigers. We found that the tiger meta-population in the Satpura-Maikal landscape has high genetic variation and very low genetic subdivision. Individual-based Bayesian clustering algorithms reveal two highly admixed genetic populations. We attribute this to forest connectivity and high gene flow in this landscape. However, deforestation, road widening, and mining may sever this connectivity, impede gene exchange, and further exacerbate the genetic division of tigers in central India.
Expansion of industrial plantations continues to threaten Malayan tiger habitat
DOI:10.3390/rs9070747 URL [本文引用: 1]
Exploring the motivations associated with the poaching and trafficking of Amur tigers in the Russian Far East
Genetic structure of the Amur tiger (Panthera tigris altaica) population: Are tigers in Sikhote-Alin and Southwest Primorye truly isolated?
DOI:10.1111/1749-4877.12175
PMID:26663570
[本文引用: 4]
We used molecular genetic analyses to noninvasively identify individual Amur tigers and define subpopulations of tigers in the Russian Far East. We identified 63 individuals after genotyping 256 feces, 7 hair and 11 blood samples collected within southern, central and northern Sikhote-Alin, as well as Southwest Primorye. Analysis of nuclear DNA at 9 microsatellite loci demonstrated greater genetic similarity between animals from southern and northern Sikhote-Alin (some 500 km apart) than between animals from Ussuriskii State Nature Reserve and Southwest Primorye (less than 10 km apart at their nearest point), suggesting that a true barrier exists preventing movements of tigers between Southwest Primorye and the southern Sikhote-Alin Mountains. © 2015 International Society of Zoological Sciences, Institute of Zoology/Chinese Academy of Sciences and John Wiley & Sons Australia, Ltd.
Population size estimation of Amur tigers in Russian Far East using noninvasive genetic samples
DOI:10.1644/10-MAMM-A-368.1 URL [本文引用: 1]
Tigers need cover: Multi-scale occupancy study of the big cat in Sumatran forest and plantation landscapes
The Social Organization of Tigers (Panthera tigris) in Royal Chitawan National Park, Nepal
Physiological stress response of African elephants to wildlife tourism in Madikwe Game Reserve, South Africa
DOI:10.1071/WR19045 URL [本文引用: 1]
Behavioural changes in African elephants in response to wildlife tourism
DOI:10.1111/jzo.12661 URL [本文引用: 1]
The spatial distribution and population density of tigers in mountainous terrain of Bhutan
DOI:10.1016/j.biocon.2019.07.037 URL [本文引用: 3]
Panthera tigris jacksoni population crash and impending extinction due to environmental perturbation and human-wildlife conflict
DOI:10.3390/ani11041032 URL [本文引用: 3]
The contribution of wildlife tourism to tiger conservation (Panthera tigris tigris)
DOI:10.1080/14888386.2017.1410443 URL [本文引用: 1]
Maintaining tiger connectivity and minimizing extinction into the next century: Insights from landscape genetics and spatially-explicit simulations
DOI:10.1016/j.biocon.2017.12.022 URL [本文引用: 2]
Spatiotemporal pattern and major causes of the Amur tiger population dynamics
DOI:10.3724/SP.J.1003.2009.08207
[本文引用: 1]
Based on a comprehensive literature review, we analyzed the spatiotemporal pattern of the Amur tiger (<em>Panthera tigris altaica</em>) population dynamics during the past century, and proposed a set of strategies and measures for conserving this endangered species from the perspectives of landscape ecology and sus-tainability science. The Amur tiger is a keystone species in the region of Russia Far East, Eastern Mongolia, Northeastern China, and North Korea, and its population declined dramatically during the past century, from the historical record of 3,000 to the current low level of about 500 because of different kinds of anthropo-genic disturbances. The extant tiger population is distributed mainly in the Russia Far East region, including one large habitat area along the Sikhote Mountain and two smaller habitat patches near the Russia-China border. A small number of tiger individuals are also found in several small isolated habitat patches in north-eastern China. The primary causes for the decline of the tiger population were poaching, habitat loss, and habitat fragmentation. The scarcity of prey and wars were also responsible for the decrease in the tiger popu-lation. To better conserve this endangered species, we propose the following strategies and measures: to es-tablish a long-term monitoring platform; to strictly prohibit tiger poaching and restrict forest logging, hunt-ing, and building roads and other artificial structures within the tiger distribution areas; and to build animal movement corridors among reserves and across the China-Russia border. To achieve these goals, large-scale land use planning and habitat pattern optimization are needed, and conservation goals must be integrated with the overall goal of sustainable development in the region that simultaneously considers environmental, eco-nomic, and social factors based on the principles of landscape ecology and sustainability science.
东北虎种群的时空动态及其原因分析
DOI:10.3724/SP.J.1003.2009.08207
[本文引用: 1]
作者通过对广泛的历史资料的综合分析, 研究了东北虎(Panthera tigris altaica)种群在近一个多世纪以来的时空动态及其原因, 并从景观生态学和可持续性科学的角度对东北虎的保护提出了建议。东北虎是俄罗斯远东、中国东北、蒙古东部以及朝鲜的关键种。近百年以来的多种人为干扰因素使该种群从一个世纪前的近3,000只的历史最高纪录减少到目前约500只的低水平。现存种群主要分布在俄罗斯远东地区Sikhote山脉的一个大生境区域和两个靠近中俄边境孤立的小生境斑块, 少数个体零星分布在中国境内几个小而孤立的生境斑块中。威胁东北虎种群生存的两个最主要因素是捕猎、生境丧失和破碎化。此外, 食物匮乏、战争等因素也对东北虎的生存和繁衍有一定程度的影响。我们建议建立长期监测平台; 禁止盗猎并限制在东北虎潜在栖息地采伐、狩猎以及修筑道路等人类活动; 建立相互连通的保护区域, 特别是建立中国与俄罗斯间跨国界的生态廊道。这些保护措施应该以景观生态学和可持续性科学为指导, 实现空间资源的合理配置和土地利用格局的优化, 同时考虑东北虎种群的生存和当地经济、社会发展的需求, 从而促进实现区域可持续发展的最终目标。
Physiological stress responses of tigers due to anthropogenic disturbance especially tourism in two central Indian tiger reserves
Bringing the tiger back from the brink-the six percent solution
Living on the edge: Opportunities for Amur tiger recovery in China
DOI:10.1016/j.biocon.2017.11.008 URL [本文引用: 2]
Long-distance dispersal of an Amur tiger indicates potential to restore the North-east China/Russian Tiger Landscape
DOI:10.1017/S0030605315000794 URL [本文引用: 1]
Amur tigers and leopards returning to China: Direct evidence and a landscape conservation plan
DOI:10.1007/s10980-015-0278-1 URL [本文引用: 6]
An introduction to long-term tiger-leopard observation network based on camera traps in Northeast China
DOI:10.17520/biods.2020139 URL [本文引用: 1]
东北虎豹生物多样性红外相机监测平台概述
Investigation on the population of wild Bengal tiger (Panthera tigris tigris) in Medog, Tibet
西藏墨脱县孟加拉虎种群数量调查
Research advances and perspectives of conservation genetics of threatened mammals in China
DOI:10.16829/j.slxb.150517
[本文引用: 1]
China is one of the world's mega-diverse countries. There are diverse terrestrial and marine mammals in China, including endemic and flagship species like the giant panda, snub-nosed monkey, South China tiger, Milu, and Baiji. However, habitat loss and fragmentation, poaching, pollution, and climate change imposed direct threats to mammals' survival, which resulted in the genetic diversity loss of species. Genetic diversity is the basic component of biodiversity and affects the long-term survival of species and populations. Therefore, conservation genetics was established as an important branch subject of conservation biology, aiming to investigate the population genetic variation and genetic mechanisms of species endangerment and extinction. With rapid development in research techniques and methods, great progress has been made in China in the past 40 years in the evaluation of genetic diversity and inbreeding, landscape genetics, ecological genetics, and genetic management on threatened mammals. Meanwhile, the threats to mammals' survival caused by human activities still exist. The further development of new techniques, such as high-throughput sequencing, could deepen our understanding of the genetic adaptation and endangerment processes of threatened mammals, and lead to more effective management and conservation.
中国濒危兽类保护遗传学研究进展与展望
DOI:10.16829/j.slxb.150517
[本文引用: 1]
我国是全球生物多样性大国,拥有包括大熊猫、金丝猴、华南虎、麋鹿、白鱀豚等特有物种和旗舰物种在内的丰富兽类资源。近几十年来,土地利用模式转变、盗猎、环境污染、气候变化等因素使许多兽类物种面临生存威胁,导致物种遗传多样性丧失。而遗传多样性是生物多样性的基本组成部分,决定了物种和种群能否长期生存。保护遗传学作为保护生物学的一大分支学科,旨在通过遗传学分析探明种群遗传变异和物种濒危的遗传学机制。近40年来,随着研究手段和技术的不断发展,我国兽类保护遗传学在遗传多样性和近交水平评估、景观遗传学、生态遗传学和圈养种群遗传管理等方面都取得了重要成果。然而,未来人类社会发展可能为濒危兽类带来的威胁依然存在,高通量测序等新技术的进一步发展则能够帮助我们更加深入地了解濒危物种和种群遗传适应与濒危机制,从而实现对濒危兽类的有效管理与保护。
A landscape-based conservation strategy to double the wild tiger population
DOI:10.1111/j.1755-263X.2010.00162.x URL [本文引用: 3]
Navjot’s nightmare revisited: Logging, agriculture, and biodiversity in Southeast Asia
DOI:10.1016/j.tree.2013.04.005
PMID:23764258
[本文引用: 1]
In 2004, Navjot Sodhi and colleagues warned that logging and agricultural conversion of Southeast Asia's forests were leading to a biodiversity disaster. We evaluate this prediction against subsequent research and conclude that most of the fauna of the region can persist in logged forests. Conversely, conversion of primary or logged forests to plantation crops, such as oil palm, causes tremendous biodiversity loss. This loss is exacerbated by increased fire frequency. Therefore, we conclude that preventing agricultural conversion of logged forests is essential to conserving the biodiversity of this region. Our analysis also suggests that, because Southeast Asian forests are tightly tied to global commodity markets, conservation payments commensurate with combined returns from logging and subsequent agricultural production may be required to secure long-term forest protection. Copyright © 2013 Elsevier Ltd. All rights reserved.
The many effects of carnivores on their prey and their implications for trophic cascades, and ecosystem structure and function
DOI:10.1016/j.fooweb.2016.09.002 URL [本文引用: 1]
Prey depletion as a threat to the world’s large carnivores
DOI:10.1098/rsos.160252 URL [本文引用: 2]
Range contractions of the world’s large carnivores
DOI:10.1098/rsos.170052 URL [本文引用: 1]
Lycaon pictus (amended version of 2012 assessment)
Canine distemper and wildlife
DOI:10.16829/j.slxb.150456
[本文引用: 1]
Canine distemper is a zoonotic infectious disease caused by canine distemper virus, outbreaks of this disease have occurred multiple times among domestic dogs and wildlife worldwide. Cross-species transmission of canine distemper has been threatening various wildlife populations severely such as Siberian tiger, African lion, snow leopard and giant panda. At the same time, the range of infectious hosts is still expanding. Recent studies showed that wildlife in China, especially populations in the wild, are facing serious threats from infection of canine distemper due to the constant variation of canine distemper virus and increase of stray dog population. In order to better cope with the hazard brought by canine distemper to wildlife, this study summarized the research progress on pathogenic characteristics, epidemiology, pathogenic mechanism, diagnosis and treatment of canine distemper in wildlife. Based on the above research, we proposed prevention and control measures aimed at canine distemper in wildlife from three aspects including source of infection, transmission route and susceptible animals. At present, due to the lack of scientific research and technology in nature reserves in China and the low awareness toward the risk of wildlife canine distemper transmission, little has been done in monitoring wildlife canine distemper, which undoubtedly increases the difficulty to its prevention and control. Therefore, in order to secure the existence of wildlife population in China, the research about wildlife canine distemper monitoring and epidemiology should be enhanced, establishing an effective monitoring and control system to protect wildlife population from canine distemper.
犬瘟热与野生动物
DOI:10.16829/j.slxb.150456
[本文引用: 1]
犬瘟热是一种犬瘟热病毒引起的多种动物共患传染病,在世界范围内的家犬和野生动物中多次爆发。犬瘟热的跨种间传播对多种野生动物如东北虎、非洲狮、雪豹、大熊猫等野生动物的种群安全造成严重威胁,并且感染的宿主范围仍在扩大。近年来的研究表明,犬瘟热病毒的不断变异和野生动物栖息地内流浪犬只的增多使我国野生动物尤其是野外种群正在面临犬瘟热的严重威胁。为了更好地应对犬瘟热对野生动物的危害,本文综述了犬瘟热的病原特征和在野生动物中的流行病学、致病机制、诊断与治疗以及疫苗免疫方面的研究进展,在此基础上提出了针对传染源、传播途径、易感动物三方面的控制措施来减少野生动物犬瘟热的传播风险。目前,由于国内自然保护区科研技术力量欠缺以及防控意识薄弱,对于野生动物犬瘟热的监测处于空白状态,这无疑加大了野生动物犬瘟热的防控难度。因此,为了保障我国野生动物种群安全,我们需要加强在野生动物犬瘟热监测与流行病学方面的研究工作,建立有效的监测防控体系将犬瘟热阻挡在野生动物种群之外。
Estimating abundance and density of Amur tigers along the Sino-Russian border
DOI:10.1111/1749-4877.12210
PMID:27136188
[本文引用: 1]
As an apex predator the Amur tiger (Panthera tigris altaica) could play a pivotal role in maintaining the integrity of forest ecosystems in Northeast Asia. Due to habitat loss and harvest over the past century, tigers rapidly declined in China and are now restricted to the Russian Far East and bordering habitat in nearby China. To facilitate restoration of the tiger in its historical range, reliable estimates of population size are essential to assess effectiveness of conservation interventions. Here we used camera trap data collected in Hunchun National Nature Reserve from April to June 2013 and 2014 to estimate tiger density and abundance using both maximum likelihood and Bayesian spatially explicit capture-recapture (SECR) methods. A minimum of 8 individuals were detected in both sample periods and the documentation of marking behavior and reproduction suggests the presence of a resident population. Using Bayesian SECR modeling within the 11 400 km(2) state space, density estimates were 0.33 and 0.40 individuals/100 km(2) in 2013 and 2014, respectively, corresponding to an estimated abundance of 38 and 45 animals for this transboundary Sino-Russian population. In a maximum likelihood framework, we estimated densities of 0.30 and 0.24 individuals/100 km(2) corresponding to abundances of 34 and 27, in 2013 and 2014, respectively. These density estimates are comparable to other published estimates for resident Amur tiger populations in the Russian Far East. This study reveals promising signs of tiger recovery in Northeast China, and demonstrates the importance of connectivity between the Russian and Chinese populations for recovering tigers in Northeast China. © 2016 International Society of Zoological Sciences, Institute of Zoology/Chinese Academy of Sciences and John Wiley & Sons Australia, Ltd.
Human expansion into Asian highlands in the 21st Century and its effects
DOI:10.1038/s41467-022-32648-8
PMID:36002452
[本文引用: 1]
Most intensive human activities occur in lowlands. However, sporadic reports indicate that human activities are expanding in some Asian highlands. Here we investigate the expansions of human activities in highlands and their effects over Asia from 2000 to 2020 by combining earth observation data and socioeconomic data. We find that ∼23% of human activity expansions occur in Asian highlands and ∼76% of these expansions in highlands comes from ecological lands, reaching 95% in Southeast Asia. The expansions of human activities in highlands intensify habitat fragmentation and result in large ecological costs in low and lower-middle income countries, and they also support Asian developments. We estimate that cultivated land net growth in the Asian highlands contributed approximately 54% in preventing the net loss of the total cultivated land. Moreover, the growth of highland artificial surfaces may provide living and working spaces for ∼40 million people. Our findings suggest that highland developments hold dual effects and provide new insight for regional sustainable developments.© 2022. The Author(s).
Does prey density limit Amur tiger Panthera tigris altaica recovery in northeastern China?
DOI:10.2981/12-090 URL [本文引用: 2]
Tiger predation on livestock in Gedang, Medog, Southeast Tibet
Once the distribution of tigers ( <EM>Panthera tigris</EM>) extended throughout the broad-leaved forests of southeastern Tibet. These tigers are now extinct except for one remnant population in Medog County. From May to June 2000, a survey was conducted in Gedang Village ( Xiang in Chinese), the only place in the county where losses of livestock to tigers are high. It was found that the amount of livestock killed was high in 1994 and 1995, but after that it dropped greatly, probably in part because one persistent livestock killer was shot in 1996. Between the early 1993 and late1990s, the xiang's cattle population dropped by 11 %. However, horses and mules increased by 23 % because more animals were bought elsewhere for use in transport. In 1999, the xiang lost 7.8 % of its cattle and 1.9 % of its horses to tiger predation. 4-5 tigers including a female with large cub was reported by local officials in the area that year. A total of 21 households were interviewed. Of these a household lost on average 0.8 cattle and 0.2 horses during the previous 12 months ; nine households had no losses. One reason that tigers are tempted by livestock is lack of sufficient wild prey. Widespread illegal hunting has greatly reduced tiger's principal prey population such as wild pig, takin, red goral, and mutjacs. Until recovery of wildlife populations, an effort must be made to reduce tiger predation on livestock. Conservation recommendations were as follows: (1) Villagers should herd and guard their animals cooperatively instead of permitting livestock to wander untended. (2) Overgrown abandoned fields, slopes covered with tall bracken fern, and thickets near villages should be cleared and converted to open pastures, with some of themfenced, to remove cover in which tigers can hide, and (3) Some animals could also be housed in stalls, especially in winter when much of the predation occurs.<BR>
西藏墨脱格当乡野生虎捕食家畜现状与保护建议
曾经在西藏东南的阔叶林中广泛分布的孟加拉虎, 目前仅有一个小种群残留在墨脱县境内。2000 年5~6 月间, 在大型家畜遭受野生虎捕食最严重的墨脱县格当乡展开调查,试图寻找减轻虎害的办法。结果表明: 1994~1995 年虎的捕食率达到最高, 对牛和骡马的捕食率分别是17.9 %和9.4 %; 但1996 年后, 捕食率分别降低到7.8 %和1.8 %, 这可能与1996 年当地曾捕杀过一头虎有关。1993~1999 年间, 全乡牛的数量下降了11 %, 但骡马数量上升了23 %, 这是因为当地为增加运输能力而从别处购得骡马。据反映, 目前格当乡境内大约有4~5 头虎。1997 年和1999 年, 均见母虎和小虎同行, 说明该种群尚有繁殖。在抽样的21 户居民家中, 1999 年4 月到2000 年5 月间, 66.6 %的人家有大型牲畜遭虎捕杀, 共损失牛27 头, 马12 匹。而自1993 年以来, 这21 户中共有18 户(85.8 %) 有牲畜被杀记录, 共计损失117 头。其中对牛的捕食率达19.7 %, 对马达11.9 %, 平均每户损失牛112头, 骡马0.5 匹。非法狩猎减少了虎的猎物如野猪、羚牛等的数量, 是老虎转向家畜的主要原因之一。虎害已对格当乡群众的经济造成较大负面影响。但格当乡以及周边地区保护着中国最后的野生孟加拉虎种群, 为确保虎的长期生存同时减少人—虎冲突, 建议改变目前放牧方式, 尽可能联合放牧, 增加看护; 改善放牧地条件, 清除牧场周围蕨草丛; 减少对羚牛等有蹄类的猎杀, 以减少对老虎猎物种群的破坏; 对部分家畜移入棚内试行圈养,既保护家畜, 又提高乳制品产量和增加农家肥料; 实行多种经营方法, 建议养一些山羊和家禽; 政府应该帮助安置好部分愿意外迁的居民, 这样既满足这些居民的需要, 同时也减轻对当地野生动物种群的压力。
From Amur tiger occurrence in the Greater Khingan Mountains to doing an overall conservation for its in China

{kind=link}
{kind=link}
{kind=link}
{kind=link}