



由于自然植被、景观特征以及其他众多的环境因子都具有空间嵌套性, 动物的生境选择也因空间尺度不同而存在差异(Kotliar & Wiens, 1990; Orians & Wittenberger, 1991)。Johnson (1980)提出, 动物的生境选择遵循4个基本层次, 分别为区域尺度、景观尺度、家域尺度和斑块尺度。只从单一空间尺度上研究动物的生境选择, 其结果可能无法全面准确地反映物种生境选择模式在空间尺度上的差异(Kotliar & Wiens, 1990)。因此, 从多个空间尺度研究动物在不同时期的生境选择, 可以获得更加详细而可靠的生境选择信息, 有助于全面揭示物种生境选择的空间动态和季节性变化。
绿尾虹雉(Lophophorus lhuysii)隶属鸡形目雉科虹雉属, 是中国特有大型高山雉类, 国家一级重点保护野生动物, 世界自然保护联盟(IUCN)受胁物种红色名录中的易危(VU)物种, 《濒危野生动植物种国际贸易公约》(CITES)附录I物种和《中国脊椎动物红色名录》中的濒危(EN)物种(蒋志刚等, 2016), 主要栖息于海拔3,000‒4,900 m的高山、亚高山灌丛和草甸生境。绿尾虹雉分布于四川、甘肃南部、青海南部及云南西北部(郑光美, 2015), 是我国大陆地区5个值得优先保护的中国特有鸟类物种之一(Chen, 2013)。对于绿尾虹雉的生境选择和利用, 已有学者分别在四川的夹金山(卢汰春, 1985; 何芬奇等, 1986; 卢汰春等, 1986)、小寨子沟(余翔等, 2017)、片口(钟雪等, 2020b)以及甘肃的白水江(张涛, 1995)等区域进行过研究(隆廷伦等, 1998; Wang et al, 2017), 但是, 以往研究很少考虑生境选择的层次性, 也没有对其生境选择的季节性差异进行比较。为了明确绿尾虹雉在不同空间尺度和不同季节的生境资源需求, 本研究从景观与微生境两个空间尺度、繁殖期与非繁殖期两个时间周期, 对四川卧龙国家级自然保护区内绿尾虹雉的生境选择进行了研究, 以期为该物种的生境保护和管理提供科学依据。
1 材料和方法
1.1 研究地区
四川卧龙国家级自然保护区始建于1963年, 总面积2,000 km2, 是我国最早建立的国家级自然保护区之一。该保护区位于四川省阿坝藏族、羌族自治州汶川县的西南部, 地处邛崃山脉东南坡, 主要保护对象为大熊猫(Ailuropoda melanoleuca)等珍稀野生动物及森林生态系统。保护区内生物资源极为丰富, 除大熊猫外还分布有雪豹(Panthera uncia)、红喉雉鹑(Tetraophasis obscurus)、绿尾虹雉等国家一级重点保护野生动物, 以及藏酋猴(Macaca thibetana)、白马鸡(Crossoptilon crossoptilon)和勺鸡(Pucrasia macrolopha)等国家二级重点保护野生动物。该地区位于青藏高原气候区东缘, 年均气温9.8℃, 7月最高, 平均气温17℃, 1月最低, 平均气温‒1.7℃, 年均降水量1,800 mm, 年均蒸发量874 mm。保护区地形复杂, 最低海拔1,150 m, 最高6,250 m, 落差超过5,000 m。随着海拔梯度的上升, 保护区内的生境类型由常绿阔叶林过渡到常绿落叶阔叶混交林, 再到针阔混交林, 然后是亚高山针叶林, 最后为高山灌丛、高山草甸和高山流石滩(周小平等, 2019)。
1.2 野外样线调查
根据卧龙自然保护区内的植被类型、地形条件和人为影响等因素, 结合以往研究报道的绿尾虹雉生境特征, 于2019年10月以及2020年1月、4月、7月和10月在卧龙羊角湾、魏家沟和文扎都3个地方共布设15条样线。样线海拔范围为3,300‒4,150 m, 尽可能穿越多种植被类型, 并尽量多地覆盖绿尾虹雉的潜在分布区域, 样线平均长度为3.6 km (2.5‒ 4.2 km), 单侧宽度50 m, 样线间最小间距300 m (图1)。调查过程中, 调查队员沿样线慢速行走, 沿途对绿尾虹雉的实体、鸣叫、羽毛及粪便进行搜寻和记录。绿尾虹雉的粪便长度在4‒6 cm之间, 前端粗大为圆球状, 且覆盖着白色的尿酸层, 末端逐渐变细收尾, 整体呈现幅度较小的旋转和扭曲。其粪便尺寸通常大于同域分布的红喉雉鹑和雪鹑(Lerwa lerwa), 而形状上又与白马鸡有所区别, 因此容易进行区分。此外, 当发现疑似但不易鉴定的粪便时不予记录, 以提高研究的准确性。
图1
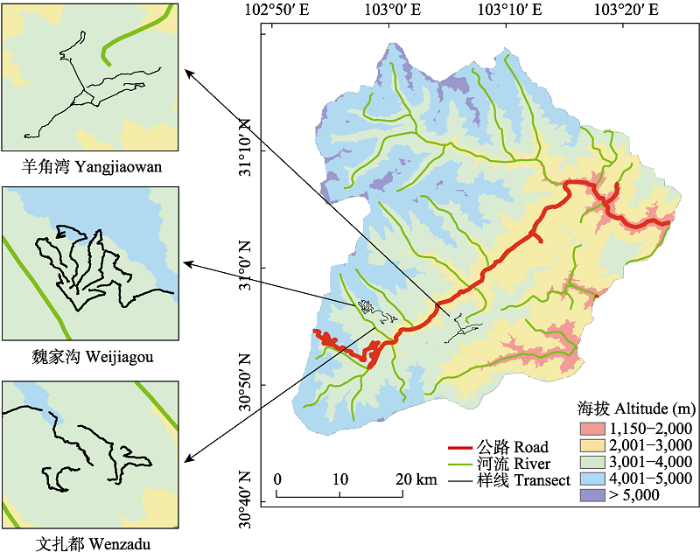
图1
四川卧龙国家级自然保护区内绿尾虹雉野外调查样线分布图
Fig. 1
Distribution map of transects for surveying Chinese monal in Wolong National Nature Reserve, Sichuan
在样线行进过程中, 一旦检测到绿尾虹雉的实体、鸣叫、羽毛及粪便, 则将所在地点视为绿尾虹雉利用的微生境, 以手持GPS (Garmin, GPSMAP 631csx)测定物种出现地点的经纬度, 并建立10 m × 10 m的样方以测定一系列反映微生境在食物资源、隐蔽条件、热量维持等方面的变量(Bibby et al, 1998; Sutherland et al, 2004)。首先, 在样方中心使用手持GPS测量海拔高度; 然后在10 m × 10 m的大样方内, 使用罗盘(DQL-5)测量坡度和坡向, 使用皮尺测量并计算乔木平均胸径, 目测样方内的乔木盖度、乔木平均高度和岩石盖度; 将样方均分为4个5 m × 5 m的中样方, 分别用皮尺测量并计算中样方内的灌木平均高度, 目测灌木盖度和雪盖度, 对样方内的牦牛粪便数量进行计数, 将4个中样方内的变量值取平均或求和后作为大样方的微生境变量; 在4个中样方的中心各设置1个1 m × 1 m的小样方, 使用皮尺测量草本层平均高度, 目测草本盖度、苔藓盖度、落叶盖度, 将4个小样方内的变量值取平均后作为大样方的微生境变量(Wang et al, 2021)。同时, 按照系统取样法, 沿样线每行走300 m设置1个对照样方, 对照样方的设置和变量记录与利用样方一致(钟雪等, 2020a)。当样方内覆盖有积雪时, 所有草本层和地表层的变量(包括苔藓、落叶、岩石和牦牛粪便数量)的测量均只考虑未被雪覆盖的部分, 不计算雪盖之下的部分。
1.3 红外相机监测
使用ArcGIS 10.2在卧龙国家级自然保护区范围内生成公里网格(1 km × 1 km)。自2017年10月起, 在海拔3,500‒4,800 m的高山区域共选择72个公里网格作为监测网格, 每一监测周期在网格中建立1‒2个红外相机监测位点, 共建立了176个有效监测的红外相机位点。在同一网格内, 同一周期的监测位点间距不小于300 m, 而不同周期的监测位点间距不小于100 m (李晟等, 2016; 施小刚等, 2017)。红外相机(易安卫士L710或L710F)通常安放于有动物活动痕迹(如粪便、足迹、毛发等)的四周, 距离痕迹处3‒5 m, 固定于乔木、灌木枝干或岩石上距离地面50‒100 cm处, 清空相机前的杂草和灌木以减少误触。红外相机设置为连续拍摄3张照片, 触发间隔设为30 s, 灵敏度中, 24 h全天工作。记录红外相机布设位点的经纬度、海拔高度、生境类型、坡向、坡度和坡位等生境信息。每隔4‒6个月更换一次电池和内存卡。
1.4 数据处理与分析
1.4.1 景观尺度
将野外样线调查中绿尾虹雉样点与红外相机拍摄所获绿尾虹雉样点相结合, 共获得繁殖期绿尾虹雉位点113个, 非繁殖期位点114个(各变量平均值见附录1)。为避免对相同区域重复取样和模型的过度拟合, 通过最小活动区面积的方法对绿尾虹雉位点进行筛选。在ArcGIS 10.2中, 以繁殖期和非繁殖期绿尾虹雉位点分别生成560 m半径的缓冲区, 当若干出现点的缓冲区相互重叠时, 随机保留其中的1个, 其余的位点剔除(Wang et al, 2017; Xu et al, 2020)。最终繁殖期共保留67个位点, 非繁殖期共保留64个位点(图2)。同时, 在研究区域随机生成100个对照样点, 相邻两个样点之间距离 > 560 m。
图2
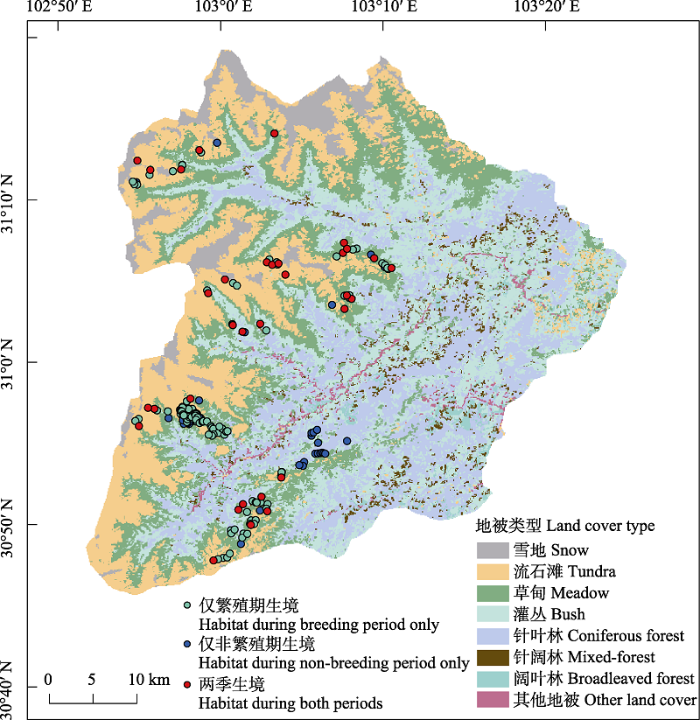
图2
四川卧龙国家级自然保护区内调查发现的不同时期绿尾虹雉生境
Fig. 2
Chinese monal habitat during breeding and non-breeding period detected in Wolong National Nature Reserve, Sichuan
植被数据来源于全国森林二类调查成果(
主成分分析(principal components analysis, PCA)能够将高维的原始数据投影到低维的相互正交的主成分上, 在实现数据降维的同时可最大化地保留原始数据的信息量, 并能通过相互正交的主成分有效克服变量间的相关性或多重共线性问题(Wang et al, 2018)。因此, 首先对所有变量进行主成分分析, 提取特征根(eigenvalue) > 1的主成分, 保留最大方差法标准旋转后的因子得分进入后续分析。使用逻辑斯蒂回归模型分析绿尾虹雉景观尺度的生境选择及其影响因素。在进行逻辑斯蒂回归分析时, 纳入各主成分作为单级因子(不考虑变量间二级以上的变量交互作用), 建立不同组合的备选模型(包括零模型)。使用AICc值来评价所有可能的模型结果, AICc值越小则模型准确性越高。若在所有模型结果中某单一模型ΔAICc > 2则视其为最优模型, 使用该模型计算参数估计值; 否则, 表明多个模型对于变量有相近的解释效能, 对所有ΔAICc ≤ 2的备选模型采用多模型平均的方法求出参数估计值及其标准误(Burnham & Anderson, 2002)。
1.4.2 微生境尺度
在野外样线调查过程中, 共获得繁殖期(3‒8月)绿尾虹雉位点52个, 对照样方101个; 非繁殖期(9月至翌年2月)绿尾虹雉位点69个, 对照样方81个(各变量平均值见附录2)。对所有变量进行主成分分析, 提取特征根 > 1的主成分, 保留最大方差法标准旋转后的因子得分, 作为降维后的微生境变量进入后续分析。用二项式逻辑斯蒂回归模型分别对繁殖期和非繁殖期的绿尾虹雉利用样方和对照样方的差异进行比较, 分析其微生境选择偏好和影响利用的重要变量。在进行逻辑斯蒂回归分析时, 纳入各主成分作为单级因子(不考虑变量间二级以上的变量交互作用), 建立不同组合的备选模型(包括零模型)。使用AICc值来评价模型结果, 并进行模型平均。逻辑斯蒂回归模型分析在R 4.0.5 (R Core Team, 2021)中进行, 使用了glmulti (Calcagno, 2020)、MuMIn (Bartoń, 2020)两个插件。统计分析中, 视P < 0.05为统计显著。
2 结果
2.1 景观尺度
2.1.1 景观变量主成分分析
表1 繁殖期和非繁殖期绿尾虹雉景观尺度生境因子主成分分析载荷
Table 1
| 景观变量 Landscape variable | 繁殖期 Breeding period | 非繁殖期 Non-breeding period | ||||||
|---|---|---|---|---|---|---|---|---|
| 主成分1 PC1 | 主成分2 PC2 | 主成分3 PC3 | 主成分4 PC4 | 主成分1 PC1 | 主成分2 PC2 | 主成分3 PC3 | 主成分4 PC4 | |
| 海拔 Altitude | 0.492 | 0.624 | ‒0.417 | ‒0.060 | 0.528 | 0.573 | ‒0.438 | ‒0.038 |
| 坡度 Slope | ‒0.223 | ‒0.259 | ‒0.178 | 0.784 | ‒0.275 | ‒0.206 | ‒0.151 | 0.782 |
| 坡向 Aspect | 0.079 | 0.342 | 0.224 | 0.681 | 0.097 | 0.310 | 0.166 | 0.688 |
| 阔叶林比例 Proportion of broad-leaved forest | ‒0.044 | 0.011 | 0.767 | ‒0.102 | ‒0.046 | 0.040 | 0.755 | ‒0.097 |
| 针叶林比例 Proportion of coniferous forest | ‒0.348 | ‒0.763 | ‒0.101 | ‒0.033 | ‒0.393 | ‒0.743 | ‒0.126 | ‒0.058 |
| 针阔混交林比例 Proportion of mixed-forest | ‒0.213 | ‒0.379 | 0.434 | ‒0.123 | ‒0.251 | ‒0.305 | 0.455 | ‒0.152 |
| 灌丛比例 Proportion of bush | ‒0.621 | ‒0.239 | 0.287 | 0.176 | ‒0.618 | ‒0.221 | 0.298 | 0.143 |
| 草甸比例 Proportion of meadow | ‒0.356 | 0.825 | ‒0.256 | ‒0.043 | ‒0.285 | 0.853 | ‒0.262 | 0.023 |
| 流石滩比例 Proportion of tundra | 0.849 | 0.200 | ‒0.139 | ‒0.038 | 0.839 | 0.180 | ‒0.127 | ‒0.042 |
| 雪地比例 Proportion of snow | 0.648 | ‒0.098 | 0.036 | 0.001 | 0.642 | ‒0.140 | 0.028 | ‒0.021 |
| 其他地被比例 Proportion of other land cover | ‒0.073 | ‒0.098 | 0.642 | 0.177 | ‒0.062 | ‒0.087 | 0.658 | 0.195 |
| 特征根 Eigenvalue | 2.124 | 2.095 | 1.624 | 1.174 | 2.166 | 1.949 | 1.643 | 1.183 |
| 贡献率 Contribution (%) | 19.311 | 19.048 | 14.760 | 10.670 | 19.694 | 17.719 | 14.933 | 10.756 |
| 累计贡献率 Cumulative contribution (%) | 19.311 | 38.360 | 53.120 | 63.790 | 19.694 | 37.413 | 52.346 | 63.102 |
2.1.2 景观尺度生境选择
表2 繁殖期和非繁殖期绿尾虹雉景观尺度生境选择逻辑斯蒂回归模型参数(加粗表示P < 0.05)
Table 2
| 主成分 Principal component | 参数估计值 Parameter estimates | 标准误 Standard error | 显著性 P | |
|---|---|---|---|---|
| 繁殖期 Breeding period | 截距 Intercept | ‒0.836 | 0.244 | <0.001 |
| PC1 | 0.718 | 0.192 | <0.001 | |
| PC2 | 1.412 | 0.257 | <0.001 | |
| PC3 | ‒0.807 | 0.384 | 0.035 | |
| PC4 | ‒0.860 | 0.245 | <0.001 | |
| 非繁殖期 Non-breeding period | 截距 Intercept | ‒0.779 | 0.225 | <0.001 |
| PC1 | 0.730 | 0.188 | <0.001 | |
| PC2 | 1.115 | 0.226 | <0.001 | |
| PC3 | ‒0.744 | 0.348 | 0.033 | |
| PC4 | ‒0.701 | 0.227 | 0.002 |
2.2 微生境尺度
2.2.1 微生境变量主成分分析
主成分分析结果显示: 繁殖期的15个主成分中, 前6个主成分的特征根 > 1, 累计贡献率达84.01%。因此, 我们提取了前6个主成分并计算出其相应的特征向量。主成分1载荷较大的变量为灌木盖度和灌木平均高度; 主成分2为乔木盖度、乔木平均高度和乔木平均胸径; 主成分3为雪盖度和牦牛粪便数量; 主成分4为坡度; 主成分5为岩石盖度; 主成分6为坡向。在非繁殖期则有所不同, 前5个主成分的特征根 > 1, 累计贡献率达72.46%。主成分1载荷较大的为乔木盖度、平均高度和平均胸径; 主成分2为灌木盖度、灌木平均高度、草本盖度和落叶盖度; 主成分3为岩石盖度和牦牛粪便数量; 主成分4为苔藓盖度、主成分5为坡向和雪盖度(表3)。
表3 繁殖期和非繁殖期绿尾虹雉微生境尺度生境因子主成分分析载荷
Table 3
| 微生境变量 Microhabitat factors | 繁殖期 Breeding period | 非繁殖期 Non-breeding period | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 主成分1 PC1 | 主成分2 PC2 | 主成分3 PC3 | 主成分4 PC4 | 主成分5 PC5 | 主成分6 PC6 | 主成分1 PC1 | 主成分2 PC2 | 主成分3 PC3 | 主成分4 PC4 | 主成分5 PC5 | |
| 海拔 Altitude | ‒0.495 | ‒0.269 | ‒0.271 | ‒0.401 | 0.442 | 0.054 | ‒0.603 | ‒0.472 | 0.032 | 0.414 | 0.180 |
| 坡度 Slope | ‒0.062 | 0.002 | ‒0.026 | 0.836 | 0.195 | 0.272 | ‒0.023 | ‒0.281 | 0.389 | ‒0.502 | ‒0.041 |
| 坡向 Aspect | 0.197 | 0.028 | 0.026 | 0.001 | ‒0.072 | 0.855 | ‒0.050 | 0.038 | 0.107 | ‒0.175 | 0.866 |
| 雪盖度 Snow coverage | ‒0.277 | ‒0.032 | ‒0.801 | ‒0.401 | ‒0.232 | 0.003 | ‒0.018 | ‒0.149 | ‒0.112 | 0.365 | 0.607 |
| 乔木盖度 Tree coverage | 0.082 | 0.952 | ‒0.049 | 0.045 | ‒0.014 | 0.048 | 0.928 | 0.084 | 0.065 | 0.022 | ‒0.029 |
| 乔木平均高度 Average height of trees | 0.197 | 0.964 | ‒0.060 | 0.022 | ‒0.035 | ‒0.013 | 0.956 | 0.129 | 0.076 | ‒0.024 | ‒0.031 |
| 乔木平均胸径 Average diameter at breast height of tree | 0.228 | 0.948 | ‒0.058 | 0.003 | ‒0.023 | ‒0.019 | 0.889 | 0.163 | 0.051 | ‒0.056 | 0.017 |
| 灌木盖度 Shrub coverage | 0.801 | ‒0.026 | ‒0.159 | 0.198 | ‒0.046 | 0.305 | ‒0.014 | 0.897 | 0.133 | ‒0.142 | 0.016 |
| 灌木平均高度 Average height of shrub | 0.869 | 0.145 | ‒0.078 | 0.177 | ‒0.170 | 0.122 | 0.165 | 0.866 | ‒0.025 | 0.056 | ‒0.078 |
| 草本盖度 Herbage coverage | ‒0.506 | ‒0.182 | 0.674 | 0.162 | ‒0.390 | ‒0.106 | ‒0.321 | ‒0.667 | ‒0.558 | ‒0.088 | ‒0.024 |
| 草本平均高度 Average height of herbage | 0.152 | 0.014 | 0.094 | 0.732 | ‒0.216 | ‒0.355 | 0.104 | 0.099 | 0.615 | ‒0.313 | ‒0.205 |
| 苔藓盖度 Moss coverage | 0.627 | 0.301 | 0.069 | ‒0.106 | 0.254 | ‒0.274 | ‒0.075 | 0.004 | 0.113 | 0.811 | ‒0.020 |
| 落叶盖度 Litter coverage | 0.768 | 0.395 | ‒0.067 | ‒0.150 | 0.028 | 0.081 | 0.331 | 0.633 | ‒0.154 | 0.212 | ‒0.073 |
| 岩石盖度 Rock coverage | ‒0.020 | ‒0.036 | ‒0.019 | 0.065 | 0.930 | ‒0.076 | ‒0.087 | ‒0.159 | 0.851 | 0.092 | 0.070 |
| 牦牛粪便数量 Count of yak dung | ‒0.300 | ‒0.150 | 0.776 | ‒0.337 | ‒0.138 | 0.069 | ‒0.192 | ‒0.361 | ‒0.585 | ‒0.121 | ‒0.175 |
| 特征根 Eigenvalue | 4.721 | 2.238 | 1.977 | 1.445 | 1.206 | 1.014 | 4.548 | 1.946 | 1.860 | 1.472 | 1.043 |
| 贡献率 Contribution (%) | 31.476 | 14.922 | 13.180 | 9.633 | 8.040 | 6.758 | 30.230 | 12.971 | 12.400 | 9.816 | 6.951 |
| 累计贡献率 Cumulative contribution (%) | 31.476 | 46.398 | 59.578 | 69.211 | 77.251 | 84.009 | 30.323 | 43.293 | 55.693 | 65.509 | 72.461 |
2.2.2 微生境选择分析
繁殖期使用6个主成分建立了64个备选模型, 其中有8个模型的ΔAICc < 2, 因此对这8个模型进行全模型平均。根据平均后的模型参数, 只有主成分5的P值达到统计显著, 表明其是影响绿尾虹雉繁殖期微生境选择的最重要的因子(表4)。结合主成分分析因子载荷矩阵(表3), 在繁殖期内, 绿尾虹雉主要偏好选择岩石盖度较高的微生境。而非繁殖期使用5个主成分建立了32个备选模型, 共有4个模型的ΔAICc < 2。根据平均后的模型参数, 只有主成分2与其微生境选择显著相关(表4)。结合主成分分析的因子载荷(表3), 绿尾虹雉在非繁殖期偏好选择草本盖度较高而灌木盖度、灌木平均高度和落叶盖度均较低的微生境。
表4 繁殖期和非繁殖期绿尾虹雉微生境尺度生境选择逻辑斯蒂回归模型参数(加粗表示P < 0.05)
Table 4
| 主成分 Principal component | 参数估计值 Parameter estimates | 标准误 Standard error | 显著性 P | |
|---|---|---|---|---|
| 繁殖期 Breeding period | 截距 Intercept | ‒0.706 | 0.179 | <0.001 |
| PC4 | 0.246 | 0.210 | 0.243 | |
| PC5 | 0.420 | 0.176 | 0.018 | |
| PC2 | ‒0.133 | 0.217 | 0.541 | |
| PC6 | 0.023 | 0.090 | 0.797 | |
| PC3 | ‒0.023 | 0.095 | 0.808 | |
| 非繁殖期 Non-breeding period | 截距 Intercept | ‒0.155 | 0.170 | 0.365 |
| PC2 | ‒0.380 | 0.175 | 0.031 | |
| PC4 | 0.307 | 0.196 | 0.120 | |
| PC3 | 0.058 | 0.129 | 0.656 | |
| PC1 | ‒0.054 | 0.127 | 0.671 | |
| PC5 | 0.023 | 0.088 | 0.797 |
3 讨论
鸟类对生境的选择是在长期的进化过程中对不断变化环境条件的一种适应行为, 其选择机制和主要影响因子不但受到环境非生物因素和种间、种内相互作用的影响(Bibby et al, 1998; Sutherland et al, 2004), 而且存在着一定程度的空间尺度依赖性(Johnson, 1980)。本研究在以往野外观测描述的基础上(何芬奇等, 1986; 卢汰春等, 1986), 进一步从景观和微生境两个空间尺度, 对绿尾虹雉在繁殖期和非繁殖期的生境选择进行了定量分析和比较, 丰富了该物种的基础生物和生态学资料。研究结果显示, 在不同空间尺度上, 绿尾虹雉生境选择模式具有明显的差异。在景观尺度上, 影响绿尾虹雉生境选择的生态因子要多于微生境尺度, 并呈现出了一定的分层嵌套模式(徐基良等, 2006), 这可能暗示了绿尾虹雉对景观尺度的生境有更高的要求, 而在适宜其生存的景观内, 其微生境的选择和利用则具有较高的灵活性。
在景观尺度下, 绿尾虹雉在繁殖期和非繁殖期都偏好选择平均海拔较高的生境, 表明这一物种已经对高海拔山区的低温低氧环境形成了适应。近期的基因组研究也提供了相应的进化证据(Cui et al, 2019)。绿尾虹雉在不同时期均喜好草甸和流石滩比例较高、森林和灌丛比例较低的生境, 这与以往的野外观察(何芬奇等, 1986; 刘梦瑶等, 2013)和近年的模型预测研究(余翔等, 2017; 钟雪等, 2020b)一致, 表明该物种高度依赖于林线以上的草甸和流石滩生境。然而, 绿尾虹雉夜间常常需要寻找较为高大的乔木或灌木夜栖, 因此针叶林和灌丛并非对其毫无作用, 本研究也发现绿尾虹雉的景观尺度生境内都有一定比例(0.06‒0.13)的针叶林和灌丛(附录1)。绿尾虹雉的主要食物为草本植物, 其对阳坡生境的偏好可能与研究区域内草甸多位于阳坡有关, 光照充足的草甸生境不仅能为绿尾虹雉不同时期提供足够的食物资源, 满足其食物需求, 还能为绿尾虹雉的繁殖和个体发育提供所必需的阳光照度(陈冬梅等, 2017)。
与景观尺度不同, 绿尾虹雉在微生境尺度上的生境选择存在明显的季节性变化, 并且这些变化主要与该物种不同时期的资源需求和行为模式有关。在繁殖期, 绿尾虹雉只表现出了对岩石覆盖率更高的微生境的显著偏好, 这是因为雄鸟在求偶期具有破晓时站在凸出的岩石上鸣叫吸引雌性的行为习性, 高大的裸岩更利于雄鸟进行求偶展示(何芬奇等, 1986); 另一方面, 可能是因为绿尾虹雉有将巢筑于岩壁缝隙或洞穴的习性(何芬奇等, 1986), 裸岩较多的微生境更符合其巢址选择的偏好; 此外, 还可能跟绿尾虹雉春、夏季有时会在岩洞中夜栖有关(卢汰春等, 1986)。我们在野外调查中也曾在一些崖壁下发现大量的绿尾虹雉的粪便和羽毛, 支持了这种可能性。在非繁殖期, 食物获取相对繁殖期更为困难, 为获取充足的食物资源, 绿尾虹雉更加偏好草本盖度较高的微生境。而灌木植物, 不论是常绿的革叶灌木还是冬季落叶的落叶灌丛, 在这一时期所能提供的食物都极为有限, 因此绿尾虹雉在非繁殖期倾向于回避灌木植物发达的微生境。此外, 绿尾虹雉还偏好落叶盖度较低的微生境, 这是由于绿尾虹雉常使用足和喙挖掘土壤中的植物根和茎为食, 数量稀少的落叶能更好地显露食物和便于挖掘, 提高食物的获取效率。
在繁殖期, 成群牦牛在绿尾虹雉栖息地内自由活动和觅食的现象非常普遍。很多学者在不同地区的野外调查中都报道了这一现象, 并且认为牦牛放牧会导致直接干扰和栖息地丧失, 是绿尾虹雉保护的主要威胁(隆廷伦等, 1998; 余翔等, 2017; 钟雪等, 2021)。然而, 近年有研究认为, 牦牛粪便的存在能够增加无脊椎动物的丰富度和可获得性, 在一定程度上有利于改善食虫和杂食性鸟类的生境适宜性(Liu et al, 2019)。本研究发现, 牦牛粪便数量并不会显著影响绿尾虹雉的微生境选择。而且在野外调查中, 我们曾拍摄到绿尾虹雉在牦牛粪便周围刨食的照片, 也发现牦牛粪便上有绿尾虹雉用爪刨过的痕迹。因此, 我们推测牦牛对于绿尾虹雉而言可能并不仅仅是传统认为的直接干扰和威胁, 建议开展进一步的研究以揭示牦牛放牧对于绿尾虹雉种群和生境的实际影响程度和机制, 从而为绿尾虹雉生境内的放牧活动的规范化管理提供理论依据。
附录 Supplementary Material
附录1 繁殖期和非繁殖期绿尾虹雉景观尺度生境特征比较
Appendix 1 Comparison of Chinese monal landscape habitat during breeding and non-breeding period
附录2 繁殖期和非繁殖期绿尾虹雉微生境特征比较
Appendix 2 Comparison of Chinese monal microhabitat during breeding and non-breeding period
参考文献
Preliminary report on the effects of artificial illumination on reproduction of Lophophorus lhuysii
光照影响绿尾虹雉繁殖效果初报
Conservation priority for endemic birds of mainland China based on a phylogenetic framework
DOI:10.5122/cbirds.2013.0021 URL [本文引用: 1]
The first draft genome of Lophophorus: A step forward for Phasianidae genomic diversity and conservation
DOI:10.1016/j.ygeno.2018.07.016 URL [本文引用: 1]
HBW and Birdlife International Illustrated Checklist of the Birds of the World: Non-Passerines
Modelling invasion for a habitat generalist and a specialist plant species
DOI:10.1111/j.1472-4642.2008.00486.x URL [本文引用: 1]
Study on the breeding ecology of the Chinese monal
绿尾虹雉的繁殖生态研究
Niche differentiation among three middle-sized carnivores in Wolong Nature Reserve
卧龙保护区三种中型食肉动物的生态位差异
Red List of China’s Vertebrates
中国脊椎动物红色名录
The comparison of usage and availability measurements for evaluating resource preference
DOI:10.2307/1937156 URL [本文引用: 2]
Habitat selection studies in avian ecology: A critical review
DOI:10.1093/auk/118.2.557 URL [本文引用: 1]
Multiple scales of patchiness and patch structure: A hierarchical framework for the study of heterogeneity
DOI:10.2307/3545542 URL [本文引用: 2]
Camera-trapping survey on the mammal diversity of the Laohegou Nature Reserve, Sichuan Province
四川省老河沟自然保护区兽类多样性红外相机调查
Research status of conservation biology on Lophophorus lhuysii
绿尾虹雉保护生物学研究现状
Dung-associated arthropods influence foraging ecology and habitat selection in Black-necked Cranes (Grus nigricollis) on the Qinghai-Tibet Plateau
DOI:10.1002/ece3.4904 URL [本文引用: 1]
Observational study on the ecology of Chinese monal in winter
绿尾虹雉冬季生态的跟踪观测研究
Field investigation of Chinese monal in Sichuan Province
四川绿尾虹雉的野外考察
Ecological studies on Chinese monal
绿尾虹雉生态学研究
Study on the habitat selection of alpine musk deer in the Helan Mountains in autumn
贺兰山马麝秋季生境选择
Spatial and temporal scales in habitat selection
DOI:10.1086/285138 URL [本文引用: 1]
R: A Language and Environment for Statistical Computing
https://www.R-project.org. (accessed on 2021-12-21)
Camera-trapping surveys of the mammal and bird diversity in Wolong National Nature Reserve, Sichuan Province
DOI:10.17520/biods.2017193 URL [本文引用: 1]
利用红外相机调查四川卧龙国家级自然保护区鸟兽多样性
DOI:10.17520/biods.2017193
[本文引用: 1]

卧龙国家级自然保护区位于邛崃山脉东南坡, 地处邛崃山自然保护区群的核心地带。2014-2016年, 我们在卧龙保护区内使用红外相机技术开展大中型兽类与鸟类的本底调查。经过10,961个相机工作日的调查, 完成有效调查位点83个, 覆盖58个公里网格。共鉴定出分属6目15科的32种野生兽类和4种家畜, 探测数总计2,095次, 其中国家一、二级重点保护野生动物分别有5、11种, 被IUCN红色名录评估为濒危(EN)、易危(VU)、近危(NT)的动物分别有4、6、4种。野生兽类中, 食肉目物种数最多, 共6科16种, 其次为偶蹄目(4科8种)和啮齿目(2科5种)。偶蹄目是有效拍摄数最多的类群(占总探测数的63.33%), 其次为食肉目(20.01%)和啮齿目(5.73%)。食肉目中相对多度最高的物种是猪獾(Arctonyx collaris, RAI = 12.23), 偶蹄目中最高的物种是毛冠鹿(Elaphodus cephalophus, RAI = 32.21)。记录到雪豹(Panthera uncia)、狼(Canis lupus)、豺(Cuon alpinus) 3种大中型顶级食肉兽类。鸟类探测数655 次, 鉴定出分属5目18科的59种鸟类, 其中国家一、二级重点保护野生动物分别有3、9种, 保护区新记录鸟类4种。鸡形目是探测数最高的类群, 占全部鸟类探测数的56.76%。本次调查了解了区内兽类和鸟类的物种组成、空间分布和相对多度, 提供了区内大中型兽类和鸟类群落的初步本底信息, 为后续的保护管理规划和长期监测提供了数据支持和指导。
Research advances of Galliformes since 1990 and future prospects
DOI:10.1186/s40657-018-0124-7 URL [本文引用: 1]
Niche partitioning among three montane ground-dwelling pheasant species along multiple ecological dimensions
DOI:10.1111/ibi.12842 URL [本文引用: 1]
Predicting suitable habitat of the Chinese monal (Lophophorus lhuysii) using ecological niche modeling in the Qionglai Mountains, China
DOI:10.7717/peerj.3477 URL [本文引用: 2]
Overlap and selection of dust-bathing sites among three sympatric montane galliform species
DOI:10.1642/AUK-18-44.1 URL [本文引用: 1]
Multi-scale analysis on wintering habitat selection of Reeves’s pheasant (Syrmaticus reevesii) in Dongzhai National Nature Reserve, Henan Province, China
DOI:10.1016/S1872-2032(06)60031-0 URL [本文引用: 1]
白冠长尾雉越冬期栖息地选择的多尺度分析
Predicting range shifts of the Chinese monal (Lophophorus lhuysii) under climate change: Implications for long-term conservation
DOI:10.1016/j.gecco.2020.e01018 URL [本文引用: 1]
Landscape-scale habitat selection by Buff-throated Partridges Tetraophasis szechenyii during the breeding season
繁殖期黄喉雉鹑在景观尺度上的生境选择
Population density estimation and habitat suitability assessment of Lophophorus lhuysii during breeding season in Xiaozhaizigou National Nature Reserve, Sichuan Province
四川小寨子沟国家级自然保护区绿尾虹雉种群密度调查与栖息地评价
Distribution and ecology of Chinese monals in Baishuijiang Nature Reserve, Gansu
甘肃白水江自然保护区绿尾虹雉分布与生态的初步观察
Population density estimation and microhabitat selection of Golden Pheasant in Sichuan Baishuihe National Nature Reserve
四川白水河国家级自然保护区红腹锦鸡种群密度及微生境选择
Population survey and habitat quality assessment of Lophophorus lhuysii in the Wolong National Nature Reserve
卧龙国家级自然保护区绿尾虹雉种群分布和生境质量评价
Assessment of Chinese monal habitat in Sichuan Piankou Nature Reserve with MaxEnt model
基于MaxEnt模型的四川片口自然保护区绿尾虹雉栖息地评价
The distribution of Budorcas taxicolor in Wolong National Nature Reserve
卧龙自然保护区羚牛(Budorcas taxicolor)的分布格局

{kind=link}
{kind=link}
{kind=link}
{kind=link}