



幼鸟与成鸟在季节性长途迁徙的时间和行为方面可能存在明显的差异(Péron & Grémillet, 2013)。研究表明, 幼鸟几乎总是比成鸟更晚到达度夏/繁殖地(Hockey et al, 1998; Stewart et al, 2002)。造成这一差异的原因目前有两种假设: 一种假设认为由于身体状况的差异, 幼鸟比成鸟更晚离开越冬地, 并且在中途停歇地停留的时间更长、更频繁。如, Ueta和Higuchi (2002)的研究发现, 由于能量摄取效率的差异, 虎头海雕(Haliaeetus pelagicus)、黑脸琵鹭(Platalea minor)和白枕鹤(Antigone vipio)的幼鸟与成鸟相比, 需要在中途停歇地进行更长时间的觅食, 以此来增加脂肪的储存。更多的研究也证实了这一假设, 认为由于翅膀形态的差异, 幼鸟在中途停歇地的觅食效率和飞行效率均低于成鸟, 所以它们中途停歇的时间更长、停歇更频繁(Bowlin, 2007; Paxton et al, 2008; Mellone et al, 2013)。另一种假设认为成鸟繁殖压力大, 所以它们更早到达繁殖地以增加筑巢和繁殖的成功率。研究表明, 提前到达繁殖地的成鸟更能定居在高质量的领地, 更早地孵出幼鸟, 且幼鸟体重更重、数量更多(Smith & Moore, 2005)。鉴于幼鸟和成鸟迁徙行为的差异, 有必要针对幼鸟的迁徙策略开展研究。
东方白鹳(Ciconia boyciana)是国家I级重点保护野生动物(
气候条件会影响鸟类的迁徙持续时间和迁徙行为(Shamoun-Baranes et al, 2017; Becciu et al, 2019), 其中风向和风速对大型翱翔鸟类的迁徙影响较大(Liechti, 2006; Mellone et al, 2012; Rotics et al, 2016; Vidal-Mateo et al, 2016), 如欧洲白鹳(Ciconia ciconia), 其春/秋迁徙均依赖顺风条件(Shamoun-Baranes et al, 2003)。顺风条件对同种鸟类的不同迁徙路线影响不同(Alerstam, 1979; Nilsson et al, 2014), Fan等(2020)发现顺风对东方白鹳长江越冬群体幼鸟的春/秋迁徙飞行速度有影响, 但对渤海湾越冬群体幼鸟尚未有报道。
本研究基于卫星追踪数据和美国环境预测中心(National Center for Environmental Prediction, NCEP)的气象数据, 以东方白鹳渤海湾越冬群体幼鸟为研究对象, 探究其春/秋迁徙策略之间的差异, 并研究风向和风速与迁徙策略之间的关系。此外, 本研究选择均在黑龙江流域出生的幼鸟, 对长江越冬和渤海湾越冬两个群体之间迁徙策略和利用风的方式方面的差异进行了比较分析。
1 材料与方法
1.1 卫星追踪
2016-2018年间, 课题组在中国黑龙江省洪河国家级自然保护区(133°40° E, 47°47° N)和俄罗斯阿穆尔河流域(131°42° E, 47°52° N)共追踪了102只东方白鹳幼鸟。本文选择14只在渤海湾越冬的幼鸟进行卫星追踪研究, 在鸟体背部安装防水型太阳能供电的背包式GPS/GSM发射器, 详见附录1, 设备重量均小于个体重量的3%, 不会干扰追踪鸟类的正常活动(Millspaugh & Marzluff, 2001)。设备每10-180 min记录1次GPS位点(在野外测试中, 水平精度为9.6 ± 5.6 m), 每8-24 h通过手机基站传输1次数据。追踪期间共获得渤海湾越冬群体22条完整的迁徙路线(秋季迁徙N = 13, 春季迁徙N = 9)。由于os 002个体秋季迁徙数据缺失, os 003、os 004、os 005、os 008、os 013等5只个体春季迁徙数据不完整而本研究未采用。
1.2 定位数据处理及行为划分
对获得的位点数据集进行筛选, 去除无效和定位精度过低的数据。参照Wang等(2018)的方法, 将水鸟迁徙过程划分为“飞行”和“非飞行”两种状态。根据“首次通过时间”划分飞行状态和停歇数据集, 对停歇数据集进行二次验证, 将其中满足下列条件的位点重新划分为“飞行”状态: (1)净位移距离(GPS位置与初始位置之间的最短距离)大于150 km, 且净位移速度(净位移距离/位移时间)大于150 km/d; (2)净位移距离大于300 km, 且净位移速度大于75 km/d; 不符合判定条件的被归类为“非飞行”状态。将两种状态用不同的颜色标记, 使用谷歌地球使迁徙路径可视化, 以确认每个位点的到达和离开时间。如果“非飞行”状态持续时间大于48 h, 则该范围被视为1个停歇地。按照这样的划分标准, 将每只追踪个体的春季迁徙或秋季迁徙过程划分为飞行和停歇状态不断变化的连续过程, 并获得每种状态对应的时间段。
1.3 迁徙参数定义
本文采用了9个迁徙参数来量化迁徙策略: 迁徙开始时间、迁徙结束时间、迁徙距离(km)、迁徙持续时间(d)、迁徙速度(km/d)、停歇位点数、停歇持续时间(d)、迁徙飞行时间(d)和迁徙飞行速度(km/d)①(①李祥煌 (2019) 灰雁(Anser anser)迁徙策略的卫星追踪研究. 硕士学位论文, 中国科学院生态环境研究中心, 北京.)。
迁徙开始时间是离开度夏/繁殖地或越冬地的时间, 即判断为迁徙开始位点所对应的时间; 迁徙结束时间为到达度夏/繁殖地或越冬地的时间, 即判断为迁徙结束位点所对应的时间; 迁徙距离为候鸟春/秋迁徙过程中累积的飞行距离(不包括在中途停歇地活动的距离); 迁徙持续时间指候鸟在春/秋迁徙过程中, 从离开度夏/繁殖地或越冬地到抵达越冬地或度夏/繁殖地所花费的时间, 包括中途停歇的时间; 迁徙速度为迁徙距离除以迁徙持续时间, 表示该个体迁徙过程的平均速度; 停歇位点数是指在候鸟迁徙过程中, 被判断为中途停歇地的个数; 停歇持续时间为追踪个体在中途停歇地停留的时长; 迁徙飞行速度是迁徙距离除以迁徙飞行时间, 而迁徙飞行时间是迁徙持续时间减去停歇持续时间, 表示追踪个体处于飞行状态下的速度。
1.4 气象学数据及计算
为计算东方白鹳迁徙途中顺风和逆风的状态, 本文采用Shamoun-Baranes等(2003)的研究方法, 使用R 3.6包“RNCEP” (Kemp et al, 2012), 从NCEP提取数据, 获取了850 mb压力水平下的风向和风速数据, 该R包可以对气象数据集进行检索并对其进行组织和可视化。该数据基于实时观测数据, 利用高时空分辨率进行建模, 并根据观测数据或模型的相对影响程度, 将数据分为4个质量等级(Kalnay et al, 1996), 本研究使用的850 mb的风分量U和V (分别为沿纬度和经度方向的瞬时风速)是4个质量等级中最可靠的A类变量。我们将该风分量数据插值到东方白鹳运动的时空数据中, 用于分析风向和风速与东方白鹳幼鸟迁徙策略的关系。
东方白鹳昼夜迁徙飞行瞬时速度的研究表明, 东方白鹳主要在白天飞行, 夜晚休息, 并且迁徙飞行速度在14:00 (UTC + 8)达到峰值②(② 樊淑娟 (2020) 基于卫星追踪研究东方白鹳(Ciconia boyciana)的迁徙生态学. 硕士学位论文, 中国科学技术大学, 合肥.)。因此, 本研究仅使用东方白鹳在9:00-18:00 (UTC + 8)之间且迁徙飞行速度超过14 km/h的数据(Bengtsson et al, 2014)。为了减少短时停歇和数据缺失导致的差异, 本研究对日迁徙飞行速度进行了估算。日迁徙飞行速度(km/h)是每天从9:00至18:00 (UTC + 8)所有迁徙飞行速度的平均值。飞行方向定义为每个飞行阶段的起点和终点之间的矢量方向。将日迁徙飞行速度与850 mb压力处风的U和V变量进行关联分析(Shamoun-Baranes et al, 2003)。
利用在时空上最接近每个GPS点的U和V风分量来计算顺/逆风分量(和鸟类飞行方向相同/相反) (Shamoun-Baranes et al, 2003)。850 mb风的数据与海拔1,500 m处的风向和风速有关, 同时与较低海拔处的风速也存在可预测的幂律关系(Şen et al, 2012), 通常从离地面1,000 m开始, 海拔每增加100 m, 风速增加0.1 m/s (Parent & Ilinca, 2011)。因此, 利用850 mb风数据建模可以很好地研究东方白鹳在地面以上350-960 m处飞行时利用的风速和风向, 其中东方白鹳在春季和秋季迁徙中顺风的相对收益为正值, 逆风为负值。
1.5 统计分析
本研究使用配对t检验或Wilcoxon符号秩检验, 检验每个迁徙参数(迁徙开始和结束时间除外)在春/秋迁徙之间的差异。当迁徙参数满足正态分布和方差齐性时, 使用配对t检验进行分析; 当迁徙参数不满足正态分布或方差齐性时, 使用Wilcoxon符号秩检验进行分析。同时, 使用Wilcoxon符号秩检验分析春/秋迁徙之间和顺/逆风条件下日迁徙飞行速度的差异。用广义加性模型(generalized additive model, GAM)检验日迁徙飞行速度与风的关系。所有建模和统计均在R 3.6中完成(R Development Core Team, 2017)。
2 结果
2.1 迁徙路线与停歇地
渤海湾越冬个体(N = 13)秋季从俄罗斯远东和中国东北的度夏地开始迁徙, 迁徙途中主要在黑龙江省松花江流域、辽宁省盘锦市湿地、河北省唐山市曹妃甸区和天津市北大港水库进行停歇, 最后到达渤海湾越冬。翌年春季, 个体(N = 9)从渤海湾开始迁徙, 停歇于松花江流域, 最后返回度夏地(图1)。
图1
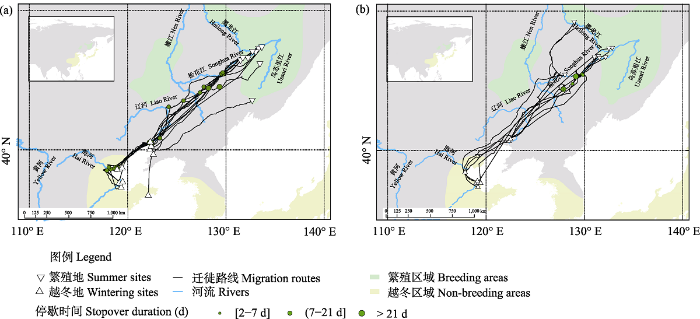
图1
东方白鹳渤海湾越冬群体幼鸟迁徙路线和停歇位点图。(a)秋季迁徙(N = 13); (b)春季迁徙(N = 9)。
Fig. 1
Migration routes and stopover sites of juvenile Oriental Storks overwintering in Bohai Bay. (a) Autumn migration (N = 13); (b) Spring migration (N = 9).
2.2 春秋季迁徙参数
渤海湾越冬群体幼鸟通常在10月下旬离开度夏地, 11月下旬到达越冬地。春季迁徙通常开始于4月下旬, 结束于5月上旬(其中特殊个体os 011于7月16日离开越冬地, 7月22日到达度夏地)。通过比较春季和秋季迁徙策略(表1, 图2; 追踪个体迁徙参数表见附录2), 发现尽管迁徙距离(图2a)在两个季节间没有显著差异, 但春季迁徙飞行速度明显快于秋季(P < 0.05; 图2e), 从而春季迁徙飞行时间更短(P < 0.05; 图2f); 与此同时, 春季停歇位点数和秋季相似, 但春季停歇时间(P = 0.05)更短, 春季迁徙持续时间明显短于秋季(P < 0.05; 图2b), 春季迁徙速度更快(P < 0.05)。
表1 东方白鹳渤海湾越冬群体幼鸟秋季和春季迁徙参数统计表。加粗字体表示有显著性差异。
Table 1
| 迁徙参数 Parameters | 秋季 Autumn (mean ± SD, N = 13) | 春季 Spring (mean ± SD, N = 9) | P |
|---|---|---|---|
| 迁徙开始时间 Departure date | 24 October ± 8 | 23 April ± 35 | |
| 迁徙结束时间 Arrival date | 21 November ± 17 | 4 May ± 31 | |
| 迁徙持续时间 Migration duration (d) | 28.0 ± 21.2 | 11.2 ± 8.7 | < 0.05 |
| 迁徙距离 Migration distance (km) | 1,566.9 ± 325.9 | 1,533.8 ± 424.8 | 0.69 |
| 迁徙速度 Migration speed (km/d) | 92.4 ± 61.7 | 211.1 ± 118.2 | < 0.05 |
| 迁徙飞行速度 Travel speed (km/d) | 185.5 ± 72.0 | 280.4 ± 62.0 | < 0.05 |
| 迁徙飞行时间 Travel duration (d) | 10.3 ± 6.5 | 5.9 ± 2.5 | < 0.05 |
| 停歇时间 Stopover duration (d) | 17.8 ± 18.2 | 5.4 ± 9.7 | 0.05 |
| 停歇位点数 Number of stopovers | 1.8 ± 1.5 | 0.7 ± 1.1 | 0.07 |
图2
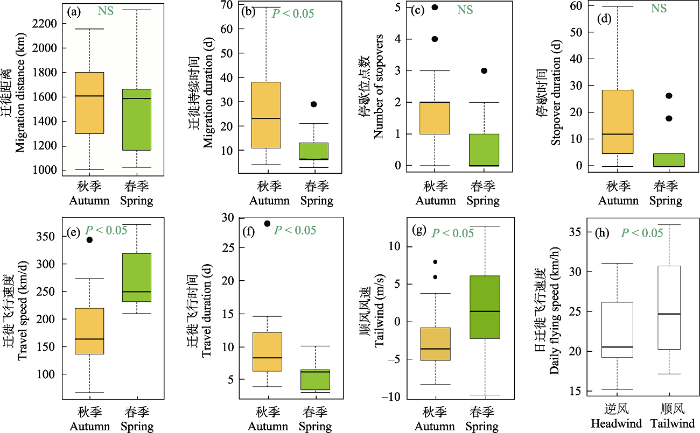
图2
东方白鹳渤海湾越冬群体幼鸟春季(绿色; N = 9)和秋季(橙色; N = 13)主要迁徙参数箱线图。“NS”表示不显著。
Fig. 2
Box plots of the main migration parameters of the spring (green; N = 9) and autumn (orange; N = 13) migrations of the juvenile Oriental Storks wintering in the Bohai Bay. “NS” indicates no significant difference.
2.3 风向和风速对迁徙飞行速度的影响
图3
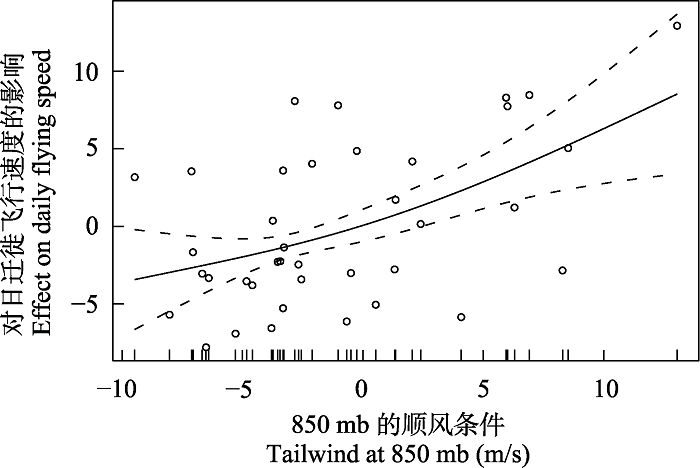
图3
850 mb的顺风(X轴)与日迁徙飞行速度(Y轴)的最佳拟合函数图(GAM模型)。空心圆点表示原始数据点, 虚线表示协变量周围的两个标准误差边界。沿X轴的垂直线表示850 mb顺风数据点的分布。
Fig. 3
GAM plot from the best fitting function of tailwind at 850 mb (X-axis) on the daily flying speed (Y-axis). Circles represent the orgin data points. The dashed line represents the two standard error boundaries around the covariate. The vertical line along the X-axis represents the distribution of the 850 mb tailwind data points.
3 讨论
3.1 东方白鹳幼鸟春秋季迁徙参数差异
比较渤海湾越冬群体春/秋迁徙策略差异可知, 该群体春/秋迁徙距离相似, 但春季迁徙飞行速度显著快于秋季, 且春季停歇时间短于秋季。因此, 东方白鹳春季迁徙持续时间显著短于秋季。
与已有研究数据相比①(①樊淑娟 (2020) 基于卫星追踪研究东方白鹳(Ciconia boyciana)的迁徙生态学. 硕士学位论文, 中国科学技术大学, 合肥.), 长江越冬群体幼鸟的迁徙距离约为渤海湾越冬群体幼鸟的1.7倍, 两个群体秋季迁徙的开始时间、结束时间、持续时间和停歇时间差异不大; 但两个群体的春季迁徙开始时间、持续时间和停歇时间有明显差异, 长江越冬群体幼鸟比渤海湾越冬群体幼鸟提早迁徙1个月(3月28日 ± 26天, 4月23日 ± 35天), 于相似时间到达(5月4日 ± 31天, 5月4日 ± 41天)。长江越冬群体幼鸟的迁徙持续时间为渤海湾越冬群体幼鸟的3倍(36.8 ± 31.8天, 11.2 ± 8.7天), 停歇时间为5倍(25.5 ± 29.9天, 5.4 ± 9.7天)。东方白鹳春季迁徙的顺风条件优于秋季, 春季迁徙的飞行速度快于秋季, 两个群体之间的迁徙速度相似(附录3)。两个群体幼鸟春季迁徙持续时间差异较大, 原因为长江越冬群体幼鸟先出发1个月, 到达渤海湾后, 进行停歇并和渤海湾越冬群体一起出发迁徙至度夏地。渤海湾越冬群体的迁徙距离短, 能够更好地预测度夏地的情况, 所以春季迁徙开始时间较晚(Coppack & Both, 2002)。
3.2 风向和风速对迁徙策略的影响
东方白鹳是主要利用上升热气流迁徙的翱翔鸟类(Flack et al, 2018), 风向条件可以解释东方白鹳渤海湾越冬群体幼鸟日迁徙飞行速度在春/秋季之间的差异。顺风强度(850 mb)与平均日迁徙飞行速度呈显著正相关, 由于春季的顺风条件显著优于秋季的逆风条件, 这使得春季迁徙飞行速度显著快于秋季。这一结果也表明, 春季和秋季迁徙时的迁徙飞行速度差异约为100 km/d, 可以用这两个季节风速的差别来定量理解, 即假定每天飞行8 h, 风速差异约为4 m/s, 则每小时14 km, 每天为112 km, 和日迁徙飞行速度差异形成的日迁徙飞行距离差异基本相似。虽然该群体春秋两季迁徙距离相似, 但由于风向条件使得春季迁徙飞行时间显著短于秋季。
东方白鹳的长江越冬群体幼鸟迁徙距离长, 个体在春季和秋季都要利用顺风, 由于春季的顺风(4.4 ± 5.6 m/s)条件明显优于秋季(0.5 ± 5.6 m/s), 在秋季迁徙过程中几乎没有从顺风条件中获得净收益, 所以春季的日迁徙飞行速度比秋季快(Fan et al, 2020)。相对而言, 渤海湾越冬群体幼鸟迁徙距离短, 春季利用顺风迁徙可更快到达度夏地; 秋季逆风迁徙, 迁徙飞行速度慢, 迁徙飞行时间和停歇时间长。因此, 尽管翱翔鸟类主要依赖上升热气流迁徙以节约能量, 但顺风条件也是其迁徙的有利因素。
致谢
感谢野外捕捉团队的贡献以及中国洪河国家级自然保护区和俄罗斯Bastak保护区对野外捕捉工作提供的支持; 感谢谢彦波老师对数据分析提供的帮助, 孟凡娟老师对文章英文部分的润色, 黄筑同学对本文的文字和图标多次帮助修改; 陈艺雯同学对文章的文字润色。感谢本文责任编委及匿名专家提出的诸多建设性意见。
附录 Supplementary Material
附录1 2016-2018年追踪的14只东方白鹳的详细信息
Appendix 1 Detailed information of 14 Oriental Storks tracked during 2016-2018
附录2 东方白鹳渤海湾越冬群体秋季和春季个体迁徙参数
Appendix 2 Individual migration parameters in autumn and spring of Oriental Storks wintering population in Bohai Bay
附录3 东方白鹳幼鸟长江越冬群体和渤海湾越冬群体秋季和春季迁徙参数
Appendix 3 Autumn and spring migration parameters of juvenile Oriental Storks overwintering population in Yangtze River and Bohai Bay
参考文献
Wind as selective agent in bird migration
DOI:10.2307/3676347 URL [本文引用: 1]
Environmental effects on flying migrants revealed by radar
DOI:10.1111/ecog.03995
[本文引用: 1]

Migratory animals are affected by various factors during their journeys, and the study of animal movement by radars has been instrumental in revealing key influences of the environment on flying migrants. Radars enable the simultaneous tracking of many individuals of almost all sizes within the radar range during day and night, and under low visibility conditions. We review how atmospheric conditions, geographic features and human development affect the behavior of migrating insects and birds as recorded by radars. We focus on flight initiation and termination, as well as in-flight behavior that includes changes in animal flight direction, speed and altitude. We have identified several similarities and differences in the behavioral responses of aerial migrants including an overlooked similarity in the use of thermal updrafts by very small (e.g. aphids) and very large (e.g. vultures) migrants. We propose that many aerial migrants modulate their migratory flights in relation to the interaction between atmospheric conditions and geographic features. For example, aerial migrants that encounter crosswind may terminate their flight or continue their migration and may also drift or compensate for lateral displacement depending on their position (over land, near the coast or over sea). We propose several promising directions for future research, including the development and application of algorithms for tracking insects, bats and large aggregations of animals using weather radars. Additionally, an important contribution will be the spatial expansion of aeroecological radar studies to Africa, most of Asia and South America where no such studies have been undertaken. Quantifying the role of migrants in ecosystems and specifically estimating the number of departing birds from stopover sites using low-elevation radar scans is important for quantifying migrant-habitat relationships. This information, together with estimates of population demographics and migrant abundance, can help resolve the long-term dynamics of migrant populations facing large-scale environmental changes.
Movements, home-range size and habitat selection of mallards during autumn migration
DOI:10.1371/journal.pone.0100764 URL [本文引用: 1]
Sex, wingtip shape, and wing-loading predict arrival date at a stopover site in the Swainson’s Thrush (Catharus ustulatus)
DOI:10.1093/auk/124.4.1388 URL [本文引用: 1]
Predicting life-cycle adaptation of migratory birds to global climate change
Cyclical helping hands: Seasonal tailwinds differentially affect migrating Oriental Storks (Ciconia boyciana) travel speed
DOI:10.1186/s40657-020-00196-8 URL [本文引用: 3]
From local collective behavior to global migratory patterns in White Storks
DOI:10.1126/science.aap7781 URL [本文引用: 1]
Observation on behavior of adult oriental White Stork in nesting period
DOI:10.1007/BF02855541 URL [本文引用: 1]
What selective pressures have driven the evolution of deferred northward migration by juvenile waders
DOI:10.2307/3677117 URL [本文引用: 1]
The NCEP/NCAR 40-year reanalysis project
DOI:10.1175/1520-0477(1996)077<0437:TNYRP>2.0.CO;2 URL [本文引用: 1]
Quantifying flow-assistance and implications for movement research
DOI:10.1016/j.jtbi.2012.05.026 URL [本文引用: 1]
Birds: Blowin’ by the wind
DOI:10.1007/s10336-006-0061-9 URL [本文引用: 1]
Interspecific comparison of the performance of soaring migrants in relation to morphology, meteorological conditions and migration strategies
DOI:10.1371/journal.pone.0039833 URL [本文引用: 1]
The trans-equatorial loop migration system of Eleonora’s falcon: Differences in migration patterns between age classes, regions and seasons
Seasonal modulation of flight speed among nocturnal passerine migrants: Differences between short- and long-distance migrants
DOI:10.1007/s00265-014-1789-5 URL [本文引用: 1]
Anti-icing and de-icing techniques for wind turbines: Critical review
DOI:10.1016/j.coldregions.2010.01.005 URL [本文引用: 1]
Movement patterns and stopover ecology of Wilson’s warblers during spring migration on the lower Colorado River in southwestern Arizona
DOI:10.1525/cond.2008.8602 URL [本文引用: 1]
Tracking through life stages: Adult, immature and juvenile autumn migration in a long-lived seabird
DOI:10.1371/journal.pone.0072713 URL [本文引用: 1]
R: A Language and Environment for Statistical Computing
The challenges of the first migration: Movement and behaviour of juvenile vs. adult White Storks with insights regarding juvenile mortality
DOI:10.1111/1365-2656.12525
PMID:27046512
[本文引用: 1]
Migration conveys an immense challenge, especially for juvenile birds coping with enduring and risky journeys shortly after fledging. Accordingly, juveniles exhibit considerably lower survival rates compared to adults, particularly during migration. Juvenile white storks (Ciconia ciconia), which are known to rely on adults during their first fall migration presumably for navigational purposes, also display much lower annual survival than adults. Using detailed GPS and body acceleration data, we examined the patterns and potential causes of age-related differences in fall migration properties of white storks by comparing first-year juveniles and adults. We compared juvenile and adult parameters of movement, behaviour and energy expenditure (estimated from overall dynamic body acceleration) and placed this in the context of the juveniles' lower survival rate. Juveniles used flapping flight vs. soaring flight 23% more than adults and were estimated to expend 14% more energy during flight. Juveniles did not compensate for their higher flight costs by increased refuelling or resting during migration. When juveniles and adults migrated together in the same flock, the juvenile flew mostly behind the adult and was left behind when they separated. Juveniles showed greater improvement in flight efficiency throughout migration compared to adults which appears crucial because juveniles exhibiting higher flight costs suffered increased mortality. Our findings demonstrate the conflict between the juveniles' inferior flight skills and their urge to keep up with mixed adult-juvenile flocks. We suggest that increased flight costs are an important proximate cause of juvenile mortality in white storks and likely in other soaring migrants and that natural selection is operating on juvenile variation in flight efficiency.© 2016 The Authors. Journal of Animal Ecology © 2016 British Ecological Society.
Individual tracking reveals first breeding of Oriental Storks at age 2 years in the wild
The effect of wind, season and latitude on the migration speed of White Storks Ciconia ciconia, along the eastern migration route
DOI:10.1034/j.1600-048X.2003.03079.x URL [本文引用: 4]
Atmospheric conditions create freeways, detours and tailbacks for migrating birds
DOI:10.1007/s00359-017-1181-9 URL [本文引用: 1]
Arrival timing and seasonal reproductive performance in a long-distance migratory landbird
DOI:10.1007/s00265-004-0855-9 URL [本文引用: 1]
Age-related differential timing of spring migration within sexes in passerines
DOI:10.1676/0043-5643(2002)114[0264:ARDTOS]2.0.CO;2 URL [本文引用: 1]
Difference in migration pattern between adult and immature birds using satellites
DOI:10.1642/0004-8038(2002)119[0832:DIMPBA]2.0.CO;2 URL [本文引用: 1]
Wind effects on the migration routes of trans-Saharan soaring raptors: Geographical, seasonal, and interspecific variation
DOI:10.1093/cz/zow008
PMID:29491895
[本文引用: 1]
Wind is among the most important environmental factors shaping birds' migration patterns. Birds must deal with the displacement caused by crosswinds and their behavior can vary according to different factors such as flight mode, migratory season, experience, and distance to goal areas. Here we analyze the relationship between wind and migratory movements of three raptor species which migrate by soaring-gliding flight: Egyptian vulture, booted eagle, and short-toed snake eagle. We analyzed daily migratory segments (i.e., the path joining consecutive roosting locations) using data recorded by GPS satellite telemetry. Daily movements of Egyptian vultures and booted eagles were significantly affected by tailwinds during both autumn and spring migrations. In contrast, daily movements of short-toed eagles were only significantly affected by tailwinds during autumn migration. The effect of crosswinds was significant in all cases. Interestingly, Egyptian vultures and booted eagles showed latitudinal differences in their behavior: both species compensated more frequently at the onset of autumn migration and, at the end of the season when reaching their wintering areas, the proportion of drift segments was higher. In contrast, there was a higher drift at the onset of spring migration and a higher compensation at the end. Our results highlight the effect of wind patterns on the migratory routes of soaring raptors, with different outcomes in relation to species, season, and latitude, ultimately shaping the loop migration patterns that current tracking techniques are showing to be widespread in many long distance migrants.
The Far East taiga forest: Unrecognized inhospitable terrain for migrating Arctic-nesting waterbirds
DOI:10.7717/peerj.4353 URL [本文引用: 1]
Differences in migration strategies between rescued juvenile and adult Oriental White Storks (Ciconia boyciana) and the conservation implications
DOI:10.1016/j.gecco.2021.e01760 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}