



物种的空间格局一直是生态学研究的基本内容之一(Watt, 1947; Wiegand et al, 2007; Velázqez et al, 2016)。由于物种的空间格局与生态过程之间存在密切关联, 因此通过定量分析物种的空间格局可以帮助了解植物种群的生物学特性(如生活史策略、喜光、耐阴和克隆特性等)、种间作用(竞争或易化)及其与环境因子之间的相互关系等重要的生态学过程(Condit et al, 2000; Lin et al, 2011; 黄运峰等, 2012; 王鑫厅等, 2020)。近些年来, 点格局分析方法作为最主要的物种空间格局定量分析方法在国内外被广泛使用(Perry et al, 2002; Wang et al, 2010; 王鑫厅等, 2012; 黄小等, 2020), 其最大的特点是可以比较灵活地选择完全空间随机模型(complete spatial randomness, CSR)、泊松聚块模型(Poisson cluster process/Neyman-Scott process, NS)、嵌套双聚块模型(nested double-cluster process, DC)等不同的零模型来定量分析实测空间格局特征, 进而比较准确地分析物种格局形成的生态过程(王鑫厅等, 2012, 2020)。此外, 点格局分析方法还克服了传统方法只能分析单一尺度物种空间分布的局限性, 使得对物种格局的研究能够在相对连续的尺度上进行(Wiegand & Moloney, 2004; 王鑫厅等, 2011)。因此, 通过基于不同零模型的点格局分析方法可以更好地帮助揭示不同环境条件下植物种群空间格局特征及其形成的潜在生态学机制。
水文过程是湿地最基本的生态过程, 湿地环境中水文条件的变化会显著影响植物的生长发育、繁殖分配和竞争关系, 从而决定植物群落的物种组成及其空间分布格局的形成(杨娇等, 2014; Zhang et al, 2014; Shi et al, 2015)。首先, 湿地植物对水位变化的响应最直观地反映在植物的个体大小性状上, 如单株生物量、株高、节间距、分枝数、基径等形态特征。研究发现, 随着水位的升高, 植物的个体大小一般会降低, 如某些对水分敏感的挺水植物毛秆野古草(Arundinella hirta)等会通过降低个体大小来适应水淹胁迫(Zeng et al, 2006; 董海鹏等, 2022)。大量小型化的植物个体往往会高度聚集在一起, 从而使得种群格局在空间上表现为强聚集分布或斑块化特征(王鑫厅等, 2012; Ren et al, 2020)。其次, 随着水位的升高, 湿地植物会通过降低有性繁殖分配, 增加无性繁殖体的营养分配, 以确保个体自我更新。例如, 随水位或土壤含水量的增加, 淡水沼泽湿地植物对有性繁殖的投入大大减少, 植物趋向于通过无性繁殖快速占领空间, 从而影响种群密度及其空间格局(Shi et al, 2015; Bai et al, 2021; 董海鹏等, 2022)。再者, 植物间的互作类型(竞争或促进)及互作强度亦会随着环境条件的变化而改变, 从而造成植物空间格局的变化。胁迫梯度假说(stress gradient hypothesis)认为, 随着环境胁迫的增加, 植物间的促进作用强度将增加, 而竞争作用将减弱(Bertness & Callaway, 1994; Callaway & Walker, 1997)。研究表明, 在湿地生态系统中, 当水位对植物生长产生胁迫时, 物种间的竞争关系会逐渐减弱或者消失, 甚至转化为促进作用(杨娇等, 2014)。例如, 随着水淹程度的增加, 湿地植物的种内竞争作用减弱, 而促进作用增强, 从而形成聚集分布(Pennings et al, 2005; 于国磊, 2011)。目前, 关于异质生境下湿地不同水位环境如何通过改变植物繁殖分配、个体性状和竞争关系共同影响植物种群空间格局的研究仍然较少。
三江平原地处我国东北地区, 是由黑龙江、乌苏里江与松花江3条大江在东北平原内部汇流并经过不断的冲积形成的。同时, 由于特殊的地形和水文环境条件, 三江平原也是我国最大的淡水沼泽分布区之一(赵魁义, 1990)。在三江平原的沼泽分布区内, 最主要的优势物种为毛薹草(Carex lasiocarpa)、漂筏薹草(C. pseudocuraica)、狭叶甜茅(Glyceria spiculosa)、小叶章(Deyeuxia angustifolia)等具有克隆习性的多年生草本植物(高福元和石福习, 2015), 这些植物在空间上一般呈带状分布于季节性积水和常年积水等不同水位环境下的沼泽湿地中(Shi et al, 2015)。因而, 不同水文情势驱动的植物个体大小、繁殖分配和植物相互关系变化可能会对三江平原沼泽湿地中不同植物种群空间格局造成显著影响。鉴于此, 本文选择三江平原季节性积水沼泽与常年积水沼泽为研究对象, 基于完全空间随机模型、泊松聚块模型和嵌套双聚块模型的点格局分析方法, 研究了不同水位环境条件下4个主要优势植物种群的空间格局类型及其形成机制, 探讨了水位环境条件变化对物种空间格局的影响, 以期揭示三江平原沼泽湿地植物种群对水位环境的响应规律以及适应策略。
1 研究方法
1.1 样地选择与取样
实验于2012年9月在中国科学院三江平原沼泽湿地生态试验站设置的沼泽湿地长期观测场进行(47°35′ N, 133°31′ E)。该样地位于别拉洪河与浓江的河间地带, 地势平坦开阔, 平均海拔56 m, 土壤主要为腐殖质沼泽土、草甸沼泽土和草甸土, 潜育层广泛发育, 在蝶形洼地形成的常年积水区和季节性积水区沼泽湿地, 由内向外植物群落一般以毛薹草群落和小叶章-毛薹草混合群落为主, 植被的总覆盖度70%-95%。季节性积水沼泽区土壤含水量在80%-120%之间, 优势种小叶章的盖度为45%-75%; 常年积水沼泽区积水深度在10-45 cm之间, 优势种毛薹草的盖度为60%-90%。该区属温带大陆性季风气候, 年平均气温2.5℃, 年降水量558 mm, 主要集中于7-8月, 占全年降水量的65%以上(Shi et al, 2015)。
本研究在参照摄影定位法的基础上, 采用邻近格子法测定了栖息于常年积水区和季节性积水区沼泽湿地群落中4个优势物种毛薹草、漂筏薹草、狭叶甜茅和小叶章的空间分布(王鑫厅等, 2011; 高福元和石福习, 2015)。测定时, 在季节性积水区和常年积水区各设置1个具有代表性的4 m × 4 m正方形群落样方, 先用竹杆将每个4 m × 4 m 的样方分割成256个25 cm × 25 cm的亚样方, 再用25 cm × 25 cm的小样方框(框内小格子的面积为5 cm × 5 cm)以逆时针方向依次放置256次, 记录样方内主要植物植株个体或株丛的相对坐标位置。每个物种分别选择6-10株不同个体, 测定其在不同水位环境条件下的种群密度、单株生物量、株高和基径等个体大小特征。
1.2 点格局分析
式中, A为样方面积; λ为模型估计参数, 指样方内物种个体密度; dij为两随机点间的距离; wij为边界效应修正的权重; n是样方内点事件的数量; I为指示函数, 当dij ≤ r时, I = 1.0, 当dij > r时, I = 0。
当L(r) = 0时, 个体显示为完全空间随机分布, L(r) > 0时为聚集分布, L(r) < 0时为均匀分布。在应用L(r)函数进行点格局分析时, 一般会将实测值与选定的零模型通过Monte-Carlo方法绘制置信区间来判断目标点格局的分布状况, 如果实测值在包迹线以上, 则符合聚集分布; 在包迹线以内, 则符合随机分布; 在包迹线以下, 则符合均匀分布。本研究中, 设定步长为1 cm, Monte-Carlo拟合20次, 置信水平为95%, 置信区间通过最大值和最小值获得。
1.3 零模型选择
1.3.1 完全空间随机模型
完全空间随机模型是最简单、最常用的一种零模型, 其实质是均质泊松过程(homogeneous Poisson process, HP)。在均质泊松过程中, 假设任何一点(或个体)在研究区域内任何一个位置上出现(或发生)的机会是相同的, 物种的空间分布不受任何生物或非生物过程影响, 属于空间上完全随机。均质泊松过程的Ripley’s K函数同上文定义。
1.3.2 泊松聚块模型
泊松聚块模型又称作均质托马斯过程(homogeneous Thomas process, TH), 主要模拟物种子代个体受扩散限制而聚集分布于母株周围的情况, 描述的是一种聚块机制(Diggle, 2003; 黄运峰等, 2012)。在均质托马斯过程中, 假设母体事件以完全空间随机过程发生, 而每一个母体在其周围按一定的概率分布产生随机数量的子代个体, 且这些子代个体在空间分布上遵循某双变量概率密度函数。如果子代个体的数量遵循泊松分布且其在空间的位置相对母体而言符合双变量高斯分布, 那么, 子代个体的发生就符合泊松聚块过程(Wiegand & Moloney, 2004; Lin et al, 2011; 王鑫厅等, 2012)。泊松聚块模型的Ripley’s K函数表达式如下:
其中, r为尺度, ρ为母体Poisson分布的密度或强度, σ2为高斯分布的方差。
1.3.3 嵌套双聚块模型
且
其中, 参数r、ρ和σ2的意义同泊松聚块模型, 下角标1代表第一代, 下脚标2代表第二代。
1.4 数据处理
2 结果
2.1 种群密度和个体大小差异
图1
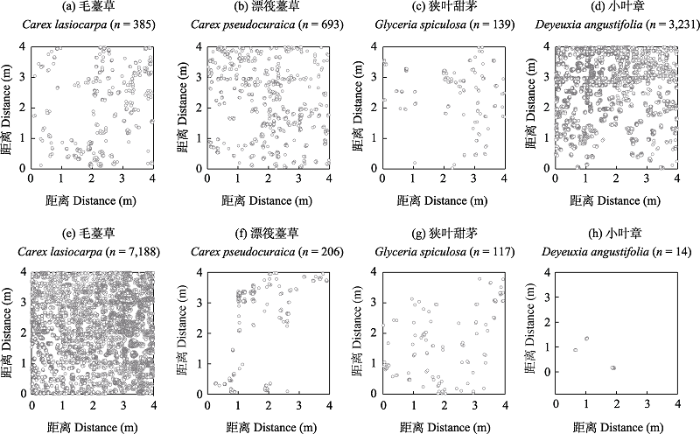
图1
三江平原4个优势物种在季节性积水沼泽(a-d)和常年积水沼泽(e-h)的个体分布点位图
Fig. 1
Individual distribution locus map of four dominant species in seasonal inundated marsh (a-d) and perennial inundated marsh (e-h) in the Sanjiang Plain
图2
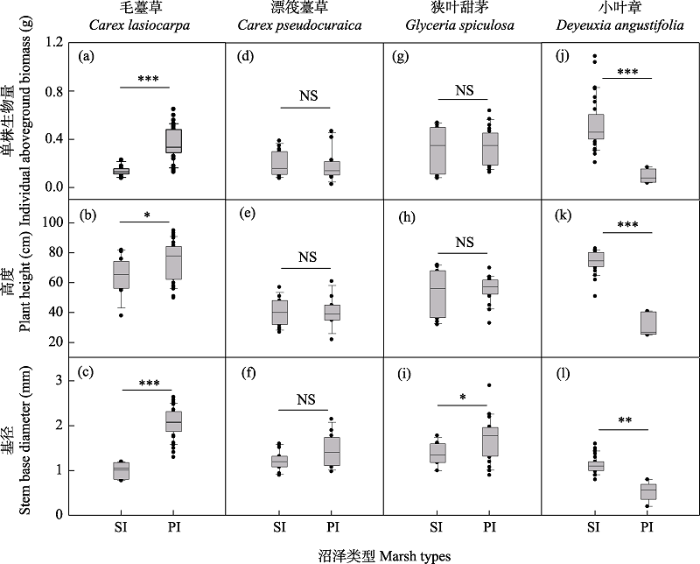
图2
三江平原季节性积水沼泽(SI)和常年积水沼泽(PI) 4个优势物种的单株生物量、植株高度和基径变化。NS: P > 0.05; * P < 0.05; ** P < 0.01; *** P < 0.001。
Fig. 2
Variations in individual aboveground biomass, plant height and stem base diameter of four dominant species in seasonal inundated (SI) marsh and perennial inundated (PI) marsh in the Sanjiang Plain. NS, P > 0.05; * P < 0.05; ** P < 0.01; *** P < 0.001.
2.2 季节性积水环境中的点格局分析
在季节性积水沼泽中, 在0-200 cm尺度范围内, 毛薹草、漂筏薹草、狭叶甜茅和小叶章基本上都偏离了完全随机模型, 且不同物种的偏离程度有所不同(图3)。其中, 狭叶甜茅偏离完全随机模型的程度最大(图3c), 其次是毛薹草和漂筏薹草(图3a, b), 小叶章随着尺度的增大聚集强度呈逐渐增大趋势(图3d); 通过泊松聚块模型模拟发现, 毛薹草、漂筏薹草、狭叶甜茅和小叶章种群分别在0-14 cm (图3e)、0-21 cm (图3f)、0-35 cm (图3g)和0-21 cm (图3h)尺度上偏离了泊松聚块模型, 在15-200 cm (图3e)、22-200 cm (图3f)、36-200 cm (图3g)和22-200 cm (图3h)尺度上符合泊松聚块模型; 进一步通过嵌套双聚块模型模拟发现, 在小尺度范围内偏离了泊松聚块模型的毛薹草、漂筏薹草、狭叶甜茅和小叶章种群在0-200 cm尺度上均符合嵌套双聚块模型(图3i-l)。
图3
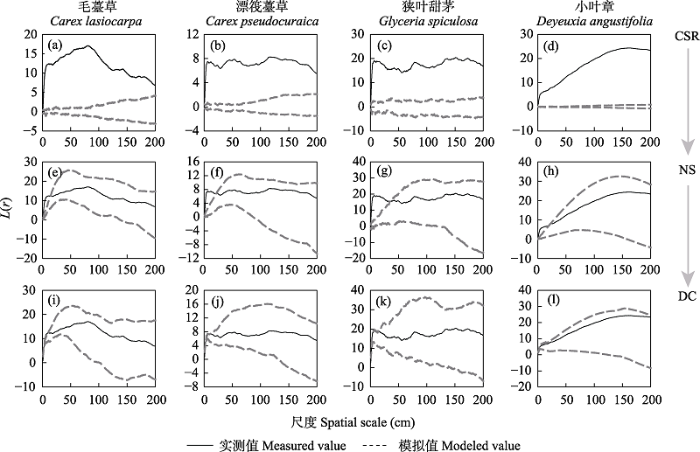
图3
4个优势物种在季节性积水环境中基于完全空间随机模型(CSR)、泊松聚块模型(NS)和嵌套双聚块模型(DC)的点格局分析
Fig. 3
Point pattern analysis of four dominant species based on complete spatial randomness (CSR), Poisson cluster process (NS) and nested double-cluster process (DC) in seasonal inundated (SI) marsh
2.3 常年积水环境中的点格局分析
在常年积水沼泽中, 毛薹草、漂筏薹草、狭叶甜茅和小叶章也均偏离了完全随机模型, 且不同物种偏离程度不同(图4)。其中, 小叶章偏离完全随机模型的程度最大(图4d), 其次是漂筏薹草和狭叶甜茅(图4b, c), 毛薹草偏离完全随机模型的程度最小(图4a); 通过泊松聚块模型模拟发现, 毛薹草、漂筏薹草、狭叶甜茅分别在0-17 cm (图4e)、0-37 cm (图4f)、0-35 cm (图4g)尺度上偏离了泊松聚块模型, 在18-200 cm (图4e)、38-200 cm (图4f)、36-200 cm (图4g)尺度上符合泊松聚块模型, 而小叶章种群则在0-200 cm尺度上完全符合泊松聚块模型(图4h)。进一步通过嵌套双聚块模型模拟发现, 在小尺度范围内偏离了泊松聚块模型的毛薹草、漂筏薹草和狭叶甜茅也均在0-200 cm尺度上符合嵌套双聚块模型(图4i-k)。
图4
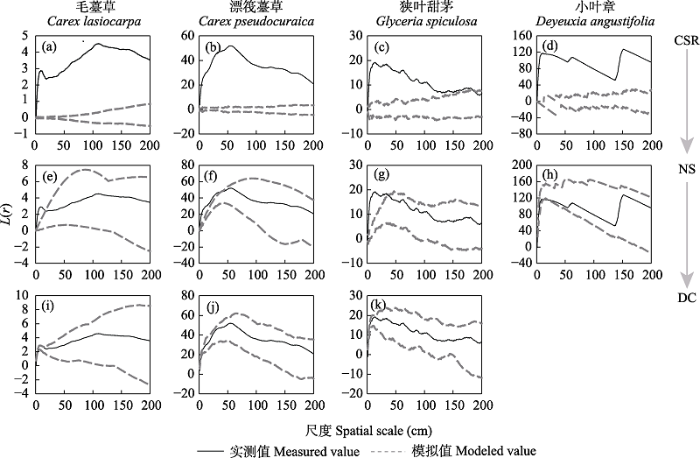
图4
4个优势物种在常年积水环境中基于完全空间随机模型(CSR)、泊松聚块模型(NS)和嵌套双聚块模型(DC)的点格局分析
Fig. 4
Point pattern analysis of four dominant species based on complete spatial randomness (CSR), Poisson cluster process (NS) and nested double-cluster process (DC) in perennial inundated (PI) marsh
3 讨论
3.1 三江平原沼泽湿地主要植物聚集分布格局及形成机制
本文应用点格局分析中的完全空间随机模型模拟了三江平原沼泽湿地毛薹草、漂筏薹草、狭叶甜茅和小叶章4个主要植物种群在小尺度范围内的空间格局类型。研究发现, 不管是季节性积水环境还是常年积水环境, 这4个植物种群在0-200 cm尺度范围内基本都偏离了完全空间随机模型, 说明三江平原沼泽湿地主要植物种群表现为聚集分布, 且聚集距离主要在0-50 cm之间, 随着尺度的增大, 聚集强度有逐渐减弱的趋势, 这与之前的研究结果基本一致(高福元和石福习, 2015; Ren et al, 2020)。实际上, 三江平原沼泽湿地植物种群的这种集体性聚集分布往往使之在空间上呈现出斑块化特征(高福元和石福习, 2015; Ren et al, 2020; Bai et al, 2021)。然而, 利用简单的完全空间随机模型仅能检验植物种群在不同尺度下的空间格局类型, 无法描述植被聚集分布或斑块化的形成机制。因此, 很早就有学者通过泊松聚块模型和嵌套双聚块模型来检验植物种群在空间上呈斑块化现象的潜在机制, 并取得了很好的效果(王鑫厅等, 2011, 2012)。
本研究选择泊松聚块模型模拟了扩散限制作用对毛薹草、漂筏薹草、狭叶甜茅和小叶章物种空间格局的影响。结果表明, 在季节性积水和常年积水沼泽中, 4个主要植物种群在50-200 cm尺度范围内均符合泊松聚块模型, 说明扩散限制作用是三江平原沼泽湿地植物空间分布最重要的生态过程。扩散限制作为中性理论的一个重要基础, 对植物群落结构及空间分布具有显著的影响, 是造成物种空间聚集的重要原因之一(Levine & Murrell, 2003; Wiegand et al, 2007; Lin et al, 2011; 黄运峰等, 2012)。然而, 与种子扩散限制引起的树木空间聚集分布不同的是, 在湿地生态系统中, 草本植物在小尺度范围内的集体性聚集分布可能与其营养繁殖特性引起的扩散限制有很大关系(高福元和石福习, 2015)。由于上述4个物种都是具有克隆习性的挺水植物(王丽等, 2009; Shi et al, 2015; Bai et al, 2021), 它们一般会通过根状茎等无性繁殖方式在母体周围产生新个体而呈现出聚集分布, 所以三江平原沼泽湿地优势种群表现出的集体性聚集分布格局当为母体营养繁殖的结果(高福元和石福习, 2015; Ren et al, 2020), 这与之前在草原生态系统的研究结果也基本一致(王鑫厅等, 2011, 2012; 李月飞等, 2019)。因此, 在三江平原沼泽湿地中, 植物这种特殊的无性繁殖方式引起的扩散限制可能是决定植物种群形成聚集分布的最主要因素。
但是, 泊松聚块模型并不能完全模拟4个主要优势植物种群在0-200 cm尺度范围内的空间分布, 除了常年积水环境中的小叶章以外, 在季节性积水和常年积水沼泽中的所有物种在小尺度范围内均偏离了泊松聚块模型, 说明除了植物无性繁殖引起的扩散限制作用外, 可能还有其他生态过程影响着三江平原沼泽湿地的物种空间格局。在本研究中, 由于选择的是地表平坦、尺度较小的生境均质群落片断, 这就排除了生境异质性对物种聚集分布的影响, 因此, 进一步选择嵌套双聚块模型模拟了物种空间格局形成的多聚块机制。结果表明, 季节性积水环境中的4个物种以及常年积水环境中的毛薹草、漂筏薹草和狭叶甜茅, 在0-200 cm尺度上均符合嵌套双聚块模型, 说明这些物种在大聚块中均分布着较高密度的小聚块, 存在多聚块机制。研究表明, 具有克隆繁殖特性的植物一般会通过营养繁殖在母体周围产生分株种群, 分株种群又继续繁殖形成下一级分株系统, 所有的分株系统在空间上就构成了一种多聚块的斑块分布特征(高福元和石福习, 2015)。因此, 合理选择具有不同生态指示意义的零模型可以更好地揭示水位环境胁迫条件下植物种群的空间分布格局及其形成机制。
3.2 不同水位环境对沼泽湿地植物空间格局的影响
湿地环境中水文条件的不稳定性及空间分布不均造成的生境异质性被认为是影响植物生长繁育和空间格局变化的首要非生物因子(Loreti et al, 2016; 董海鹏等, 2022)。首先, 本研究结果表明, 随着水位的升高, 4个优势物种的种群密度发生了显著变化, 说明水位条件对不同植物的繁殖分配方式造成了较大影响(Bai et al, 2021; 董海鹏等, 2022), 种群密度的变化可能会通过种内竞争变异引起物种聚集强度的变化(Pennings et al, 2005; 于国磊, 2011; 杨娇等, 2014)。研究表明, 种内竞争变弱会引起植物种群内部个体之间产生正向的生态关系, 容易导致种群自身分布格局向聚集分布转变, 种群的聚集强度往往会增大; 相反, 种内竞争增强则会引起种群内部之间产生负向的生态关系或无关, 种群格局则会由聚集分布向随机分布或均匀分布转变, 聚集强度往往会降低(赵成章等, 2010; 郝红敏等, 2017)。从季节性积水区到常年积水区, 毛薹草种群密度增加了18.7倍, 说明其根茎繁殖能力显著增强, 这与之前的盆栽实验结果一致(王丽等, 2009; Shi et al, 2015)。毛薹草种群密度的增大导致其种群内部的个体竞争大大增加, 所以随水位的升高其偏离完全随机模型的程度降低, 聚集强度变弱; 相反, 高水位胁迫使得小叶章种群密度变为仅有0.88株/m2, 说明高水位条件会显著抑制小叶章的营养繁殖投入(Xie et al, 2009; Shi et al, 2015; 董海鹏等, 2022)。所以, 随着水位的升高, 小叶章种群内部的个体竞争变弱, 其偏离完全随机模型的程度变大, 聚集强度也变高。此外, 高水位胁迫条件也不同程度地降低了漂筏薹草和狭叶甜茅种群密度, 但仅观测到漂筏薹草增加了其聚集强度, 狭叶甜茅的聚集强度变化不大, 这可能是由于随着水位的上升狭叶甜茅种群密度比漂筏薹草种群密度降低较少的原因。因此, 在沼泽湿地生态系统中, 植物营养繁殖投入变异及其引起的种内竞争关系强弱变化可能是植物种群空间格局响应不同水位环境的重要因素之一。
除种群密度的影响外, 三江平原沼泽湿地季节性积水和常年积水环境条件下主要植物种群这种集体性的多聚块和斑块化分布格局也符合胁迫梯度假说。研究表明, 在环境胁迫条件下, 同种个体之间往往可以通过相互帮助来共同抵御外界不利条件, 且这种易化或促进作用与环境胁迫的强度成正相互作用关系(Bertness & Callaway, 1994; Callaway & Walker, 1997), 这也就意味着环境胁迫程度越强, 植物种群越容易形成空间上的聚块以及多聚块格局。例如, 近期研究发现, 分布于内蒙古的大针茅(Stipa grandis)草原原生群落中竞争是主要的相互作用, 所以大针茅种群在小尺度范围内呈均匀分布, 而在长期过度放牧群落中植物正相互作用逐渐占据主导地位, 从而导致种群表现为强聚集分布格局, 且存在多聚块机制(王鑫厅等, 2020)。三江平原沼泽湿地中, 季节性积水和常年积水环境都存在不同程度的水位缺氧胁迫, 因此, 所有物种基本都符合嵌套双聚块模型, 即在大聚块中分布较高密度的小聚块, 这种格局是种群易化作用的结果。值得注意的是, 小叶章种群在季节性积水环境中符合嵌套双聚块模型, 存在多聚块机制。但是在常年积水环境中, 小叶章种群仅符合泊松聚块模型, 说明在高水位胁迫条件下其空间格局的大聚块中不存在较高密度的小聚块, 这主要是因为与其他3种植物相比, 小叶章缺少呼吸根, 在高水位环境条件下幼苗个体很难进行大规模的营养繁殖(Colmer, 2003; Xie et al, 2009; Ren et al, 2020), 在4 m × 4 m的取样范围内, 小叶章种群仅存在3个大聚块, 也间接证明了这一点。因此, 环境胁迫造成的种群易化作用对沼泽湿地不同水位环境对植物种群空间格局的影响也具有非常重要的作用。
此外, 植物个体性状的表型可塑性变异亦被认为是影响植物种群空间格局响应环境胁迫的重要生物因素。例如, 在严重退化的内蒙古典型羊草(Leymus chinensis)群落中, 植物个体会趋于小型化, 这主要是由于为了抵御过度放牧的选择压力, 大量小型化的羊草种群个体会聚集在一起, 从而使得种群格局表现为嵌套双聚块过程(王鑫厅等, 2012)。本研究中, 随着水位的升高, 三江平原沼泽湿地植物的个体大小存在3种变异(图2): (1)小叶章种群植物个体也同羊草一样表现出了小型化现象。但是, 这种小型化与内蒙古典型草原植物在放牧压力下的小型化原因不同。三江平原沼泽湿地小叶章植物个体小型化并不是种群密度增大引起的, 主要是由于高水位胁迫造成植物个体缺氧影响了植物的发育, 从而最终影响种群的空间格局(Xie et al, 2009; Shi et al, 2015; Ren et al, 2020; 董海鹏等, 2022)。(2)随水位的增加, 毛薹草种群植物个体显著增大, 趋于大型化。早期的水位控制实验已表明, 水分增多会明显促进毛薹草植物的根茎繁殖, 同时也会增加其地上和地下生物量(王丽等, 2009; Zhang et al, 2014; Shi et al, 2015)。高水位胁迫下, 毛薹草大量繁殖形成的多聚块及其个体大型化可能有利于植物个体之间增加通气组织对氧气的传输性, 以便更好地适应沼泽湿地缺氧环境(Colmer, 2003)。(3)随水位的增加, 漂筏薹草和狭叶甜茅植物个体并没有发生明显的小型化或大型化趋势, 这可能与这2种植物自身的水位生态位有关(Shi et al, 2015; Bai et al, 2021)。因此, 环境胁迫造成的植物表型可塑性变异并不一定只是个体小型化, 而可能存在多种变异途径, 在今后的研究当中需引起更多关注。
总之, 本研究基于3种零模型对不同水位环境条件下的三江平原沼泽湿地4种主要优势植物毛薹草、漂筏薹草、狭叶甜茅和小叶章种群进行了小尺度点格局分析, 发现三江平原沼泽湿地植物普遍存在着多聚块过程, 水位环境胁迫主要通过扩散限制、种内竞争、易化作用以及个体大小变异等生物因素共同决定着沼泽湿地植物种群空间分布格局的形成。
附录 Supplementary Material
附录1 使用泊松聚块模型和嵌套双聚块模型的单变量分析结果
Appendix 1 Results of univariate analyses using Poisson cluster process and the univariate double-cluster model process
参考文献
Functional traits response to flooding depth and nitrogen supply in the helophyte Glyceria spiculosa (Gramineae)
DOI:10.1016/j.aquabot.2021.103449 URL [本文引用: 5]
Positive interactions in communities
DOI:10.1016/0169-5347(94)90088-4 URL [本文引用: 2]
Contribution to the discussion of Dr. Ripley’s paper
Competition and facilitation: A synthetic approach to interactions in plant communities
DOI:10.1890/0012-9658(1997)078[1958:CAFASA]2.0.CO;2 URL [本文引用: 2]
Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots
Spatial patterns in the distribution of tropical tree species
DOI:10.1126/science.288.5470.1414
PMID:10827950
[本文引用: 1]

Fully mapped tree census plots of large area, 25 to 52 hectares, have now been completed at six different sites in tropical forests, including dry deciduous to wet evergreen forest on two continents. One of the main goals of these plots has been to evaluate spatial patterns in tropical tree populations. Here the degree of aggregation in the distribution of 1768 tree species is examined based on the average density of conspecific trees in circular neighborhoods around each tree. When all individuals larger than 1 centimeter in stem diameter were included, nearly every species was more aggregated than a random distribution. Considering only larger trees (>/= 10 centimeters in diameter), the pattern persisted, with most species being more aggregated than random. Rare species were more aggregated than common species. All six forests were very similar in all the particulars of these results.
Effect of water level difference in heterogeneous habitats on sexual reproductive allocation of Deyeuxia angustifolia
异质生境水位差异对小叶章有性繁殖分配的影响
Small-scale point pattern analysis based on different null models for detecting spatial patterns of dominant species in Sanjiang Plain, China
基于不同零模型的三江平原沼泽湿地主要物种小尺度点格局分析
Structure and dynamic characteristics of shrub patch of Artemisia ordosica in the cross areas of wind water erosion
水蚀风蚀交错带黑沙蒿灌丛斑块种群结构及动态特征研究
Structure and spatial distribution pattern of a native Metasequoia glyptostroboides population in Hubei
DOI:10.17520/biods.2019283
[本文引用: 1]
Metasequia glyptostroboides is an endemic and endangered species in China, and has therefore been heavily researched. M. glyptostroboides’ distribution seems to have a recognizable spatial pattern, with a clear potential mechanism. To test this, we analyzed the structure and diameter classes and height level classes from survey data of parent M. glyptostroboides trees with a pairwise correlation function g (r) and three null models (complete spatial randomness, heterogeneous Poisson process, and antecedent condition) in a point pattern analysis. The results of the spatial distribution patterns showed that: (1) There was 5,661 individuals of the native M. glyptostroboides population in the study area. Within this population, 31 of which were dead, 40 trees were on the verge of death, and 465 trees were classified as “weak”, with morphological characteristics and growth conditions associated with these trees such as shoot breakage, ant damage, and lightning strike. (2) Both the diameter-class structure and the height-class structure were spindle-shaped, with poor natural renewal; (3) Based on Complete Spatial Randomness, the native M. glyptostroboides population showed a random distribution at all scales, the middle- aged trees showed an aggregation distribution on a small scale (r < 3,300 m), and the adult trees and the older trees appeared an aggregation distribution on a larger scale (r < 4,700 m). The aggregation scales of the population and the different age groups were 0-3,000 m, 0-2,100 m, 0-2,900 m and 0-2,500 m, respectively, and followed a weak random distribution and uniform distribution. (4) Based on Complete Spatial Randomness, there were positive correlations between the three growth stages at all scales that weakened when habitat heterogeneity was excluded (only a positive correlation between 0-2,800 m). In summary, the M. glyptostroboides population is decreasing gradually with poor population regeneration. The population is presented mainly as cluster distribution with positive relationship among all age classes, due to habitat heterogeneity, diffusion limitation and intra-species competition according to the existing spatial pattern.
水杉原生种群结构及空间分布格局
DOI:10.17520/biods.2019283
[本文引用: 1]
水杉(Metasequoia glyptostroboides)是我国特有濒危极小种群物种, 其种群的状态一直被国内外学者广泛关注。分析现存水杉原生种群结构和空间分布格局及其空间关联性, 可以从空间格局角度深入认识水杉原生种群结构和分布格局及可能的形成机理。本文基于湖北利川境内水杉原生种群的野外调查数据, 分析其径级和高度级结构, 同时运用点格局分析中的成对相关函数g(r)以及3个零模型(完全空间随机模型、异质泊松模型、先决条件模型)分析水杉原生种群空间分布格局、各龄级空间分布格局及空间关联性。结果表明: (1)分布于研究区域内的水杉原生种群个体数共5,663株, 已死亡33株, 现存活5,630株, 其中40株濒临死亡, 465株处于衰弱状态, 部分个体呈现不同的形态特征和生长状况, 断梢和蚁害最为常见, 雷击是最致命的危害。(2)水杉原生种群结构分析显示, 其径级结构和高度级结构均呈纺锤型, 自然更新不良。(3)基于完全空间随机模型, 水杉原生种群在各尺度下均呈现聚集分布, 中龄树在较小尺度(r < 3,300 m)上呈现聚集分布, 成年树和老龄树在较大尺度(r < 4,700 m)上呈现聚集分布; 排除生境异质性影响后, 聚集尺度均减小, 所以种群及不同龄级的聚集尺度分别为0-3,000 m、0-2,100 m、0-2,900 m和0-2,500 m, 随后呈现为微弱的随机分布和均匀分布。(4)基于完全空间随机模型, 3个龄级之间在所有尺度均为正关联; 在排除生境异质性影响后, 不同龄级的正关联尺度减小, 均在0-2,800 m呈正关联。综上, 水杉原生种群个体数量正呈现逐步减少的趋势, 种群主要呈聚集分布, 各个龄级间具有正向的关系, 从现有的空间格局来看, 生境异质性、扩散限制和种内竞争是导致该格局的主要原因。
Spatial pattern of trees in tropical lowland rain forest in Bawangling of Hainan Island, China
DOI:10.3724/SP.J.1258.2012.00269 URL [本文引用: 4]
海南岛霸王岭热带低地雨林树木的空间格局
DOI:10.3724/SP.J.1258.2012.00269
[本文引用: 4]
树木空间格局及其形成过程是物种共存及生物多样性维持机制研究的一个重要方面。该文以海南岛两个1 hm <sup>2</sup>的典型热带低地雨林老龄林森林动态样地为基础, 通过4个点格局模型(均质Poisson过程、异质Poisson过程、均质Thomas过程和异质Thomas过程)模拟扩散限制和生境异质性作用对树木空间分布格局的影响, 并分析不同空间尺度下(< 2 m, 2-5 m, 5-10 m, 10-15 m, 15-20 m和20-25 m)不同作用的相对重要性。结果表明: 热带低地雨林的所有树木总体上呈现聚集分布的空间格局, 随着尺度的增大, 聚集强度逐渐减小。树种在模拟空间分布格局最优模型中的比例由高到低分别是: 均质Thomas过程, 均质Poisson过程、异质Thomas过程和异质Poisson过程。扩散限制作用是形成热带低地雨林树木空间分布格局最重要的生态过程, 其次是完全随机作用以及生境异质性和扩散限制的联合作用, 而生境异质性的作用最小。不同空间尺度上模拟各树种空间分布格局的最优模型比例差异显著, 扩散限制作用能够在多数空间尺度上模拟多个树种的空间分布格局, 其次为随机作用; 生境异质性和扩散限制的联合作用主要在小尺度(0-5 m)影响树种分布, 而生境异质性在较大尺度(15-25 m)上影响树种的空间分布格局。
The community-level consequences of seed dispersal patterns
DOI:10.1146/ecolsys.2003.34.issue-1 URL [本文引用: 1]
Point pattern analysis of an Artemisia scoparia seedling population under different soil conditions in a desert steppe
荒漠草原不同土壤条件下猪毛蒿幼苗种群的点格局分析
Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation
DOI:10.1007/s00442-010-1718-x URL [本文引用: 4]
Plant responses to flooding stress
DOI:S1369-5266(16)30088-7
PMID:27322538
[本文引用: 1]
Most plant species cannot survive prolonged submergence or soil waterlogging. Crops are particularly intolerant to the lack of oxygen arising from submergence. Rice can instead germinate and grow even if submerged. The molecular basis for rice tolerance was recently unveiled and will contribute to the development of better rice varieties, well adapted to flooding. The oxygen sensing mechanism was also recently discovered. This system likely operates in all plant species and relies on the oxygen-dependent destabilization of the group VII ethylene response factors (ERFVIIs), a cluster of ethylene responsive transcription factors. An homeostatic mechanism that controls gene expression in plants subjected to hypoxia prevents excessive activation of the anaerobic metabolism that could be detrimental to surviving the stress.Copyright © 2016 Elsevier Ltd. All rights reserved.
Plant zonation in low-latitude salt marshes: Disentangling the roles of flooding, salinity and competition
DOI:10.1111/jec.2005.93.issue-1 URL [本文引用: 2]
Illustrations and guidelines for selecting statistical methods for quantifying spatial pattern in ecological data
DOI:10.1034/j.1600-0587.2002.250507.x URL [本文引用: 1]
R: A Language and Environment for Statistical Computing
Response of individual sizes and spatial patterns of Deyeuxia angustifolia to increasing water level gradient in a freshwater wetland
DOI:10.1007/s11356-020-08283-5 URL [本文引用: 7]
Modelling spatial patterns
Plant zonation patterns reflected by the differences in plant growth, biomass partitioning and root traits along a water level gradient among four common vascular plants in freshwater marshes of the Sanjiang Plain, Northeast China
DOI:10.1016/j.ecoleng.2015.04.054 URL [本文引用: 10]
An evaluation of the state of spatial point pattern analysis in ecology
DOI:10.1111/ecog.2016.v39.i11 URL [本文引用: 1]
Responses of Carex lasiocarpa clonal reproduction to water regimes at different growth stages
不同生长阶段毛苔草(Carex lasiocarpa)克隆繁殖对水文情势的响应
Species associations in an old-growth temperate forest in Northeastern China
DOI:10.1111/jec.2010.98.issue-3 URL [本文引用: 1]
Population spatial pattern of Stipa grandis and its response to long-term overgrazing
DOI:10.17520/biods.2019276 URL [本文引用: 3]
典型草原大针茅种群空间格局及对长期过度放牧的响应
DOI:10.17520/biods.2019276
[本文引用: 3]
种群空间格局是生态学研究的基本问题之一。典型草原带由于过度放牧退化严重, 原生群落罕见, 探讨原生群落的种群空间格局具有重要生态学意义。大针茅(Stipa grandis)草原是典型草原区广泛分布的主要群落类型, 1979年围封的大针茅样地, 是目前保存完整的大针茅草原原生群落。本文选择大针茅草原原生群落和长期过度放牧群落, 应用O-Ring函数结合不同零假设模型分析了大针茅种群的空间格局。结果表明: 在原生群落中大针茅种群在小尺度范围内呈均匀分布, 而在长期过度放牧群落中则表现为聚集分布。这说明在大针茅草原原生群落中竞争是主要的相互作用, 而在长期过度放牧群落中正相互作用居主导, 验证了胁迫梯度假说; 同时证明长期过度放牧改变了种群空间格局。
Point pattern analysis based on different null models for detecting spatial patterns
DOI:10.3724/SP.J.1003.2012.08163 URL [本文引用: 9]
基于不同零模型的点格局分析
DOI:10.3724/SP.J.1003.2012.08163
[本文引用: 9]
在种群空间格局研究中, 定量分析格局及其形成过程已成为生态学家的主要目标。在量化分析的众多方法中, 点格局分析是最常用的方法, 而在选择零模型时, 完全空间随机模型以外的复杂零模型很少使用, 实际上, 这些零模型可能有助于认识格局的内在特征。为此, 我们在研究实例中, 选择完全空间随机模型(complete spatial randomness)、泊松聚块模型(Poisson cluster process)和嵌套双聚块模型(nested double-cluster process)对典型草原处于不同恢复演替阶段的羊草(Leymus chinensis)种群空间格局进行了分析。结果发现: 完全空间随机模型仅能检测种群在不同尺度下的格局类型; 而通过泊松聚块模型和嵌套双聚块模型检验表明, 在恢复演替的初期阶段, 羊草种群在小尺度范围内偏离泊松聚块模型, 而在整个取样范围内完全符合嵌套双聚块模型; 随着恢复演替时间的推移, 在恢复演替的后期, 在整个取样尺度上, 羊草种群与泊松聚块模型相吻合。这是很有意义的生态学现象。这一实例表明在应用点格局分析种群空间格局时, 仅通过完全空间随机模型的检验来分析格局特征, 或许很难论证复杂的生态过程, 而选择一些完全空间随机模型以外的较复杂的零模型, 可能发现一些有价值的生态学现象, 对揭示格局掩盖下的内在机制有所裨益。
Point pattern analysis of dominant populations in a degraded community in Leymus chinensis + Stipa grandis steppe in Inner Mongolia, China
DOI:10.3724/SP.J.1258.2011.01281 URL [本文引用: 4]
羊草 + 大针茅草原退化群落优势种群空间点格局分析
DOI:10.3724/SP.J.1258.2011.01281
[本文引用: 4]
草原退化的特征主要表现为群落生产力的大幅下降和植物个体的小型化, 同时, 退化的草原生态系统与一定强度的放牧压力保持平衡而相对稳定。该文应用摄影定位法测定了羊草+大针茅草原退化群落中4个优势种群羊草(Leymus chinensis)、米氏冰草(Agropyron michnoi)、大针茅(Stipa grandis)、糙隐子草(Cleistogenes squarrosa)的空间格局。点格局分析结果表明: 在空间分布格局上, 4个优势种群均偏离完全随机模型和泊松聚块模型, 而符合嵌套双聚块模型, 也就是说, 其空间格局表现为聚集分布, 且在大聚块中分布着较高密度的小聚块。严重退化的草原群落中优势种群的嵌套双聚块空间分布格局当属一种集体行为, 是种群适应过度放牧压力的一种表现形式, 为过度放牧导致的退化草原群落的主要特征之一。这种生态学现象同植物个体小型化一样, 是种群易化(正相互作用)的结果。在过度放牧的胁迫下, 种群通过改变个体性状及个体在空间的分布状况实现自我帮助, 以抵御外界的放牧压力达到自我保护, 从而维持退化草原生态系统与放牧压力间的相对平衡。
Pattern and process in the plant community
DOI:10.2307/2256497 URL [本文引用: 1]
Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering
DOI:10.1890/06-1350.1 URL [本文引用: 2]
Rings, circles, and null models for point pattern analysis in ecology
DOI:10.1111/oik.2004.104.issue-2 URL [本文引用: 4]
Increased nutrient supply facilitates acclimation to high-water level in the marsh plant Deyeuxia angustifolia: The response of root morphology
DOI:10.1016/j.aquabot.2008.12.004 URL [本文引用: 3]
Research progress in response of plants in wetlands to water level change
湿地植物对水位变化的响应研究进展
Effects of waterlogging on intraspecific interactions of the clonal herb Alternanthera philoxeroides
DOI:10.3724/SP.J.1258.2011.00973 URL [本文引用: 2]
水淹对克隆植物空心莲子草种内关系的影响
DOI:10.3724/SP.J.1258.2011.00973
[本文引用: 2]
为检验“水淹程度可以改变植物种内关系的类型和强度”的假说, 将克隆植物空心莲子草(Alternanthera philoxeroides) 的3种不同密度的植株(每盆种植1、4或16株)置于4种不同的水淹处理下(水位分别为–20 (不水淹)、0、20或40 cm), 研究不同水淹程度对空心莲子草种内关系的影响。随着植株密度和水淹程度的增加, 空心莲子草的生长显著减慢, 但密度效应在不同的水淹处理下显著不同。在不发生水淹的情况下, 植株密度对生长的负面(竞争)效应最强; 在水位为0和20 cm的情况下,植株密度对生长的效应仍为负面的, 但影响强度相对减小; 而在水位为40 cm的情况下, 空心莲子草植株的生物量随着植株密度的增大而倾向于增加。进一步分析相对邻体效应时发现, 随着水淹程度的增加, 相对邻体效应显著增加, 并且数值从负值(不水淹)逐渐变为正值(40 cm水位下)。这些结果支持胁迫梯度假说, 表明水淹可以影响植物的种内关系, 即随着水淹程度的增加, 植物种内竞争作用减弱, 而易化作用增强。
Different responses of sexual and asexual reproduction of Arundinella hirta to flooding
DOI:10.1007/s11515-005-0012-5 URL [本文引用: 1]
Effects of hydrology and competition on plant growth in a freshwater marsh of northeast China
DOI:10.1080/02705060.2013.825821 URL [本文引用: 2]
Fine-scale spatial patterns of Stellera chamaejasme population in degraded alpine grassland in upper reaches of Heihe, China
黑河上游高寒退化草地狼毒种群小尺度点格局分析
DOI:10.3773/j.issn.1005-264x.2010.11.009
[本文引用: 1]
植被斑块化是自然界的一种普遍现象, 斑块的形成和变化对植物种群格局的形成和变化具有重要影响。在黑河上游祁连山北坡高寒退化草地, 采用点格局分析方法, 研究了小尺度上狼毒(Stellera chamaejasme)种群的种群密度、组成格局以及分布格局。结果表明: 随着狼毒种群分盖度的增大, 种群密度、领地密度和组成格局呈现规律性的变化, 斑块内部狼毒种群的数量出现增减交替变化趋势, 组成格局规律明显; 狼毒种群的分布格局表现出与尺度关联的变化趋势, 在31%–40%分盖度下, 狼毒种群在所有尺度上表现为随机分布, 在41%–50%、51%–60%、61%–70%、71%–80%分盖度下随着尺度增大, 分布格局的基本模式为: 随机—聚集—随机或均匀—随机—聚集—随机分布, 在聚集状态下, 聚集强度不同。以成株为核心的斑块内部种群表现为随机分布或均匀分布, 相对于外部表现为聚集分布, 随着成株个体数量的逐渐增多, 种群竞争关系由种间竞争转化为种内竞争, 促进了斑块扩张与合并、斑块增多与吞并, 从而实现了种群扩散。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}