



2021年10月12日, 在昆明召开的联合国《生物多样性公约》第十五次缔约方大会上, 中国宣布正式设立三江源、大熊猫、东北虎豹、海南热带雨林和武夷山等首批国家公园, 其中武夷山国家公园主要保护热带、亚热带重要森林生态系统。武夷山国家公园(27°31°-27°55° N, 117°24°-117°59° E)位于福建省武夷山市、建阳区、邵武市、光泽县的交界处, 规划总面积1,001.41 km2 (贾岚等, 2021)。公园蕴藏着丰富的野生动植物资源, 不但物种多样性高、特有种多, 还是很多物种形成和分化的中心, 是一批动植物的模式标本产地(陈昌笃, 1999), 已记录高等植物2,800余种, 野生脊椎动物558种, 昆虫类6,849种(游文茂, 2020)。
武夷山国家公园位于武夷山脉的北端, 其所处的地理位置是闽江和赣江的源头, 也是闽江水系、汀江水系与赣江水系、信江水系的天然分水岭(张春英等, 2008)。国家公园及其周边水系网络复杂多样、分布密集, 公园主要是以建溪上游支流为主, 由北向南依次为崇阳溪、九曲溪和麻阳溪。崇阳溪为闽江支流建溪的上游, 发源于武夷山脉铜钹山, 干流河道162 km; 九曲溪为崇阳溪上游主要支流, 发源于武夷山公园内的桐木关, 长62.8 km, 自西向东流至武夷宫汇入崇阳溪; 麻阳溪是崇阳溪中游主要支流, 发源于建阳区的黄坑乡黄毛岗山, 河道长136 km (姚有则等, 2000; 张玉等, 2010; 何美峰等, 2012)。国家公园西侧为北溪, 北溪是富屯溪的支流, 发源于光泽县的大岐山, 干流河道全长70.54 km, 自北向西南流至城关回龙潭处汇入富屯溪(光泽县地方志编纂委员会, 1994)。崇阳溪、麻阳溪、九曲溪、北溪最终汇入闽江。国家公园北侧的铅山河是信江的一级支流, 发源于赣闽边界武夷山桐木关的右侧, 主河道长82 km, 于凤来村汇入信江( 张凯强 (2011) 铅山河生态基流确定及生态调度模型初步研究. 硕士学位论文, 南昌大学, 南昌.)。
历史上对于武夷山地区的鱼类有过一些调查, 如《福建鱼类志》(上卷)记述了1975-1979年武夷山国家公园所在的南平市和建阳区明确有标本记录的鱼类分别为45种和52种(福建鱼类志编写组, 1984)。丁桂枝和郑闽泉(1990)调查总结了武夷山市建溪和富屯溪的鱼类资源, 记录鱼类138种。何美峰等(2012)于2007-2008年调查了崇阳溪的鱼类资源, 采集到鱼类73种。李万宝(2014)于2007-2012年对崇阳溪鱼类资源进行了调查, 共记录鱼类75种。宋小晶等(2017)讨论了华东武夷山-仙霞岭地区整体的鱼类区系和动物地理, 共记录鱼类282种, 但并未具体谈及武夷山地区。祝于红等(2017)发现武夷山脉西坡3个自然保护区中武夷山自然保护区的鱼类多样性最高。总的来说, 上述研究工作仅限于鱼类物种组成和区系的定性分析, 缺少定量研究; 或仅涉及到现在武夷山国家公园范围的部分区域, 且仅从物种丰富度角度开展了研究, 缺少与鱼类功能多样性相关的研究。
武夷山国家公园作为武夷山脉生物多样性热点地区, 围绕其鱼类多样性开展系统性的调查与分析工作, 对于了解武夷山国家公园及其周边地区鱼类物种组成和多样性现状, 探讨武夷山脉在我国淡水鱼类区系中的地位, 以及后续鱼类保护工作的开展具有重要意义和参考价值。
1 材料与方法
1.1 标本采集和数据收集
作者于2021年旱季(4月)和雨季(7月)分别对武夷山国家公园及其周边水域进行了鱼类多样性调查。流经武夷山国家公园内的河流包括九曲溪和麻阳溪上游河段, 而崇阳溪、北溪、铅山河以及麻阳溪中下游河段为武夷山国家公园的周边水域(图1)。在急流、缓滩、洄水湾、深潭等鱼类栖息生境共设置采样点47处(图1)。基于国家公园的管理规定和可涉水条件, 尽量将不同河段的鱼类栖息生境类型点位保持一致。本次调查以地笼作为主要采集方式, 调查点位间统一地笼的规格、数量和放置时间; 同时辅以手抄网、撒网等其他方式进行物种丰富度的调查。在深潭只采用地笼收集鱼类标本。进行鱼类多样性比较和分析时, 为统一捕捞努力, 主要以地笼调查的渔获结果作为数据基础。手抄网、撒网等其他形式采集的结果, 以及附近农贸市场进行的调查和标本收集, 作为鱼类物种组成的重要补充。
图1
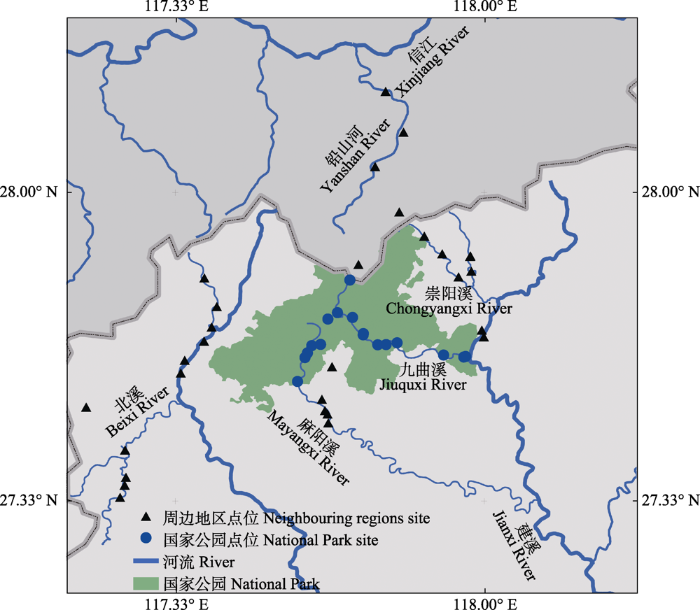
图1
武夷山国家公园及其周边地区调查点分布图
Fig. 1
Sampling sites in Wuyishan National Park and its adjacent areas
采集所得鱼类鉴定计数后, 部分放归河流, 剩余个体使用无水乙醇或10%的甲醛溶液固定, 作为存证标本长期保存于中国科学院动物研究所鱼类标本馆备查。同时, 作者对该馆保藏的武夷山地区的历史标本进行了数据采集, 并检索了已发表的历史文献、报告等。
1.2 物种鉴定与测量
标本鉴定主要参考《中国动物志•硬骨鱼纲•鲤形目(中卷)》(陈宜瑜, 1998)、《中国动物志•硬骨鱼纲•鲤形目(下卷)》(乐佩琦, 2000)以及《福建鱼类志》(福建鱼类志编写组, 1984)等志书, 物种有效学名依据《中国内陆鱼类物种与分布》(张春光和赵亚辉, 2016)、《中国生物物种名录(第二卷): 动物•脊椎动物(V)•鱼类》(张春光等, 2020)和Catalog of Fishes (
每种鱼类选取成鱼标本20尾进行测量, 标本量小于20尾的全部测量。测量工具为150 mm的电子游标卡尺, 精确到0.01 mm。
1.3 鱼类多样性分析
1.3.1 α多样性
式中, S为野外采集鱼类的总物种数, N为野外采集鱼类总个体数, Pi为第i种鱼类的个体数占鱼类总数的比例。
群落中个体数量占采集鱼类总个体数超过10%的为优势种, 其中占比最高的为第一优势种, 占比第二的为第二优势种, 占比第三的为第三优势种(李雪健等, 2020)。
1.3.2 功能多样性
鱼类功能多样性主要是关注与鱼类群落功能密切相关的功能性状, 例如鱼类的体型、口裂大小、口位、食性、胸鳍长等, 对这些功能性状赋值和量化, 以此来反映群落中物种功能的差异性(Tilman et al, 2001; Petchey & Gaston, 2002)。功能性状值的确定依据群落中鱼类功能特点的数值和范围来定义(Petchey & Gaston, 2006)。选择与鱼类栖息地、摄食、运动相关的功能特征值, 即体长、体宽、眼径、头长、口裂、胸鳍长、尾柄高等(Villéger et al, 2008)。根据测量个体计算平均性状值(Villéger et al, 2010), 平均性状值经log(x + 1)标准化后(Pease et al, 2012), 利用主成分分析(principal components analysis, PCA)对测量的鱼类功能特征值进行选择(Mao et al, 2021) (表1)。
表1 武夷山国家公园及其周边鱼类功能特征值
Table 1
| 功能特征值 Functional traits | 计算方法 Method | 生态学意义 Ecological meaning | 参考文献 Reference |
|---|---|---|---|
| 胸鳍相对长度 Relative pectoral fin length | 胸鳍长/体长 Pectoral fin length/Standard length | 比值越大, 则转向越强, 灵活性越高 The greater the ratio, the stronger the steering and the higher the flexibility | 1984 |
| 腹鳍相对长度 Relative pelvic fin length | 腹鳍长/体长 Pelvic fin length/Standard length | 比值越大, 则转向越强, 灵活性越高 The greater the ratio, the stronger the steering and the higher the flexibility | 本文 This paper |
| 尾鳍形状 Caudal fin shape | 尾鳍面积/(体长 × 体高) Caudal fin area/(Standard length × Body depth) | 比值越高, 游泳加速度越快 The higher the ratio of persistence and acceleration of swimming, the faster the swimming acceleration | 1984 |
| 尾柄相对高度 Relative caudal peduncle depth | 尾柄高/体高 Caudal peduncle depth/Body depth | 比值越高, 尾鳍摆动能力越强 The higher the ratio, the stronger the tail fin swinging ability | 本文 This paper |
| 口裂相对大小 Relative mouth opening | 口裂 × 口高/(体宽 × 体高) (Mouth depth × Mouth width)/(Body width × Body depth) | 最大捕食能力, 比值越高, 能力越强 Maximum predation ability, the higher the ratio, the stronger the ability | 2003 |
| 眼睛相对大小 Relative eye size | 眼径/头长 Eye diameter/Head length | 对食物的可视范围 Visual scope to food | 2006 |
| 体型指标I Body shape I | 体高/体长 Body depth/Standard length | 水体中的栖息位置 Habitat location in water body | 2000 |
| 体型指标II Body shape II | 体高/体宽 Body depth/Body width | 水体中的栖息位置 Habitat location in water body | 2000 |
| 眼睛相对位置 Relative position of eye | 眼径/头高 Eye diameter/Head depth | 在水体中的垂直位置 Vertical position in water body | 1979 |
| 食性 Feeding habits | 肉食性、杂食性(偏肉食性、偏植食性)、植食性 Carnivorous, omnivorous (partial carnivorous, partial phytophagous), phytophagous | 对食物的偏好 Food preference | - |
| 口位 Mouth position | 上位、亚上位、端位、亚下位、下位 Superior, sub-superior, terminal, sub- inferior, inferior | 不同位置表示获得食物组成不一样 Different location means different food composition | - |
式中, SFic表示群落i中鱼类占据的生态位空间, Rc表示功能特征值的绝对值。
式中, Sp为物种数, Wi、Wj分别为物种i和物种j的相对丰度, dist(i, j)为物种间的欧式距离。l为分支长, PEWl为分支长权重。
功能离散度(FDiv)表示在性状空间中鱼类功能性状散布的最大离散水平。FDiv指数越低, 说明群落中物种的生态位处于饱和状态, 物种之间的竞争越激烈。计算公式如下:
式中, Xik为物种i的性状k的值, gk为性状k的重心, dGi为物种i到性状k的欧氏距离,
2 结果
2.1 武夷山国家公园及其周边地区鱼类区系组成
根据野外调查、历史文献及馆藏标本记录, 并进行分类学厘定后得出, 武夷山国家公园及其周边区域共有鱼类115种, 隶属于5目18科62属。除去国外引入的尼罗罗非鱼(Tilapia niloticus)和欧洲鳗鲡(Anguilla anguilla), 土著鱼类为113种, 隶属于5目17科61属(附录1)。
从目级水平看, 调查区域主要以鲤形目鱼类为主, 有4科47属82种, 占物种总数的72.6%; 其次为鲈形目, 有5科5属16种, 占14.2%; 鲇形目5科6属11种, 占9.7%; 合鳃目2科2属2种, 占1.8%, 鳗鲡目为1科1属2种, 占1.8%。科级水平多样性较高, 共有17科, 其中鲤科鱼类物种最多, 共有66种, 占土著鱼类总物种数的58.4%; 其次为爬鳅科(10种), 占8.9%; 其他依次为虾虎鱼科(7种)、鲿科(7种)、花鳅科(5种)、鳢科(3种)、鮨鲈科(3种)、鳗鲡科(2种)、斗鱼科(2种), 只包含1种的科有鮡科、胭脂鱼科、鲇科、胡子鲇科、钝头鮠科、沙塘鳢科、刺鳅科和合鳃科。
在我国鲤科鱼类的12个亚科中, 该区域分布有10个。其中鮈亚科鱼类最多, 有12属23种, 占鲤科鱼类总科数的34.9%; 鲌亚科7属12种, 占18.2%; 鲃亚科4属7种, 占10.6%; 鱊亚科2属7种, 占10.6%; 鲴亚科2属5种, 占7.6%; 雅罗鱼亚科4属4种, 占6.1%;鱼丹亚科1属3种, 占4.6%; 鲤亚科和鲢亚科各2属2种, 各占3.0%, 鳅鮀亚科1属1种, 占1.5%。
从水系来看, 武夷山国家公园北坡分布有83种鱼类, 隶属于5目16科53属; 南坡分布有99种鱼类, 隶属于5目17科60属(附录1); 南北坡鱼类均以鲤科中的鮈亚科和鲌亚科鱼类为主, 但南坡鲤科中的鲃亚科、爬鳅科种类比北坡多, 南北坡鱼类区系具有一定的差异。武夷山国家公园的北坡和南坡各自特有的种类较少, 无特有属, 北坡包含长江水系特有鱼类5种, 占北坡总数的6.0%, 南坡闽江特有鱼类4种, 占南坡总数的4.0%。
调查区域分布有55种中国特有种(附录1), 占总种数的48.7%。该区域还分布有国家II级重点保护水生野生动物胭脂鱼(Myxocyprinus asiaticus)和花鳗鲡(Anguilla marmorata)。在该区域分布的鱼类中, 被评定为极危(CR)的物种有1种, 为胭脂鱼, 评定为濒危(EN)的物种有2种, 为日本鳗鲡(Anguilla japonica)和花鳗鲡; 受胁物种占总种数的2.7%, 占比较少。近危(NT)物种有4种, 为台湾白甲鱼(Onychostoma barbatula)、暗鳜(Siniperca obscura)、叉尾斗鱼(Macropodus opercularis)和圆尾斗鱼(M. chinensis)。评定为无危(LC)的物种有75种, 占66.4%, 数据缺乏(DD)的20种, 占17.7%, 还有11种鱼类在红色名录中未被评定。
2.2 野外调查
2.2.1 野外调查到的物种组成
野外调查共采集到鱼类70种, 隶属于5目16科47属, 占本区域全部淡水鱼类物种数的59.8%; 旱季采集到鱼类62种, 隶属于5目13科39属, 雨季采集到60种, 隶属于12科37属。其中中国特有种32种, 占实地调查鱼类总数的45.7%; 长江特有鱼类2种, 占2.9%; 闽江水系特有鱼类2种, 占2.9%。实地采集到受胁鱼类1种, 为日本鳗鲡。
从目级水平看, 鲤形目鱼类3科32属48种, 占物种总数的68.6%; 鲇形目5科5属9种, 占12.9%; 鲈形目4科4属10种, 占14.3%; 合鳃目2科2属2种, 占2.9%; 鳗鲡目1科1属1种, 占1.4%。科级水平多样性较高, 共有16科, 其中鲤科鱼类最多(40种), 占土著鱼类总种数的57.1%; 其次为虾虎鱼科(6种), 占8.57%; 鲿科5种, 占7.1%; 爬鳅科和花鳅科鱼类各4种, 各占5.7%; 鲇科、胡子鲇科、斗鱼科、钝头鮠科、鮡科、鮨鲈科、鳢科、沙塘鳢科、刺鳅科、鳗鲡科、合鳃科均只有1种, 各占1.4%。本次调查共采集到鲤科中的10个亚科鱼类。其中鮈亚科鱼类最多, 有11属18种, 占鲤科鱼类种数的45.0%; 鲃亚科、鲌亚科鱼类次之(各3属5种), 各占12.5%; 鱼丹亚科1属3种, 占7.5%; 鱊亚科2属3种, 占7.5%; 雅罗鱼亚科2属2种, 占5.0%; 鲴亚科、鲤亚科、鲢亚科和鳅鮀亚科鱼类均只有1属1种, 各占2.5%。
2.2.2 优势种组成
武夷山国家公园内的第一优势种为拟腹吸鳅(Pseudogastromyzon fasciatus), 占采集总个体数的28.5%; 第二优势种为武夷光唇鱼(Acrossocheilus wuyiensis); 第三优势种为半刺光唇鱼(A. hemispinus)。国家公园周边地区的第一优势种为点纹银鮈(Squalidus wolterstorffi), 占鱼类总数的21.6%; 第二优势种为福建小鳔鮈(Microphysogobio fukiensis), 占15.1%; 第三优势种为高体鳑鲏(Rhodeus ocellatus)。
5条河流中, 铅山河的优势种为点纹银鮈、张氏小鳔鮈(Microphysogobio zhangi)和拟腹吸鳅, 其中第一优势种点纹银鮈的个体数占铅山河鱼类总数的70.2%。崇阳溪的3个优势种分别为点纹银鮈、拟腹吸鳅和长鳍马口鱼(Opsariichthys evolans), 第一优势种点纹银鮈个体数占崇阳溪鱼类总数的16.6%。九曲溪的3个优势种为武夷光唇鱼、拟腹吸鳅、斑纹台鳅(Formosania stigmata), 第一优势种武夷光唇鱼占九曲溪鱼类总数的27.7%。麻阳溪的优势种为拟腹吸鳅、半刺光唇鱼、长鳍马口鱼, 第一优势种拟腹吸鳅占麻阳溪鱼类总数的16.9%。福建小鳔鮈、拟腹吸鳅、高体鳑鲏为北溪的3个优势种, 其中福建小鳔鮈为第一优势种, 占北溪鱼类总数的26.2%。从表2可知, 旱季的第一优势种为点纹银鮈; 雨季的则为福建小鳔鮈。
表2 武夷山国家公园鱼类的优势种季节变化
Table 2
| 物种 Species | 整体 Total | 旱季 Dry season | 雨季 Rainy season |
|---|---|---|---|
| 拟腹吸鳅 Pseudogastromyzon fasciatus | ++ | ++ | ++ |
| 长鳍马口鱼 Opsariichthys evolans | + | + | +++ |
| 福建小鳔鮈 Microphysogobio fukiensis | ++++ | + | ++++ |
| 点纹银鮈 Squalidus wolterstorffi | +++ | ++++ | + |
| 高体鳑鲏 Rhodeus ocellatus | + | +++ | + |
++++ 第一优势种; +++ 第二优势种; ++ 第三优势种; + 存在
++++ First dominant species; +++ Second dominant species; ++ Third dominant species; + existence
2.2.3 生态类型
图2
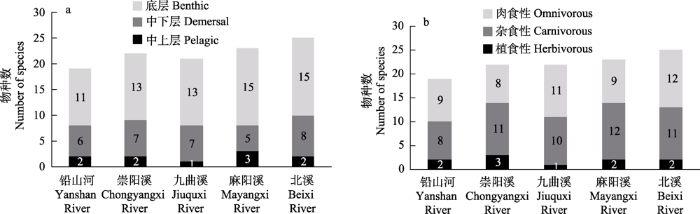
图2
武夷山国家公园5条河流鱼类按栖息水层(a)及食性(b)的生态分布
Fig. 2
Ecological distribution of fish in five rivers of Wuyishan National Park by habitation space (a) and food preference (b)
2.3 鱼类α多样性
2.3.1 鱼类物种多样性在空间尺度上的变化
图3
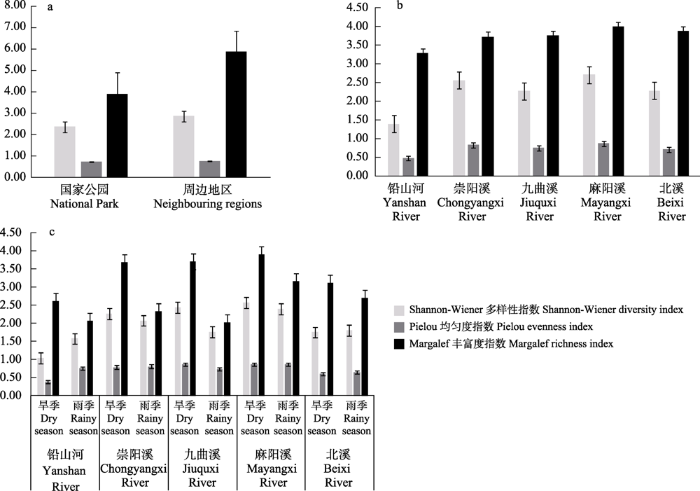
图3
武夷山国家公园及周边地区的鱼类群落多样性指数及其季节差异
Fig. 3
Fish diversity indices and their seasonal differences in Wuyishan National Park and its adjacent areas
2.3.2 鱼类物种多样性在时间尺度上的变化
旱季和雨季分别采集到鱼类43种和30种, 综合所有河流分析, 5条河流的丰富度指数都表现出旱季高于雨季(图3c); 铅山河、北溪旱季的多样性指数和均匀度指数都低于雨季, 崇阳溪、九曲溪、麻阳溪旱季的多样性指数高于雨季。5条河流中物种多样性指数和丰富度指数的最高值均出现在麻阳溪的旱季, 分别为2.57和3.91; 最低值出现在铅山河的旱季(1.04)和九曲溪的雨季(2.02); 均匀度指数最高值出现在九曲溪的旱季(0.86), 最低值为铅山河的雨季(0.38)。所有河流旱季和雨季的均匀度指数(P = 0.245, > 0.05)和丰富度指数(P = 0.655, > 0.05)以及多样性指数差异不显著(P = 0.058, > 0.05), 说明鱼类多样性的分布无明显的季节差别。
2.4 鱼类功能多样性
对功能多样性指数空间格局进行分析(表3), 结果表明武夷山国家公园(0.0300)和周边地区(4.8762)的功能丰富度相差较大, 功能均匀度和功能离散度差异不明显。从河流来看, 功能丰富度最高值出现在北溪(0.1115), 最低值为九曲溪(0.0069); 功能均匀度差异不显著, 调查区域内的5条河流的功能均匀度均在0.5-0.7之间, 其中最高值为崇阳溪(0.6633), 最低值为铅山河(0.5696)。功能离散度最高的为九曲溪(0.7928), 最低为铅山河(0.4912)。
表3 武夷山国家公园鱼类功能多样性在空间尺度上的变化
Table 3
| 河流 River | 功能丰富度 Functional richness (FRic) | 功能均匀度 Functional evenness (FEve) | 功能离散度 Functional divergence (FDiv) |
|---|---|---|---|
| 国家公园 National park | 0.0300 | 0.5383 | 0.8092 |
| 周边地区 Neighbouring regions | 4.8762 | 0.5030 | 0.6688 |
| 铅山河 Yanshan River | 0.0191 | 0.5696 | 0.4912 |
| 崇阳溪 Chongyangxi River | 0.0797 | 0.6633 | 0.6696 |
| 九曲溪 Jiuquxi River | 0.0069 | 0.5734 | 0.7928 |
| 麻阳溪 Mayangxi River | 0.0322 | 0.5760 | 0.7875 |
| 北溪 Beixi River | 0.1115 | 0.5940 | 0.7398 |
对此次调查涉及到水系的鱼类功能多样性指数进行季节动态比较(表4)。结果表明: 所有河流的功能丰富度都明显表现出雨季比旱季高, 最高值出现在北溪的雨季(6.6307), 最低值则出现在铅山河的旱季(0.0004)。铅山河和北溪的功能均匀度表现出雨季高于旱季, 而崇阳溪、麻阳溪、九曲溪则相反。
表4 武夷山国家公园鱼类功能多样性在时间尺度上的变化
Table 4
| 河流 River | 季节 Season | 功能丰富度 Functional richness (FRic) | 功能均匀度 Functional evenness (FEve) | 功能离散度 Functional divergence (FDiv) |
|---|---|---|---|---|
| 铅山河 Yanshan River | 旱季 Dry season | 0.0004 | 0.6182 | 0.5798 |
| 雨季 Rainy season | 0.0054 | 0.7521 | 0.8153 | |
| 崇阳溪 Chongyangxi River | 旱季 Dry season | 0.0078 | 0.7054 | 0.8469 |
| 雨季 Rainy season | 1.1193 | 0.5439 | 0.7350 | |
| 九曲溪 Jiuquxi River | 旱季 Dry season | 0.0022 | 0.6488 | 0.8255 |
| 雨季 Rainy season | 0.6105 | 0.5561 | 0.7485 | |
| 麻阳溪 Mayangxi River | 旱季 Dry season | 0.0094 | 0.6547 | 0.8121 |
| 雨季 Rainy season | 4.7039 | 0.4958 | 0.7498 | |
| 北溪 Beixi River | 旱季 Dry season | 0.0156 | 0.5991 | 0.9068 |
| 雨季 Rainy season | 6.6307 | 0.6087 | 0.6162 |
功能离散度在河流季节动态上则呈现出除了铅山河是雨季高于旱季, 其他的河流都是旱季高于雨季的特征。
3 讨论
3.1 鱼类物种组成
3.1.1 鱼类区系的特点
武夷山国家公园及其周边地区鱼类种类繁多(附录1), 物种数量约占中国淡水鱼类总数的1/10 (张春光和赵亚辉, 2016)。除鮈亚科、鲌亚科、鱊亚科和雅罗鱼亚科等江河平原鱼类, 该区域还分布着鲤科中的鲃亚科、花鳅科中的沙鳅亚科、鮡科、爬鳅科、胡子鲇科、钝头鮠科、斗鱼科、刺鳅科等暖水性鱼类, 虽缺乏鲤科中野鲮亚科的种类, 但仍呈现出较为明显的东洋界鱼类特点。陈宜瑜等(1986)将南岭-武夷山作为东洋界中的华东小区和华南小区的分界线; 唐文乔等(2022)进一步将南岭-武夷山-洞宫山脉作为东洋界的南东亚亚区华南小区与华东小区的分界线, 南坡的闽江属于华南小区, 北坡的信江水系为华东小区。根据附录1统计, 武夷山国家公园北坡信江水系鱼类83种, 南坡闽江水系淡水鱼类99种。相比较而言, 南北坡共有鱼类67种, 占总淡水鱼类种数的59.3%, 南坡闽江水系有不见于北坡信江水系的纵纹原缨口鳅(Vanmanenia caldwelli)、原缨口鳅(V. stenosoma)、裸腹原缨口鳅(V. gymnetrus)、斑纹台鳅(Formosania stigmata)、缨口台鳅(F. lacustre)、台湾白甲鱼、武夷光唇鱼、长鳍犁头鳅(Lepturichthys dolichopterus)等广泛分布于东南沿海、台湾岛、海南岛诸水系以及珠江的鱼类。北坡有不见于南坡的吻鮈(Rhinogobio typus)、短须颌须鮈(Gnathopogon imberbis)、双色小鳔鮈(Microphysogobio bicolor)、张氏薄鳅(Leptobotia tchangi)、长臀疯鲿(Tachysurus analis)、大斑原缨口鳅(Vanmanenia maculate)等长江中下游地区的鱼类。此外闽江水系的江河平原鱼类相较于信江有所下降, 而爬鳅科、鲃亚科、钝头鮠科等东洋区典型类群的比重相对增加。结合现状调查, 崇阳溪、九曲溪、麻阳溪和北溪在鱼类组成上基本相似, 以鲤科鮈亚科、鲃亚科以及爬鳅科鱼类为主。半刺光唇鱼、武夷光唇鱼、台湾白甲鱼、刺鲃(Spinibarbus caldwelli)等鱼类只在闽江水系的崇阳溪、麻阳溪、九曲溪等溪流中采集到, 中华纹胸鮡(Glyptothorax sinensis)则只在北溪溪流中采集到。而铅山河主要是以鮈亚科、爬鳅科、虾虎鱼科等鱼类为主体成分, 相对于另外4条河流, 其独有鱼类为大斑原缨口鳅, 另外鲃亚科鱼类占比下降, 只采集到克氏光唇鱼(Acrossocheilus kreyenbergii), 而钝头鮠科、鮡科等鱼类在此次调查中没有采集到。由此可见武夷山国家公园南北两侧的鱼类区系在物种水平上差异明显, 使得南北坡分别表现出明显的东洋区南亚亚区华南小区、东洋区东亚亚区华东小区鱼类特点。
3.1.2 新记录种
本次调查旱季和雨季采集到鱼类共70种, 只占了历史鱼类种数的59.8%, 这与祝于红等(2017)近年在武夷山进行调查的结果类似, 推测与海拔、地形等因素有关。本次野外调查亦发现一批调查区域的新记录种, 总计有14种, 包括小鳔鮈属中的张氏小鳔鮈、双色小鳔鮈, 鳑鲏属中的方氏鳑鲏(Rhodeus fangi), 鳈属中的江西鳈近缘种(Sarcocheilichthys aff. kiangsiensis), 原鲌属中的达氏原鲌(Chanodichthys dabryi), 花鳅属中的长吻花鳅(Cobitis dolichorhynchus), 原缨口鳅属中的大斑原缨口鳅, 疯鲿属中的细身疯鲿(Tachysurus gracilis)和叉尾疯鲿近似种(T. cf. tenuifurcatus)以及吻虾虎鱼属中的雀斑吻虾虎鱼(Rhinogobius lentiginis)、长汀吻虾虎鱼(R. changtinensis)、武义吻虾虎鱼(R. wuyiensis)、黑吻虾虎鱼(R. niger)、网纹吻虾虎鱼(R. reticulatus), 不但丰富了我们对武夷山国家公园鱼类物种多样性的认识, 也为武夷山国家公园鱼类保护提供了更新的基础性数据。
3.1.3 濒危物种
武夷山国家公园共分布受威胁鱼类3种(附录1), 仅占鱼类总数的2.7%, 较中国整体受威胁淡水鱼类的比例(20.4%)低(曹亮等, 2016)。这些受威胁的鱼类中, 野生花鳗鲡、日本鳗鲡已多年无分布记录, 胭脂鱼在2007-2012年的崇阳溪鱼类资源调查中有一定的种群资源, 但之后包括本研究在内的多次调查均未采集到(李万宝, 2014)。日本鳗鲡在本次调查中虽有记录, 但根据对当地渔政、渔民的走访问询, 在当地有日本鳗鲡养殖场, 关于其是养殖逃逸物种还是野生物种尚有存疑。所以当前在武夷山地区 基本没有自然分布的受威胁鱼类。因此从这个角度来看, 武夷山国家公园覆盖的范围对于受威胁鱼类的保护有限。此外, 数据缺乏和未评估的物种占土著鱼类种数的27.4%。
3.2 α多样性
3.2.1 α多样性的空间变化
此次调查的鱼类α多样性在国家公园及其周边地区空间上表现出差异性(图3a)。国家公园内海拔最高为2,158 m, 最低为300 m, 落差大, 再加上山势较陡, 溪流河面窄, 水体底质多为砾石, 水量充沛, 流速快, 使得武夷山国家公园水生环境生境异质性低(张春英等, 2008)。而水生生境的异质性程度可影响鱼类的物种数和丰富度, 导致异质性程度低则物种数和丰富度也较低(Yan et al, 2011)。因此在国家公园内分布的鱼类主要是一些适应山区溪流的小型鱼类, 优势种如拟腹吸鳅、斑纹台鳅、纵纹原缨口鳅等。周边地区包括崇阳溪的中下游以及铅山河和北溪流域, 随着海拔的降低, 相应的溪流流速降低和生境的异质性增加, 形成缓流、洄水湾、深潭等生境(何美峰等, 2012)。国家公园周边地区在鱼类组成上与国家公园有一定差异性, 除了分布拟腹吸鳅、纵纹原缨口鳅等溪流性鱼类之外, 还分布了一些广适种如银鮈(Squalidus argentatus)、鳗尾䱀(Liobagrus anguillicauda)、中华纹胸鮡等。生境异质性程度低是国家公园鱼类的多样性、丰富度以及均匀度都低于周边地区的主要原因(图3a)。
武夷山国家公园鱼类多样性在水系空间分布上也表现出了一定的波动性(图3b)。麻阳溪的多样性、均匀度以及丰富度在5条河流中最高, 而九曲溪的3个物种多样性指数比大部分点位都位于国家公园周边地区的崇阳溪要低。九曲溪是武夷山国家级重点风景名胜区主要区域, 九曲竹筏漂流对水体搅动较大, 影响了河流河床(张玉等, 2010)。随着游客数量的增加, 九曲溪流域整体受到土地利用开发和旅游业发展的干扰不断增加(张玉等, 2010; 李灵等, 2011)。鱼类作为水生生境的重要组成部分, 其多样性和丰度在一定程度上能反映水体的健康状态(黄亮亮等, 2013)。九曲溪鱼类多样性相较于麻阳溪受人为活动的影响程度较大, 但相较于国家公园周边地区的北溪和铅山河, 其鱼类多样性和丰富度又处于较高的水平。铅山河的多样性、均匀度和其他4条河流相差较大, 原因可能为受到水资源利用的影响, 同时铅山河流域内有开采的铜矿厂, 洗矿废水的排入造成重金属污染(江西省上饶市水利电力勘测设计院, 2010②(② 江西省上饶市水利电力勘测设计院 (2010) 铅山河修编报告.); 张凯强, 2011③(③ 张凯强 (2011) 铅山河生态基流确定及生态调度模型初步研究. 硕士学位论文, 南昌大学, 南昌.))。
3.2.2 α多样性的季节差异
旱季和雨季的鱼类多样性呈现一定的季节动态变化(图3c)。5条河流的多样性指数和丰富度指数普遍表现出旱季高于雨季, 鱼类的多样性受到鱼类生活史的影响, 绝大多数鱼类的繁殖期集中在春末, 如只在旱季采集到的黑鳍鳈(Sarcocheilichthys nigripinnis)、小鳈(S. parvus)、克氏光唇鱼等的繁殖期集中在4月左右(福建鱼类志编写组, 1984), 且处于繁殖期的高体鳑鲏成为旱季优势种, 渔获量占了旱季总数的17.8% (表2)。这时有大量仔稚鱼群体的补入, 可维持鱼类的多样性和丰度, 可能是导致旱季鱼类多样性较高的原因(叶昆等, 2021)。均匀度指数在雨季高于旱季, 原因可能为雨季采集到的渔获物(707尾)和旱季(749尾)相差不大, 而雨季的物种数(30种)明显少于旱季(43种), 从而导致雨季的均匀度高于旱季(图3c)。
3.3 功能多样性
3.3.1 功能多样性的空间变化
功能多样性是将鱼类行使繁殖、摄食、运动等方面的功能器官量化, 以此来反映群落内物种间功能特征的整体差别或多样性(Petchey & Gaston, 2006)。功能丰富度可反映鱼类在群落中占据的生态位, 同时可评价群落生产力、对环境波动的缓冲能力等, 功能丰富度降低, 表明群落内的生产力和对外界干扰因素的抵抗力降低(帅方敏等, 2017)。另外稀有物种独特的功能特征值也会使功能丰富度的数值极大膨胀, 这种独特的功能特征值的消失会导致功能多样性的降低, 并导致功能同质化(Laliberté & Legendre, 2010; Chua et al, 2019)。国家公园周边地区、崇阳溪和北溪这三个区域的生境异质性高, 在鱼类口位和食性功能性状组成上, 分布着口下位的藻食性鱼类, 例如斑纹台鳅; 同时还分布着口端位的杂食性鱼类, 例如高体鳑鲏; 还分布着口裂大的肉食性鱼类, 例如鲇(Silurus asotus)、黄颡鱼(Tachysurus sinensis)、盎堂疯鲿(T. ondon)等。这些鱼类的分布使得功能性状多样化, 特有的功能特征值使得功能丰富度增高(Chua et al, 2019), 因此国家公园周边地区、崇阳溪和北溪的功能丰富度呈现较高水平(表3)。而全部河段位于国家公园内的九曲溪和上游河段位于国家公园内的麻阳溪在栖息生境方面单一, 且分布的鱼类优势种主要是以口下位杂食性(偏植食性)鱼类为主, 如拟腹吸鳅、台湾白甲鱼、武夷光唇鱼等(图2b)。功能丰富度数值的变化反映出国家公园鱼类栖息环境单一, 导致鱼类群落的生产力低, 不足以为鱼类提供多样化的栖息环境。
功能均匀度相对较高的栖息地更稳定, 对群落中的资源利用也更充分(Massicotte et al, 2014; 帅方敏等, 2017)。淡水鱼类的群落结构和功能多样性模式与栖息地类型、栖息水层、人为活动、鱼类食性组成等非生物或生物因素相关(Comte et al, 2016)。不同类型的栖息地为鱼类提供不同的食物资源, 也丰富了鱼类对资源的利用方式(Mason et al, 2005)。5条河流的功能均匀度差异不明显(表3), 除了崇阳溪处于较高水平, 其他4条河流基本处于相同水平。崇阳溪中上层、中下层鱼类分别占其所有鱼类的9.1%和31.8% (图2a), 相较于其他河流不同水层的鱼类物种数均衡。另外, 植食性、杂食性、肉食性的鱼类物种数均衡性也高于其他河流(图2b)。国家公园内的功能均匀度也要高于周边地区, 推测是因为其周边地区是当地居民生产生活区域, 对土地利用程度较大, 加大了对国家公园周边地区的人为干扰, 另外土地覆盖率的变化也会使得河流的功能均匀度降低(Barbosa et al, 2020)。
功能离散度的变化可反映群落中物种的竞争状态, 功能离散度小, 表明物种间竞争激烈, 生态位空间相对紧张(Villéger et al, 2008; 帅方敏等, 2017)。从研究结果来看, 国家公园内的功能离散度高于周边地区, 另外九曲溪、麻阳溪上游河段在调查点位上和国家公园有重复, 因此功能离散度基本处于同一水平, 相差不大(表3)。但却比在国家公园周边地区的北溪、崇阳溪、铅山河高, 这三条河流在不同程度上受到水质的污染和人为活动的干扰, 使得鱼类所需要的食物来源和生存空间被压缩, 物种间的竞争加剧(兰永清, 2003; 张凯强, 2011④(④ 张凯强 (2011) 铅山河生态基流确定及生态调度模型初步研究. 硕士学位论文, 南昌大学, 南昌.); 陈莎等, 2021)。
综合以上分析结果可知, 影响武夷山国家公园功能多样性的因素主要是生境异质性, 其次是人为活动带来的干扰, 例如土地的利用、水质的污染等。全部河段处于国家公园内的九曲溪以及上游河段处于国家公园内的麻阳溪主要受到生境异质性的限制, 功能多样性较低, 但国家公园范围的划定也为这些区域提供了保护屏障, 受到的人为活动干扰较少, 其群落之间的物种竞争相对周边地区的北溪、崇阳溪、铅山河要少。
3.3.2 功能多样性的季节变化
功能多样性在季节上呈现出一定的变化规律(表4)。各河流雨季的功能丰富度均高于旱季, 表明旱季的鱼类在功能性状组成上更相似(Chua et al, 2019)。从食性方面看, 旱季杂食性(偏植食)的鱼类数量占比较高, 而雨季肉食性鱼类的种类和数量均有所增加; 从运动方面看, 雨季运动能力强的鱼类成为优势种(表2), 优势种体型较大, 可进行长距离的游泳、摄食等行为, 使得群落中的资源得到更加有效的利用(Villéger et al, 2008; 张晓妆等, 2019)。植食性、杂食性、肉食性等不同食性的鱼类数量和占比影响着鱼类对群落中食物的利用状况, 另外, 水资源的下降也会导致群落中资源的利用和生存空间的竞争加剧(张金屯和范丽宏, 2011; 张倩等, 2022)。武夷山国家公园属于亚热带季风气候, 5-6月为丰水期, 丰水期过后, 饵料和水位的下降使得雨季(7月)的鱼类生存空间压缩(张玉等, 2010)。这可能是功能均匀度和功能离散度旱季普遍高于雨季的原因。
3.4 鱼类多样性保护的对策
为修复武夷山国家公园及其周边水域的水生生境, 提高鱼类多样性, 恢复鱼类资源, 提出如下保护建议:
(1)生态修复, 增加栖息环境多样化。
根据调查研究结果, 鱼类的物种丰富度和多样性沿着河流的海拔呈纵向梯度降低, 可见武夷山国家公园及其周边地区的鱼类空间分布特征是由自然生境和人为活动干扰所致。从水系上看, 国家公园内外水体不宜相互割裂, 相应的保护设计应该进行统筹考虑。在这样的背景下, 加大维护武夷山国家公园特别是国家公园毗邻区水体自然生境的力度, 减少人为活动的干扰, 对公园外重要水域进行必要的生态修复, 有利于提高鱼类栖息生境的多样化。另外, 可根据河道的水文情况和底质对受污染较大的周边地区河流进行水质生态修复。譬如: 可基于铅山河水域内水生动植物的情况, 选择适应能力强且有净化水质功能的水生植物, 再辅以水生动物, 以此构建“水生植物-微生物-藻类-水生动物”食物链, 来达到水质净化的目的, 尽可能提升水生生态系统的完整性(刘青等, 2020)。
恢复河流连通性是恢复鱼类栖息生境的方法之一。水利设施的建设改变了河流的连通性, 河段从流水生境转为静水生境, 如宸前水电站建成后, 从宸前至白茶埠的11 km从流水河段转为静水河段, 导致原有的鱼类产卵场消失, 改变了区域内的鱼类群落结构(李万宝, 2014)。调查区域内有中型以上水库11座(张天明和黄常斌, 2002), 根据当地水利局工作人员介绍, 仅在麻阳溪就有20多座水电站。水利设施的建立还会影响鱼类生活史, 例如每年的6-9月是幼鳗溯河期, 很多幼鳗被阻拦于河坝前而被大量捕捞, 无法完成生活史, 导致现在中上游很难再看见日本鳗鲡、花鳗鲡等珍稀濒危鱼类。可结合鱼类的生物学特性和生态习性, 适当拆除效率低下的水电站和修建过鱼通道设施, 来恢复水利设施上下游的鱼类种群交流。过鱼通道在地势地形允许的情况下, 仿自然的旁路式鱼道应是首选。
(2)科学且有计划地开展增殖放流。
合理的增殖放流是补充鱼类种群资源和数量的重要手段, 可以改善因人为活动干扰带来的水生生态破坏, 延续珍稀濒危鱼类种群, 保持鱼类多样性(杨君兴等, 2013)。针对武夷山国家公园及其周边水域的鱼类多样性现状, 在增加对经济鱼类放流的同时, 也应重视对花鳗鲡、胭脂鱼等珍稀濒危鱼类的种群恢复。鼓励政企合作, 攻克珍稀濒危或重要土著鱼类人工繁育难关, 培育当地土著鱼类为亲鱼, 作为增殖放流苗种来源。在增殖放流过程中还应针对水域的不同特性选择不同种类和数量, 严防外来物种入侵。当地渔政相关部门应借鉴相关养殖鱼类培育经验, 在已有的人工培育物种基础上加强人工繁殖技术发展, 以保护珍稀濒危鱼类、发展当地经济渔业为向导, 加强珍稀濒危鱼类(花鳗鲡、胭脂鱼)以及其他土著鱼类种质资源保护和培育, 积极开展珍稀濒危鱼类的人工繁育工作和土著鱼类的增殖放流工作。同时还应做好增殖放流效果评价, 监测放流种群的存活状况以及放流工作对野外种群恢复的效果。
(3) 扩大水域保护范围。
国家公园的设立为水生生态系统建立了保护屏障, 缓解了鱼类资源衰退, 减少了人类活动对鱼类及其生境的影响。国家公园覆盖的范围应该涵盖鱼类资源丰富的水域, 崇阳溪的鱼类多样性仅次于麻阳溪, 因此为了保护武夷山地区鱼类多样性, 可考虑适当扩大禁渔区范围, 将崇阳溪水域划入国家公园实施禁止捕捞野生鱼类的范畴, 增大鱼类保护力度。在调查中, 崇阳溪附近市场发现大量捕捞售卖河流野生鱼类的情况。从1960年代开始, 处于武夷山国家公园周边地区的崇阳溪和北溪的渔获物在闽江水系中占一定的比重, 长期的过度捕捞使得水体中大型的经济鱼类数量减少, 开始以捕捞小型鱼类和低龄鱼类为主(何美峰等, 2012; 李万宝, 2014)。长江“十年禁渔”政策的实施为铅山河鱼类多样性的保护提供了保障。建议在国家公园周边地区的河流(崇阳溪、北溪)制订合理的休渔禁渔制度, 控制渔业作业时间和规模。禁渔制度可协调多个县市共同推广, 禁渔期的时间应涵盖洄游性鱼类的洄游季节。
(4)合理调度旅游安排。
九曲溪是武夷山风景名胜区的主要区域, 游客数量的增加加大了对水域生态的干扰, 不利于水生生态系统的自身恢复。武夷山游客数量在每年10月和11月(枯水期)到达高峰值, 其次为4月(平水期); 在旅游旺季, 行筏人数达到全年的23.35% (张玉等, 2010)。针对武夷山旅游季游客的数量, 应考虑河流水位和水体受干扰程度, 结合河流的平水期(4月)、枯水期(10-12月)、丰水期(5-6月)和鱼类的产卵期(4-5月)合理协调游客数量。建议在旅游旺季限制游客数量, 规定游玩时间, 合理安排旅游高峰期和休渔期错峰。
附录 Supplementary Material
附录1 武夷山国家公园及其周边地区淡水鱼类物种名录
Appendix 1 List of freshwater fish species in the Wuyishan National Park and its adjacent areas
参考文献
Influence of land-use classes on the functional structure of fish communities in southern Brazilian headwater streams
DOI:10.1007/s00267-020-01274-9
PMID:32130463
[本文引用: 1]

Changes in landscape composition caused by conversion of natural habitats into human-altered ecosystems can directly influence the physical characteristics of stream networks. Such impacts can modify the functional structure of fish communities, although the exact consequences of anthropic land-use changes can be context-dependent. This study investigated the influence of different land-use classes on the functional structure of fish communities in 32 headwater streams from southern Brazil. Trait composition and indices of functional diversity of the fish community were related to four land-use classes: native forest vegetation, silviculture, agriculture, and urban areas. Streams surrounded by larger areas of native forest were characterized by the predominance of foraging specialist species like grazers. However, as native vegetation is replaced by agriculture and urban areas, specialist species are replaced by species with generalist diet like detritivores. In streams surrounded by larger areas of agriculture, functional richness and divergence increased, while functional evenness decreased. Most likely, these changes were induced by alterations in the water quality, indicated by increased electrical conductivity and water temperature in streams with more agriculture areas. In conclusion, the conservation of the native forest vegetation is essential to maintain habitat characteristics and ecological processes in streams and to avoid the loss of specialist species in fish communities.
Fishing effects on diversity, size and community structure of the benthic invertebrate and fish megafauna on the bay of Biscay coast of France
Comparison of feeding guild structure and ecomorphology of intertidal fish assemblages from central California and central Chile
Evaluating the status of China’s continental fish and analyzing their causes of endangerment through the red list assessment
通过红色名录评估研究中国内陆鱼类受威胁现状及其成因
DOI:10.17520/biods.2015331
[本文引用: 1]
根据已有的基础资料, 采用IUCN评估等级和标准, 对中国目前已鉴定的1,443种内陆鱼类受威胁现状进行了评估。评估结果显示, 1,443种内陆鱼类中, 灭绝3种、区域灭绝1种、极危65种、濒危101种、易危129种、近危101种、无危454种和数据缺乏589种。同已有的IUCN评估结果相比, 本次被评估的物种数目多, 受威胁物种大幅度增加, 其数目达295种, 占已知中国内陆鱼类总数的20.44%, 低于全球平均值(29%)。属于灭绝等级的鱼类是大鳞白鱼(Anabarilius macrolepis)、异龙鲤(Cyprinus yilongensis)和茶卡高原鳅(Triplophysa cakaensis); 属于区域灭绝等级的鱼类是长颌北鲑(Stenodus nelma)。鲤科是受威胁物种数最多的科, 其中裂腹鱼亚科和鲤亚科的种类受威胁程度最高。长江上游和珠江上游受威胁物种最多, 是受威胁最严重的地区。中国内陆鱼类受威胁的主要因素为河流筑坝、生境退化或丧失、酷渔滥捕和引进外来种。列入数据缺乏等级的鱼类较多, 占中国内陆鱼类的40.82%, 表明对中国内陆鱼类物种多样性了解不充分, 需要加强野外调查以积累基础资料。
Biodiversity in the Wuyi Mountains and its importance in China
论武夷山在中国生物多样性保护中的地位
Structural characteristics of zooplankton and phytoplankton communities and its relationship with environmental factors in a typical tributary reservoir in the Three Gorges Reservoir Region
三峡库区典型支流水库浮游动植物群落结构特征及其与环境因子的关系
Ichthyofauna of the Pearl River with a discussion on zoogeographical divisions for freshwater fishes
珠江的鱼类区系及其动物地理区划的讨论
Loss of endemic fish species drives impacts on functional richness, redundancy and vulnerability in freshwater ecoregions of Sundaland
Resource partitioning and functional diversity of worldwide freshwater fish communities
Fish fauna characteristics of Jianxi River and Futunxi River and suggestions on resource protection and utilization
建溪、富屯溪鱼类区系特征及其资源保护利用意见
Community organization in fishes as indicated by morphological features
Study on fish composition and diversity in Chongyang River in Fujian Province
闽江崇阳溪鱼类组成及多样性研究
Diversity and evenness: A unifying notation and its consequences
Development and application of IBI based on fish to assess the river’s health in the East Tiaoxi River
东苕溪鱼类生物完整性评价河流健康体系的构建与应用
Analysis of ecological environment changes and driving forces in Wuyishan National Park
武夷山国家公园生态环境变化及其驱动力分析
The relationships between mouth size and shape and body length for 18 species of marine fishes and their trophic implications
A distance-based framework for measuring functional diversity from multiple traits
A new framework for measuring functional diversity (FD) from multiple traits has recently been proposed. This framework was mostly limited to quantitative traits without missing values and to situations in which there are more species than traits, although the authors had suggested a way to extend their framework to other trait types. The main purpose of this note is to further develop this suggestion. We describe a highly flexible distance-based framework to measure different facets of FD in multidimensional trait space from any distance or dissimilarity measure, any number of traits, and from different trait types (i.e., quantitative, semi-quantitative, and qualitative). This new approach allows for missing trait values and the weighting of individual traits. We also present a new multidimensional FD index, called functional dispersion (FDis), which is closely related to Rao's quadratic entropy. FDis is the multivariate analogue of the weighted mean absolute deviation (MAD), in which the weights are species relative abundances. For unweighted presence-absence data, FDis can be used for a formal statistical test of differences in FD. We provide the "FD" R language package to easily implement our distance-based FD framework.
Present situation and protection countermeasures of fishery resources in Futunxi River
富屯溪渔业资源现状及保护对策
Influence of different land use to soil quality in Jiuquxi ecological protection area
九曲溪生态保护区不同土地利用方式对土壤质量的影响
Investigation and protection on fish resource of Chongyang River in Jianyang City
建阳市崇阳溪鱼类资源现状调查与保护对策
Species diversity of freshwater fish and assessment on watershed health in the Irtysh River and Ulungur River basins in Xinjiang, China
DOI:10.17520/biods.2019071
[本文引用: 1]
We conducted yearly field surveys between 2013 and 2016 in Altay Prefecture, Xinjiang Uygur Autonomous Region to understand the current status of fish species diversity and its change in the Irtysh River and the Ulungu River basins. The Margalef richness index, the Shannon-Wiener diversity index, and the Pielou evenness index were applied to assess the species diversity and temporal and spatial variation of freshwater fishes, based on the data collected from our field investigations, museum collection, and research literature. There were 23 native fish species distributed in the Irtysh River and the Ulungu River basins. And 19 of them were collected with 15 exotic species during our surveys. The fish fauna is dominated by cyprinid fishes. High proportions of endemic and endangered fish are distinct characteristics of fish composition in Altay Prefecture. Our results showed that the fish diversity in the basins was generally stable between 2013 and 2016. Species richness in the Irtysh River was higher than one in the Ulungu River. Additionally, the Fish Index of Biological Integrity (F-IBI) of 34 sampling sites indicated that the river health of most sites in the Irtysh River basin was at the “sub-health” or “general” level, while that of most ones in the Ulungu River basin was “healthy”. Water infrastructures, exotic species, and overexploitation of fish resources were key factors to influence freshwater fishes in the region. Aiming at the protection of fish biodiversity and improvement of river healthy, several ways should be applied in future including ecological scheduling of multi-scale coupling of reservoir group, construction of fish pass, scientific management of aquaculture to control the exotic species, artificial enhancement and releasing of endemic and endangered fish, and in situ conservation.
新疆阿勒泰地区额尔齐斯河和乌伦古河流域鱼类多样性演变和流域健康评价
DOI:10.17520/biods.2019071
[本文引用: 1]
为了解阿勒泰地区额尔齐斯河和乌伦古河流域的鱼类多样性现状和历史演变, 本研究于2013-2016年间在该流域的鱼类多样性进行了连续调查, 并结合历史资料和标本, 以Margalef丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数分析评估了流域内鱼类的多样性水平和时空变化。该流域历史上分布有土著鱼类23种, 当前记录到19种, 流域内还有外来鱼类15种。阿勒泰鱼类的区系组成以鲤科种类为主, 其中特有和珍稀濒危物种占比高, 具有重要的保护价值。多样性指数计算结果显示, 2013-2016年鱼类多样性情况整体稳定, 额尔齐斯河鱼类物种数多于乌伦古河。研究还基于鱼类生物完整性指数(Fish Index of Biological Integrity, F-IBI)对34个采集点的河流健康状况进行了评价, 结果显示额尔齐斯河流域大多数调查点的健康状况处于“亚健康”或“一般”水平, 乌伦古河流域多数调查点的健康状况处于“健康”水平。水利工程、外来物种、过度捕捞是影响阿勒泰地区鱼类多样性的重要因素。未来应通过水利工程的联合调度、下泄合理生态流量、布设鱼类通道、规范养殖渔业、严控外来物种、本地土著鱼类的人工增殖放流, 以及合理的就地保护措施对阿勒泰地区的鱼类多样性加以保护, 提升水体健康程度。
Advances in theory and technology of ecological restoration of mountain rivers
山区河流生态修复理论与技术研究进展
How does fish functional diversity respond to environmental changes in two large shallow lakes?
Functional richness, functional evenness and functional divergence: The primary components of functional diversity
Riverscape heterogeneity explains spatial variation in zooplankton functional evenness and biomass in a large river ecosystem
Ecological methodology
Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment
Functional diversity (FD), species richness and community composition
Functional diversity: Back to basics and looking forward
DOI:10.1111/j.1461-0248.2006.00924.x
PMID:16706917
[本文引用: 2]
Functional diversity is a component of biodiversity that generally concerns the range of things that organisms do in communities and ecosystems. Here, we review how functional diversity can explain and predict the impact of organisms on ecosystems and thereby provide a mechanistic link between the two. Critical points in developing predictive measures of functional diversity are the choice of functional traits with which organisms are distinguished, how the diversity of that trait information is summarized into a measure of functional diversity, and that the measures of functional diversity are validated through quantitative analyses and experimental tests. There is a vast amount of trait information available for plant species and a substantial amount for animals. Choosing which traits to include in a particular measure of functional diversity will depend on the specific aims of a particular study. Quantitative methods for choosing traits and for assigning weighting to traits are being developed, but need much more work before we can be confident about trait choice. The number of ways of measuring functional diversity is growing rapidly. We divide them into four main groups. The first, the number of functional groups or types, has significant problems and researchers are more frequently using measures that do not require species to be grouped. Of these, some measure diversity by summarizing distances between species in trait space, some by estimating the size of the dendrogram required to describe the difference, and some include information about species' abundances. We show some new and important differences between these, as well as what they indicate about the responses of assemblages to loss of individuals. There is good experimental and analytical evidence that functional diversity can provide a link between organisms and ecosystems but greater validation of measures is required. We suggest that non-significant results have a range of alternate explanations that do not necessarily contradict positive effects of functional diversity. Finally, we suggest areas for development of techniques used to measure functional diversity, highlight some exciting questions that are being addressed using ideas about functional diversity, and suggest some directions for novel research.
Functional diversity of freshwater fishes and methods of measurement
淡水鱼类功能多样性及其研究方法
Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics
Freshwater fish fauna and zoogeographical divisions in the Wuyi-Xianxialing mountains of eastern China
DOI:10.17520/biods.2017207
[本文引用: 1]
The watershed of the Wuyi-Xianxialing Mountains comprises the Poyang Lake water system of the Yangtze River basin and independent coastal rivers of eastern China, including the Qiantang River, Oujiang River, Minjiang River, and Hanjiang River. It is a key area for the conservation of biodiversity in Chinese ecosystems. In this study, fish surveys were conducted in 111 upstream sections of five river basins draining the Wuyi-Xianxialing Mountains and over 13,000 specimens were collected. The fish fauna of this area, on the basis of our investigations and records from historical literature, included 282 species, of which 239 were freshwater fishes, belonging to 101 genera, 23 families, and 6 orders. The orders of Cypriniformes, Perciformes, and Siluriformes supported the majority of fish composition in this area, with 181, 29, and 22 species, respectively, accounting for 75.73%, 12.13%, and 9.21% of total freshwater fish found. There were 109 fish species that were the major component of fish composition of the river plains of East Asia and 51 species of warm water fish with Southeast Asian origin, accounting for 45.61% and 21.34% of the total freshwater fishes, respectively. There were many species shared among the five river basins, and the average faunal resemblance of any two rivers was more than 50%. For the four substantive rivers in the eastern region, from north to south, the proportion of endemic fishes in the river plains of East Asia gradually decreased, and the proportion of warm water fishes of Southeast Asian origin gradually increased. Clustering analysis of fish fauna demonstrates that Poyang Lake, Qiantang River, Oujiang River, and Minjiang River are clustered into one class, while Hanjiang River forms the other class. This analysis indicates that the Wuyi-Xianxialing Mountains do not form a strong barrier for freshwater fishes, and that the whole district of the Wuyi-Xianxialing Mountains should be a component of the Oriental Region. In view of the differences between the north and the south, this paper argues that Hanjiang River in the southeast belongs to the South China Area of the South-east Asiatic Subregion, while Poyang Lake, Qiantang River, Oujiang River, and Minjiang River belong to the East China Area.
华东武夷山-仙霞岭地区淡水鱼类区系特征及其动物地理区划
DOI:10.17520/biods.2017207
[本文引用: 1]
武夷山脉-仙霞岭是长江鄱阳湖水系与我国东部独流入海的钱塘江、瓯江、闽江和韩江等水系的分水岭, 是中国陆地生物多样性保护的关键地区之一。本文对武夷山-仙霞岭地区5个水系的111个上游江段鱼类进行了采集, 获得标本13,000余号, 并结合文献资料对该地区的鱼类区系作了分析。结果显示, 该地区共分布有鱼类282种, 其中纯淡水土著鱼类239种, 隶属于6目23科101属。鲤形目、鲈形目、鲇形目各有181种、29种和22种, 分别占纯淡水鱼类的75.73%、12.13%和9.21%, 是本地区鱼类组成的主体。东亚特有的江河平原鱼类和南亚暖水性鱼类各有109种和51种, 分别占纯淡水鱼类的45.61%和21.34%。5个水系之间鱼类的共有种较多, 平均相似度均在50%以上。但东部4条独流入海河流, 自北往南江河平原鱼类所占比例逐渐下降, 而南亚暖水性鱼类则逐渐增加。鱼类区系的聚类分析显示, 鄱阳湖、钱塘江、瓯江和闽江聚为一类, 韩江单独构成另一类。分析表明, 武夷山脉-仙霞岭并没有对淡水鱼类形成很强的阻隔作用, 整个区域都是东洋区的组成部分。但鉴于已经出现的一些南北差异, 本文认为可以将东南部的韩江水系划归南东亚亚区的华南小区, 鄱阳湖水系与钱塘江、瓯江和闽江水系划归华东小区。
Species diversity and zoogeographical analysis of freshwater fishes in central and southern mountainous area of East China
华东中南部山区淡水鱼类物种多样性及动物地理区划
Diversity and productivity in a long-term grassland experiment
Plant diversity and niche complementarity had progressively stronger effects on ecosystem functioning during a 7-year experiment, with 16-species plots attaining 2.7 times greater biomass than monocultures. Diversity effects were neither transients nor explained solely by a few productive or unviable species. Rather, many higher-diversity plots outperformed the best monoculture. These results help resolve debate over biodiversity and ecosystem functioning, show effects at higher than expected diversity levels, and demonstrate, for these ecosystems, that even the best-chosen monocultures cannot achieve greater productivity or carbon stores than higher-diversity sites.
New multidimensional functional diversity indices for a multifaceted framework in functional ecology
DOI:10.1890/07-1206.1
PMID:18724739
[本文引用: 4]
Functional diversity is increasingly identified as an important driver of ecosystem functioning. Various indices have been proposed to measure the functional diversity of a community, but there is still no consensus on which are most suitable. Indeed, none of the existing indices meets all the criteria required for general use. The main criteria are that they must be designed to deal with several traits, take into account abundances, and measure all the facets of functional diversity. Here we propose three indices to quantify each facet of functional diversity for a community with species distributed in a multidimensional functional space: functional richness (volume of the functional space occupied by the community), functional evenness (regularity of the distribution of abundance in this volume), and functional divergence (divergence in the distribution of abundance in this volume). Functional richness is estimated using the existing convex hull volume index. The new functional evenness index is based on the minimum spanning tree which links all the species in the multidimensional functional space. Then this new index quantifies the regularity with which species abundances are distributed along the spanning tree. Functional divergence is measured using a novel index which quantifies how species diverge in their distances (weighted by their abundance) from the center of gravity in the functional space. We show that none of the indices meets all the criteria required for a functional diversity index, but instead we show that the set of three complementary indices meets these criteria. Through simulations of artificial data sets, we demonstrate that functional divergence and functional evenness are independent of species richness and that the three functional diversity indices are independent of each other. Overall, our study suggests that decomposition of functional diversity into its three primary components provides a meaningful framework for its quantification and for the classification of existing functional diversity indices. This decomposition has the potential to shed light on the role of biodiversity on ecosystem functioning and on the influence of biotic and abiotic filters on the structure of species communities. Finally, we propose a general framework for applying these three functional diversity indices.
Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation
Ecomorphological analysis of fish taxocenes in rainforest streams of northern Borneo
Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed, the Qingyi Stream, China
Overview of the artificial enhancement and release of endemic freshwater fish in China
中国淡水鱼类人工增殖放流现状
Fish community characteristics in the confluence area of Wanhe estuary in Anqing Section of the Yangtze River
长江安庆段皖河口汇流区鱼类群落特征
Practice and exploration of forest resources conservation in Wuyishan National Park
武夷山国家公园森林资源保护实践与探索
Fractal features of river system landscape in the world natural reserve
武夷山自然保护区水系景观分形特征
Development of species functional diversity and its measurement methods
物种功能多样性及其研究方法
Functional diversities of fish community in Peng’an Section of the middle reach of Jialing River
嘉陵江中游蓬安段鱼类群落功能多样性研究
Function analysis and flood control countermeasures of flood control reservoirs in Jianxi River and Futunxi River basin in the upper reaches of Minjiang River
闽江上游建溪、富屯溪流域防洪水库作用分析及防洪对策
海州湾鱼类群落功能多样性的时空变化
DOI:10.13287/j.1001-9332.201909.035
[本文引用: 1]
功能多样性是联系物种、生态环境和生态系统功能的基础.从功能多样性的角度研究群落结构,有助于更好地理解生物多样性与生态系统功能之间的关系.本研究根据2011—2017年(除2012年外)春、秋季海州湾渔业资源底拖网调查数据,选取反映鱼类摄食、运动、生态适应性、繁殖行为和种群动态特征的13个功能性状,利用功能丰富度指数、功能均匀度指数、功能离散度指数和群落特征加权平均数指数(CWM),研究了海州湾鱼类群落功能多样性的季节、年际和空间的变化.结果表明: 海州湾鱼类群落功能多样性指数具有显著的季节差异,其中秋季功能丰富度指数显著高于春季,春季功能离散度指数显著高于秋季,鱼类洄游是造成功能多样性指数季节变化的主要原因;CWM分析表明,春季鱼类群落优势种为营养级较高、运动能力较强、生长系数、恢复力和脆弱性较高的冷/暖温性鱼类,秋季则相反,春、秋季鱼类群落均以产浮性卵鱼类为主;鱼类群落功能多样性指数呈现一定的年际变化,其中春、秋季各功能多样性指数在不同年际均呈波动趋势,表明海州湾鱼类群落稳定性较低;鱼类群落功能多样性指数呈现显著的空间差异,其中20 m以深远岸水域功能离散度指数较高.海州湾鱼类群落功能多样性具有明显的时空变化特征,鱼类生态位与其对资源的利用均随季节、年际和空间而变化.
武夷山风景名胜区水资源变化特征及可持续规划
Standard of Morphological Measurements and Description for Cyprinidae, Based on an Example of Gobionine Fishes (Teleostei: Cypriniformes)
鲤科鱼类形态学测度和物种描述规范——以鮈亚科鱼类为例

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}