



20世纪伟大的博物学家和生物地理学家爱德华·奥斯本·威尔逊(Edward Osborne Wilson, 1929-2021)与杰出的理论和数学生态学家罗伯特·赫尔默·麦克阿瑟(Robert Helmer MacArthur, 1930-1972)共同开创的岛屿生物地理学理论是他们青年时代的巅峰之作。但是, 由于威尔逊后来在昆虫学和社会生物学等领域的巨大成就, 加之麦克阿瑟的英年早逝, 在很多介绍威尔逊学术成就的出版物中, 往往没有详细交代他们在岛屿生物地理学理论上的合作佳话。在威尔逊刚辞世之际, 回顾他在岛屿生物地理学领域的宝贵科学遗产, 评述其对推动科学革命的贡献, 一方面为了缅怀他勇敢探索、富于创新和谦逊合作的优秀个人品质, 另一方面也启迪我们要拥有勇于挑战学术高峰的科学精神。
岛屿生物地理学理论最早在Evolution期刊发表(MacArthur & Wilson, 1963), 随后又以专论形式系统阐述(MacArthur & Wilson, 1967)。他们的最终目标是发展一个基于迁入、局部灭绝、物种形成等基本生态和进化过程的动态生物地理学的一般理论, 用来解释各种各样的孤岛现象, 包括物种丰富度模式、物种拓殖定居能力和生态位转换, 以及孤立生物类群的进化。多年来, 尽管受到一些批评, 岛屿生物地理学理论为理解生物多样性的形成和维持机制奠定了可预测的理论基础, 更开辟了20世纪中期科学革命结构转变的新范式(从静态描述到动态定量) (Kuhn, 1996), 从而为生物地理学、生态学和进化生物学等提供了真正具有变革性的见解。
以下, 我们简要回顾威尔逊的物种进化知识启蒙教育、南太平洋岛屿考察、生物类群循环理论(theory of taxon cycles)、岛屿生物地理学理论及其控制实验验证, 最后, 简单总结岛屿生物地理学理论的科学价值。
1 物种扩散和进化知识的启蒙
威尔逊成为一名科学家的动力源于对自然界中生物(包括其毕生研究的蚂蚁类群)不可抗拒的热情。他一开始对鸟类感兴趣, 后来右眼在钓鱼时不幸误伤失明, 无法远视, 就转向研究昆虫。正如达尔文的自然选择理论受益于亚历山大·冯·洪堡(Alexander von Humboldt)、詹姆斯·赫顿(James Hutton)、查尔斯·莱伊尔(Charles Lyell)、托马斯·罗伯特·马尔萨斯(Thomas Robert Malthus)等那个时代其他有远见的科学家一样, 威尔逊攻读研究生时(1950年)就对古生物学家威廉·迪勒·马修(William Diller Matthew)提出的北温带是新生代物种起源中心(Matthew, 1915), 以及后来菲利普·达林顿(Philip Darlington)将物种起源中心确定为旧世界热带地区(Darlington, 1948)的学术观点产生浓厚兴趣。据威尔逊回忆, 马修和达林顿这史诗般的全球疆域的生物地理学观点一直刻在他的脑海中, 极大地启发了他的科学想象力(Wilson, 1994)。
与大多数博物学家类似, 威尔逊也选择新世界热带地区开始自己的职业生涯。23岁时(1953年), 哈佛大学的研究生奖学金资助了他在古巴和墨西哥的科学考察。他最具影响力的导师之一达林顿是著名的昆虫学家和动物地理学家, 同时也是他在哈佛大学的同事。当他准备第一次热带采集之旅时, 达林顿为这位前途无量的研究生提供了极具帮助的建议。
2 结缘南太平洋岛屿
1954年3月, 达林顿向威尔逊提供了一个千载难逢的机会——去南太平洋的新几内亚群岛采集蚂蚁标本, 他毫不犹豫地接受了这份礼物。正如威尔逊在自传中所述(Wilson, 1994): “一百年前, 年轻的华莱士在那里开始动物分布研究, 并将动物地理学转化为一门科学学科, 而我也正在这个领域开始工作。谁知道这段经历会如何改变我成为一个动物地理学家的想法?”
1954年11月24日, 威尔逊离开波士顿开始了10个月的南太平洋远行。12月, 先在斐济的维提拉武岛采集到了几种当地特有的蚂蚁种类。短暂停留后, 前往美拉尼西亚群岛最南端的新喀里多尼亚, 这里的动植物是在近数百万年间陆续由澳洲往东或是由所罗门群岛经由新赫布里底群岛(New Hbrides)往南迁移而来, 彼此交流, 进化出了独特的生态谱系。在努美亚附近, 他采集的粗角蚁属(Cerapachys)和束蚁属(Sphinctomyrmex)是人类首次采集到的蚂蚁新物种。他发现, 粗角蚁属工蚁为了征服猎物, 以类似热带大陆地区的行军蚁的方式, 集体出猎。行军蚁分布于亚洲和澳洲, 无法渡过珊瑚海(Coral Sea)到达新喀里多尼亚定居。粗角蚁属蚂蚁在此地所采取的行军蚁生活方式, 或许是它们在这里如此“成功”拥有优势地位, 而罕见于其他大陆的原因。威尔逊当时对这个现象的描述和推测, 事后被证明对于追踪蚂蚁进化过程非常重要。他在回忆录中写道: “这个想法在当时并未成形, 之所以记录它们, 纯粹是因为兴趣。然而, 三年后在重新建立行军蚁的进化起源时, 这份野外记录派上了大用场” (Wilson, 1994)。
1955年1月初, 威尔逊到达新赫布里底群岛(现在的瓦努阿图), 这里是前往西太平洋更偏远岛屿的驿站, 向北, 由纯热带的所罗门群岛接收来自亚洲的生物; 向南, 由亚热带的新喀里多尼亚接收来自澳洲的生物。那时, 从来没有人在这里采集过蚂蚁, 因此, 他的每一条记录都是新的发现。2月, 他专门去西澳大利亚的埃斯佩兰斯(Esperance)寻找最原始的大眼响蚁(Nothomyrmecia macrops), 用来研究蚂蚁的社会生活起源, 可惜没有找到。3月份, 转战“最标准”的热带地区新几内亚, 先在布朗河谷采集到了50种蚂蚁, 其中很多是未知的新种; 之后, 在苏加利高原山脚的雨林, 找到了以集群形式寄生在其他种类蚂蚁窝巢中的种类。
4月, 他到达新几内亚东北胡安半岛(Huon Peninsula)的沙拉威吉山脉(Sarawaget Range), 在这个从未被昆虫学家造访过的大山里采集并研究蚂蚁。这次山地之旅虽然在蚂蚁采集上没有什么特别之处, 但在返程的路上, 威尔逊却在蚂蚁的分歧进化问题上灵光乍现。他在思考新几内亚的蚂蚁分布现象。虽然每公里间的森林外貌结构变化非常小, 但蚂蚁群落的结构变化却相当惊人。这些变异来源有些是栖息地的根本变化, 但是, 许多变异并不容易解释, 尤其是地理距离增加所造成的物种变化。1958年, 他专门就此写了一篇文章(Wilson, 1994):
无论从比较生态学的哪一个层面来评价, 新几内亚蚂蚁群落最大的特点莫过于物种超级丰富。……除了单纯的数量外, 个别物种不一致的片断分布增加了整个群落的复杂度。没有任何两个地点具有完全相同的动物群落。想想看, 上百种蚂蚁因此而发生进化, 显然, 整个新几内亚动物群落的时空结构必定表现出万花筒般的面貌。这对个别蚂蚁物种的进化所产生的效果一定是非常可观的, 很可能促进了局域种群在遗传方面的分歧进化…….。
威尔逊非常高兴自己能在未经训练的采集和长途考察中感悟出一个事后被证明应用广泛的生态学模式, 也就是生物类群循环理论。完成新几内亚群岛考察后, 他在澳洲昆士兰雨林继续调查了一周。之后, 在斯里兰卡寻找到了全世界最罕见的西蒙原臭蚁(Aneuretus simoni)。至此, 热带岛屿的野外工作结束。
3 生物类群循环理论的提出
对新几内亚和美拉尼西亚蚂蚁的考察研究, 从根本上改变了威尔逊的科学生涯, 使其从一个传统的系统学家和博物收藏家转变为一个非常有洞察力和口才出众的生物地理学家、进化生物学家和生物多样性保护者。从南太平洋岛屿回来不久, 他就开始整理和发表考察成果, 其中, 有关生物类群循环的两篇论文成为了他与麦克阿瑟关于物种平衡理论的序篇。1959年, 威尔逊提出了初步的理论和分析, 然后成功地将这一理论扩展为更全面、更完整的岛屿生物历史和生态动态理论(Wilson, 1961)。
事实上, 20世纪50年代后期的生物地理学已具备了这种革命性思想成熟的时机, 理论发展到极限, 积累了一堆例外和问题, 未能将观察到的格局与其他学科结合起来。然而, 生态学、进化生物学和种群生物学的新进展, 使这种综合成为可能。正如威尔逊在阐述他的理论时所描述的那样, “普遍适应的本质和主要生物迁移扩散背后的机制显然是现代进化理论的重大问题之一。我们现在的知识可以并且应该能对这些因果过程进行更精细的分析。”
威尔逊将生物类群循环定义为3个阶段的进化和生态变化序列。第一阶段的物种是那些在群岛或区域具有相对连续地理分布范围的物种, 它们的性状无地理差异。岛屿种群已适应大陆来源地的海滨栖息地, 因此易于扩散, 并可预适应和拓殖定居于具有类似栖息地特征的岛屿边缘。随后, 这些拓殖种群就会进入“进化陷阱” (Mayr, 1942; Darlington, 1957), 因为海滨栖息地不仅是边缘的, 而且是短暂的, 容易受到演替或大规模风暴等灾难性事件的破坏。然而, 一些拓殖种群可能会通过适应和入侵美拉尼西亚岛体内部的雨林, 从而逃离海滨生境, 并与拓殖和祖先种群隔离, 最终为群岛种群的进化分歧创造条件。如果进化分歧持续到后代种群中形成新种的程度, 那么它们就进入生物类群循环的第二阶段。这些岛屿特有种经历了性状更替和生态位转化, 因此变得越来越特化于岛屿内部生境。
如果分化和特化进展到这个程度, 岛屿本土特有种就容易在群岛内和整个群岛灭绝, 从而进入生物类群循环的第三阶段。曾经广布的岛内本土特有种的地理范围缩小, 成为局限于一两个岛屿的残余种。威尔逊假设: “对大多数种群来说, 下一个也是最后一个步骤是局域灭绝(整个群岛), 或极端特有化, 并可能因第一和第二阶段物种的入侵产生的新竞争而加速。”他用非常清晰而又深刻的术语描述了这一系列相互关联的进化和生态现象的驱动力: 种间相互作用。在海滨及其边缘和临时栖息地的不同拓殖过程之后, 种间相互作用驱动了岛内物种行为、栖息地和地理分布的改变, 以及种群与特有种的分化和灭绝。
Wilson (1961)通过描述物种饱和数量如何随岛屿面积和隔离度的变化, 进一步论证了生物类群循环的地理背景。他将种面积关系, 至少部分归因于岛屿面积和物种灭绝间的反比关系。需要指出的是, 威尔逊对种面积关系的数学建模相对简单, 但基于相同的公式, MacArthur和Wilson (1963)的幂函数或双对数模型将其作为数学核心。无论如何, 威尔逊确实观察到幂函数模型的幂在动物类群间和动物分布区间的差异, 他认为变异残差(在种面积关系中分散)是由人为干扰导致的岛屿间栖息地差异造成的, 或者仅仅是由于岛屿动物取样完整性不同导致的。这就是威尔逊的初始理论的数学运算应用程度, 两篇论文都没有提供描述循环过程平衡的数学公式。
虽然生物类群循环理论是描述性的, 但其概念框架的前景是不可否认的, 在科学革命中发挥了至关重要的先驱作用, 为岛屿生物地理学提出了一个引人注目的新范式, 超越了传统的岛屿生物地理学静态理论, 证明了生态和进化过程是动态的、反复的和相互关联的。
4 与麦克阿瑟的第一次合作: 岛屿物种平衡理论
1961年, 在纽约举行的美国科学促进会会议上, 拉里·斯洛博金(Larry Slobodkin)将麦克阿瑟(两人都是哈钦森的研究生)介绍给威尔逊。威尔逊和麦克阿瑟有很多共同点, 包括对博物学和生态学的热爱, 以及对生物多样性和地理普适性解释的渴望(Wilson 1994)。威尔逊觉得生物地理学处于“大混乱”状态, “新思想的时机已经成熟”, 就给麦克阿瑟分享他关于岛屿、生物类群循环和自然平衡的想法。麦克阿瑟对生物地理学也很着迷, 他很快同意与威尔逊合作, 用自己的概念综合和数学抽象能力补充威尔逊的理论结构。
他们第一次合作的目标在论文最后一段有明确的说明: “处理可能适用于其他动物的通用平衡准则, 以及平衡状态的一些生物学含义” (MacArthur & Wilson, 1963)。这确实是一个非常恰当的模型描述。无疑, 他们的模型具备实现已经酝酿了几十年的科学革命的潜力。但他们认为, 首先有必要在一篇严谨的论文中阐明他们模型的基本前提, 因此就促成了他们第一篇合作论文在Evolution发表。
但是, 这篇具有革命性的论文的战略意义有限。虽然其隐含目标是进一步从定性和定量两方面发展出生物类群循环理论, 但他们首次合作关注的仅是鸟类群落的物种丰富度格局及其平衡成因, 或如他们所说, 迁入、灭绝和成种等反复过程间“非常动态的平衡”。事实上, 他们在第一次合作中过于保守, 以至于将推论局限于一个特定的类群——印度-澳洲岛屿鸟类群落。除了有限的生物类别, 显然, 包括群岛内和跨群岛的物种组成(即镶嵌分布和群落嵌套)、性状和生态位转化、生态释放, 以及岛内和岛间种群局域特化等, 都是超出这篇论文以外的有趣格局。
正如他们在摘要中所述, 该论文的主要目的是“表达平衡状态的标准和含义, 而不是将它们扩展到印度澳洲鸟类群落之外。”威尔逊和麦克阿瑟都是杰出的博物学家, 他们之前的研究清楚说明了种间差异和相互作用在塑造岛屿地理格局方面的重要性。然而, 在他们的模型中, 物种被视为“黑箱”。他们强调: 我们知道它们是不同的, 但在动态平衡的假设下, 这些差异可能不是解释物种丰富度格局的必要条件。
他们构建的物种中性模型(species-neutral model)是革命性的。这一成功很大程度上源于麦克阿瑟和威尔逊采用图形和数学互补形式巧妙地解释了两物种丰富度的普适模式。对大多数科学家来说, 由此产生的模型既容易理解又易于解释, 而且经得起逻辑推敲和可证伪预测的考验。例如, 他们不仅证明了动态平衡如何解释物种-面积关系和物种-隔离度关系, 而且还证明了为什么尽管物种组成发生了变化, 物种丰富度仍然相对稳定。此外, 他们证明了如何采用物种中性模型的数学形式估计类群平均扩散距离等种群性质, 动物多样性平衡被打破(例如飓风或火山爆发)后岛屿物种饱和度的动态, 以及再次达到均衡所需的时间。模型的图形形式还能够发展出一套关于岛屿群落生物地理动态(例如迁入、灭绝和周转率应随面积、隔离度、气候以及源区物种数量而变化)的定性和可证伪的预测, 这对他们理论的评估和未来发展同样重要。
为了证明物种平衡理论, 他们从威尔逊的物种适应区视角, 利用Dammermann (1948)关于1883年火山爆发后喀拉喀托(Krakatau)鸟类动态的详细记录, 在动态平衡背景下, 计算并检验他们的理论预测。结果发现, 到1933年, 喀拉喀托已被6种广布的本地猛蚁所拓殖。喀拉喀托火山的案例很好地证明了岛屿物种通过快速而反复的拓殖和积累, 群落趋于饱和, 最终灭绝平衡了迁入——这就形成了生物类群循环的中心前提, 成就了岛屿生物地理学的平衡理论。
喀拉喀托火山的事实也令人信服地证明了借助天然事件开展实验的效用, 同时, 也为巧妙设计岛屿生物地理学野外人工控制实验播下了创造性的种子, 包括由威尔逊和他的研究生丹尼尔·辛伯洛夫(Daniel Simberloff)所设计的经典的红树林岛礁节肢动物去除实验。
5 与麦克阿瑟的第二次合作: 岛屿生物地理学理论
1963年论文发表后, 威尔逊与麦克阿瑟成了亲密朋友。1964年12月, 他们决定写一本关于岛屿生物地理学的全面书籍, 目的是创造新的模型, 并将他们的理论模型扩展到尽可能多的生态学领域。3年后, 他们的跨时代著作The Theory of Island Biogeography出版。威尔逊后来在评价他们专著的成功时说: “麦克阿瑟和我完成了大部分我们要做的事情。以种群生物学为基础, 我们开始统一, 或至少开始统一生物地理学和生态学。”
这本专著是自达尔文和华莱士以来最全面的生物地理学理论著作。除了两章导论和发展平衡理论的第3章, 其他5章探讨了广泛的主题, 包括: r/k选择及其与进化的相关性, 以及岛屿种群的建立(第4章); 种群统计学、种间相互作用和生态位动态(第5章); 扩散曲线、群岛几何学和生物交换(第6章); 以及拓殖定居和适应性扩散之后的进化变化(第7章)。在专著中, 平衡模型最具革命性的特征, 但他们的目标远比这个更宏大、更全面。在该书出版之前, 他们给当时在密歇根大学的斯洛博金寄送了一份草稿和注释, 提出了他们的最终目标, 那就是, 不仅改变岛屿生物地理学, 而且改变整个生物地理学领域(Lomolino & Brown, 2009)。
与威尔逊关于生物类群循环的原始论文形成鲜明对比的是, 专著The Theory of Island Biogeography没有包括一幅物种分布图。可以猜想, 这是有意而为, 重在强调普适性和综合价值, 而不是过分描述和解释特殊细节。不幸的是, 他们混淆了最终目标, 在发展普适理论时, 仍以物种和过程为基础, 即物种生物地理学的基本过程(如迁入、灭绝和成种)有何不同。因此, 他们在结论中回到了非常普适的主题, 呼吁生物地理学领域“按照种群生态学和遗传学的首要原则重新制定……暂时不关注传统的高等类群分布和地质变化的作用问题……转而详细研究选定物种。物种的生物地理学需要理论和实验, 而这些理论和实验在很大程度上必须是新颖的”。
图1
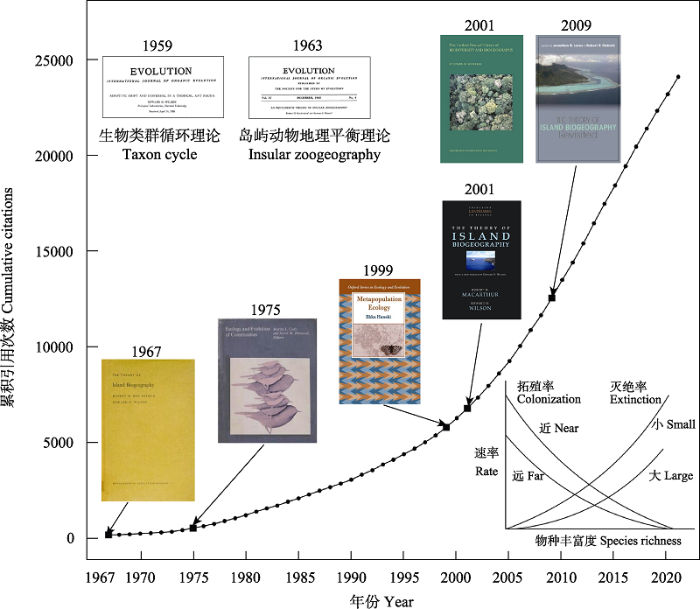
图1
《岛屿生物地理学理论》专著的累积引用次数。图示重要前期理论基础、后续纪念专辑与衍生理论(如: 集合种群理论、群落生态学的中性理论)。
Fig. 1
The cumulative citations of The Theory of Island Biogeography. In this figure, two important foundation works of this theory, the memorial monographs, and the derived theories (e.g., metapopulation theory, and the neutral theory in community ecology) are highlighted.
6 寻找岛屿生物地理学理论的实验证据
虽然岛屿生物地理学构想很好地按照他们设定的目标发展, 但终归是停留在书本上和口头上的理论。威尔逊是一个忠实的野外生态学家, 他非常渴望在现实的岛屿上检验他们的新理论。但是, 由于路途的遥远, 回到美拉尼西亚是不可能的。于是, 就产生了在离家近的地方建立海岛生物学自然实验室的想法。在这个梦想的诱惑下, 威尔逊很快想到佛罗里达群岛是一个合乎逻辑的去处, 因为有众多的岛礁可供选择。
威尔逊认为, 研究实验岛屿生物地理学最好的方法是从许多生态相似但面积和距离不同的小岛开始, 然后把它们变成微型的喀拉喀托火山。也就是说, 找到一种方法来根除动物群落, 然后跟踪物种重新拓殖的过程。第一次选择的地点是地处墨西哥湾的干龟群岛(Dry Tortugas)。1965年夏天, 他与研究生系统调查了岛礁, 在上面发现了为数不多的植物和节肢动物。他们的想法是让现状继续, 等待飓风完全清除了岛屿生物后, 跟踪观察植物和节肢动物的重新拓殖过程。如有神助, 当年就有两次飓风席卷了干龟群岛。当1966年返回调查时发现, 几个月前观察的岛礁没有了陆地生物。这样, 他们的研究就开始了。
然而, 威尔逊对这个实验的前景越来越不满意。因为岛屿和动植物群落太少, 飓风太少也难以预测, 并且无法控制。于是, 他又去了佛罗里达湾的红树植物岛(Mangrove Islet), 这里没有干龟群岛的缺点, 但有其他不足, 那就是飓风不可能把所有的节肢动物都从茂密的红树林叶子上带走(Wilson, 2009)。
威尔逊与博士生丹尼尔选择了位置最有利和最适合的红树植物岛, 请专业的灭虫专家史蒂夫·坦德里希(Steve Tendrich)在不损害植被的情况下, 使用甲基溴熏蒸法“去除”节肢动物。之后, 丹尼尔开始每月对每个红树植物岛逐厘米进行检查的艰苦过程, 同时, 在分类学家的帮助下, 将这些小岛上的甲虫、树皮虱、蛾类、蜘蛛、螨虫和其他节肢动物分类到种水平(Simberloff & Wilson, 1969)。
两年内, 所有红树植物岛上的物种数量差不多恢复到了灭绝前的水平。如预期的那样, 最遥远的红树植物岛一开始的物种数很少, 后来又恢复到原来的低水平, 从而证明了物种平衡的存在。然而, 令人惊讶的是, 红树植物岛间物种组成存在差异, 同一岛上的物种在灭绝前后也存在差异(Simberloff & Wilson, 1971)。此外, 大多数物种重新定居的速度和广泛而频繁的周转, 也基本符合麦克阿瑟-威尔逊平衡模型。
7 岛屿生物地理学理论: 科学革命的典范
麦克阿瑟和威尔逊的物种平衡理论强烈地影响了生物地理学、生态学和进化生物学等相关理论的科学思维(Kuhn, 1996)。岛屿生物地理学理论解决了20世纪该领域的前范式困境, 即, 由独立个案到多时期和多站点, 以及从扩散论向起源-扩散-适应中心思想转变, 这些思想达到了生物地理学传统静态理论的极限。作为回应, 大胆而富有远见的威尔逊与麦克阿瑟开创性地提出了生物类群循环理论和岛屿物种平衡理论, 从而化解了这场科学危机。
麦克阿瑟和威尔逊的物种平衡理论之所以能够得到迅速和压倒性的赞誉, 在很大程度上可归功于3个因素: (1)相对简单性和由此产生的美学吸引力; (2)及时性, 即, 在具备必要的科学工具的情况下取得先进性; (3)真实、协作性质, 即, 现实性。
岛屿生物地理学理论提出后, 我们关于多尺度生物地理过程的理解取得了巨大进步(图1), 在理论方面, 为分析和解释宏观群落提供了生态过程新模型(Cody & Diamond, 1975), 并衍生形成了集合种群理论(Hanski, 1999)和中性理论(Hubbell, 2001), 在实践方面, 有效地推动了生物多样性保护和自然保护地规划(Diamond et al, 1976)。2017年, 在The Theory of Island Biogeography发表50年之际, 世界各地的生态学家和生物地理学家通过各种方式纪念这一伟大成果对生态学和进化学等领域的贡献。2016年的国际岛屿生物学大会在总结该领域50年来发展成果的基础上, 通过问卷调查, 系统梳理了岛屿生物地理学未来的发展路线图, 遴选出了50个亟需重点解决的科学问题(Patiňo et al, 2017)。可以预见, 生物地理学正在经历一波新的范式转变, 或者更恰当地说是范式整合(Whittaker & Fernández-Palacios, 2007; Losos & Ricklefs, 2009; Warren et al, 2015; Whittaker et al, 2017)。但无论如何, 新的范式应该与威尔逊的原始理论共享一些基本属性(Pickett et al, 2007; Lomolino & Brown, 2009)。
参考文献
The fauna of Krakatau, 1883-1933
The geographical distribution of cold-blooded vertebrates
DOI:10.1086/396077 URL [本文引用: 2]
Island biogeography and conservation: strategy and limitations
The reticulating phylogeny of island biogeography theory
DOI:10.1086/648123 URL [本文引用: 2]
An equilibrium theory of insular zoogeography
DOI:10.1111/evo.1963.17.issue-4 URL [本文引用: 3]
Climate and evolution
Perspectives in biogeography
DOI:10.4039/Ent95299-3 URL [本文引用: 1]
A roadmap for island biology: 50 fundamental questions after 50 years of the Theory of Island Biogeography
DOI:10.1111/jbi.2017.44.issue-5 URL [本文引用: 1]
Experimental zoogeography of islands: Defaunation and monitoring techniques
DOI:10.2307/1934855 URL [本文引用: 1]
Experimental zoogeography of islands: A two-year record of colonization
DOI:10.2307/1933995 URL [本文引用: 1]
Mammals and the nature of continents
DOI:10.2475/ajs.241.1.1 URL [本文引用: 1]
Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson
DOI:10.1111/ele.12398
PMID:25560682
[本文引用: 1]

The study of islands as model systems has played an important role in the development of evolutionary and ecological theory. The 50th anniversary of MacArthur and Wilson's (December 1963) article, 'An equilibrium theory of insular zoogeography', was a recent milestone for this theme. Since 1963, island systems have provided new insights into the formation of ecological communities. Here, building on such developments, we highlight prospects for research on islands to improve our understanding of the ecology and evolution of communities in general. Throughout, we emphasise how attributes of islands combine to provide unusual research opportunities, the implications of which stretch far beyond islands. Molecular tools and increasing data acquisition now permit re-assessment of some fundamental issues that interested MacArthur and Wilson. These include the formation of ecological networks, species abundance distributions, and the contribution of evolution to community assembly. We also extend our prospects to other fields of ecology and evolution - understanding ecosystem functioning, speciation and diversification - frequently employing assets of oceanic islands in inferring the geographic area within which evolution has occurred, and potential barriers to gene flow. Although island-based theory is continually being enriched, incorporating non-equilibrium dynamics is identified as a major challenge for the future. © 2014 John Wiley & Sons Ltd/CNRS.
Adaptive shift and dispersal in a tropical ant fauna
DOI:10.1111/evo.1959.13.issue-1 URL [本文引用: 1]
The nature of the taxon cycle in the Melanesian ant fauna
DOI:10.1086/282174 URL [本文引用: 2]
Island biogeography in the 1960s
Island biogeography: Taking the long view of nature’s laboratories
DOI:10.1126/science.aam8326 URL [本文引用: 1]

{kind=link}
{kind=link}