



竹类植物具有重要的经济和生态价值, 很早就被人类认识和研究(McClure, 1966)。根据目前在国际上广为接受的APG IV (Angiosperm Phylogeny Group IV)系统, 竹亚科隶属于被子植物单子叶分支禾本目禾本科, 是禾本科的12个亚科之一, 与早熟禾亚科、稻亚科一起构成BOP (Bambusoideae-Oryzoideae-Pooideae)分支(GPWG, 2001; GPWG II, 2012; Soreng et al, 2017; Saarela et al, 2018)。近年来的分子系统发育研究支持将竹亚科分为莪莉竹族(Olyreae)、青篱竹族(Arundinarieae)和箣竹族(Bambuseae) 3个族, 它们又分别属于草本竹类分支(herbaceous bamboos)、温带木本竹类分支(temperate woody bamboos, TWB)和热带木本竹类分支(tropical woody bamboos), 其中热带木本竹类又可进一步分为旧世界热带木本竹分支(paleotropical woody bamboos, PWB)和新世界热带木本竹分支(neotropical woody bamboos, NWB); 中国仅有旧世界热带木本竹类和温带木本竹类(Kelchner & BPG, 2013; Guo et al, 2019)。据最新统计, 全世界共有竹类127属约1,680种(Clark & Oliveira, 2018)。中国是世界上竹类资源最丰富的国家, 其物种数、分布面积、蓄积量等均居世界各国之首, 《中国植物志》(Flora Reipublicae Popularis Sinicae, FRPS)和Flora of China (FOC)记载的竹类达34属534种(Li et al, 2006)。
竹类是公认的禾本科植物分类困难类群。究其原因, 首先是竹类标本采集要求高, 过程较为繁琐, 一般需要地下茎、秆节、秆箨、分枝和完整的叶子(包括叶鞘、叶舌、叶耳和叶片)等部位的标本, 缺少某些关键性状(如秆箨)会增加鉴定的难度和降低鉴定的准确性, 因此很多植物标本采集者往往会忽视竹类标本的采集(Soderstrom & Young, 1983)。其次, 竹类植物具有特殊的生物学特性。长期以来, 植物分类学基于繁殖器官的特征来描述和鉴别物种, 但由于竹类特殊的开花习性, 绝大多数木本竹类均为一次性开花, 开花后死亡, 且开花周期具有异质性(3-120年) (Janzen, 1976), 采集的营养体标本很难与常常仅包括繁殖体的模式标本进行对比鉴定。因此, 竹类物种的鉴定问题以及较少的标本记录对其研究形成了极大的阻碍, 并在很大程度上制约着竹类植物的培育、利用和保护。
植物标本是植物研究者在长期的科研活动中采集和保存的凭证材料, 体现着人类对某个类群的认识, 也是自然遗产的实体记录; 信息完备的植物标本是物种分类、分布及系统演化等学科的重要资料(Bridson & Forman, 1998), 其携带的信息有着重要的研究价值。最早的标本主要服务于物种发现和志书编写, 随着标本数目增加和相关信息的数字化, 基于植物标本开展的研究形式也越来越多样化。如张玉雪等(2018)利用国家标本资源共享平台(National Specimen Information Infrastructure, NSII,
对于竹类这样一个有巨大价值但研究困难的类群, 关于其馆藏标本及地理分布的研究却鲜见报道。本研究利用中国竹类植物馆藏标本的采集信息, 以省级(直辖市、自治区等)行政区划为基本单元, 分析了中国竹类植物的采集现状和采集偏差; 同时结合气候数据, 对中国竹类的地理分布状况进行研究, 以期为今后的竹类植物标本采集和物种保护等工作提供基础资料。
1 材料与方法
1.1 数据来源
本研究的数据来源主要包括: (1)国家标本资源共享平台; (2)中国数字植物标本馆(Chinese Virtual Herbarium, CVH,
1.2 数据标准化
随着植物分类学研究的不断深入, 植物科学名称的使用往往会发生变化; 且不同的采集者对标本采集信息的记录风格和完整度不一, 造成了现有标本信息不全乃至含混。因此, 本研究对获得的标本数据进行了标准化处理, 具体方法如下: (1)利用网站Taxonomic Name Resolution Service v4.1 (TNRS,
1.3 复份标本的处理
《国际藻类、菌物和植物命名法规》第八条规定: 一个采集的多个制备, 如果未注明其是同一份标本的组成材料时, 则这些制备互相构成复份标本(Turland et al, 2018)。一份完整的标本通常包含植物的各部分器官, 但由于竹类植物的特殊性, 各部位的材料往往难以展示在同一张台纸上, 而该类群的标本在采集和装订过程中很少注明是复份标本还是属于同一份标本的不同部分。因此, 为了研究的方便, 本研究将采集号、采集人、采集时间以及采集地点完全相同的标本视为一份标本, 只保留其中一条标本信息。
1.4 采集完整度的评估
标本采集偏差是评价标本采集完整度的重要参数, 具体可分为两种: 一是受地理、交通、气候等条件和采集人的记录习惯等因素的影响, 在采集过程中会出现采集地点不均衡, 部分地区遗漏或采集数量过少等影响数据完整性的情况, 即采集地点偏差; 二是由于采集对象的类群特点、分布频率、物候和采集难易程度不同等, 导致采集者对某些类群重复采集, 而某些类群采集很少, 即采集类群偏差。本研究利用比值法和斜率法分别对其进行评估, 并比较了这2种方法评估结果的差异。
(1)比值法。该方法利用观察丰富度和估计丰富度的差值与估计丰富度的比值大小来评估采集的完整度, 是生物多样性领域常用的方法之一。估计物种的丰富度, 常用的有Chao, Jackknife和Bootstrap等模型, 其中Chao 1估计量最早由Chao于1984年提出, 其基本假设认为如果在一个群体中随机抽样, 当稀有的物种(采集份数仅为1)依然不断被发现时, 则表明还有稀有的物种没有被发现; 直到所有物种至少被抽到2次(采集份数至少为2), 则表明不会再有新的物种被发现(Chao, 1984)。其计算公式为:
其中, Sobs表示样本中观察到的物种数, F1表示只有1个个体的物种数, F2表示只有2个个体的物种数。
本文采用标本实际记载物种数和Chao 1估计量的差值与Chao 1估计量的比值来评估竹类各属标本以及中国各省级行政区的采集完整度, 即采集完整度 = (观测值 - 理论值)/理论值, 比值越小代表采集越不完整。根据实际情况, 在本研究中我们限定比值< -0.5时为采集不完整; [-0.5, -0.3]为采集较完整; > -0.3为采集完整。
(2)斜率法。该方法基于物种累积曲线(species accumulation curve, SAC), 当抽样数目达总标本数的90%时对应的曲线斜率作为采集完整度的判断标准(Tittensor et al, 2010; Chu et al, 2014)。物种累积曲线最早是生态学中用于估计物种数量的一种方法, 原理是当采集的标本逐渐增多时, 新种发现的速率就会下降, 如果标本数量无限大时, 物种累积曲线就趋近于物种库总数。
本研究将竹类各属(采集类群偏差)以及中国各省级行政区(采集地点偏差)的全部标本记录进行随机抽样并统计每次获得的物种数目, 抽样数目以总标本数的1%为梯度依次递增, 并且每个数目进行10,000次重抽样。此后, 以每次抽样的标本数目作为横坐标, 以10,000次抽样的平均物种数为纵坐标分别绘制各属以及各省区的物种累积曲线, 并计算抽样数目为总标本数90%时曲线所对应的斜率。当曲线斜率值接近0时, 表明该属或该地区的采集较完整, 当斜率值接近1时, 则表明采集不完整, 可能有新种存在。本研究以0.005作为斜率阈值对采样完整度进行评估, 当斜率≥ 0.005时表示采集不完整, 反之则表明采集完整。
1.5 气候因子变量数据筛选
从全球气候数据库WorldClim (
1.6 不同竹类分支的MaxEnt模型分析
将获得的中国竹类标本数据按照最新的竹亚科系统学结果分成旧世界热带木本竹类和温带木本竹类2个分支(Kelchner & BPG, 2013), 把竹类标本的分布点经纬度信息和环境图层导入MaxEnt软件(Phillips et al, 2006)。在模型分析中, 最大迭代次数设置为10,000, Bootstrap重复运算20次, 启用random seed、勾选绘制响应曲线(response curves)和刀切法(Jackknife检验)来分析影响竹类分布状况的环境变量, 其中25%的数据作为训练数据(training data), 75%的数据用于建立模型。MaxEnt软件会自动绘制ROC曲线(receiver operating characteristic curve)并计算曲线下的面积AUC (areas under the receiver operating characteristic curves)来对模型的拟合效果进行评价。当AUC < 0.5时, 表明模型的预测效果很差, 在0.6-0.9之间表明模型预测效果一般, AUC > 0.9说明模型预测的效果很好(王运生等, 2007; Liu et al, 2005。本文模型拟合的AUC均> 0.9。
2 结果
2.1 我国竹类植物标本基本情况
初步统计表明, 中国竹类植物标本藏量为51,194份, 本研究获得其中44,694份(87%)标本的信息。通过异名处理后, 剩余35,024条有效信息; 剔除采集地点缺失数据和在中国行政区以外的采集记录, 剩余30,156份标本。统计发现, 中国共有108家单位的标本馆(室)收藏有竹类标本, 馆藏量从1-13,369份不等, 其中南京大学生命科学学院植物标本室的竹类植物标本数量最高, 为13,369份, 占全国总馆藏量的1/4; 剔除复份标本后, 发现馆藏量排名前15位的标本馆保存了86%的竹类标本(图1A)。除黑龙江省、吉林省和澳门特别行政区外, 中国各省级行政区域均有竹类标本的采集记录, 但新疆维吾尔自治区、内蒙古自治区和辽宁省的馆藏标本均采自栽培竹类。从各省区的标本数量来看, 馆藏竹类植物标本的采集主要集中于长江以南地区, 采集超过1,000份的省区有10个, 其中以云南最多, 浙江次之, 这两省采集的标本份数均超过4,000份, 占中国竹类标本的1/3 (图2)。而长江以北地区的馆藏竹类植物标本主要采集于黄河以南区域, 黄河以北地区竹类标本记录稀少, 如青海省竹类标本不足10份。
图1
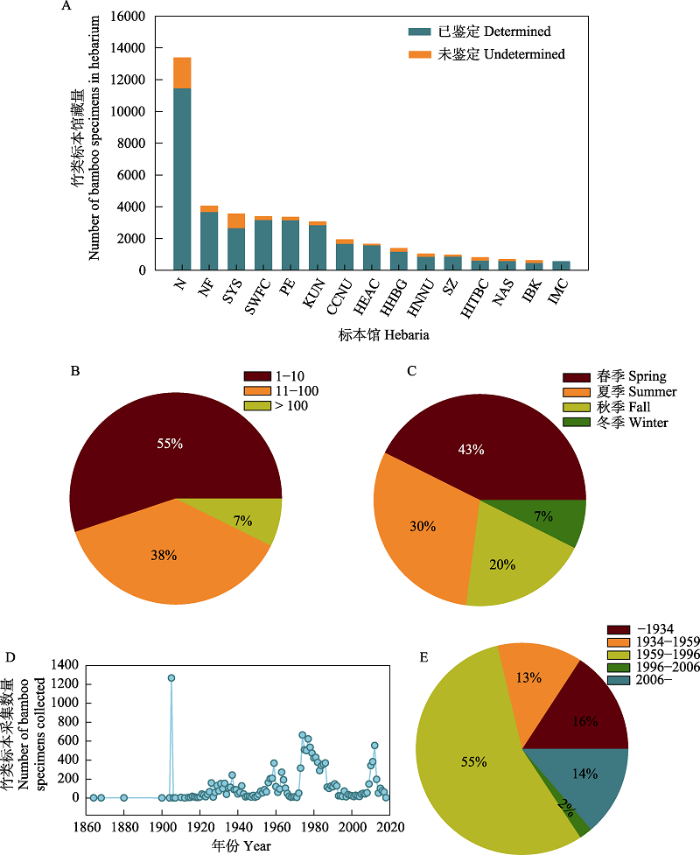
图1
中国馆藏竹类标本数据统计。A: 中国馆藏竹类植物标本数量前15的标本馆馆藏量(标本馆代码见附录2); B: 国产竹类标本采集份数统计; C: 国产竹类标本采集季节统计; D: 国产竹类标本采集年份统计; E: 国产竹类各时期标本采集量统计。
Fig. 1
Statistical depiction of bamboo specimen data collected in China. A, Herbarium statistics of top 15 herbaria with bamboo specimens collected in China (The code of the herbarium is shown in Appendix 2); B, Status of bamboo specimens in China at the species level; C, Season of collection of bamboo specimens in China; D, Year of collection of bamboo specimens in China; E, Relative proportions of bamboo specimens collected at various periods in China.
图2
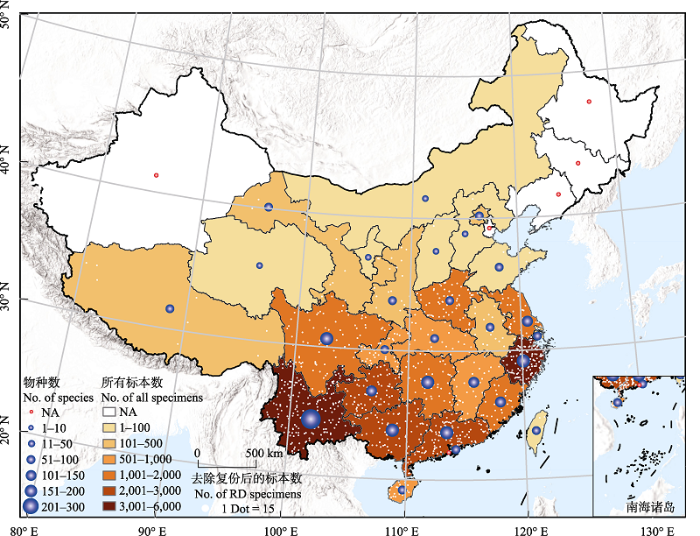
图2
中国各省级行政区竹类标本采集份数和物种数的分布。白色圆点表示去除复份标本后的标本数量, 每个白点代
Fig. 2
Distribution of bamboo specimens and species in different provinces of China. The white dots indicate the number of specimens that removing duplicate (RD) specimens. Each white dot represents 15 specimens.
最新的文献记载中国产竹类38属(Vorontsova, 2016), 但标本记录仅有33属, 有5个属在共享数据库中没有标本, 包括须弥筱竹属(Himalayacalamus) (耿伯介, 1983)、多枝竹属(Holttumochloa, 我国新分布属) (Wong, 1993)、纪如竹属(Hsuehochloa, 2018年发表的新属) (Zhang et al, 2018)、冷箭竹属(Sarocalamus, FRPS中被处理为异名) (Stapleton, 2004)和华箬竹属(Sasamorpha, FRPS中被处理为异名) (Nakai, 1931), 这些类群标本数据的缺失主要是相应标本馆未能对标本鉴定信息进行适时更新造成的。从属级水平看, 平均标本数大于10份的属有21个, 仅占所有属的62% (表1); 从物种水平看, 馆藏标本物种的平均采集量为30.5份, 且各物种的实际采集量间存在着很大差异。55%的物种采集量不足10份, 其中15%的物种仅采集了1份标本; 仅有7%的物种采集量在100份以上(图1B), 如湖南刚竹(Phyllostachys carnea)采集量仅有1份, 而同属的桂竹(P. bambusoides)采集量则高达910份。
表1 中国国产竹类各属标本采集情况统计与分析
Table 1
| 属 Genus | 文献记录物种数 Number of species based on literature | 标本记录物种数 Number of species based on specimens | 标本数 Specimens | 每个物种平均标本数 Mean specimens per species | Chao 1 | 比值 Ratio | 斜率 Slope |
|---|---|---|---|---|---|---|---|
| 酸竹属 Acidosasa | 10 | 6 | 32 | 5.3 | 7 | -0.143 | 0.003 |
| 悬竹属 Ampelocalamus | 14 | 9 | 98 | 10.9 | - | - | 0.007 |
| 箣竹属 Bambusa | 90 | 62 | 2,188 | 35.3 | 65 | -0.046 | 0.003 |
| 巴山竹属 Bashania | 6 | 0 | 0 | - | - | - | - |
| 单枝竹属 Bonia | 4 | 3 | 35 | 11.7 | 3 | 0.000 | 0.000 |
| 空竹属 Cephalostachyum | 7 | 5 | 91 | 18.2 | - | - | 0.000 |
| 方竹属 Chimonobambusa | 38 | 21 | 926 | 44.1 | 21 | 0.000 | 0.000 |
| 香竹属 Chimonocalamus | 10 | 11 | 263 | 23.9 | - | - | 0.000 |
| 牡竹属 Dendrocalamus | 37 | 31 | 1,146 | 37.0 | 33 | -0.061 | 0.002 |
| 镰序竹属 Drepanostachyum | 4 | 1 | 2 | 2.0 | - | - | 0.000 |
| 箭竹属 Fargesia | 87 | 56 | 831 | 14.8 | 59 | -0.051 | 0.003 |
| 铁竹属 Ferrocalamus | 2 | 1 | 22 | 22.0 | - | - | 0.000 |
| 贡山竹属 Gaoligongshania | 1 | 1 | 5 | 5.0 | - | - | 0.000 |
| 井冈寒竹属 Gelidocalamus | 12 | 8 | 76 | 9.5 | 8 | 0.000 | 0.003 |
| 巨竹属 Gigantochloa | 15 | 9 | 55 | 6.1 | 9 | 0.000 | 0.003 |
| 须弥筱竹属 Himalayacalamus | 2 | 0 | 0 | - | - | - | - |
| 多枝竹属 Holttumochloa | 1 | 0 | 0 | - | - | - | - |
| 纪如竹属 Hsuehochloa | 1 | 0 | 0 | - | - | - | - |
| 箬竹属 Indocalamus | 32 | 20 | 1,317 | 65.9 | 22 | -0.091 | 0.002 |
| 大节竹属 Indosasa | 16 | 15 | 378 | 25.2 | - | - | 0.000 |
| 梨藤竹属 Melocalamus | 6 | 3 | 72 | 24.0 | - | - | 0.006 |
| 梨竹属 Melocanna | 1 | 1 | 8 | 8.0 | - | - | 0.000 |
| 新小竹属 Neomicrocalamus | 1 | 1 | 8 | 8.0 | - | - | 0.000 |
| 少穗竹属 Oligostachyum | 18 | 8 | 61 | 7.6 | - | - | 0.005 |
| 刚竹属 Phyllostachys | 61 | 52 | 5,868 | 112.8 | 53 | -0.019 | 0.001 |
| 苦竹属 Pleioblastus | 18 | 15 | 778 | 51.9 | - | - | 0.006 |
| 茶秆竹属 Pseudosasa | 17 | 14 | 577 | 41.2 | 22 | -0.364 | 0.006 |
| 泡竹属 Pseudostachyum | 1 | 1 | 65 | 65.0 | - | - | 0.000 |
| 冷箭竹属 Sarocalamus | 3 | 0 | 0 | - | - | - | - |
| 赤竹属 Sasa | 9 | 10 | 83 | 8.3 | 28 | -0.643 | 0.013 |
| 华箬竹属 Sasamorpha | 3 | 0 | 0 | - | - | - | - |
| 𥯨簩竹属 Schizostachyum | 13 | 9 | 368 | 40.9 | 9 | 0.000 | 0.000 |
| 业平竹属 Semiarundinaria | 5 | 3 | 12 | 4.0 | - | - | 0.021 |
| 倭竹属 Shibataea | 7 | 7 | 110 | 15.7 | - | - | 0.003 |
| 唐竹属 Sinobambusa | 10 | 8 | 146 | 18.3 | - | - | 0.002 |
| 筱竹属 Thamnocalamus | 2 | 1 | 7 | 7.0 | 1 | 0.000 | 0.000 |
| 泰竹属 Thyrsostachys | 2 | 2 | 23 | 11.5 | - | - | 0.010 |
| 玉山竹属 Yushania | 70 | 51 | 609 | 11.9 | 53 | -0.038 | 0.003 |
| 总计 Total | 636 | 445 | 16,334 | 36.7 | 476 | -0.065 | 0.003 |
对竹类标本的采集月份进行统计, 结果显示标本采集工作主要集中于春夏两季, 其采集量占全部标本的73%, 而冬季最少, 仅占总量的7% (图1C)。对采集年份的统计结果表明, 标本采集量呈现4个明显的波峰(图1D)。结合中国禾本科等研究历史, 我们将采集时间段划分为5个阶段: 1934年以前为第一阶段, 以国外采集者为主, 采集量占16%; 第二阶段为1934-1959年(FRPS启动前), 采集量占13%; 第三阶段为1959-1996年(FRPS启动后到竹亚科分册完成), 采集量最多, 采集量占55%; 第四阶段为1996-2006年(FRPS竹亚科分册完成到FOC竹亚科部分完成), 采集量仅占2%; 第五阶段为2006年至今(FOC竹亚科部分完成后), 采集量占14% (图1E)。统计标本所处的生活史阶段, 发现中国馆藏竹类植物标本主要以营养性状为主, 比例高达69%, 仅有1% (将近1,000份)的标本同时具有花果性状, 分布在27属124竹种中, 而温带木本竹类的具花标本数量远高于旧世界热带木本竹类, 且有花标本主要采集于FRPS竹亚科分册出版(1996年)之前。
2.2 采集完整度评估
物种丰度观测值和预测值最大的均为云南省, 远超其他省份; 位于黄河以北的各省区观测值和预测值都偏小, 其中内蒙古、宁夏和青海3省标本采集总数非常少, 均不超过10份(图2)。各省市标本的观测值和预测值的差值与预测值的比值在-0.610到0之间波动, 平均值为-0.299, 其中安徽、北京和台湾为采集不完整的省区(其中台湾极可能是因为本研究未能收录其相关标本馆数据之故); 而甘肃、广东、海南、河北、山西、四川和西藏为采集较完整区域, 其余地区为采集完整区域。比值大小和标本采集份数的相关性分析结果表明二者之间的相关性较弱(附录1A, R2 = 0.21)。此外, 地理邻近区域的采集情况也参差不齐, 如湖南和湖北彼此接壤, 但标本采集数量和物种数却都存在较大差异。斜率法结果表明, 安徽、北京、海南、河北、内蒙古、青海、上海、山西、台湾和西藏为采集不完整区域(其中台湾可能是受本研究收集数据影响), 其余省区采集完整(表2)。与比值法相比, 在标本采集总量大的情况下, 二者结果差异不明显。
表2 中国各省级行政区竹类植物标本采集情况统计
Table 2
| 省份 Province | 物种数 Species | Chao 1 | 比值 Ratio | 斜率 Slope | 省份 Province | 物种数 Species | Chao 1 | 比值 Ratio | 斜率 Slope |
|---|---|---|---|---|---|---|---|---|---|
| 安徽 Anhui | 46 | 118 | -0.610 | 0.005 | 内蒙古 Inner Mongolia | 3 | - | - | 0.067 |
| 北京 Beijing | 14 | 32 | -0.563 | 0.005 | 宁夏 Ningxia | 2 | - | - | 0.000 |
| 重庆 Chongqing | 45 | 48 | -0.063 | 0.002 | 青海 Qinghai | 1 | - | - | 0.200 |
| 福建 Fujian | 91 | 134 | -0.321 | 0.004 | 陕西 Shaanxi | 37 | 48 | -0.229 | 0.004 |
| 甘肃 Gansu | 25 | 42 | -0.405 | 0.004 | 山东 Shandong | 12 | 14 | -0.143 | 0.004 |
| 广东 Guangdong | 124 | 179 | -0.307 | 0.003 | 上海 Shanghai | 22 | 30 | -0.267 | 0.006 |
| 广西 Guangxi | 132 | 170 | -0.224 | 0.003 | 山西 Shanxi | 6 | 11 | -0.455 | 0.008 |
| 贵州 Guizhou | 99 | 133 | -0.256 | 0.003 | 四川 Sichuan | 112 | 189 | -0.407 | 0.004 |
| 海南 Hainan | 48 | 89 | -0.461 | 0.005 | 台湾 Taiwan | 25 | 63 | -0.603 | 0.006 |
| 河北 Heibei | 9 | 13 | -0.308 | 0.008 | 香港 Hong Kong | 25 | 25 | 0.000 | 0.001 |
| 河南 Henan | 46 | 56 | -0.179 | 0.002 | 西藏 Xizang | 31 | 55 | -0.436 | 0.005 |
| 湖北 Hubei | 48 | 64 | -0.250 | 0.003 | 云南 Yunnan | 255 | 280 | -0.089 | 0.002 |
| 湖南 Hunan | 101 | 135 | -0.252 | 0.004 | 浙江 Zhejiang | 125 | 157 | -0.204 | 0.003 |
| 江苏 Jiangsu | 59 | 77 | -0.234 | 0.003 | 总计 Total | 446 | 478 | -0.067 | 0.001 |
| 江西 Jiangxi | 60 | 76 | -0.211 | 0.004 |
通过比值法评估各属的采集完整度时, 由于以悬竹属(Ampelocalamus)为代表的23个属的物种标本数据量不符合计算条件, 无法计算Chao 1值。剩余的15个属中仅有赤竹属(Sasa)的比值< -0.5, 为采集不完整的属, 茶杆竹属(Pseudosasa)为采集较完整的属, 其余13个属为采集完整的属。值得注意的是, 仅有赤竹属和茶秆竹属两个属的Chao 1估计量超过了文献记载数, 其余各属的估计量均小于文献记载数(表1)。比值大小和各属标本采集份数的相关性分析结果表明二者之间的相关性较弱(附录1B, R2 = 0.006)。斜率法评估的结果显示悬竹属、梨藤竹属(Melocalamus)、少穗竹属(Oligostachyum)、苦竹属(Pleioblastus)、茶秆竹属、冷箭竹属、赤竹属、业平竹属(Semiarundinaria)、泰竹属(Thyrsostachys)等9个属采集不完整。比较发现, 在标本采集量不足时, 两种方法都存在较大误差甚至失效, 但斜率法的适用性优于比值法。
2.3 气候因子对竹类植物地理分布的影响
根据竹类植物馆藏标本经纬度信息绘制竹类分布图, 发现旧世界热带木本竹类分布于长江以南地区, 而温带木本竹类可分布到黄河流域, 且黄河以北还有零星分布(图3A)。MaxEnt模型分析的刀切法结果显示(图3B), 旧世界热带木本竹类的分布受最冷月最低温的影响最大, 其次是年温度变化范围, 而水分条件的影响较小; 温带木本竹类的分布主要受年均降水量的影响, 其次是最冷月最低温。对比发现, 最冷月最低温是影响两大分支分布的共同影响因素, 水分对温带木本竹类的影响比对旧世界热带木本竹类明显, 而温度对旧世界热带木本竹类的限制性更强。MaxEnt模型模拟结果表明, 旧世界热带木本竹类同等检测特异性和敏感性的阈值(equal test sensitivity and specificity cloglog threshold)为0.4813, 其适生区域主要为我国珠江流域以南的地区(图4绿色区域); 温带木本竹类同等检测特异性和敏感性的阈值为0.4817, 其适生区在长江流域地区(图4红色部分); 两者之间存在一带状的重叠区, 为两者的共同适生区(图4黄色区域), 值得注意的是云南大部分区域和四川盆地均为其适生重叠区域。
图3
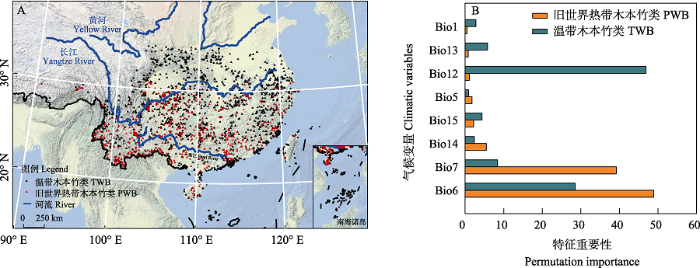
图3
国产竹类地理分布图(A)及影响因子重要性(B)。Bio1: 年均温; Bio5: 最热月最高温; Bio6: 最冷月最低温; Bio7: 年温度变化范围; Bio12: 年均降水量; Bio13: 最湿月降水量; Bio14: 最干月降水量; Bio15: 降水量季节性变异系数。
Fig. 3
Distribution map of bamboos in China (A) and permutation importance of the climatic variables (B). PWB, Paleotropical woody bamboos; TWB, Temperate woody bamboos; Bio1, Annual mean temperature; Bio5, Max temperature of warmest month; Bio6, Min temperature of the coldest month; Bio7, Temperature annual range; Bio12, Annual precipitation; Bio13, Precipitation of wettest month; Bio14, Precipitation of driest month; Bio15, Coefficient of variation of precipitation seasonality.
图4
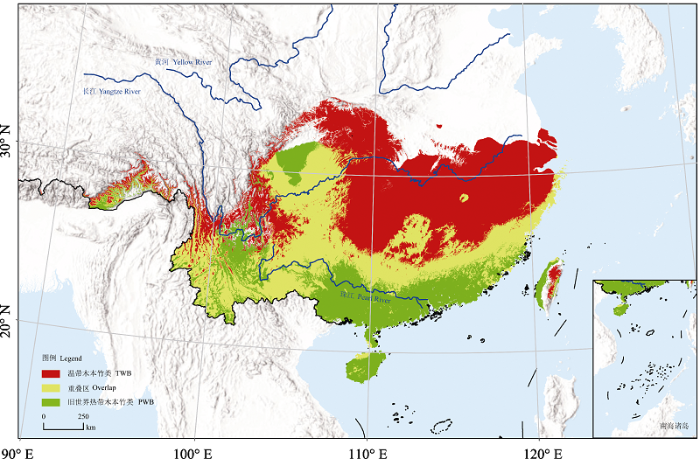
图4
MaxEnt模型对国产竹类高适生区的模拟结果
Fig. 4
Simulation results of the MaxEnt model of suitable areas for bamboos in China. PWB, Paleotropical woody bamboos; TWB, Temperate woody bamboos.
3 讨论
本研究收集整理了全国竹类植物馆藏标本的采集信息, 首先将其标准化处理后对标本的基本信息进行分析, 再利用比值法和斜率法对标本的采集类群偏差和采集地点偏差进行评估, 最后基于MaxEnt模型对国产竹类两大分支的适生区和影响其分布的主要环境因子进行评估。
3.1 竹类标本馆藏历史及现状分析
统计分析发现, 国内标本馆对竹类标本的收集和保藏存在很大的不均衡性, 竹类标本主要收藏于南京大学生命科学学院植物标本室等15个标本馆内。究其原因, 首先这些标本馆在不同时期均有研究团队开展过竹类相关研究, 积累了大量的研究材料或凭证标本; 其次竹类标本采集专业性强, 分类鉴定困难, 没有专门从事竹类研究人员的标本馆一般不会或很少保存竹类标本。南京大学是FRPS竹亚科分册完成前我国竹类植物分类研究的中心, 耿以礼教授和耿伯介教授分别是我国禾本科和竹亚科研究的开拓者, 耿伯介教授和王正平教授是FRPS竹亚科分册的卷编辑。通过采集信息我们还发现标本采集量的变化与团队的发展密切相关, 在收藏的类群上与专家研究的类群和当地的资源均有很强的相关性。基于此, 我们认为竹类标本的馆藏过于集中, 这一方面极大地方便了该类群的研究者, 使其从少数几个标本馆中即可获得全国大量竹类植物的分类与分布信息; 但另一方面也容易产生偏向性, 使本来就具有较高入门标准的竹类分类学研究更加边缘化和专业化。
将竹类标本采集的阶段性特征放到中国近代植物分类学研究的历程中, 我们发现竹类标本的采集很好地代表了国内植物分类学研究的历史阶段。首先, 在鸦片战争之后, 西方的植物采集者活跃于中国各地开展采集活动和研究工作, 如莫古礼(F. M. McClure)等。其次, 20世纪以来, 随着第一代中国植物学研究机构的建立, 不少类群(包括禾本科在内)开始了由中国植物学者主导的研究工作, 并逐渐积累植物标本, 其中耿以礼教授1934年学成归国, 专注于中国禾本科的研究。新中国成立后, 启动了植物志书的编纂项目, 标本采集和收藏活动蓬勃开展, 逐渐形成目前的规模和馆藏状况。随后在FRPS和FOC等志书编纂完成之后, 标本采集热度降低, 采集量明显减少。最近十余年, 随着分类学研究进入分子生物学时代, 发现了研究中存在的诸多问题, 需要继续采集标本, 同时分子生物学研究也需要采集凭证标本, 因此标本采集的热度又逐渐恢复, 采集量逐年增加。对竹类标本采集史的梳理为我们认识中国植物分类学研究的历程提供了一个生动的例证。
对比文献记录和标本记录的属种信息, 我们发现两者之间存在较大差异。通过文献查证和实地考察之后, 可将差异归结于以下两个原因: 一是部分竹类标本还未实现数字化和数据共享, 如四川农业大学都江堰分校竹标本室保存有易同培教授等人收藏的4,500余份标本, 多为高山竹类且包含许多模式标本, 但目前尚未进行数字化; 中国科学院华南植物园植物标本馆保存的2,000余份竹类标本也仅有151份标本信息公布在网络共享平台上; 二是由于许多竹类物种的建立与分类变动, 其标本信息未及时更新, 如研究发现国产锐药竹属(Oxytenanthera)物种均应归类到巨竹属(Gigantochloa)中(耿伯介, 1984)。标本是植物研究者积累的宝贵财富, 只有得到合理运用才能发挥其最大的价值。国家标本资源共享平台致力于标本数字化和资源共享, 其重要性已逐步显现。首先, 各标本馆在运行过程中应该充分利用这些平台资源, 这不仅能提高标本的使用效率, 也更有利于标本价值的提升; 其次, 标本数字化是一个持续不断的过程, 馆藏量的逐年增加和与时俱进的分类修订, 使得共享平台需要建立一套行之有效的数字化信息管理系统, 以促进标本数字化信息的实时更新。通过这些工作, 才能使新的分类学研究成果及时体现在标本信息上, 促进分类学成果的运用和分类学知识的传播。
杨永(2012)在分析中国植物标本馆馆藏规模时引用了Baum的估算结果, 认为全球高等植物各物种的标本采集量平均为952份, 笔者根据NYBG (
3.2 采集完整度分析
从采集地点偏差评估结果来看, 比值法和斜率法虽然没有得到完全一致的结果, 但也都表明大部分省区采集完整度高。我们认为, 以省级行政区为研究单位虽然较为粗放, 结论稍显粗糙, 但仍可以为将来的竹类标本采集提供一些宏观指导。从采集类群偏差评估结果来看, 采集主要集中在少数几个常见属内, 平均标本数大于10份的属有21个, 仅占所有属的62%, 部分属标本采集量不足, 导致Chao值法和斜率法存在较大误差。对比文献记载数、标本记载数和Chao 1估计量, 我们认为竹类标本采集在属级水平存在明显的偏好性和不完整性。两种方法在采集地点偏差和采集类群偏差的评估中没有得出一致的结论, 从各自的理论依据来看, 两种方法都会受到其他变量的影响, 斜率法容易受到采集物种数和标本数的影响, 而比值法对稀有物种数更加敏感。当标本采集量过少时, 两种方法都存在较大误差, 但在标本采集量充足时, 两种方法得到的结果差异不大。综合两种方法的评估结果可以看出, 国产馆藏竹类植物标本的采集整理工作任重道远。我们需要综合运用两种方法得出的结果, 重点关注采集不完整的区域和属, 并加大采集保藏和鉴定工作的力度。
3.3 适生区模拟分析
基于标本的采集信息, 在最新分子系统学框架的指导下, 我们绘制了国产竹类植物两大分支的分布图, 发现其分布和标本采集都呈现出很强的地域性。结合气候因子进行模型模拟的结果表明, 温度限定了竹类植物两大分支各自的分布北界, 水分条件对温带木本竹类的限制作用更强, 而温度对旧世界热带木本竹类的限制性更强; 对竹类适生区的模拟结果发现, 竹类两大分支在国内的适生区出现了明显的分化, 但仍有部分重叠。值得注意的是, 两大分支在四川盆地和云南南部的适生度都很高, 但在低海拔的盆地或河谷区域, 旧世界热带木本竹类仍然占据主要生态位。四川盆地在纬度范围内处于温带木本竹类的适生区内, 但模拟结果却发现该地是旧世界热带木本竹类和温带木本竹类的适生重叠区, 其盆地的中心位置是旧世界热带木本竹类的适生区, 该结果最早被吴征镒等(1980)在《中国植被》中对中国竹林分区的划分结果中体现, 该研究中单独将四川盆地划为混生竹区。其次云南大部分区域也属于两大分支的适生重叠区, 这与我们野外考察的结论相吻合, 同时也揭示了云南省竹类资源丰富的原因。依据标本数据, 我们得以从更深层次解释宏观层面所观察到的地理分布状况, 或可为其他植物类群的研究提供一种新的模式。
综上所述, 植物标本记录了特定类群在时间和空间地理上的分布格局, 相关的信息一方面可以促进物种灭绝风险评估、可持续利用和综合保护, 另一方面也可助力大尺度生物多样性分布格局及全球变化对多样性的影响研究。
致谢
感谢莫智琼女士在数据分析中提供的帮助;感谢审稿专家提出的宝贵意见!
附录 Supplementary Material
附录1 比值与标本采集量的相关性检测。A: 各省区比值与标本采集量的相关性; B: 竹类各属比值与标本采集量的相关性。
Appendix 1 Correlation analysis of ratio and specimen collections. A, Correlation analysis of ratio and specimen collections at the province level; B, Correlation analysis of ratio and specimen collections at the generic level.
附录2 标本馆索引接受的主要标本馆名称及国际标准代码
Appendix 2 Major herbaria and their standard code, adopted from Index Herbariorum
参考文献
The taxonomic name resolution service: An online tool for automated stan-dardization of plant names
DOI:10.1186/1471-2105-14-16
PMID:23324024
[本文引用: 1]

Background: The digitization of biodiversity data is leading to the widespread application of taxon names that are superfluous, ambiguous or incorrect, resulting in mismatched records and inflated species numbers. The ultimate consequences of misspelled names and bad taxonomy are erroneous scientific conclusions and faulty policy decisions. The lack of tools for correcting this 'names problem' has become a fundamental obstacle to integrating disparate data sources and advancing the progress of biodiversity science.;Results: The TNRS, or Taxonomic Name Resolution Service, is an online application for automated and user-supervised standardization of plant scientific names. The TNRS builds upon and extends existing open-source applications for name parsing and fuzzy matching. Names are standardized against multiple reference taxonomies, including the Missouri Botanical Garden's Tropicos database. Capable of processing thousands of names in a single operation, the TNRS parses and corrects misspelled names and authorities, standardizes variant spellings, and converts nomenclatural synonyms to accepted names. Family names can be included to increase match accuracy and resolve many types of homonyms. Partial matching of higher taxa combined with extraction of annotations, accession numbers and morphospecies allows the TNRS to standardize taxonomy across a broad range of active and legacy datasets.;Conclusions: We show how the TNRS can resolve many forms of taxonomic semantic heterogeneity, correct spelling errors and eliminate spurious names. As a result, the TNRS can aid the integration of disparate biological datasets. Although the TNRS was developed to aid in standardizing plant names, its underlying algorithms and design can be extended to all organisms and nomenclatural codes. The TNRS is accessible via a web interface at http://tnrs.iplantcollaborative.org/ and as a RESTful web service and application programming interface. Source code is available at https://github.com/iPlantCollaborativeOpenSource/TNRS/.
The Herbarium Handbook, 3rd edn), pp. 184-242
标本馆手册(第3版), pp. 184-242
Nonparametric estimation of the number of classes in a population
A hidden species-area curve
DOI:10.1007/s10651-013-0247-2 URL [本文引用: 1]
Diversity and evolution of the new world bamboos (Poaceae: Bambusoideae: Bambuseae, Olyreae)
Potential distribution of wild Camellia oleifera based on ecological niche modeling
DOI:10.17520/biods.2016164 URL [本文引用: 1]
基于生态位模型预测野生油茶的潜在分布
Phylogeny and subfamilial classification of the grasses (Poaceae)
DOI:10.2307/3298585 URL [本文引用: 1]
New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins
DOI:10.1111/j.1469-8137.2011.03972.x URL [本文引用: 1]
Genome sequences provide insights into the reticulate origin and unique traits of woody bamboos
DOI:10.1016/j.molp.2019.05.009 URL [本文引用: 1]
Distributional responses to climate change for alpine species of Cyananthus and Primula endemic to the Himalaya-Hengduan Mountains
DOI:10.1016/j.pld.2019.01.004 URL [本文引用: 1]
Very high resolution interpolated climate surfaces for global land areas
DOI:10.1002/(ISSN)1097-0088 URL [本文引用: 1]
Why bamboos wait so long to flower?
DOI:10.1146/annurev.es.07.110176.002023 URL [本文引用: 1]
Higher level phylogenetic relationships within the bamboos (Poaceae: Bambusoideae) based on five plastid markers
DOI:10.1016/j.ympev.2013.02.005
PMID:23454093
[本文引用: 2]
Bamboos are large perennial grasses of temperate and tropical forests worldwide. Two general growth forms exist: the economically and ecologically important woody bamboos (tribes Arundinarieae and Bambuseae), and the understory herbaceous bamboos (tribe Olyreae). Evolutionary relationships among the 1400+described species have been difficult to resolve with confidence. Comparative analysis of bamboo plastid (chloroplast) DNA has revealed three to five major lineages that show distinct biogeographic distributions. Taxon sampling across tribes and subtribes has been incomplete and most published data sets include a relatively small number of nucleotide characters. Branching order among lineages is often poorly supported, and in more than one study herbaceous bamboos form a clade within the woody bamboos. In this paper, the Bamboo Phylogeny Group presents the most complete phylogeny estimation to date of bamboo tribes and subtribes using 6.7 kb of coding and noncoding sequence data and 37 microstructural characters from the chloroplast genome. Quality of data is assessed, as is the possibility of long branch attraction, the degree of character conflict at key nodes in the tree, and the legitimacy of three alternative hypotheses of relationship. Four major plastid lineages are recognized: temperate woody, paleotropical woody, neotropical woody, and herbaceous bamboos. Woody bamboos are resolved as paraphyletic with respect to Olyreae but SH tests cannot reject monophyly of woody species (Arundinarieae+Bambuseae).Published by Elsevier Inc.
A revision of the genera of bamboos from the world (III)
世界竹亚科各属的考订(之三)
A revision of the genera of bamboos from the world (V)
世界竹亚科各属的考订(之五)
Selecting thresholds of occurrence in the prediction of species distributions
DOI:10.1111/eco.2005.28.issue-3 URL [本文引用: 1]
Applied Logistic Regression Analysis
The bamboos: A fresh perspective
DOI:10.2307/1294068 URL [本文引用: 1]
Hokkaido Teikoku Daigaku Nogakubu Kiyo
Maximum entropy modeling of species geographic distributions
DOI:10.1016/j.ecolmodel.2005.03.026 URL [本文引用: 1]
A 250 plastome phylogeny of the grass family (Poaceae): Topological support under different data partitions
DOI:10.7717/peerj.4299 URL [本文引用: 1]
A guide to collecting bamboos
DOI:10.2307/2399010 URL [本文引用: 1]
A worldwide phylogenetic classification of the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications
DOI:10.1111/jse.12262 URL [本文引用: 1]
Sarocalamus, a new Sino-Himalayan bamboo genus (Poaceae-Bambusoideae)
Global patterns and predictors of marine biodiversity across taxa
DOI:10.1038/nature09329 URL [本文引用: 1]
Analysis of geographical deviation of record collection of Rosaceae plant specimens in national specimen resource sharing platform
国家标本资源共享平台蔷薇科植物标本记录采集地理偏差分析
Statistical analyses of vascular plant specimen data from the herbarium of South China Botanical Garden, Chinese Academy of Sciences (IBSC)
中国科学院华南植物园标本馆维管植物标本数据统计分析
Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models
DOI:10.1360/biodiv.060280 URL [本文引用: 1]
ROC曲线分析在评价入侵物种分布模型中的应用
Four new genera of bamboos (Gramineae: Bambusoideae) from Malesia
DOI:10.2307/4118719 URL [本文引用: 1]
Geographical Sampling Bias in the Collections of Chinese Plants and Its Impacts on the Analysis of Biodiversity Patterns
中国植物采集的地理偏差及其对生物多样性格局分析的影响
Holdings of type specimens of plants in herbaria of China
DOI:10.3724/SP.J.1003.2012.05076 URL [本文引用: 1]
我国植物模式标本的馆藏量
Genome skimming herbarium specimens for DNA barcoding and phylogenomics
DOI:10.1186/s13007-017-0271-6 URL [本文引用: 1]
A new genus of temperate woody bamboos (Poaceae, Bambusoideae, Arundinarieae) from a limestone montane area of China
DOI:10.3897/phytokeys.109.27566 URL [本文引用: 1]
The analysis of geographical bias of the collection of Chinese Rhododendraceae plants based on the data of National Specimen Information Infratructure
基于国家标本资源共享平台数据的中国杜鹃花科植物标本采集地理偏差分析

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}