


叶和根是植物地上和地下进行物质分配和交换的重要场所。其中叶片是植物地上部分进行光合作用的关键部位, 而比叶面积、叶干物质含量作为重要的叶功能性状, 在一定程度上反映了叶片对环境的适应性和获取资源的能力(Wright et al, 2004)。对热带森林木本植物功能性状的研究发现, 比叶面积与树高呈正相关, 而与叶干物含量呈负相关(Wright et al, 2007)。细根是植物地下根系与土壤产生交互作用的重要器官, 其功能性状如比根长和根平均直径能体现根系对营养资源的吸收和运输能力(Holdaway et al, 2011)。地中海地区的木本植物细根功能性状的研究表明, 比根长与比叶面积呈正相关, 而与根平均直径呈负相关(De la Riva et al, 2018)。因此, 在一些特定环境和地区植物的根叶功能性状会通过权衡关系或协同效应来适应环境变化, 使植物采取相应的生态策略来维持生长。虽然目前有关不同植物根叶功能性状差异的研究有很多, 但对不同菌根类型的根叶功能性状的特征仍少有研究。
菌根是植物根系与土壤中的真菌相结合形成的一种共生体, 一方面菌根真菌通过根外菌丝从土壤中获取营养物质供给植物生长; 另一方面植物根系可以提供菌根真菌生长所需的碳水化合物或根系分泌物等有机物质(Ekblad et al, 2013)。菌根类型主要以外生菌根(ectomycorrhiza, EM)和丛枝菌根(arbuscular mycorrhiza, AM)为主, 其中EM真菌会在植物根系的表面形成外延菌丝和菌套以及在根系皮层细胞间形成哈蒂氏网等, 其中哈蒂氏网和菌套具有固定土壤水分和养分的作用(Ekblad et al, 2013); 外延菌丝能在土壤中形成菌丝网, 使植物根系的吸收面积显著增加, 以此提高植物根系获取资源的能力。有研究表明未被EM菌丝侵染的根呈细长状态, 主要依靠根系生长获取养分。但随着EM菌丝侵染程度和包裹的加强, 吸收根的根长生长会受到限制(Taylor & Alexander, 2005; Segnitz et al, 2020)。AM真菌则主要通过侵染根的皮层细胞而在胞内形成丛枝或泡囊结构, 使其根系直径大、皮层较厚, 因此对根系伸长生长的影响较小, 且植物的根系直径会受到AM真菌侵染的影响而改变, 从而影响养分运输(Kiers et al, 2011; Comas et al, 2014)。由于不同菌根真菌侵染会导致根系形态产生差异, 使吸收根的功能性状发生变异, 最终影响菌根的吸收效率和植物获取资源的策略。
植物功能性状也存在种内变异, 通常由环境异质性和遗传变异共同作用所致(Albert et al, 2010)。以往大多数关于植物功能性状的研究主要以种间的性状变异为主, 而忽略由环境因素等导致的种内变异, 然而也有研究发现在某些环境下植物功能性状的种内变异程度与种间相等(Jung et al, 2010; Messier et al, 2010)。所以对不同菌根类型的树种而言, 其功能性状的种内与种间变异如何对研究菌根真菌与植物功能性状之间的关系有所帮助。本文以江西德兴新岗山亚热带森林生物多样性与生态系统功能实验研究平台中相同林龄相同密度(林龄5年, 密度均为400株/亩)的14个亚热带常见树种纯林为研究对象, 利用微根管及相应的图像分析等方法获取14个树种的根叶功能性状及地上地下生长指标数据, 拟探讨如下问题: (1)不同菌根类型树种间根叶功能性状及生长指标的差异?(2)不同菌根类型树种根叶功能性状及生长指标的种间与种内变异有何异同?(3)驱动不同菌根类型树种生长的关键根叶功能性状有何异同?
1 材料与方法
1.1 研究区域概况
研究区位于江西省德兴市新岗山镇(29°08°- 29°11° N, 117°90°-117°93° E)的中国亚热带森林生物多样性与生态系统功能实验研究平台(简称BEF-China)。BEF-China样地A的年平均气温和年降水量分别为16.7℃和1,821 mm, 海拔范围为105-275 m, 坡度为0-45°, 属丘陵地带。自然植被以亚热带常绿落叶阔叶混交林为主。样地所在区域是典型的亚热带季风气候, 红黄壤是该地区主要的土壤类型(马克平,2013)。
1.2 实验设计
样地A于2009年栽植一年生实生苗, 以1亩(25.82 m × 25.82 m)为基本单元样方, 共计271个样方, 占地约18 ha。1亩的基本样方中包括16个亚热带常见树种单一种植样方(即纯林), 其中EM树种和AM树种各8种。其中盐肤木( Rhus chinensis)的根系属于串根类型, 根系取样后容易死亡, 而甜槠(Castanopsis eyrei)的根系容易遭受蚂蚁、老鼠等动物啃食而导致植株大量死亡, 因而本研究上述两个物种均未取样。本研究共选取了7个EM树种和7个AM树种为研究对象(表1)。
表1 本研究选取的14个树种的基本特征
Table 1
| 树种 Tree species | 菌根类型 Mycorrhiza type | |
|---|---|---|
| 白栎 | Quercus fabri | 外生菌根 EM |
| 青冈 | Cyclobalanopsis glauca | 外生菌根 EM |
| 短柄枹栎 | Quercus serrata | 外生菌根 EM |
| 苦槠 | Castanopsis sclerophylla | 外生菌根 EM |
| 细叶青冈 | Cyclobalanopsis myrsinifolia | 外生菌根 EM |
| 石栎 | Lithocarpus glaber | 外生菌根 EM |
| 锥栗 | Castanea henryi | 外生菌根 EM |
| 无患子 | Sapindus saponaria | 丛枝菌根 AM |
| 枫香 | Liquidambar formosana | 丛枝菌根 AM |
| 复羽叶栾 | Koelreuteria bipinnata | 丛枝菌根 AM |
| 乌桕 | Triadica sebifera | 丛枝菌根 AM |
| 蓝果树 | Nyssa sinensis | 丛枝菌根 AM |
| 木荷 | Schima superba | 丛枝菌根 AM |
| 南酸枣 | Choerospondias axillaris | 丛枝菌根 AM |
EM: Ectomycorrhiza; AM: Arbuscular mycorrhiza
1.3 根叶功能性状及生长指标的测定方法
2014年5月, 在14个单种样方的中心以梅花五点法将微根管均匀设置在两棵树之间, 即每个样方中设置5根微根管, 共70根。微根管内径为7 cm, 长度为90 cm, 安装的角度与地面成30°, 插入长度约为70-75 cm (垂直深度35-45 cm左右)。半年后用BTC-100微根管根系生态监测系统对每根微根管四个面的根系生长过程进行影像收集, 每根微根管每个侧面收集约40-45张图片, 总共收集了11,760张图片。2014年9月, 在每一个单种样方中随机选取除微根管监测植株之外的8棵植株进行根叶样品采样, 共计112株植株。每棵植株选取当年生向阳面的健康成熟叶片约10-50片, 用CanoScan图像扫描仪对采集叶片进行扫描, 再用ImageJ图像处理软件测量扫描图像的叶面积, 用千分位电子天平称叶片鲜重, 随后烘干至恒重称重, 计算叶片的比叶面积和叶干物质含量。比叶面积(SLA, cm²/g)是叶面积与叶干重的比值, 叶干物质含量(LDMC, g/g)是叶干重与叶鲜重的比值。此外, 每棵植株的地径统一采用胸径尺在距地面5 cm以上的位置测定其周长后换算而成(精确到0.01 cm); 而树高则使用5 m长的测量杆和钢卷尺对每一株植株进行准确测量(精确到0.1 m)。其中树高生长速率为各树种的平均树高与林龄(5年, 即2009-2014)的比值, 地径生长速率为各树种平均地径与林龄(同上)的比值。
根系样品的采样方法是: 采用主根溯源法在目标植株的根部挖取3-5条完整的根系, 装入自封袋内, 放入便携式冷藏箱内带回实验室。使用去离子水浸泡后, 小心去除根系上的泥土和沙石等。洗净后的根系按根序级别进行分级: 最细的根划分为1级根, 2个1级根交叉后的根系划分为2级根, 以此类推一直延伸到5级根, 其中1-3级根为吸收根, 4-5级根为运输根(Pregitzer et al, 2002)。用WinRHIZO根系扫描仪对分级后的根系进行扫描, 然后利用自带软件进行图片分析, 获取吸收根和运输根的根长和平均直径, 同时将扫描后的根系样品烘干至恒重称重, 获得分级的根系干重。比根长(SRL, m/g)是根长与根系干重的比值(刘娜娜等, 2014)。同时, 利用WinRHIZO TRON MF图像处理软件对微根管图片进行分析处理后获取根长和平均直径, 通过比对采样的根系平均直径与微根管图片软件获取的根系平均直径数据, 发现微根管监测的根系平均直径与实际采样的吸收根根系平均直径相关性很高(r= 0.88), 因此采用实际采样的吸收根比根长来计算细根生物量。根系生物量(RBDv, g/m3)通过单位土壤体积的根长密度(RLDv,m/m3)与比根长(SRL, m/g)来计算(字洪标等, 2018):
式中, L表示通过微根管图片分析得到的细根根长(m); A为所分析的微根管图片面积(m2); DOF为微根管到周围土壤的距离, 一般范围为0.2-0.3 cm, 由于研究中的目标树种细根直径较小, 因此本研究的计算中DOF取0.25 cm。
1.4 数据处理与分析
1.4.1 不同菌根类型树种根叶功能性状及生长指标的差异
利用二级因子方差分析和多重比较对不同树种与菌根真菌类型间的叶片功能性状、地上地下生长指标进行差异显著性检验; 同时利用三因素方差分析对不同菌根树种之间的不同根序级别的根系功能性状进行差异显著性分析。
1.4.2 不同菌根类型树种根叶功能性状及生长指标的种内和种间变异
运用混合线性模型对不同树种根叶功能性状及生长指标的方差进行分解。对每个指标构建只有截距的随机效应模型(树种作为随机效应), 将各指标的变异分解为种内变异和种间变异, 按照以下公式计算(Auger & Shipley, 2013):
其中, yij是第i个树种的第j个个体的根叶功能性状或地上地下生长指标值, μ i是解释树种水平根叶功能性状及地上地下生长指标变异的随机方差项, ε ij是残差。
1.4.3 根叶功能性状与不同菌根类型树种生长的关系
为使混合线性模型中的变量接近正态分布, 对每个树种的根叶功能性状及地上地下生长数据进行对数转换, 以不同树种的根叶功能性状指标为固定效应, 树种为随机效应, 用生长指标作为因变量来构建不同菌根树种的混合线性模型, 对所有模型的边际决定系数(marginal R2)和条件决定系数(conditional R2)进行估计, 前者表示仅由固定效应解释的方差, 后者为固定效应和随机效应共同解释的方差(Nakagawa & Schielzeth, 2013)。
本研究利用Excel 2016和R 3.5.3完成数据的统计分析与绘图, 其中数据分析主要使用R软件中的lme4、lmerTest和ggplot2包等完成(Nakagawa & Schielzeth, 2013), 显著性水平P = 0.05。
2 结果
2.1 不同菌根树种的叶片功能性状变异
从表2可知, 树种和菌根类型及其交互作用对比叶面积和叶干物质含量均有显著影响。多重比较结果表明, 南酸枣的比叶面积(185.67 cm²/g)显著高于其他13个树种; 苦槠的比叶面积最低(65.04 cm²/g), 与白栎、青冈、短柄枹栎、细叶青冈和石栎间无显著差异, 但显著低于其余8个树种(图1A)。白栎和青冈的叶干物质含量最高(均为0.54 g/g), 与苦槠、细叶青冈和石栎之间无明显差异, 却显著高于其余9个树种; 南酸枣的叶干物质含量最低(0.36 g/g), 与其余13个树种均存在显著差异(图1B)。总体上, EM树种的比叶面积显著地低于AM树种, 但其叶干物质含量显著地高于AM树种(图2A, D)。
表2 树种和菌根类型对叶功能性状及生长指标影响的双因素方差分析(F值)
Table 2
| 叶功能性状及生长指标 Leaf functional traits and growth indices | 树种 Tree species | 菌根类型 Mycorrhizal type | 树种 × 菌根类型 Tree species × mycorrhizal type |
|---|---|---|---|
| 比叶面积 Specific leaf area | 12.03*** | 113.47*** | 30.23*** |
| 叶干物质含量 Leaf dry matter content | 32.04*** | 155.43*** | 81.65*** |
| 树高生长速率 The growth rate of height | 21.21*** | 29.37*** | 26.77*** |
| 地径生长速率 The growth rate of basal diameter | 6.27*** | 32.78*** | 9.74*** |
| 细根生物量 Fine root biomass | 29.85*** | 3.78* | 28.75*** |
*P<0.05; ***P<0.001。
图1
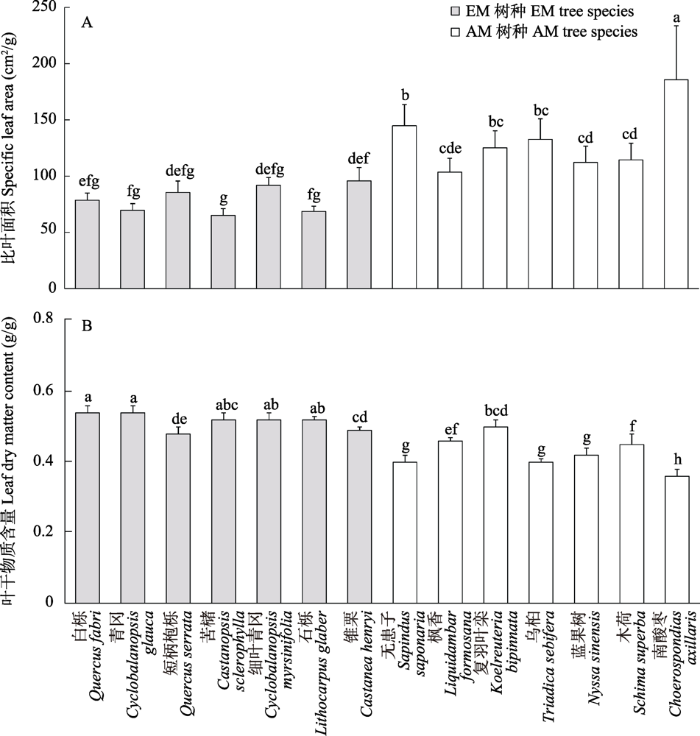
图1
14个树种的叶片功能性状变异。不同小写字母表示不同树种间的叶功能性状差异显著(P < 0.05)。
Fig. 1
Variations in leaf functional traits with 14 tree species. Different lowercase letters represent significant differences between different tree species for leaf functional traits at P< 0.05.
图2
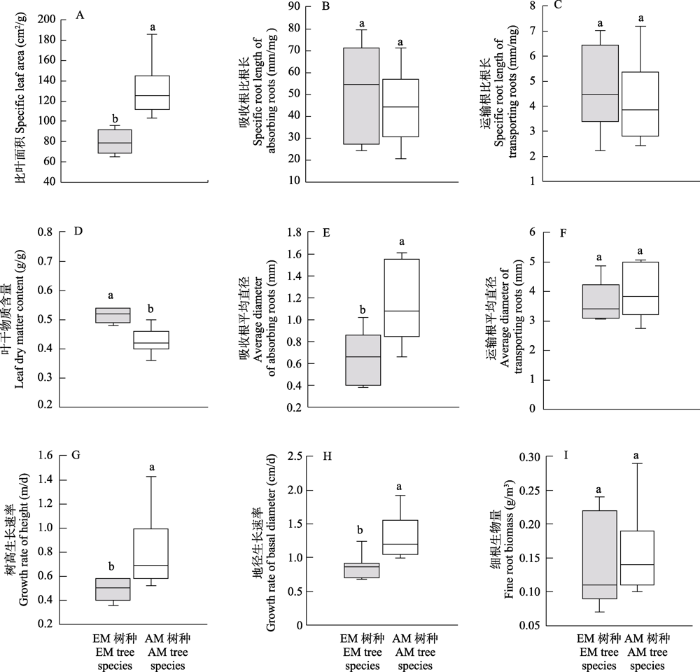
图2
不同菌根树种之间叶片和根功能性状及生长指标的差异。不同小写字母表示不同树种间的叶和根功能性状及生长指标差异显著(P < 0.05)。
Fig. 2
Variations in leaf and root functional traits, above- and belowground growth indices with different mycorrhizal tree species, different lowercase letters represent significant differences of leaf and root functional traits, above- and belowground growth indices between different tree species at P< 0.05.
2.2 不同菌根树种的根系功能性状变异
树种、根功能类型、菌根类型及其交互作用对比根长和平均直径均有显著影响(表3)。短柄枹栎的吸收根比根长最大(79.49 m/g), 与除白栎、石栎、锥栗、无患子、复羽叶栾外的8个树种有显著差异; 复羽叶栾的运输根比根长最大(7.2 m/g), 显著地高于苦槠、乌桕和南酸枣; 乌桕的吸收根和运输根比根长均最小(分别为20.68 m/g和2.43 m/g) (图3A)。蓝果树的吸收根平均直径最大(1.61 mm), 与除无患子、乌桕、南酸枣外的10个树种之间存在显著差异, 锥栗的吸收根平均直径最小(0.38 mm); 乌桕的运输根平均直径最大(5.07 mm), 显著地大于除南酸枣、青冈、苦槠、蓝果树、木荷外的其余8个树种, 而复羽叶栾的运输根平均直径最小(2.76 mm) (图3B)。此外, 相同树种吸收根的比根长均显著高于运输根, 而平均直径则正好相反。总体上, EM树种吸收根和运输根的比根长平均值大于AM树种, 但两类树种间并无显著差异(图2B, C), 仅AM树种吸收根的平均直径显著地大于EM树种(图2E, F)。
表3 树种、根功能类型和菌根类型对根功能性状影响的多因素方差分析(F值)
Table 3
| 因素类型 Factor type | 比根长 Specific root length | 平均直径 Average diameter |
|---|---|---|
| 树种 Tree species | 10.80*** | 14.05*** |
| 根功能类型 Root functional type | 683.40*** | 1,462.96*** |
| 菌根类型 Mycorrhizal type | 2.06* | 3.37* |
| 树种 × 根功能类型 Tree species × root functional type | 15.12** | 79.10*** |
| 树种 × 菌根类型 Tree species × mycorrhizal type | 2.54** | 1.91* |
| 根功能类型 × 菌根类型 Root functional type × mycorrhizal type | 156.42*** | 293.21*** |
| 树种 × 根功能类型 × 菌根类型 Tree species × root functional type × mycorrhizal type | 64.47*** | 79.11*** |
*P< 0.05; **P< 0.01; ***P< 0.001。
图3
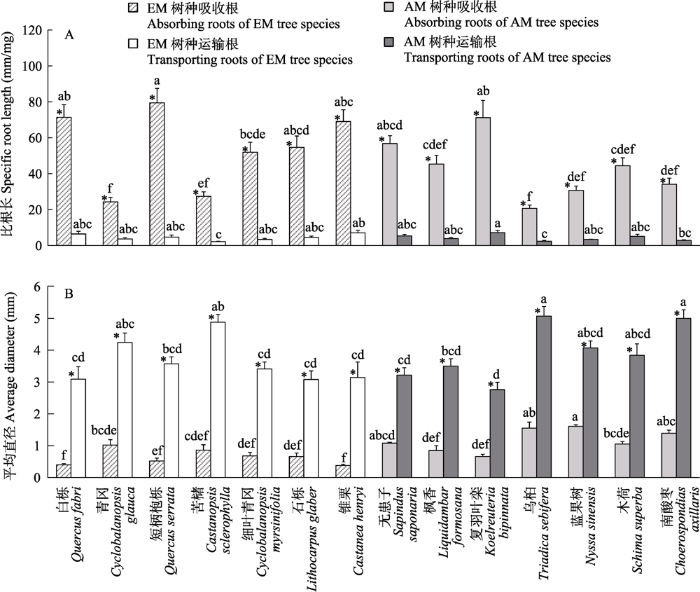
图3
14个树种的根功能性状变异。不同小写字母表示不同树种间的根功能性状差异显著(P < 0.05), *表示同一树种不同功能类型的根功能性状差异显著(P < 0.05)。
Fig. 3
Variations in root traits with 14 tree species. Different lowercase letters represent significant differences of root traits between different tree species at P< 0.05. * Significant differences between different root function types of the same tree species atP< 0.05.
2.3 不同菌根树种的地上地下生长指标的差异
树高生长速率、地径生长速率、细根生物量在树种和菌根类型及其交互作用中有明显差异(表2)。南酸枣的树高生长速率最大(1.43), 显著大于其余13个树种, 而白栎的树高生长速率最小(0.36)(图4A); 地径生长速率最大为南酸枣(1.92), 显著大于除木荷外的其他12个树种, 短柄枹栎的地径生长速率最小(0.68), 显著小于锥栗、蓝果树、木荷、南酸枣(图4B)。乌桕具有最高的细根生物量(0.29 g/m³), 与除青冈外的12个树种有显著差异; 短柄枹栎最低(0.07 g/m³), 显著低于除白栎、细叶青冈、石栎、锥栗、无患子、复羽叶栾外的其余7个树种(图4C)。总体上, AM树种的树高和地径生长速率显著地高于EM树种, 但细根生物量在不同菌根类型的树种间无显著差异(图2G, H, I)。
图4
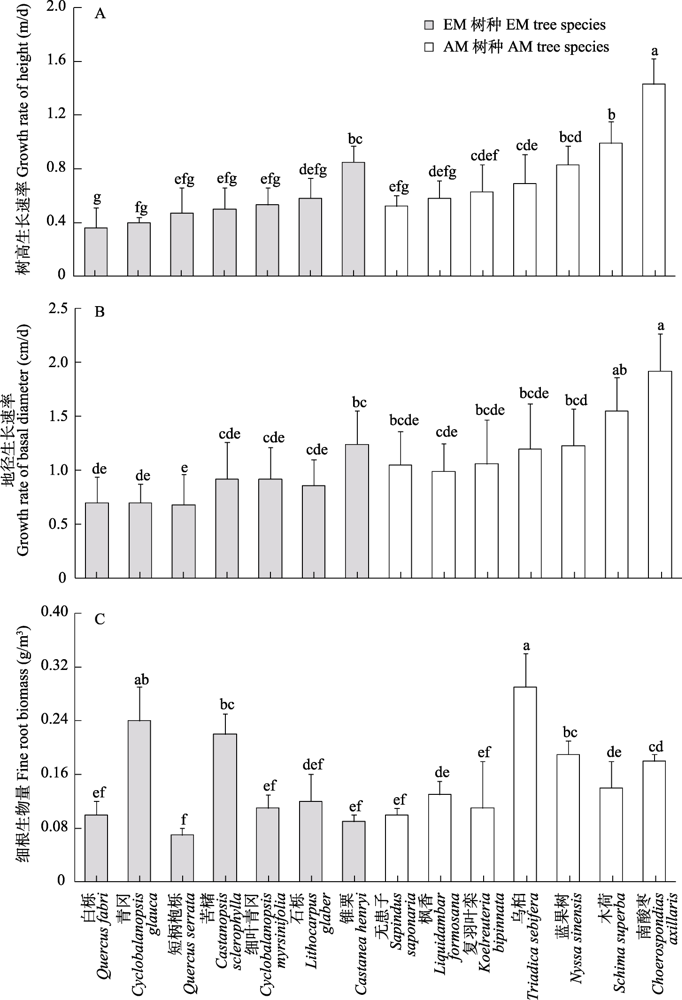
图4
14个树种地上地下生长指标的差异。不同小写字母表示不同树种间差异显著(P < 0.05)。
Fig. 4
Variations in above- and belowground growth indices of 14 tree species. Different lowercase letters represent significant differences of above- and belowground growth indices between different tree species at P< 0.05.
2.4 不同菌根树种根叶功能性状及生长指标的种内和种间变异
对于EM树种来说, 叶片功能性状及地上生长指标的种间变异均小于种内变异(图5), 其中地径生长速率(种间变异和种内变异分别为0.03和0.97)的种内变异在叶片功能性状及地上生长指标中最大, 其次是叶干物质含量(分别为0.27和0.73); 吸收根和运输根的根功能性状种间变异均小于种内变异, 最后细根生物量的种间变异(0.89)大于种内变异(0.11)。对于AM树种来说, 比叶面积和地径增长速率的种间变异均小于种内变异, 而叶干物质含量和树高生长速率则相反; 除吸收根的平均直径(种间和种内变异分别为0.53和0.47)和细根生物量(种间和种内分别为0.97和0.03)外, 其余根功能性状的种间变异均小于种内变异。总之, 除细根生物量外, EM树种的其他根叶功能性状及生长指标的种间变异均小于种内变异; 而AM树种的叶片干物质含量、吸收根平均直径、树高生长速率及细根生物量的种间变异大于种内变异。
图5
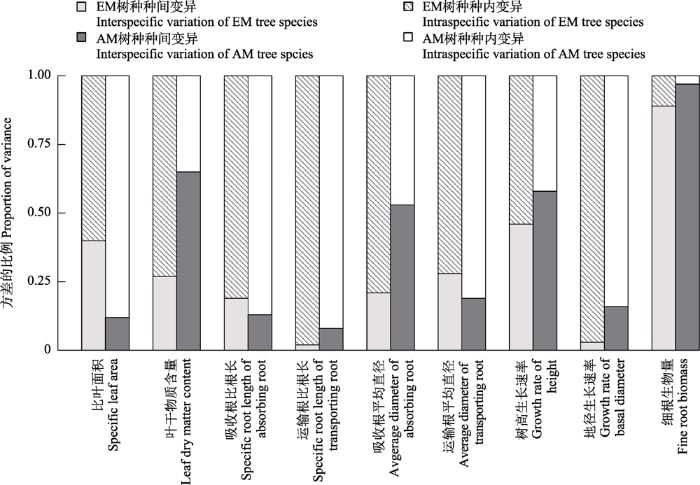
图5
不同菌根树种根叶功能性状及生长指标的变异来源
Fig. 5
Sources of variations in leaf and root functional traits, above- and belowground growth indices of different mycorrhizal type tree species
2.5 根叶功能性状与不同菌根树种生长的关系
对于EM树种来说, 树高生长速率与叶干物质含量呈显著负相关, 与运输根平均直径呈显著正相关, 二者的固定效应解释了67%的变异(表4); 而地径生长速率则只与运输根平均直径呈显著正相关, 固定效应解释了96%的变异; 二者随机效应(树种)只解释了很小的变异。细根生物量仅与吸收根比根长呈显著负相关, 固定效应解释了总变异的35%, 而树种解释了58%的变异。
表4 根叶功能性状与不同菌根树种生长关系的混合线性模型
Table 4
| 随机效应 Random effects | 因变量 Dependent variables | 截距 Intercept | 固定效应 Fixed effects | 条件决定 系数 Conditional R2 | 边际决定 系数 Marginal R2 | ||
|---|---|---|---|---|---|---|---|
| 叶干物质含量 LDMC | 吸收根比根长 SRL of absorbing root | 运输根平均直径 Average diameter of transporting root | |||||
| EM树种 Ectomycorrhizal tree | 树高生长速率 Growth rate of height | ‒0.23 | ‒0.11 | - | 0.40 | 0.69 | 0.67 |
| 地径生长速率 Growth rate of basal diameter | 0.29 | - | - | 0.30 | 0.97 | 0.96 | |
| AM树种 Arbuscular mycorrhizal tree | 细根生物量 Fine root biomass | ‒0.12 | - | ‒0.21 | - | 0.93 | 0.35 |
| 树高生长速率 Growth rate of height | 0.22 | ‒0.21 | 0.09 | - | 0.65 | 0.25 | |
| 地径生长速率 Growth rate of basal diameter | 0.56 | ‒0.13 | 0.07 | - | 0.44 | 0.24 | |
| 细根生物量 Fine root biomass | ‒0.21 | ‒0.18 | 0.94 | 0.34 | |||
每个因变量的样本量均为56 (n = 56), 条件决定系数减去边际决定系数即为菌根树种(随机效应)单独解释的变异量。
Dependent variables n = 56, variations (conditional R2-marginal R2) are explained individually by tree species with different mycorrhizal fungi types. LDMC: Leaf dry matter content, SRL: Specific root length.
AM树种的树高生长速率和地径生长速率均与叶干物质含量呈显著负相关, 与吸收根比根长呈显著正相关, 二者的固定效应分别解释了25%和24%的变异, 随机效应(树种)分别解释了40%和20%的变异。细根生物量与吸收根比根长呈显著负相关, 并解释了34%的变异, 树种解释了60%的变异。总之, EM树种的地上生长主要受运输根平均直径的影响; 而AM树种的地上生长则主要受叶片干物质含量和吸收根比根长的影响。吸收根比根长会影响两种菌根树种的地下生长, 但树种类型对两种菌根地下生长的影响更大。
3 讨论
3.1 不同菌根树种的根叶功能性状差异
本研究结果显示, 与EM树种对比, AM树种的比叶面积高、叶干物质含量低, 这与全球不同纬度区域下(包括北方、温带和寒带) 2,940个不同菌根树种的根叶功能性状研究结果相似(Averill et al, 2019)。由叶经济型谱性状可知比叶面积高且叶干物质含量低的树种拥有高的净光合速率, 使叶片能够快速获取养分, 有利于提高植物地上部分的生产力, 使树高、地径生长速率显著增加, 让植物在适应环境的同时也能保持较强的资源获取能力(Shipley, 2002; 唐青青等, 2016)。此外, 由于本研究中AM树种绝大部分是落叶树种, 叶片生长倾向于获取性策略(Kloeke et al, 2012)。其次, AM树种的根系平均直径显著大于EM树种, 这与帽儿山天然次生林20个不同阔叶树种细根形态和美国东北部33个木本树种根系特征的研究结果一致(师伟等, 2008; Comas et al, 2014), 原因可能与菌根类型有关。菌根真菌是植物根系获取资源的主要来源, 而AM树种获取资源主要通过AM真菌侵染根的皮层细胞, 在细胞内形成丛枝或泡囊, 使根平均直径增加、皮层增厚, 以此提高菌根的吸收效率。此外, AM树种在未受到菌根真菌侵染前, 主要依靠根毛来获取资源, 但由于根毛较粗且伸长距离有限, 导致其根系养分利用效率较低、生长速率缓慢, 根长较短(Wells & Eissenstat, 2002)。
对EM树种而言, 其比叶面积低于AM树种, 但叶干物质含量较高。EM树种光合作用所获取的资源多用于叶片的构建, 以提高叶片对外界环境的耐受性, 但用于植物生长的养分却较少, 导致EM树种的树高、地径生长速率显著低于AM树种(Wright et al, 2005; Reich, 2014)。而EM树种的根平均直径小于AM树种, 是由于EM真菌侵染后会在根尖形成菌套, 使吸收根生长受到限制, 依靠外延菌丝来增加根系的吸收面积和范围, 从而提高植物根系的吸收效率(Taylor & Alexander, 2005)。此外, EM真菌仅包裹植物根尖, 不影响皮层厚度, 因此不需要投入更多成本用于构建较粗的直径(Druebert et al, 2009), 即以较少的细根生物量投入换取更大的根系吸收面积。综上所述, AM树种叶片通常采取获取性策略, 根系偏向保守型策略; 但EM树种却截然相反。
3.2 不同菌根树种的根叶功能性状及生长指标的种内和种间变异
本研究发现, EM树种的根叶功能性状及地上生长指标的种内变异大于种间变异, 这可能与趋同效应有关。它是系统演化过程中的一种机制, 能通过植物个体间相互影响来进行相同或相近的活动(Twieg et al, 2010)。种间变异小说明物种本身的作用比较小, 可能会弱化树种特性所带来的影响(Prinzing et al, 2001); 而种内变异大说明植物主要通过形成同种个体特征的差异, 利用趋异性的生态策略来降低竞争强度、加强对环境的适应能力, 使具有相似功能性状的植物个体能在局域群落中相互共存(Fordyce, 2006)。其次, AM树种地上指标的种内和种间变异程度类似。一方面, 由于本研究中AM树种为不同科属的植物, 它们与AM真菌所建立的共生关系具有特异性, 使AM真菌的功能、种类和数量发生变化(Fitter et al, 2005), 导致AM真菌促进不同植物获取资源的程度有所差距, 最终对植物的生理形态特征造成影响, 发生种间变异(Laforest-Lapointe et al, 2014)。另一方面, 环境异质性是发生种内变异的主要原因。由于不同植物个体在同一环境下所受到光照强度和时间不同会影响植物的碳素代谢, 从而影响其AM真菌群落的形成与发育, 导致AM真菌对不同植物养分吸收的促进作用有所差异(Heijden et al, 2003)。
AM和EM树种细根生物量的种间变异大于种内变异, 主要是根系的吸收效率不同所致。而菌根真菌类型和侵染程度是影响吸收效率的主要原因(Cornelissen et al, 2001), 其中EM树种主要依靠根系表面形成的根外菌丝来拓展获取资源的范围, 而AM真菌会侵入根的皮层细胞, 使根直径增加, 同时还需要根毛与土壤接触来获取养分。另一方面, 细根具有很强的可塑性, 不同菌根树种的细根数量和形态特征在相同环境下会发生相应变化, 而细根的数量和形态等方面的性状特征又决定了植物对资源的吸收和利用能力(Bu et al, 2017)。因此, 不同菌根树种细根数量的差异能够影响其根系对土壤资源的利用及植物与土壤交互过程中的养分循环, 使其在资源利用效率和养分循环效率上产生差异。
3.3 根叶功能性状与不同菌根类型树种生长关系
本研究中叶干物质含量越低、运输根平均直径越大, EM树种地上部分的生长速率越快。EM树种会通过外延菌丝扩张来增加根系的吸收面积以获取土壤中的水分和养分, 这种特殊的资源获取方式能保证植物高效地获取资源, 导致吸收根在获取水分和养分上的重要性下降(Twieg et al, 2010), 使EM树种更依赖菌根真菌来获取资源。为了能将更多资源运输到地上部分, 增加运输根直径能显著提高根系的运输能力, 而降低叶干物质含量能保证EM菌丝和光合作用产生的养分多用于植物生长, 减少对叶片建成的投入, 促使植物树高和地径生长速率显著增加(Segnitz et al, 2020)。而对于AM树种来说, 叶干物质含量越低、吸收根比根长越大, 越能促进其地上生长。一方面是因为叶干物质含量低能保证获取的资源都投入植物生长(Díazet al, 2004)。另一方面, 虽然AM真菌侵染能使根系运输能力增强, 但因为AM树种获取资源还需与根毛接触, 而根毛较粗且伸长有限, 会使植物生长受到限制, 因此需要延长吸收根的根长以获取更多资源, 再将养分更好地运输到地上部分, 以保证树高和地径生长(Turrini et al, 2018)。此外, 对于两种菌根树种来说吸收根比根长越小, 细根生物量越大, 这可能反映了植物在生长策略上的权衡过程。吸收根比根长越小说明根系吸收水分和养分的能力较弱, 因此需要通过分配更多碳资源用于产生更多细根, 增加细根生物量, 满足植物生长与繁殖的需要(Westoby et al, 2002)。
参考文献
A multi-trait approach reveals the structure and the relative importance of intra- versus interspecific variability in plant traits
DOI:10.1111/j.1365-2435.2010.01727.x URL [本文引用: 1]
Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest
DOI:10.1111/jvs.2013.24.issue-3 URL [本文引用: 1]
Global imprint of mycorrhizal fungi on whole-plant nutrient economics
Interspecific and intraspecific variation in specific root length drives aboveground biodiversity effects in young experimental forest stands
DOI:10.1093/jpe/rtw096 URL [本文引用: 1]
Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: Implications for the evolution of belowground strategies
DOI:10.1002/ece3.2014.4.issue-15 URL [本文引用: 2]
Carbon cycling traits of plant species are linked with mycorrhizal strategy
DOI:10.1007/s004420100752
PMID:24577702
[本文引用: 1]

Ecosystem carbon cycling depends strongly on the productivity of plant species and the decomposition rates of the litter they produce. We tested the hypothesis that classifying plant functional types according to mycorrhizal association explains important interspecific variation in plant carbon cycling traits, particularly in those traits that feature in a hypothesized feedback between vegetation productivity and litter turnover. We compared data from standardized 'screening' tests on inherent potential seedling relative growth rate (RGR), foliar nutrient concentrations, and leaf litter decomposability among 83 British plant species of known mycorrhizal type. There was important variation in these parameters between mycorrhizal plant types. Plant species with ericoid mycorrhiza showed consistently low inherent RGR, low foliar N and P concentrations, and poor litter decomposability; plant species with ectomycorrhiza had an intermediate RGR, higher foliar N and P, and intermediate to poor litter decomposability; plant species with arbuscular-mycorrhiza showed comparatively high RGR, high foliar N and P, and fast litter decomposition. Within the woody species subset, differentiation in RGR between mycorrhizal types was mostly confounded with deciduous versus evergreen habit, but the overall differentiation in litter mass loss between mycorrhizal types remained strong within each leaf habit. These results indicate that, within a representative subset of a temperate flora, ericoid and ectomycorrhizal strategies are linked with low and arbuscular-mycorrhizal species with high ecosystem carbon turnover. The incorporation of mycorrhizal association into current functional type classifications is a valuable tool in the assessment of plant-mediated controls on carbon and nutrient cycling.
Root traits across environmental gradients in Mediterranean woody communities: Are they aligned along the root economics spectrum?
DOI:10.1007/s11104-017-3433-4 URL [本文引用: 1]
The plant traits that drive ecosystems: Evidence from three continents
DOI:10.1658/1100-9233(2004)015[0295:TPTTDE]2.0.CO;2 URL [本文引用: 1]
Beech carbon productivity as driver of ectomycorrhizal abundance and diversity
The production and turnover of extramatrical mycelium of ectomycorrhizal fungi in forest soils: Role in carbon cycling
DOI:10.1007/s11104-013-1630-3 URL [本文引用: 2]
Biodiversity and ecosystem function in soil
DOI:10.1111/fec.2005.19.issue-3 URL [本文引用: 1]
The evolutionary consequences of ecological interactions mediated through phenotypic plasticity
DOI:10.1242/jeb.02271 URL [本文引用: 1]
Linking above- and belowground traits to soil and climate variables: An integrated database on China's grassland species
Different arbuscular mycorrhizal fungi alter coexistence and resource distribution between co-occurring plant
DOI:10.1046/j.1469-8137.2003.00688.x URL [本文引用: 1]
Species- and community-level patterns in fine root traits along a 120000-year soil chronosequence in temperate rain forest
DOI:10.1111/jec.2011.99.issue-4 URL [本文引用: 1]
Intraspecific variability and trait-based community assembly
DOI:10.1111/jec.2010.98.issue-5 URL [本文引用: 1]
Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis
DOI:10.1126/science.1208473 URL [本文引用: 1]
Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions
DOI:10.1111/j.1466-8238.2011.00667.x URL [本文引用: 1]
Intraspecific variability in functional traits matters: Case study of Scots pine
DOI:10.1007/s00442-014-2967-x
PMID:24850418
[本文引用: 1]
Although intraspecific trait variability is an important component of species ecological plasticity and niche breadth, its implications for community and functional ecology have not been thoroughly explored. We characterized the intraspecific functional trait variability of Scots pine (Pinus sylvestris) in Catalonia (NE Spain) in order to (1) compare it to the interspecific trait variability of trees in the same region, (2) explore the relationships among functional traits and the relationships between them and stand and climatic variables, and (3) study the role of functional trait variability as a determinant of radial growth. We considered five traits: wood density (WD), maximum tree height (H max), leaf nitrogen content (Nmass), specific leaf area (SLA), and leaf biomass-to-sapwood area ratio (B L:A S). A unique dataset was obtained from the Ecological and Forest Inventory of Catalonia (IEFC), including data from 406 plots. Intraspecific trait variation was substantial for all traits, with coefficients of variation ranging between 8% for WD and 24% for B L:A S. In some cases, correlations among functional traits differed from those reported across species (e.g., H max and WD were positively related, whereas SLA and Nmass were uncorrelated). Overall, our model accounted for 47% of the spatial variability in Scots pine radial growth. Our study emphasizes the hierarchy of factors that determine intraspecific variations in functional traits in Scots pine and their strong association with spatial variability in radial growth. We claim that intraspecific trait variation is an important determinant of responses of plants to changes in climate and other environmental factors, and should be included in predictive models of vegetation dynamics.
Comparison of adaptive strategies to phosphorus-deficient soil between dominant species Artemisia frigida and Stipa krylovii in typical steppe of Nei Mongol
内蒙古典型草原优势种冷蒿和克氏针茅对土壤低磷环境适应策略的比较
DOI:10.3724/SP.J.1258.2014.00085
[本文引用: 1]
为探究典型草原植物长期共存的生理生态机制, 以典型草原的优势物种克氏针茅(Stipa krylovii)和冷蒿(Artemisia frigida)为材料, 采用基质培养方法, 通过比较不同供磷浓度对二者生物量、根系形态、质子分泌、酸性磷酸酶和有机酸分泌以及磷吸收利用效率的影响, 探讨克氏针茅和冷蒿对土壤磷缺乏的适应策略。研究结果表明: 冷蒿主要通过根系分泌酸性磷酸酶和酸化根际来适应低磷环境; 而克氏针茅主要是通过根系分泌有机酸(主要是苹果酸)来适应土壤磷缺乏。在低磷条件下, 克氏针茅和冷蒿的磷吸收效率没有显著差异, 但克氏针茅的磷利用效率显著高于冷蒿。随着供磷浓度增加, 二者的磷吸收速率增加, 磷利用效率降低。在生物量、地上部分性状以及根系生长方面, 克氏针茅和冷蒿对磷供给的响应都表现先增长后降低的趋势; 克氏针茅的生物量在外源供0.25 mmol·L<sup>-1</sup>磷时达到最大, 而冷蒿的生物量在外源供0.50 mmol·L<sup>-1</sup>磷时达到最大, 表明冷蒿对磷的生理需求高于克氏针茅。因此, 克氏针茅和冷蒿具备各自不同的适应土壤有效磷缺乏的生理策略, 这可能是它们在土壤贫瘠的温带典型草原长期共存的重要机制。
Plant functional traits—Concepts, applications and future directions
DOI:10.1360/N052014-00244 URL [本文引用: 1]
植物功能性状研究进展
Studies on biodiversity and ecosystem function via manipulation experiments
生物多样性与生态系统功能的实验研究
How do traits vary across ecological scales? A case for trait-based ecology
DOI:10.1111/j.1461-0248.2010.01476.x
PMID:20482582
[本文引用: 1]
Despite the increasing importance of functional traits for the study of plant ecology, we do not know how variation in a given trait changes across ecological scales, which prevents us from assessing potential scale-dependent aspects of trait variation. To address this deficiency, we partitioned the variance in two key functional traits (leaf mass area and leaf dry matter content) across six nested ecological scales (site, plot, species, tree, strata and leaf) in lowland tropical rainforests. In both traits, the plot level shows virtually no variance despite high species turnover among plots and the size of within-species variation (leaf + strata + tree) is comparable with that of species level variation. The lack of variance at the plot level brings substantial support to the idea that trait-based environmental filtering plays a central role in plant community assembly. These results and the finding that the amount of within-species variation is comparable with interspecific variation support a shift of focus from species-based to trait-based ecology.
A general and simple method for obtaining R2 from generalized linear mixed-effects models
DOI:10.1111/mee3.2013.4.issue-2 URL [本文引用: 2]
Fine root architecture of nine north American trees
DOI:10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2 URL [本文引用: 1]
The niche of higher plants: Evidence for phylogenetic conservatism
The world-wide ‘fast-slow' plant economics spectrum: A traits manifesto
DOI:10.1111/1365-2745.12211 URL [本文引用: 1]
Ectomycorrhizal fungi drive positive phylogenetic plant-soil feedbacks in a regionally dominant tropical plant family
Fine root morphology of twenty hardwood species in Maoershan natural secondary forest in northeastern China
帽儿山天然次生林20个阔叶树种细根形态
DOI:10.3773/j.issn.1005-264x.2008.06.002
[本文引用: 1]
细根在森林生态系统C分配和养分循环过程中发挥着重要作用。细根形态不但影响养分和水分的吸收, 而且与细根寿命和周转有密切关系。因此, 研究森林树种的细根形态对了解根系结构与功能、预测寿命与周转具有重要理论意义。该文根据细根分枝等级划分方法, 研究了东北帽儿山天然次生林20个阔叶树种1~5级根直径、根长和比根长等形态指标。结果表明, 20个树种中, 除5个树种1级根直径略大于2级和比根长略小于2级根外, 其余15个树种均表现为1级根直径和根长最小、比根长最高, 随着根序增加, 直径和根长增加, 而比根长降低。20个阔叶树种前3级根的累积根长均占前5级根总根长的80%以上。9个内生菌根侵染的树种的平均直径、根长和比根长均大于11个外生菌根侵染的树种。
Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: Relationship with daily irradiance
DOI:10.1046/j.1365-2435.2002.00672.x URL [本文引用: 1]
Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests
DOI:10.17520/biods.2015200 URL [本文引用: 1]
亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异
DOI:10.17520/biods.2015200
[本文引用: 1]
不同物种间的功能性状差异是自然生态系统中物种共存的基础, 而物种内个体间的性状变异对物种的共存和分布同样具有重要作用。本文以湖北星斗山自然保护区亚热带常绿落叶阔叶混交林内28种主要树种(通过物种多度排序获得, 其中常绿和落叶树种各14种)为研究对象, 探讨不同叶习性树种的4种功能性状(比叶面积、叶干物质含量、叶面积和比茎密度)在种间和种内的差异程度。结果表明: (1)常绿和落叶树种在4种功能性状上均存在显著差异, 常绿树种的比叶面积和叶面积显著低于落叶树种, 但叶干物质含量和比茎密度则显著高于落叶树种; (2)比叶面积的变化主要来源于叶习性(57.49%), 叶面积变化主要来源于种间(66.80%)和种内变异(27.52%), 叶干物质含量的变化主要来源于种间(38.12%)和种内(33.88%)变异, 但比茎密度的变化主要来源于种内变异(51.50%), 其次为种间变异(32.52%); (3)常绿和落叶树种种间水平的性状相关性可能掩盖各功能性状之间的相关性。种内变异能够显著影响群落间的植物功能性状差异, 但不同功能性状的种内变异程度存在差异。
The ectomycorrhizal symbiosis: Life in the real world
DOI:10.1017/S0269-915X(05)00303-4 URL [本文引用: 2]
Local diversity of native arbuscular mycorrhizal symbionts differentially affects growth and nutrition of three crop plant species
DOI:10.1007/s00374-017-1254-5 URL [本文引用: 1]
Ectomycorrhizal fungal succession in mixed temperate forests
DOI:10.1111/nph.2007.176.issue-2 URL [本文引用: 2]
Beyond the roots of young seedlings: The influence of age and order on fine root physiology
DOI:10.1007/s00344-003-0011-1 URL [本文引用: 1]
Plant ecological strategies: Some leading dimensions of variation between species
DOI:10.1146/annurev.ecolsys.33.010802.150452 URL [本文引用: 1]
Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests
DOI:10.1093/aob/mcl066 URL [本文引用: 1]
Assessing the generality of global leaf trait relationships
Global-scale quantification of relationships between plant traits gives insight into the evolution of the world's vegetation, and is crucial for parameterizing vegetation-climate models. A database was compiled, comprising data for hundreds to thousands of species for the core 'leaf economics' traits leaf lifespan, leaf mass per area, photosynthetic capacity, dark respiration, and leaf nitrogen and phosphorus concentrations, as well as leaf potassium, photosynthetic N-use efficiency (PNUE), and leaf N : P ratio. While mean trait values differed between plant functional types, the range found within groups was often larger than differences among them. Future vegetation-climate models could incorporate this knowledge. The core leaf traits were intercorrelated, both globally and within plant functional types, forming a 'leaf economics spectrum'. While these relationships are very general, they are not universal, as significant heterogeneity exists between relationships fitted to individual sites. Much, but not all, heterogeneity can be explained by variation in sample size alone. PNUE can also be considered as part of this trait spectrum, whereas leaf K and N : P ratios are only loosely related.
The worldwide leaf economics spectrum
Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
Effects of nitrogen addition on root dynamics in an alpine meadow, Northwestern Sichuan
DOI:10.17521/cjpe.2017.0255 URL [本文引用: 1]
氮肥添加对川西北高寒草甸植物群落根系动态的影响
DOI:10.17521/cjpe.2017.0255
[本文引用: 1]
该文以川西北高寒草甸为研究对象, 采用微根管法研究了不同施氮(N)水平下高寒草甸植物群落根系现存量、生产量、死亡量和周转率的变化及其与土壤理化性质的相互关系。结果表明: N添加显著增加了土壤速效氮(AN)含量, 降低了土壤pH值, 但是对土壤有机质(SOM)和全氮(TN)含量无显著影响。在0-10 cm土层, 平均根系现存量和累积根系生产量无显著变化, 累积根系死亡量在N10处理下显著降低了206.1 g·m <sup>-2</sup>, 根系周转率在N30处理下显著提高了17%; 在10-20 cm土层, N添加处理的平均根系现存量和累积根系生产量分别显著降低了195.3和142.3 g·m <sup>-2</sup> (N10)、235.8和212.1 g·m <sup>-2</sup> (N20)、198.0和204.4 g·m <sup>-2</sup> (N30), 累积根系死亡量和周转率无显著变化。此外, 累积根系生产量、死亡量和周转率与AN含量相关性较大, 而平均根系现存量与SOM、AN和TN含量相关性较大。综上所述, N添加对高寒草甸的影响主要通过改变土壤可利用N含量, 进而影响根系的动态特征、空间分布格局和周转以及碳分配特征。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}