


多物种共存机制是群落生态学的核心研究内容, 是生物多样性保护的基础, 其关键是生物间的相互作用。从19世纪的Logistic种群增长模型(种内直接相互作用, Verhuls, 1838), 到20世纪的Lotka-Volterra竞争模型(种内和种间的直接相互作用, Lotka, 1925; Volterra, 1926), 再到21世纪的当代物种共存理论(生态位差异与适合度差异, Chesson, 2000, 2018; 储诚进等, 2017)的近200年研究中, 生物间的直接相互作用一直是人们理解自然系统的关键机理和主体思想。除了成对物种之间的直接相互作用, 如竞争、互惠(如传粉、植物种子的动物传播等行为)、促进、(拟)寄生、捕食等, 人们很早就意识到多物种之间的间接相互作用(Darwin, 1859)。受还原论思维的影响, 人们总是将高层级的、复杂的对象分解为较低层级的、简单的对象来处理。应用于多物种共存的研究, 人们首先将复杂群落系统分解还原为简单的两两物种之间的相互作用(配对相互作用), 然后再将这些配对组装起来重新认识原先的复杂系统。在这个“一分一合”的过程中, 诸多与复杂系统相关的信息随之丢失, 其中最为重要的一个方面即为生物间的间接相互作用。这是因为直接相互作用无法考虑除成对物种以外的其他有机体的影响, 而在复杂系统里, 有机体常常存在于相互作用的网络并彼此交织在一起(图1)。另一方面, 人们因为理论模型在考虑间接相互作用之后变得过于复杂并认为间接相互作用的强度可能很弱, 而常常在研究中忽略间接相互作用。
图1
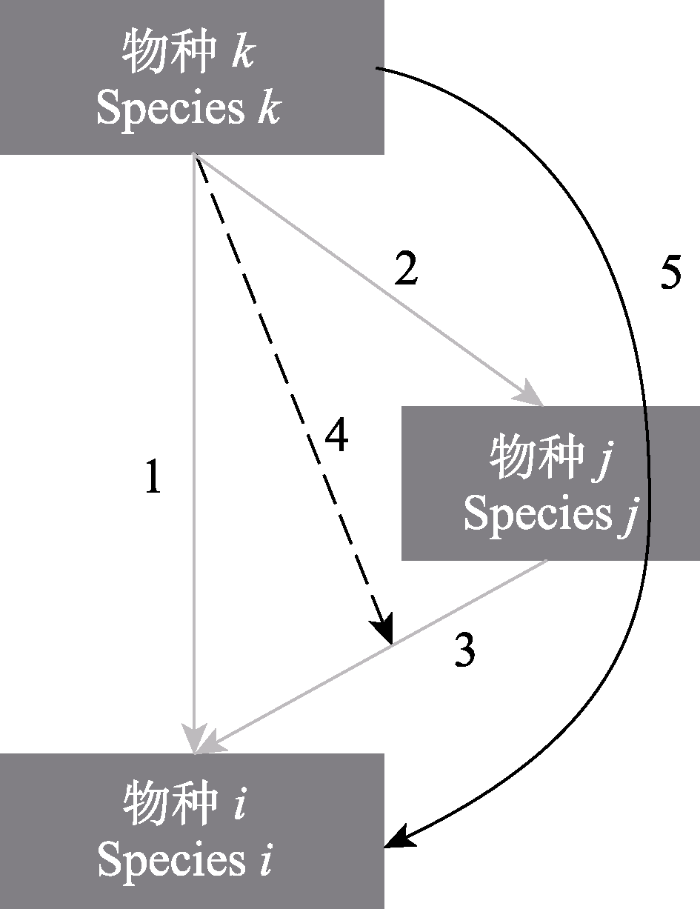
图1
包含直接和间接相互作用的生态网络。灰色箭头为直接配对相互作用(箭头1-3), 黑色箭头为间接相互作用(箭头4-5)。在间接相互作用中, 箭头4表示高阶相互作用, 即物种k影响的是物种j和i之间的相互作用, 箭头5表示链式相互作用, 即物种k先影响物种j的密度进而影响物种i。可见, 物种k对物种i既存在直接相互作用(箭头1), 也存在间接相互作用(箭头4和5)。箭头表示作用方向, 为简单起见, 只绘出了单向作用。
Fig. 1
The ecological network including direct (arrows 1-3) and indirect interactions (arrows 4-5) between species. Arrow 4 indicates that species k may indirectly affect species i by modifying the per capita effect of species j on species i (higher-order interactions, HOIs). Arrow 5 indicates that species k may indirectly affect species i by changing population density of species j (interaction chains). Therefore, species k may have both direct (arrow 1) and indirect (arrows 4-5) effects on species i. For simplicity, we only display direct and indirect effects of species j and k on species i.
复杂系统中忽略间接相互作用会导致至少两个方面的重要不足: (1)无法全面深入理解物种共存和生物多样性维持的机制, 配对相互作用的实验和理论工作很难外推至群落水平; (2)无法准确预测多样性如何响应生物/非生物环境变化, 其中间接相互作用是此类预测的关键不确定来源。理论工作和微宇宙实验证实了间接相互作用的普遍性和对群落及生态系统功能的重要性(Kerr et al, 2002; van Veen et al, 2005; Soliveres et al, 2015; Bairey et al, 2016; Gallien et al, 2017; Grilli et al, 2017; Letten & Stouffer, 2019), 然而鲜有来自复杂真实系统的研究。正如Levine等(2017)所言, 生态学极少有哪个方面能像间接相互作用这般将革新人们对生物多样性维持和分布机制的认识。明确考虑间接相互作用将有效完善和丰富群落生态学的理论框架, 夯实生态系统生态学的群落学基础。
基于Lotka-Volterra模型, 本文首先介绍了两类不同的间接相互作用: 链式相互作用和高阶相互作用。由于链式相互作用本质上还是配对相互作用, 因此我们重点介绍高阶相互作用, 包括跨营养级和同一营养级内高阶相互作用的研究历史和概况以及高阶相互作用定义的演变(种群水平), 进而介绍最新发展的基于个体的高阶相互作用(个体水平), 最后提出间接相互作用未来可能的研究重点和难点, 尤其是高阶相互作用的内在机理以及与功能性状之间的可能联系。
1 间接相互作用
相比两两物种的直接相互作用, 间接相互作用指一个物种通过中间物种对目标物种产生的间接影响, 因此不包括通过非生物因子对目标物种产生的间接影响(Wootton, 1994a; Abrams, 1995)。间接相互作用按照中间物种的介导方式可分为两类: (1)密度介导的间接相互作用(density-mediated indirect interactions, Abrams, 1995), 如物种k通过改变物种j的密度对物种i产生的间接相互作用(图1箭头5), 又称为链式相互作用(interaction chains); (2)性状介导的间接相互作用(trait-mediated indirect interactions, Abrams, 1995; Werner & Peacor, 2003), 如物种k通过改变物种j的性状而改变物种j对物种i的直接作用强度(图1箭头4), 也称高阶相互作用 (higher-order interactions, HOIs)。Lotka-Volterra模型 (简称L-V模型)作为生态学里最为重要的经典模型, 可以描述绝大多数物种间的相互作用, 间接相互作用也不例外。在经典的多物种L-V模型中, 物种i的单位种群增长率是所有物种(包括i本身)密度的线性函数(Chesson, 2012):
1.1 链式相互作用
在经典的多物种L-V模型中, 链式相互作用是普遍存在的, 因为链式相互作用本质上仍然是配对物种相互作用的迭代和延伸(Levine et al, 2017)。以图1的三物种群落为例, 在不考虑高阶相互作用时该群落的动态方程是:
在等式(2.1)中, 物种i在t时刻的单位种群增长速率受其自身及其竞争者(物种j和物种k)的影响, 其中物种j的密度Nj由t时间内的dNj累积而成, 而dNj又受物种k的密度制约(当然也受物种i和j的密度制约, 等式2.2)。因此在同时考虑多物种种群动态的L-V模型中隐含着这样一条相互作用链: 物种k通过改变物种j的密度间接影响物种i的种群增长(Nk→Nj→Ni, 图1箭头5)。同理可知L-V模型(等式2)中还包含有其他的相互作用链(如Nk→Ni→Nj, Nj→Nk→Ni等)。链式相互作用的形式十分多样, 由L-V模型中的相互作用矩阵A (种内和种间相互作用系数组成的矩阵, 等式3)可以体现, 不同形式的链式相互作用对群落动态的影响也不同(Stouffer & Bascompte, 2010; Soliveres et al, 2018; Losapio et al, 2019)。
相互作用矩阵A变化形式复杂多样, 我们这里仅以两种特殊情形(A1和A2)为例展示其对群落动态影响的差异。
在A1情况下, 物种的竞争优势为: i > j > k且i > k, 即物种i是最强的竞争者, j次之, k最弱, 最终物种i会竞争排除掉物种j和k, 这种链式相互作用被称为传递性竞争(transitive competition, Gallien et al, 2017)。在A2情况下, 物种的竞争优势为: i > j > k > i, 即三者中没有最强的竞争者, 而是呈现一种类似于剪刀-石头-布的相互制约(当i的种群密度增加时, 会抑制j的种群增长, 进而缓解了j对k的竞争, 最后k对i的抑制增强, 使i的变化趋向于稳定), 使得物种i、j和k虽不能两两共存, 但三者可以同时共存, 这种链式相互作用被称为非传递性竞争(intransitive competition, Gallien et al, 2017)。非传递性竞争所带来的这种制约回环, 已被证明可以促进多物种共存 (Kerr et al, 2002; Reichenbach et al, 2007; Allesina & Levine, 2011; Rojas-Echenique & Allesina, 2011), 而且群落的稳定性与非传递性环的出现频率、数量和长度呈正相关(Laird & Schamp, 2006, 2008; Gallien et al, 2017)。由于非传递性环广泛存在于多种生物类群中(Soliveres et al, 2018), 这种链式相互作用模式还可能显著影响物种多样性和生态系统功能 (Soliveres et al, 2015; Maynard et al, 2017)。除了在同一营养级的链式相互作用, 食物网中的链式相互作用也是长期被研究者关注的。例如食物网中特定的相互作用模式(motif), 包括食物链(food chain)、似然竞争(apparent competition)及杂食环(omnivory), 它们在自然界的出现频率和对食物网稳定性的影响都有显著区别(Bascompte & Melián, 2005; Stouffer & Bascompte, 2010; 徐光华等, 2019; 王少鹏, 2020)。除了特定的相互作用模式之外, 基于直接相互作用网络所构成的整体拓扑结构, 也可以揭示链式相互作用对群落的可能影响(方强和黄双全, 2012; 宋础良, 2020)。例如Bastolla等(2009)证明互惠网络的嵌套性(nestedness)可以使物种具有更多重合的互惠合作者, 进而最大化由合作者传递的间接促进作用, 从而可以减小物种之间的竞争强度并且维持物种多样性。
1.2 高阶相互作用
从而物种k通过调节物种j的性状对物种i的单位种群增长速率的改变为:
链式相互作用(物种k通过改变物种j的密度对物种i的间接作用)的发生具有时滞性(time lag), 而高阶相互作用(物种k通过改变物种j的性状对物种i的间接作用)的发生是即时的(immediateness)。在高阶相互作用存在的情况下, 群落的动态变得更加复杂和不可预测(Wootton, 1994a), 包含高阶相互作用模型的复杂度与物种数的幂指数(平方、立方等)成正比。事实上, 自然群落的动态复杂性确实超出直接相互作用与链式相互作用所预测的范围 (Mayfield & Stouffer, 2017)。由于高阶相互作用的现象在群落中十分普遍(Wootton, 1994a), 并且其相互作用强度已被证明并不小于直接相互作用(Werner & Peacor, 2003), 所以接下来着重介绍高阶相互作用这一非常重要却又长期被忽略的相互作用类型。
2 高阶相互作用研究的发展脉络
由于高阶相互作用的定义和相关术语的使用较为混淆, 这里我们简要整理了它们之间的关系 (图2)。传统意义上的高阶相互作用, 一般又称为相互作用的调节(interaction modifications, Case & Bender, 1981; Abrams, 1983; Adler & Morris, 1994; Levine et al, 2017), 是指一个物种对另一个物种的直接作用强度受到其他物种的影响。其发生机制在于物种j在物种k存在时有性状(形态、生理、行为等) 上的可塑性变化, 并且物种j的这种可塑性变化会改变其对物种i的直接作用强度(图1箭头4), 因而又称为性状介导的间接相互作用(Abrams, 1995; Werner & Peacor, 2003; Levine et al, 2017)。因此这类定义一般认为高阶相互作用只可能发生在由三个或三个以上物种所组成的系统中, 不过也有研究认为可发生在两物种的情况(物种j或者物种k可与目标物种i为同一物种) (Case & Bender, 1981; Kleinhesselink et al, 2019)。近年来相关研究将高阶相互作用定义为系统中所有物种(包括目标物种自身)对目标物种单位种群增长速率的非线性密度制约效应(nonlinear density dependence, Bairey et al, 2016; Kleinhesselink et al, 2019; Letten & Stouffer, 2019; Xiao et al, 2020)。Kleinhesselink等(2019)将这两类定义区分为狭义高阶相互作用(hard-HOIs, 前者)和广义高阶相互作用(soft-HOIs, 后者)。狭义的高阶相互作用因一个物种对另一个物种的直接作用强度依赖于其他物种, 一定会产生非线性密度制约效应, 因而属于广义高阶相互作用的范畴。而广义高阶相互作用不仅包含狭义相互作用, 还包含种内高阶相互作用 (intraspecific HOIs)或种内非线性(intraspecific nonlinearity), 即βij,jNj2项(等式5物种j和k为同一物种时)。传统的研究强调狭义与广义相互作用的区分, 并提出了一系列检验种群动态模型中是否包含狭义高阶相互作用的方法(表1)。我们则认为狭义与广义的高阶相互作用可统一于个体水平的高阶相互作用(individual-level HOIs), 详见下文第3节。
图2
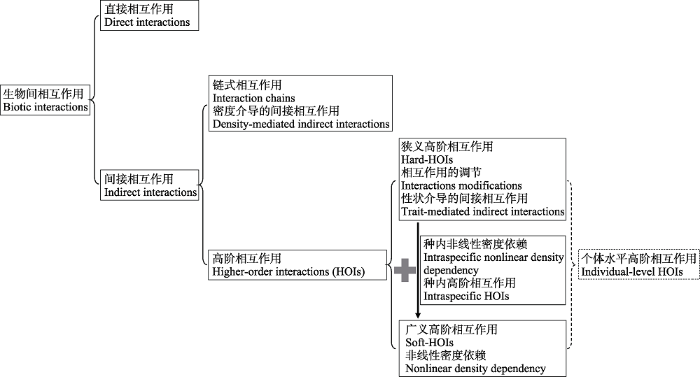
图2
生物间相互作用的类型和关系。同一方框内的不同术语为不同角度描述的同一类型的相互作用, 虚线部分为作者见解, 尚无相关文献明确说明。
Fig. 2
The types of biotic interactions. The different terms in the same box were used to describe the same type of interaction in different studies. The part in dashed line is our own opinion.
表1 检验模型是否包含狭义高阶相互作用的方法及部分常见模型检验结果。√表示模型中包含狭义高阶相互作用(模型不满足方法中等式), ×表示模型不含狭义高阶相互作用(模型满足方法中等式)。这些方法旨在将狭义高阶相互作用从广义高阶相互作用中区分出来, 其中(1)、(3)和(5)用于检验两物种或两物种以上系统是否包含狭义高阶相互作用, (2)和(4)分别是(1)和(3)用于将狭义高阶相互作用严格定义在三物种或三物种以上系统中时的情况, 因而方法(1)和(2), (3)和(4)在三个以上物种组成的系统中等效。Fi表示物种i的单位种群增长率是其自身及竞争者密度的函数, 这里给出几个常见模型的Fi函数表达式。如果函数Fi对Nj的偏导数¶Fi/¶Nj能表达成Nj的函数Gij(Nj) (方法1), 或是Ni和Nj的函数Gij(Ni, Nj) (方法2), 或是Nj和Fi自身的函数Gij(Fi, Nj) (方法3), 或是Ni和Nj以及Fi的函数Gij(Fi, Ni, Nj) (方法4), 则模型没有狭义高阶相互作用。方法(5)中, Qi表示函数Fi中所有参数的集合, jij表示除物种j外所有物种的密度均为0时函数Fi(0, …, Nj, …, 0)中的参数, Fi则是jij (j = 1, …, S)的并集。若Qi = Fi, 则模型没有狭义高阶相互作用。
Table 1
 |
2.1 跨营养级的高阶相互作用
在食物链/网研究中, 高阶相互作用通常被称为性状介导的间接相互作用。Werner和Peacor (2003)以及朱玉等(2017)详细回顾了性状介导的间接相互作用的特征、发生机制、作用途径及实验证据, 并根据性状将其分为通过行为、生理、形态、发育以及生活史等介导的间接相互作用。现有研究主要关注的是行为和生理性状, 而关于形态、发育以及生活史等方面的性状的研究实例并不多(e.g. Mopper et al, 1991; Tscharntke, 1999; van Veen & Godfray, 2013; Xi et al, 2016)。在行为性状方面, 处于食物链中间的物种在捕食风险和能量摄入两方面存在权衡, 因此其采食行为同时受到上、下营养级物种关系的调控。面对捕食者时要增加躲避时间进而能量摄入减少, 而当处于较低的资源水平时采食活动更加频繁, 被捕食风险增大(Huang & Sih, 1990; Johansson, 1995; Beckerman et al, 1997; Lima, 1998)。另一类基于行为介导的间接相互作用是通过生境利用的转变 (Messina, 1981; Werner et al, 1983; Werner & Gilliam, 1984; Wootton, 1993; Turner, 1996, 1997)。如当捕食者大口黑鲈(Micropterus salmoides)存在时, 蓝鳃太阳鱼(Lepomis macrochirus)对生境的利用从池塘中心逐渐转移至池塘沿岸植被区域, 导致池塘中心浮游动物丰度显著增加(Turner & Mittelbach, 1990)。关于生理性状的研究主要集中在植物的诱导性防御反应。例如, 美国赤杨(Alnus rubra)在被采食后产生防御反应, 通过降低叶片氮含量进而提高碳氮比, 减少了陆生植食者和水生分解者对其叶片的消耗(Jackrel & Wootton, 2015)。近年在四川红原高寒草甸的野外观测发现, 菊科植物的花序对拟寄生蜂寄生实蝇有明显的限制和选择作用, 即高阶专化作用(higher-order specialization, Xi et al, 2017)。Liao等(2020)通过对菊科植物-石蝇-拟寄生蜂三分网络的模拟分析发现, 高阶专化作用提升了石蝇物种多样性, 但降低了菊科植物和寄生蜂的物种丰富度, 强调了来自多分网络的不同物种之间的高阶相互作用对物种多样性维持的重要性。此外, 一些依赖动物扩散种子的植物会通过大年结实调控动物贮食行为而提高种子扩散效率, 因此动物行为的变化可以介导植物间的相互作用关系, 进而影响植物共存(Yang et al, 2020; 杨锡福等, 2020)。众多的实验证据表明, 性状介导的间接相互作用在水生和陆生系统中均普遍存在, 且对群落动态有显著的影响。
2.2 同一营养级的高阶相互作用
与跨营养级研究类似, 性状介导的间接相互作用也会导致同一营养级物种之间产生高阶相互作用。例如, 长叶车前(Plantago lanceolate)会抑制紫羊茅(Festuca rubra)根系的生长, 进而减弱紫羊茅对群落内其他物种的竞争强度(Padilla et al, 2013)。需要特别注意的是, 表象竞争模型 (phenomenological model of competition), 如L-V模型, 通常用竞争系数来描述一个物种对另一个物种的竞争强度, 而不考虑具体的生态学过程或潜在机制。基于消费者-资源竞争(consumer-resource competition)的机理模型(mechanistic model of competition)表明, 当资源非logistic增长或消费者对资源密度呈现非线性功能响应时, 就会产生高阶相互作用(Abrams, 1983; Kleinhesselink et al, 2019; Letten & Stouffer, 2019)。这表明, 高阶相互作用事实上是表象模型的涌现特征(emergent properties)。然而, 无论是考虑表象还是机理竞争模型, 同一营养级两个物种之间的相互作用在多大程度上受群落内其他物种的影响仍是一个至关重要的问题。
虽然跨营养级物种之间的高阶相互作用已有广泛研究, 却鲜有研究在自然群落和实验系统中去验证同一营养级高阶相互作用的普遍性和相对重要性。同一营养级高阶相互作用的实验研究可以追溯到20世纪60年代(Hairston et al, 1968; Vandermeer, 1969), 这一期间的研究主要关注的是高阶相互作用的检验方法。最初, 人们意识到简单线性的L-V竞争模型常常无法准确描述和预测物种相互作用对群落动态的影响, 因而通过在模型中加入高阶交互项对L-V模型进行扩展(Vandermeer, 1969; Wilbur, 1972; Neill, 1974)。这也是为什么关于同一营养级高阶相互作用的研究主要集中在种群水平。经典的实验设计是分别在两两配对以及多物种组合下评估物种的表现, 检验目标物种对其他物种单独的响应(配对相互作用)能否预测出目标物种对多物种组合的响应(Vandermeer, 1969; Morin et al, 1988; Worthen & Moore, 1991)。常用的统计检验方法是方差分析(ANOVA)。但需要注意的是, 对于不同的竞争模型(包括不同响应变量、数据转换、模型函数形式等, 描述种群动态和竞争的模型有很多, L-V模型只是其中一类), 对应的统计检验方法也可能不同(Case & Bender, 1981; Billick & Case, 1994; Wootton, 1994b)。检验高阶相互作用在自然系统中重要性的另外一种方法是在配对竞争模型的基础上纳入高阶相互作用的影响, 对实验或自然群落观测数据进行统计拟合(Weigelt et al, 2007; Mayfield & Stouffer, 2017)。量化高阶相互作用是理解高阶相互作用重要性的第一步, 目前依然是一个巨大的挑战。
3 基于个体的高阶相互作用
近年来高阶相互作用的研究主要是在经典的L-V模型中(仅考虑直接相互作用)引入密度制约的高阶项, 然后通过模拟比较种群动态在考虑高阶相互作用后与经典模型的差异, 进而探讨其对物种共存和生物多样性的影响(Bairey et al, 2016; Letten & Stouffer, 2019; Singh & Baruah, 2020)。一方面,种群的动态变化归根结底是个体的存活、生长和繁殖的过程 (不考虑迁入、迁出), 因而未来高阶相互作用的研究需要更加关注其对个体适合度(存活、生长和繁殖) 的影响。Mayfield和Stouffer (2017)首次在一年生草本植物群落中探索了高阶相互作用对个体种子数量的影响。另一方面, 广义高阶相互作用中的种内非线性虽不属于传统的狭义高阶相互作用的范畴, 但其本质上可以解释为个体水平的相互作用的调节: 物种i的一个个体对另一个个体的直接作用强度是依赖于该物种其他个体的。因此, 广义高阶相互作用和狭义高阶相互作用实际上可统一于个体水平的高阶相互作用(图2虚线部分)。尤为重要的是, 个体水平的高阶相互作用可以明确将个体间的差异(如个体大小、个体的空间分布)考虑进来, 对研究诸如森林群落中的直接与高阶相互作用具有非常重要的意义(Hegyi, 1974; Canham et al, 2004; Uriarte et al, 2004; Hasenauer, 2006)。以森林群落为例, Li等(2020)最近提出了在量化邻体(N个邻体分属S个物种, $N\text{=}\underset{j\text{=1}}{\overset{s}{\mathop \sum }}\,{{N}_{j}}$, Nj是物种j的多度)对目标个体im的直接与高阶相互作用中考虑个体大小与空间分布的一种新方法(图3)。N个邻体对目标个体im的直接相互作用($DI_{i_{m}}|[N]$)是每个邻体对目标个体直接相互作用之和(图3直线箭头), 并假定邻体jp对目标个体im的直接相互作用强度($\alpha_{i_{m}j_{p}}$)与邻体大小(用邻体胸径DBH度量)成正比而与邻体到目标个体的距离(d[im, jp])成反比, 且这种直接相互作用仅发生于邻体在目标个体给定半径为R的邻域内:
图3
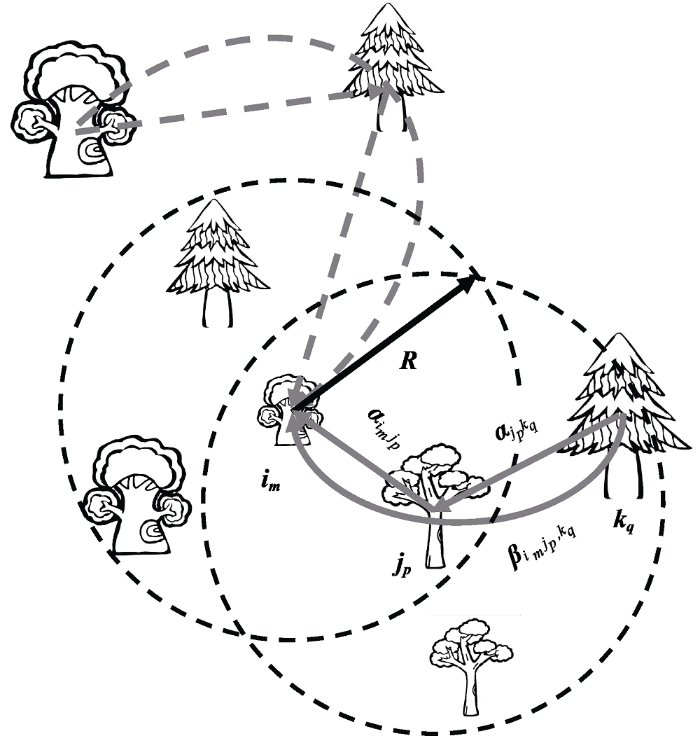
图3
邻体对目标个体的直接(直线箭头)与高阶相互作用(曲线箭头)。参数$\alpha_{i_{m}j_{p}}$表示的是邻体jp对目标个体im的直接相互作用, 参数$\beta_{i_{m}j_{p},k_{q}}$表示的是邻体kq通过个体jp对目标个体im的高阶相互作用。森林群落研究中一般假定直接相互作用发生于邻体jp在目标个体im半径为R的邻域内(实直线箭头), 因而高阶相互作用发生于当邻体jp在目标个体im半径为R的邻域内且邻体的邻体kq在邻体jp的邻域内(实曲线箭头)。虚线箭头表示邻域外不需要考虑的直接与高阶相互作用。
Fig. 3
Direct (straight arrows) and higher-order interactions (curve arrows) of neighbouring trees on a focal tree. The parameter $\alpha_{i_{m}j_{p}}$ quantifies the direct effect of a neighbour (individual p of species j) on the focal tree (individual m of species i). The direct interaction occurs only when a neighbour (jp) is located within a maximum radius (R) of im (solid straight arrows). The parameter $\beta_{i_{m}j_{p},k_{q}}$ quantifies the higher-order effect of a neighbour (individual q of species k) on the focal tree through another neighbour (individual p of species j). Higher-order interaction occurs only jp is located within the maximum radius (R) of im and kq is located within the maximum radius (R) of jp (solid curve arrows). Dashed arrows indicate direct interactions and higher-order interactions that are not considered when a neighbour is located outside the maximum radius (R) of the focal tree or its neighbour(s).
为计算方便, 将d[im, jp]大于邻域半径R的这些距离设置成无穷大以去掉邻域外个体对目标个体的影响。N邻体对目标个体的高阶相互作用是每个邻体通过其他邻体对目标个体高阶相互作用之和(图3曲线箭头), 并且邻体kq通过邻体jp对目标个体im的高阶相互作用($\beta_{i_{m}j_{p},k_{q}}$)取决于邻体kq对邻体jp的直接相互作用强度($\alpha_{j_{p}k_{q}}$)以及邻体jp对目标个体im的直接相互作用强度($\alpha_{i_{m}j_{p}}$):
同样地,如果d[im, jp]或d[jp, kq]大于邻域半径R, 就将其设置成无穷大。在不考虑个体大小与空间距离时(u = 0且v = 0), 邻体对目标个体的直接与高阶相互作用的量化可简化成仅与邻体密度相关的形式(Mayfield & Stouffer, 2017)。考虑个体大小与空间分布时(u ≠ 0且v ≠0)的量化方法可区分邻体kq通过邻体jp对目标个体im的高阶相互作用($\beta_{i_{m}j_{p},k_{q}}$)和邻体jp通过邻体kq对目标个体im的高阶相互作进而区分物种k通过物种j对物种i的高阶相互作用(βij,k)和物种j通过物种k对物种i的高阶相互作用(βik,j), 这是仅在种群水平上量化高阶相互作用所无法实现的。
4 研究展望
4.1 检验自然群落中高阶相互作用的普遍性与相对重要性
尽管最近的研究从理论层面上证明了高阶相互作用对物种共存与物种多样性维持的重要意义(Bairey et al, 2016; Grilli et al, 2017; Letten & Stouffer, 2019; Singh & Baruah, 2020), 然而鲜有研究在自然植物群落中去验证同一营养级内高阶相互作用的普遍性和相对重要性。Mayfield和Stouffer (2017)研究发现在一年生草本植物群落中, 包含高阶相互作用的模型显著提高了对个体产生种子数量的解释度, 从而首次证实高阶相互作用在自然群落中的重要性。今后的研究需要在更多不同的自然群落中采用类似于第3节中介绍的量化个体水平高阶相互作用的方法, 去检验高阶相互作用对个体适合度(存活、生长和繁殖)的影响。同时, 由于这种量化高阶相互作用的方法中的参数数量与物种数的平方成正比, 因此很难直接适用于物种丰富度较高的群落。最近已有研究表明将邻体按生活型或个体大小等分成数量较少的类群后(而非按物种分类), 再采用这种方法量化高阶相互作用, 既可简化模型的复杂度又能提高模型解释度和预测能力(Li et al, 2020; Martyn et al, 2020)。在森林群落中, 已有许多工作研究邻体直接相互作用对目标个体的存活和生长的影响(Hegyi, 1974; Lorimer, 1983; Wykoff, 1990; Monserud & Sterba, 1999; Canham et al, 2004; Uriarte et al, 2004; Hasenauer, 2006), 然而到目前为止却未见研究邻体高阶相互作用在其中所起作用的文献。基于固定监测样地的多次普查数据使得检验不同森林群落中邻体高阶相互作用对目标个体存活和生长的相对重要性成为可能。在此基础上, 可进一步在全球尺度上研究森林群落中高阶相互作用的纬度梯度格局, 并推断潜在的生物(多样性等)和非生物影响因素(气候、地形和土壤等)。
4.2 揭示高阶相互作用的内在机制
通过上述自然群落的观察研究(用包含高阶相互作用的模型拟合自然群落的观测数据)可探索高阶相互作用的普遍性和相对重要性(Where, When and What), 但难以揭示高阶相互作用发生的内在机制(How and Why) (Letten & Stouffer, 2019)。在植物群落中鲜有单种(无邻体, 无相互作用)与双种(一个邻体, 仅存在直接相互作用)的情况, 故而需要通过控制实验的手段, 比较它们和多种(多个邻体, 直接和高阶相互作用)情况下个体存活、生长和繁殖的差异, 为个体水平高阶相互作用提供更直接有力的证据。并在此基础上建立高阶相互作用强度与目标个体在有无邻体情况下性状的可塑性变化强度的联系, 从而揭示高阶相互作用性状介导的发生机制(高阶相互作用主要是通过哪种性状的可塑性变化所介导的, 这种可塑性变化的强度和方向如何, 高阶相互作用是放大还是抑制直接相互作用)。如邻体kq通过邻体jp对目标个体im的高阶相互作用, 可能是由于邻体kq通过抑制邻体jp的根系生长, 从而抑制邻体jp对目标个体im的直接作用强度(抑制jp对目标个体im根系生长的抑制)。在上述实验的基础上, 进行杀菌和去除植食性昆虫等处理, 可进一步研究多营养级之间高阶相互作用发生的内在机制。此外, 探索竞争相互作用的机理模型, 通过明确包含资源或者捕食者的动态, 有助于预测什么情况下高阶相互作用可能出现及对种群动态、群落结构和生态系统功能的影响(Abrams, 1983; Letten & Stouffer, 2019)。
4.3 高阶相互作用对多物种共存和生态系统功能的影响
揭示高阶相互作用对物种共存的影响, 需要进一步与当代物种共存理论相结合, 探索什么情况下高阶相互作用会有利于或不利于物种共存, 以及所带来的生态系统水平上的影响。例如, 不同方向和强度的高阶相互作用如何通过调节种内和种间相互作用影响物种共存以及竞争网络对物种丧失的稳健性(Singh & Baruah, 2020)。此外, 已有的关于生物多样性和生态系统功能的研究主要集中在同一个营养级内部, 如植物多样性与生态系统功能关系的研究 (Tilman et al, 1997; Loreau, 1998, 2000; Hector et al, 1999; Spehn et al, 2005)。近年来, 基于食物网的相关研究发现, 不同营养级的生物多样性也会对生态系统功能产生复杂的影响(Thébault & Loreau, 2003; Ives et al, 2005)。在此基础上, 一些研究通过在食物网中引入高阶相互作用, 发现其能影响生态系统功能以及生物多样性与生态系统功能之间的关系(Arditi et al, 2005; Goudard & Loreau, 2008; Lin & Sutherland, 2014)。然而, 目前还没有研究关注同一营养级内部高阶相互作用对生态系统功能的影响。简言之, 将间接相互作用尤其是高阶相互作用嵌入现有的各类生态学理论框架中, 或许可以更好地描述和预测人类世背景下的种群动态、群落结构和生态系统功能。
参考文献
Arguments in favor of higher order interactions
Implications of dynamically variable traits for identifying, classifying, and measuring direct and indirect effects in ecological communities
A general test for interaction modification
A competitive network theory of species diversity
Rheagogies: Modelling non-trophic effects in food webs
High-order species interactions shape ecosystem diversity
DOI:10.1038/ncomms12285
URL
PMID:27481625
[本文引用: 4]

Classical theory shows that large communities are destabilized by random interactions among species pairs, creating an upper bound on ecosystem diversity. However, species interactions often occur in high-order combinations, whereby the interaction between two species is modulated by one or more other species. Here, by simulating the dynamics of communities with random interactions, we find that the classical relationship between diversity and stability is inverted for high-order interactions. More specifically, while a community becomes more sensitive to pairwise interactions as its number of species increases, its sensitivity to three-way interactions remains unchanged, and its sensitivity to four-way interactions actually decreases. Therefore, while pairwise interactions lead to sensitivity to the addition of species, four-way interactions lead to sensitivity to species removal, and their combination creates both a lower and an upper bound on the number of species. These findings highlight the importance of high-order species interactions in determining the diversity of natural ecosystems.
Simple trophic modules for complex food webs
The architecture of mutualistic networks minimizes competition and increases biodiversity
DOI:10.1038/nature07950
URL
PMID:19396144
[本文引用: 1]
The main theories of biodiversity either neglect species interactions or assume that species interact randomly with each other. However, recent empirical work has revealed that ecological networks are highly structured, and the lack of a theory that takes into account the structure of interactions precludes further assessment of the implications of such network patterns for biodiversity. Here we use a combination of analytical and empirical approaches to quantify the influence of network architecture on the number of coexisting species. As a case study we consider mutualistic networks between plants and their animal pollinators or seed dispersers. These networks have been found to be highly nested, with the more specialist species interacting only with proper subsets of the species that interact with the more generalist. We show that nestedness reduces effective interspecific competition and enhances the number of coexisting species. Furthermore, we show that a nested network will naturally emerge if new species are more likely to enter the community where they have minimal competitive load. Nested networks seem to occur in many biological and social contexts, suggesting that our results are relevant in a wide range of fields.
Experimental evidence for a behavior-mediated trophic cascade in a terrestrial food chain
Higher order interactions in ecological communities: What are they and how can they be detected?
A neighborhood analysis of canopy tree competition: Effects of shading versus crowding
Testing for higher order interactions
Mechanisms of maintenance of species diversity
Species competition and predation
Updates on mechanisms of maintenance of species diversity
Advances in species coexistence theory
物种共存理论研究进展
On the Origin of Species by Means of Natural Selection
Progress in pollination networks: Network structure and dynamics
传粉网络的研究进展: 网络的结构和动态
The effects of intransitive competition on coexistence
DOI:10.1111/ele.12775
URL
PMID:28547799
[本文引用: 4]
Coexistence theory has been developed with an almost exclusive focus on interactions between two species, often ignoring more complex and indirect interactions, such as intransitive loops, that can emerge in competition networks. In fact, intransitive competition has typically been studied in isolation from other pairwise stabilising processes, and thus little is known about how intransitivity interacts with more traditional drivers of species coexistence such as niche partitioning. To integrate intransitivity into traditional coexistence theory, we developed a metric of growth rate when rare, Deltari , to identify and quantify the impact of intransitive competition against a backdrop of pairwise stabilising niche differences. Using this index with simulations of community dynamics, we demonstrate that intransitive loops can both stabilise or destabilise species coexistence, but the strength and importance of intransitive interactions are significantly affected by the length and the topology of these loops. We conclude by showing how Deltari can be used to evaluate effects of intransitivity in empirical studies. Our results emphasise the need to integrate complex mechanisms emerging from diverse interactions into our understanding of species coexistence.
Nontrophic interactions, biodiversity, and ecosystem functioning: An interaction web model
DOI:10.1086/523945
URL
PMID:18171154
[本文引用: 1]
Research into the relationship between biodiversity and ecosystem functioning has mainly focused on the effects of species diversity on ecosystem properties in plant communities and, more recently, in food webs. Although there is growing recognition of the significance of nontrophic interactions in ecology, these interactions are still poorly studied theoretically, and their impact on biodiversity and ecosystem functioning is largely unknown. Existing models of mutualism usually consider only one type of species interaction and do not satisfy mass balance constraints. Here, we present a model of an interaction web that includes both trophic and nontrophic interactions and that respects the principle of mass conservation. Nontrophic interactions are represented in the form of interaction modifications. We use this model to study the relationship between biodiversity and ecosystem properties that emerges from the assembly of entire interaction webs. We show that ecosystem properties such as biomass and production depend not only on species diversity but also on species interactions, in particular on the connectance and magnitude of nontrophic interactions, and that the nature, prevalence, and strength of species interactions in turn depend on species diversity. Nontrophic interactions alter the shape of the relationship between biodiversity and biomass and can profoundly influence ecosystem processes.
Higher-order interactions stabilize dynamics in competitive network models
DOI:10.1038/nature23273
URL
PMID:28746307
[本文引用: 2]
Ecologists have long sought a way to explain how the remarkable biodiversity observed in nature is maintained. On the one hand, simple models of interacting competitors cannot produce the stable persistence of very large ecological communities. On the other hand, neutral models, in which species do not interact and diversity is maintained by immigration and speciation, yield unrealistically small fluctuations in population abundance, and a strong positive correlation between a species' abundance and its age, contrary to empirical evidence. Models allowing for the robust persistence of large communities of interacting competitors are lacking. Here we show that very diverse communities could persist thanks to the stabilizing role of higher-order interactions, in which the presence of a species influences the interaction between other species. Although higher-order interactions have been studied for decades, their role in shaping ecological communities is still unclear. The inclusion of higher-order interactions in competitive network models stabilizes dynamics, making species coexistence robust to the perturbation of both population abundance and parameter values. We show that higher-order interactions have strong effects in models of closed ecological communities, as well as of open communities in which new species are constantly introduced. In our framework, higher-order interactions are completely defined by pairwise interactions, facilitating empirical parameterization and validation of our models.
The relationship between species diversity and stability: An experimental approach with protozoa and bacteria
Sustainable Forest Management: Growth Models for Europe
Discrete time models for two-species competition
DOI:10.1016/0040-5809(76)90045-9 URL PMID:1273800
Plant diversity and productivity experiments in European grasslands
DOI:10.1126/science.286.5442.1123
URL
PMID:10550043
[本文引用: 1]
At eight European field sites, the impact of loss of plant diversity on primary productivity was simulated by synthesizing grassland communities with different numbers of plant species. Results differed in detail at each location, but there was an overall log-linear reduction of average aboveground biomass with loss of species. For a given number of species, communities with fewer functional groups were less productive. These diversity effects occurred along with differences associated with species composition and geographic location. Niche complementarity and positive species interactions appear to play a role in generating diversity-productivity relationships within sites in addition to sampling from the species pool.
A simulation model for managing jack-pine stands. In: Growth Models for Tree and Stand Simulation (ed. Fries J)
pp.
Experimental studies on behaviorally mediated, indirect interactions through a shared predator
A synthesis of subdisciplines: Predator-prey interactions, and biodiversity and ecosystem functioning
Cascading effects of induced terrestrial plant defences on aquatic and terrestrial ecosystem function
Increased prey vulnerability as a result of prey-prey interactions
DOI:10.1007/BF00007398 URL [本文引用: 1]
Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors
DOI:10.1038/nature00823
URL
PMID:12110887
[本文引用: 2]
One of the central aims of ecology is to identify mechanisms that maintain biodiversity. Numerous theoretical models have shown that competing species can coexist if ecological processes such as dispersal, movement, and interaction occur over small spatial scales. In particular, this may be the case for non-transitive communities, that is, those without strict competitive hierarchies. The classic non-transitive system involves a community of three competing species satisfying a relationship similar to the children's game rock-paper-scissors, where rock crushes scissors, scissors cuts paper, and paper covers rock. Such relationships have been demonstrated in several natural systems. Some models predict that local interaction and dispersal are sufficient to ensure coexistence of all three species in such a community, whereas diversity is lost when ecological processes occur over larger scales. Here, we test these predictions empirically using a non-transitive model community containing three populations of Escherichia coli. We find that diversity is rapidly lost in our experimental community when dispersal and interaction occur over relatively large spatial scales, whereas all populations coexist when ecological processes are localized.
Mechanisms underlying higher order interactions: From quantitative definitions to ecological processes
DOI:10.1101/2020.10.31.362848
URL
PMID:33398270
[本文引用: 4]
Neutralizing antibodies (nAbs) hold promise as effective therapeutics against COVID-19. Here, we describe protein engineering and modular design principles that have led to the development of synthetic bivalent and tetravalent nAbs against SARS-CoV-2. The best nAb targets the host receptor binding site of the viral S-protein and its tetravalent versions can block entry with a potency that exceeds the bivalent nAbs by an order of magnitude. Structural studies show that both the bivalent and tetravalent nAbs can make multivalent interactions with a single S-protein trimer, observations consistent with the avidity and potency of these molecules. Significantly, we show that the tetravalent nAbs show much increased tolerance to potential virus escape mutants. Bivalent and tetravalent nAbs can be produced at large-scale and are as stable and specific as approved antibody drugs. Our results provide a general framework for developing potent antiviral therapies against COVID-19 and related viral threats, and our strategy can be readily applied to any antibody drug currently in development.
Competitive intransitivity promotes species coexistence
DOI:10.1086/506259
URL
PMID:16874628
[本文引用: 1]
Using a spatially explicit cellular automaton model with local competition, we investigate the potential for varied levels of competitive intransitivity (i.e., nonhierarchical competition) to promote species coexistence. As predicted, on average, increased levels of intransitivity result in more sustained coexistence within simulated communities, although the outcome of competition also becomes increasingly unpredictable. Interestingly, even a moderate degree of intransitivity within a community can promote coexistence, in terms of both the length of time until the first competitive exclusion and the number of species remaining in the community after 500 simulated generations. These results suggest that modest levels of intransitivity in nature, such as those that are thought to be characteristic of plant communities, can contribute to coexistence and, therefore, community-scale biodiversity. We explore a potential connection between competitive intransitivity and neutral theory, whereby competitive intransitivity may represent an important mechanism for
Does local competition increase the coexistence of species in intransitive networks
DOI:10.1890/07-0117.1
URL
PMID:18376565
[本文引用: 1]
Competitive intransitivity, a situation in which species' competitive ranks cannot be listed in a strict hierarchy, promotes species coexistence through
The mechanistic basis for higher-order interactions and non-additivity in competitive communities
DOI:10.1111/ele.13211
URL
PMID:30675983
[本文引用: 7]
Motivated by both analytical tractability and empirical practicality, community ecologists have long treated the species pair as the fundamental unit of study. This notwithstanding, the challenge of understanding more complex systems has repeatedly generated interest in the role of so-called higher-order interactions (HOIs) imposed by species beyond the focal pair. Here we argue that HOIs - defined as non-additive effects of density on per capita growth - are best interpreted as emergent properties of phenomenological models (e.g. Lotka-Volterra competition) rather than as distinct 'ecological processes' in their own right. Using simulations of consumer-resource models, we explore the mechanisms and system properties that give rise to HOIs in observational data. We demonstrate that HOIs emerge under all but the most restrictive of assumptions, and that incorporating non-additivity into phenomenological models improves the quantitative and qualitative accuracy of model predictions. Notably, we also observe that HOIs derive primarily from mechanisms and system properties that apply equally to single-species or pairwise systems as they do to more diverse communities. Consequently, there exists a strong mandate for further recognition of non-additive effects in both theoretical and empirical research.
Beyond pairwise mechanisms of species coexistence in complex communities
Beyond direct neighbourhood effects: Higher-order interactions improve modelling and predicting tree survival and growth
DOI:10.1093/nsr/nwaa009 URL PMID:32288962 [本文引用: 2]
Metacommunity robustness of plant-fly-wasp tripartite networks with specialization to habitat loss
Stress and decision making under the risk of predation: Recent developments from behavioral, reproductive, and ecological perspectives. In: Advances in the Study of Behavior: Stress and Behavior (eds Moller AP, Milinski M, Slater PJB)
pp.
Interaction modification effects on ecological networks are affected by ratio dependence and network topology
DOI:10.1016/j.jtbi.2014.08.025
URL
PMID:25167789
[本文引用: 1]
Interaction modification (IM), where one species modifies the strength of the density-mediated direct interaction between two other species, is an important ecological process, but little is known about the collective effect of multiple IM on overall community dynamics. We use stochastic bioenergetic modelling of ecological networks with different network topologies, functional responses and parameter values, to investigate the effects of IM connectance and IM strength on ecosystem properties including the evenness of species abundances and variability of system biomass. We found that the maximum system biomass that could potentially be attained by the model systems increased with IM connectance and strength when the models had nonrandom topology and prey-dependent functional responses as opposed to random topology and ratio-dependent responses. The maximum potential species evenness increased with IM strength but decreased with increasing IM connectance, when all modifications were negative. These findings underscore the importance of accounting for multiple IM across the community for understanding complex community dynamics.
Biodiversity and ecosystem functioning: A mechanistic model
Biodiversity and ecosystem functioning: Recent theoretical advances
Tests of age-independent competition indices for individual trees in natural hardwood stands
Perspectives for ecological networks in plant ecology
Identifying ‘useful’ fitness models: Balancing the benefits of added complexity with realistic data requirements in models of individual plant fitness
DOI:10.1086/711418
URL
PMID:33211565
[本文引用: 1]
The American Naturalist recently passed its sesquicentennial. Throughout this long history, it regularly encountered moments of introspection and debate over its goals, mission, identity, and audience. Here, we chronicle the history of those debates and transitions at critical moments. The Naturalist began as a popular magazine for amateur naturalists in the late 1860s. In the late 1870s, it transitioned to an increasingly academic journal for professional scientists from all branches of the natural sciences. By the turn of the century, academic specialization led to increasing fragmentation of the sciences into a multitude of societies and journals, creating an identity crisis for the once-broad-reaching American Naturalist. This identity crisis was resolved when the journal pivoted around 1910 to focus on fundamental advances in the newly emerging field of genetics. In the 1960s, the journal underwent a remarkably rapid transition to its present focus on evolution and ecology. The profound shifts in the journal's contents over this time are a reflection of the historical changes in science as a whole: from amateur naturalists, to polymath professionals, to increasingly specialized academics. This chronicle reveals the ways in which The American Naturalist has left its mark on many disciplines, many of which are today only loosely affiliated with the journal, if at all.
Higher-order interactions capture unexplained complexity in diverse communities
Competitive network determines the direction of the diversity-function relationship
Plant protection as a consequence of an ant-membracid mutualism: Interactions on goldenrod (Solidago sp.)
Modeling individual tree mortality for Austrian forest species
A new look at habitat structure: Consequences of herbivore-modified plant architecture. In: Habitat Structure: The Physical Arrangement of Objects in Space (eds Bell SS, McCoy ED, Mushinsky HR)
pp.
Competition between aquatic insects and vertebrates: Interaction strength and higher order interactions
The community matrix and interdependence of the competition coefficients
Early root overproduction not triggered by nutrients decisive for competitive success belowground
Mobility promotes and jeopardizes biodiversity in rock-paper-scissors games
DOI:10.1038/nature06095
URL
PMID:17728757
[本文引用: 1]
Biodiversity is essential to the viability of ecological systems. Species diversity in ecosystems is promoted by cyclic, non-hierarchical interactions among competing populations. Central features of such non-transitive relations are represented by the 'rock-paper-scissors' game, in which rock crushes scissors, scissors cut paper, and paper wraps rock. In combination with spatial dispersal of static populations, this type of competition results in the stable coexistence of all species and the long-term maintenance of biodiversity. However, population mobility is a central feature of real ecosystems: animals migrate, bacteria run and tumble. Here, we observe a critical influence of mobility on species diversity. When mobility exceeds a certain value, biodiversity is jeopardized and lost. In contrast, below this critical threshold all subpopulations coexist and an entanglement of travelling spiral waves forms in the course of time. We establish that this phenomenon is robust; it does not depend on the details of cyclic competition or spatial environment. These findings have important implications for maintenance and temporal development of ecological systems and are relevant for the formation and propagation of patterns in microbial populations or excitable media.
Handbook of Computations for Biological Statistics of Fish Populations
Interaction rules affect species coexistence in intransitive networks
DOI:10.1890/10-0953.1
URL
PMID:21661578
[本文引用: 1]
Intransitive communities, those in which species' abilities cannot be ranked in a hierarchy, have been the focus of theoretical and empirical research, as intransitivity could help explain the maintenance of biodiversity. Here we show that models for intransitive competition embedding slightly different interaction rules can produce opposite patterns. In particular, we find that interactions in which an individual can be outcompeted by its neighbors, but cannot outcompete its neighbors, produce negative frequency dependence that, in turn, promotes coexistence. Whenever the interaction rule is modified toward symmetry (the individual and the neighbors can outcompete each other) the negative frequency dependence vanishes, producing different coexistence levels. Macroscopically, we find that asymmetric interactions yield highest biodiversity if species compete globally, while symmetric interactions favor highest biodiversity if competition takes place locally.
Higher order interactions and species coexistence
DOI:10.1007/s12080-009-0069-x
URL
PMID:25540673
[本文引用: 3]
Although parasites represent an important component of ecosystems, few field and theoretical studies have addressed the structure of parasites in food webs. We evaluate the structure of parasitic links in an extensive salt marsh food web, with a new model distinguishing parasitic links from non-parasitic links among free-living species. The proposed model is an extension of the niche model for food web structure, motivated by the potential role of size (and related metabolic rates) in structuring food webs. The proposed extension captures several properties observed in the data, including patterns of clustering and nestedness, better than does a random model. By relaxing specific assumptions, we demonstrate that two essential elements of the proposed model are the similarity of a parasite's hosts and the increasing degree of parasite specialization, along a one-dimensional niche axis. Thus, inverting one of the basic rules of the original model, the one determining consumers' generality appears critical. Our results support the role of size as one of the organizing principles underlying niche space and food web topology. They also strengthen the evidence for the non-random structure of parasitic links in food webs and open the door to addressing questions concerning the consequences and origins of this structure.
Intransitive competition is common across five major taxonomic groups and is driven by productivity, competitive rank and functional traits
Intransitive competition is widespread in plant communities and maintains their species richness
Structural stability: Concepts, methods, and applications
结构稳定性: 概念、方法和应用
Ecosystem effects of biodiversity manipulations in European grasslands
Understanding food-web persistence from local to global scales
DOI:10.1111/j.1461-0248.2009.01407.x
URL
PMID:19968697
[本文引用: 2]
Understanding food-web persistence is an important long-term objective of ecology because of its relevance in maintaining biodiversity. To date, many dynamic studies of food-web behaviour--both empirical and theoretical--have focused on smaller sub-webs, called trophic modules, because these modules are more tractable experimentally and analytically than whole food webs. The question remains to what degree studies of trophic modules are relevant to infer the persistence of entire food webs. Four trophic modules have received particular attention in the literature: tri-trophic food chains, omnivory, exploitative competition, and apparent competition. Here, we integrate analysis of these modules' dynamics in isolation with those of whole food webs to directly assess the appropriateness of scaling from modules to food webs. We find that there is not a direct, one-to-one, relationship between the relative persistence of modules in isolation and their effect on persistence of an entire food web. Nevertheless, we observe that those modules which are most commonly found in empirical food webs are those that confer the greatest community persistence. As a consequence, we demonstrate that there may be significant dynamic justifications for empirically-observed food-web structure.
Food-web constraints on biodiversity-ecosystem functioning relationships
Plant diversity and ecosystem productivity: Theoretical considerations
Insects on common reed (Phragmites australis): Community structure and the impact of herbivory on shoot growth
Predator avoidance and community structure: Interactions among piscivores, planktivores, and plankton
Freshwater snails habitat use in response to predation
Contrasting short-term and long-term effects of predation risk on consumer habitat use and resources
A spatially explicit model of sapling growth in a tropical forest: Does the identity of neighbours matter?
Consequences of trait changes in host-parasitoid interactions in insect communities
Stable coexistence in insect communities due to density- and trait-mediated indirect effects
The competitive structure of communities: An experimental approach with protozoa
Notice sur la loi que la population poursuit dans son accroissement
Variazioni e fluttuazioni del numero d'individui in specie animali conviventi
Food web structure and functioning: Theoretical advances and outlook
食物网结构与功能: 理论进展与展望
Identifying mechanisms of competition in multi-species communities
DOI:10.1111/jec.2007.95.issue-1 URL [本文引用: 1]
The ontogenetic niche and species interactions in size-structured populations
An experimental test of the effects of predation risk on habitat use in fish
A review of trait-mediated indirect interactions in ecological communities
Competition, predation, and the structure of the ambystoma-rana sylvatica community
Indirect effects and habitat use in an intertidal community: Interaction chains and interaction modifications
The nature and consequences of indirect effects in ecological communities
Putting the pieces together: Testing the independence of interactions among organisms
Higher-order interactions and indirect effects: A resolution using laboratory drosophila communities
A basal area increment model for individual conifers in the northern Rocky Mountains
Body size response to warming: Time of the season matters in a tephritid fly
Plant-mediated resource partitioning by coexisting parasitoids
DOI:10.1002/ecy.1834
URL
PMID:28370102
[本文引用: 1]
Although it has been frequently suggested that resource partitioning of species coexisting at the same trophic level can be mediated by interactions with species at non-adjacent trophic levels, empirical evidence supporting this claim is scarce. Here we demonstrate that plants may mediate resource partitioning for two parasitoids that share the same herbivorous host. The tephritid fly Tephritis femoralis is the primary pre-dispersal seed predator of two Asteraceae species, Saussurea nigrescens and Anaphalis flavescens, both of which dominate the plant community in the alpine meadows of the Tibetan Plateau. Field surveys and molecular barcoding analyses showed that the identity of the fly's main predator depended on the plant in which the fly developed. Tephritid flies that developed in S. nigrescens were preyed upon mainly by the parasitoid wasp Pteromalus albipennis, while the parasitoid Mesopolobus sp. was the main predator of flies that developed in A. flavescens. Microcosm experiments revealed that P. albipennis could not exploit the host flies within the capitula of A. flavescens due to food limitation (capitula are too small), while Mesopolobus sp. could not exploit the host flies within the capitula of S. nigrescens due to its inability to reach the host with its ovipositor (capitula are too large). Such bottom-up control of plant species traits may facilitate the coexistence of parasitoid wasps sharing a common host in this system. We suggest that interactions between non-adjacent trophic levels may potentially promote species coexistence and diversity in biological communities.
Higher-order interactions mitigate direct negative effects on population dynamics of herbaceous plants during succession
The complexity-stability relationship: Progress in mathematical models
复杂性-稳定性研究: 数学模型的进展
Interspecific synchrony of seed rain shapes rodent-mediated indirect seed-seed interactions of sympatric tree species in a subtropical forest
DOI:10.1111/ele.13405
URL
PMID:31631473
[本文引用: 1]
Animal-mediated indirect interactions play a significant role in maintaining the biodiversity of plant communities. Less known is whether interspecific synchrony of seed rain can alter the indirect interactions of sympatric tree species. We assessed the seed dispersal success by tracking the fates of 21 600 tagged seeds from six paired sympatric tree species in both monospecific and mixed plots across 4 successive years in a subtropical forest. We found that apparent mutualism was associated with the interspecific synchrony of seed rain both seasonally and yearly, whereas apparent competition or apparent predation was associated with interspecific asynchrony of seed rain either seasonally or yearly. We did not find consistent associations of indirect interactions with seed traits. Our study suggests that the interspecific synchrony of seed rain plays a key role in the formation of animal-mediated indirect interactions, which, in turn, may alter the seasonal or yearly seed rain schedules of sympatric tree species.
Mast seeding and its relationship with animal’s hoarding behaviour
植物大年结实及其与动物贮食行为之间的关系

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}