


雪豹(Panthera uncia)是起源于青藏高原的大型猫科动物, 现仅分布在包括中国在内的12个国家(Tseng et al, 2014; McCarthy et al, 2016)。作为中亚和青藏高原山地生态系统中的顶级捕食者, 雪豹对于维持食物网结构和生态系统稳定性有重要作用(Li et al, 2016)。
由于栖息地退化、猎物资源耗竭、偷猎盗猎以及人兽冲突引发的报复性猎杀等原因, 雪豹的生存面临着巨大的挑战(Li et al, 2013a; Mallon & Jackson, 2017)。据估计, 全球雪豹的野外种群数量仅为4,678-8,745只(McCarthy et al, 2016)。中国是雪豹适宜栖息地面积最大的国家, 雪豹在我国的分布区包括新疆、西藏、青海、甘肃、宁夏、内蒙古、四川、云南等8个省区, 种群数量约占全球雪豹野外种群数量的50% (Riordan & Shi, 2016)。
食性反映了动物对食物资源的利用, 准确获得其食物组成是充分了解该物种的生活习性和食物网关系的前提, 同时也是评估物种生存状况和生态系统功能的基础(刘刚等, 2018)。由于其栖息地多位于海拔3,000 m以上的高原山地, 崎岖度大, 加之雪豹行踪隐蔽以及种群密度低等原因, 难以通过直接观察其捕食行为获得食性数据。以往的雪豹食性分析多通过观察粪便样品中食物残余的形态学特征进行物种鉴定(Schaller et al, 1988; 刘楚光等, 2003)。已有研究显示雪豹是机会主义取食者, 其食谱组成受到当地食物资源多样性的影响, 但以山地有蹄类和家畜为主, 小型哺乳动物与鸟类为辅(Bagchi & Mishra, 2006; Wang et al, 2014; Kachel et al, 2016; Shrestha et al, 2018)。其中, 岩羊(Pseudois nayaur)和北山羊(Capra sibirica)是最常被报道的雪豹猎物物种, 它们的分布范围也与雪豹具有相当高的重叠程度。而其他有蹄类例如捻角山羊(Capra falconeri)、喜马拉雅塔尔羊(Hemitragus jemlahicus)和盘羊(Ovis spp.)等虽然在雪豹食谱中所占比例较小, 但是具有区域重要性(Lovari et al, 2009; Jumabay-Uulu et al, 2014)。
传统的粪便内容物形态学食物鉴定因操作简单、成本低而仍被广泛应用。但该方法完全依赖于对毛发、骨头等未消化食物硬质残留的形态特征进行分类, 对形态相近的物种难以准确鉴定, 对无硬质残留的食物(如肌肉、软组织)无法分析; 同时, 该方法要求研究者有丰富的形态学物种鉴定经验, 而物种鉴定结果难以重复验证, 因此鉴定效率较低, 结果准确率难以保证(Pompanon et al, 2012; Akrim et al, 2018)。
近年来, 随着分子生物学技术的不断发展, 基于DNA的分析技术被逐渐应用于生态学研究中。DNA条形码(DNA barcoding)技术通过较短的、标准化的DNA序列实现了在物种水平上对生物的识别和鉴定, 具有准确、灵敏、快速、自动及通用等特性, 在生物多样性研究领域显示出巨大的价值和应用潜力(Valentini et al, 2009; 裴男才和陈步峰, 2013)。DNA宏条形码(DNA metabarcoding)技术即利用高通量测序技术获得混合样本中多种条形码基因扩增子序列并进行归类的方法, 使分析复杂成分的生物混合样品成为可能。近年来, DNA宏条形码技术被应用于动物食性分析, 极大地推动了食性研究和精细食物网分析(Taberlet et al, 2012)。相对于形态学食性分析, DNA宏条形码技术具有以下主要的优势: 不依赖于研究者的经验和主观判断, 大幅提高了食物物种辨识的准确度和可信度; 不受食物消化程度和硬质残留比例的局限, 可以分析几乎所有食物类型; 一次可产生大量数据, 对于需要大规模样品分析的研究来说, 减少了单个样品的检验花费和所需投入的精力等(Pompanon et al, 2012)。基于以上优点, DNA宏条形码技术已被应用到包括雪豹在内的多种食肉动物的食性研究中(Shehzad et al, 2012a, b; Xiong et al, 2017)。
此前我国雪豹的研究积累主要集中在青海、新疆、西藏、甘肃, 而四川雪豹的研究还很少(唐卓等, 2017)。位于我国青藏高原东缘的四川中部至川西山地地处“中国西南山地”这一全球生物多样性热点地区(Myers et al, 2000), 由于海拔相对高差大, 这一地区存在着多种不同的气候垂直带谱和植被类型, 为不同的野生动物类群提供了适宜的栖息地环境。该地区也是全球大型食肉动物物种最为丰富的地区之一(Ripple et al, 2014)。对四川雪豹的生态学研究, 有助于了解和评估青藏高原东缘雪豹种群的生活习性和生存状况, 为该地区雪豹种群和生物多样性的保护提供科学依据。
本研究利用DNA宏条形码技术对卧龙雪豹粪便样品进行分子食性分析, 为进一步了解该地区雪豹种群捕食行为、生存状况以及制定有效的保护策略提供科学依据。
1 研究方法
1.1 研究地点
卧龙国家级自然保护区(102°52′-103°24′ E、30°45′-31°25′ N)位于四川省汶川县, 属青藏高原气候区的东缘, 四川盆地西缘, 邛崃山脉中部。区内海拔1,150-6,250 m, 相对高差达5,100 m, 植被随海拔上升表现出明显的垂直分布, 依次为常绿阔叶林带(< 1,600 m)、常绿落叶阔叶混交林带(1,600-2,000 m)、针阔混交林带(2,000-2,600 m)、针叶林带(2,600-3,800 m)、高山灌丛草甸带(3,800-4,400 m)和高山流石滩稀疏植被带(4,400-5,000 m) (陈利顶等, 1999; 施小刚等, 2017)。卧龙保护区于2009年首次在区内红外相机调查中记录到雪豹分布(Li et al, 2010), 这也是雪豹在邛崃山脉的首次野外实证记录。邛崃山脉地处雪豹全球分布区的东南缘(图1), 该区域的雪豹种群相对孤立, 同时也是我国研究和了解最少的雪豹种群之一。
表1 本研究使用的引物序列
Table 1
| 引物 Primer | 序列 Sequence (5′-3′) | 产物长度 Product length | 退火温度 Annealing temperature | 参考文献 Reference |
|---|---|---|---|---|
| 16S | F: GAGAAGACCCTATGGAGC | ~380 bp | 50℃ | Xiong et al, 2016 |
| R: ATAGAAACCGACCTGGAT | ||||
| 12SV5 | F: TAGAACAGGCTCCTCTAG | ~140 bp | 60℃ | Riaz et al, 2011 |
| R: TTAGATACCCCACTATGC | ||||
| UnciB | CTATGCTTAGCCCTAAACCTAGAT AGTTAGCTCAAACAAAACTAT-C3 | - | - | Shehzad et al, 2012a |
1.2 样品采集和DNA提取
雪豹粪便样品于2018年6月采集自卧龙保护区内(图1)。采样时, 由调查人员沿野外样线进行搜索, 根据样品的形态特征包括直径、长度以及团块数等进行野外物种预判, 通过手持GPS记录样品地理信息, 填写采集表。随后用一次性医用薄膜手套拾取粪便装入50 mL无菌离心管, 并进行编号。样品采集当天先加入95%乙醇浸泡, 24小时后弃乙醇, 加入硅胶干燥保存。样品带回实验室后转移至-20℃专用冰箱低温保存。
图1
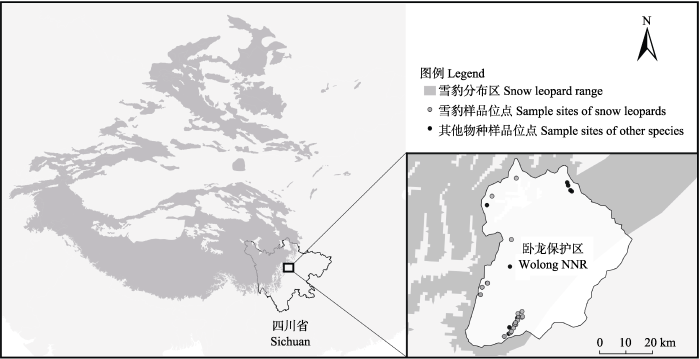
图1
四川省卧龙国家级自然保护区雪豹样品采集地点。雪豹分布区根据IUCN红色名录(http://dx.doi.org/10.2305/IUCN.UK. 2017-2.RLTS.T22732A50664030.en.)绘制。
Fig. 1
The locations of snow leopard feces collected in Wolong National Nature Reserve (NNR), Sichuan Province. Snow leopard range map is from the IUCN Red List of Threatened Species.
粪便样品的DNA提取使用2CTAB-PCI法(Vallet et al, 2007)。改进后的该方法适用于食肉动物粪便DNA提取(Xiong et al, 2016)。
1.3 粪便物种鉴定
利用引物16S-F/16S-R (表1)进行粪便线粒体DNA 16S rRNA基因片段的扩增, 扩增产物长度约为380 bp。PCR扩增体系为20 μL, 包括粪便DNA 20-40 ng、1×EasyTaq PCR SuperMix (AS111, 全式金生物技术有限公司)、16S-F/R各0.2 μM及0.4 mg/mL BSA。PCR反应程序为95℃预变性5 min; 95℃变性30 s, 50℃退火30 s, 72℃延伸30 s, 进行35个循环; 72℃延伸10 min。用纯化回收试剂盒(EP101, 全式金生物技术有限公司)对粪便DNA进行纯化, 纯化后的PCR产物由北京睿博兴科生物技术有限公司进行Sanger测序。测序结果用Chromas软件获得测序峰图, 除去杂峰部分, 得到约300 bp长度的DNA序列。通过NCBI的BLAST程序检索测序片段的匹配序列。结合研究点食肉动物物种信息, 当最匹配序列覆盖度为100%, 一致度在99%及以上, 且只对应一个物种, 并且该物种在研究地点有分布时, 认为粪便样品来自匹配序列对应的物种。
1.4 食性PCR扩增及高通量测序
1.4.1 食性分析的PCR产物扩增
由于残留在粪便样品中的食物DNA相对较少, 且存在降解和PCR抑制物等问题, 为提高食物 DNA扩增成功率, 扩增产物片段不宜过长。选用脊椎动物通用引物12SV5-F/12SV5-R (表1)进行食物DNA线粒体12S rRNA V5片段的扩增, 扩增片段长度约为140 bp。该对引物通用性广且具有较高的种属分辨率, 已应用于雪豹、豹猫(Prionailurus bengalensis)、亚洲金猫(Catopuma temminckii)等多种食肉动物的分子食性分析中(Shehzad et al, 2012a; Xiong et al, 2017)。此外, 在扩增不同样品时引物5′端增加由7个核苷酸组合而成的不同引物标签, 扩增同一样品的一对引物使用相同标签, 以区分后续混合样品高通量测序结果中的序列来源(Coissac, 2012)。
利用12SV5引物进行PCR扩增时, 能够同时扩增出捕食者(雪豹) DNA和食物DNA。为减少捕食者DNA竞争性扩增对食物DNA扩增效率的影响, 在PCR反应中加入浓度为引物20倍的雪豹DNA特异性阻抑引物UnciB (表1)以抑制雪豹DNA的扩增。有研究显示, 加入UnciB能够将雪豹DNA的扩增产物在总PCR产物中从87%降至26% (Shehzad et al, 2012a)。本研究预实验结果显示, 加入UnciB将雪豹PCR产物从77%降至37% (未发表数据)。
PCR扩增体系25 μL, 包含雪豹粪便DNA 20-40 ng、1 × EasyTaq PCR SuperMix、12SV5-F/R各0.2 μM、UnciB 2 μM及0.4 mg/mL BSA。PCR反应程序为95℃预变性5 min; 95℃变性30 s, 60℃退火30 s, 72℃延伸30 s, 进行35个循环; 72℃延伸10 min。同时在每批次扩增时加入粪便DNA提取空白和不加模板DNA的PCR空白作为对照以检验DNA污染情况。
1.4.2 高通量测序及数据处理
将所有粪便DNA样品的PCR产物进行纯化并混合, 由深圳华大基因科技有限公司进行测序。采用双端测序法(paired-end sequencing), 由Illumina HiSeq X Ten系统(Illumina Inc., San Diego, CA, USA)完成测序, 每端读长为150 bp。
测序结果使用OBITools程序包(Boyer et al, 2016;
序列的物种鉴定及分类原则包括: (1)比对结果一致度不低于98%时, 最匹配序列只对应单一物种, 且该物种在当地有分布时, 认为序列来自该物种; (2)当比对结果一致度不低于98%, 且最匹配序列对应多物种, 先排除当地无分布的物种, 如仍对应不止一种, 则鉴定结果记为涵盖这些物种的最小分类单元; (3)当比对结果一致度在95%-98%之间时, 统计最高一致度且本地分布的物种, 鉴定结果记为涵盖这些物种的最小分类单元; (4)舍弃一致度低于95%的序列; (5)合并差异小于2%的序列(Xiong et al, 2017)。
1.5 食性数据分析
对物种鉴定后的食物类群进行筛选与整理。当一个样品中某种食物序列数低于阈值(序列数低于50条或空白对照或该样品总序列数的1%), 舍弃该样品这一序列, 以排除潜在的交叉污染。此外捕食者序列及可能来自实验污染的人序列不计入食物类群。统计每份样品食物种类以及对应序列条数。
我们用出现频率(frequency of occurrence, FO)和相对序列丰度(relative read abundance, RRA)两个指标估计各食肉动物的食性构成(Deagle et al, 2019)。
FO是含某一食物类别的样品数占总样品数的百分比, 计算公式如下:
其中, Ni表示出现i类食物的样品数, N是有效样品总数。由公式可知, 当一份样品中存在一种以上食物时, 全部食物类别的FO之和大于100%。
RRA是某一食物类别的序列数在该样品总食物序列的百分比, 反映了相对生物量, 计算公式如下:
其中, T是食物类别数, N是有效样品总数, Si, j是食物类别i在样品j中的序列数。由公式可知, 全部食物类别的RRA之和为100%。
2 结果
2.1 粪便样品采集及鉴定
野外共采集疑似雪豹粪便样品38份, 粪便DNA提取及物种分子鉴定均得到有效结果, 成功率100%。其中22份(57.9%)粪便样品物种分子鉴定结果为雪豹, 其余粪便样品物种分子鉴定结果包括豹猫8份, 赤狐(Vulpes vulpes) 3份, 狼(Canis lupus) 2份, 石貂(Martes foina) 1份, 香鼬(Mustela altaica) 1份, 亚洲黑熊(Ursus thibetanus) 1份。22份雪豹粪便样品采集地点平均海拔4,179 m (最低3,401 m, 最高4,800 m) (图1)。
2.2 雪豹食性
22份卧龙雪豹粪便样品PCR产物高通量测序结果经OBITools处理后共获得1,886,353条序列。其中4份样品由于扩增失败或只有捕食者(雪豹)本身序列等原因未获得有效食性数据。有18份样品获得有效食性数据, 平均每份样品73,806条序列(3,477- 201,421条)。经进一步人工筛选和处理后, 共得到6种食物类别; 其中14份样品只有1种食物类别, 4份样品包含2种食物类别, 平均每份样品1.2种食物类别(表2)。6种食物类别分别是岩羊、牦牛(Bos grunniens)、高原鼠兔(Ochotona curzoniae)、红耳鼠兔(O. erythrotis)、鹡鸰科和雉科鸟类。其中岩羊在所有样品中总序列数为532,067条, 在NCBI GenBank数据库中最高一致度(identity)为100%; 牦牛总序列数为176,785条, 在数据库中最高一致度为100%; 高原鼠兔总序列数686条, 在数据库中最高一致度为100%; 红耳鼠兔总序列数6,496条, 在数据库中最高一致度为99%; 鹡鸰科总序列数465条, 在数据库中最高一致度为100%, 对应的潜在物种包括白鹡鸰(Motacilla alba)、灰鹡鸰(M. cinerea)以及树鹨(Anthus hodgsoni)等。雉科总序列数为1,689条, 在数据库中最高一致度为97%。
在所有食物类别中, 岩羊是雪豹食谱中最主要的成分, FO和RRA分别为66.7%和57.1%。此外, 家畜(牦牛)也是雪豹的重要取食对象, FO和RRA分别为33.3%和27.4%。同时, 高原鼠兔、红耳鼠兔、鹡鸰科和雉科在雪豹食性中占了较小的比例, 其序列各自仅在1份雪豹样品中有出现。各食物类别的FO和RRA统计值无显著差异(Wilcoxon符号秩检验, Z = -1.577, N = 6, P = 0.115)。
3 讨论
3.1 粪便物种鉴定
由于雪豹的行踪隐蔽以及种群密度低等原因, 对其行为生态的研究较为困难。野外痕迹调查和红外相机调查法(马鸣等, 2005; Li et al, 2013b; Alexander et al, 2015)是了解雪豹分布、种群数量和密度的主要方法, 但是无法回答雪豹食性等生态学问题。基于粪便DNA的分析方法为准确获得种群数量、遗传结构以及食物组成提供了有力的分析手段(Janecka et al, 2008; 周芸芸等, 2014; Shrestha et al, 2018; Chetri et al, 2019)。本研究使用的线粒体16S rRNA片段已被用于多种食肉动物的物种鉴定, 包括四川北部山地森林中的豹猫和亚洲金猫(Xiong et al, 2017)。本研究物种鉴定结果显示, 除雪豹外, 同域分布的食肉动物还包括狼、赤狐、亚洲黑熊、豹猫、石貂和香鼬, 与该区域红外相机调查的结果基本一致(施小刚等, 2017), 证实了该引物对食肉动物物种具有良好的分辨力。本研究中采集的38份粪便样品, 均由具有丰富野生动物监测经验的人员在野外根据粪便形态判定为雪豹粪便, 而分子物种鉴定发现其中仅有58%为雪豹粪便, 其余均为其他食肉动物的粪便。这提示依据粪便形态进行物种判定的准确率较低, 如果仅依靠此方法挑选样品并进行后续食性分析, 很可能会导致结果产生较大偏差。以往由于技术条件限制, 大部分粪便物种鉴定和食性分析均基于形态学特征。在今后的研究中, 使用分子遗传学方法进行非损伤样品(如粪便、毛发等)的物种鉴定应成为必需步骤, 以有效减少野外采集过程中物种误判的风险(Wang et al, 2014; Akrim et al, 2018)。
3.2 雪豹食物组成
与之前在我国青海省进行的雪豹食性研究结果相比(Schaller et al, 1988; 刘楚光等, 2003), 本研究发现卧龙地区雪豹主要以有蹄类为食物, 而较少依赖于中小型哺乳类(表3)。这可能与野生有蹄类(岩羊)和家畜(牦牛)在保护区中的数量较多有关, 这一结果也和青海三江源雪豹食性研究结果较为相似(李娟, 2012)。
从中亚及青藏高原周边其他国家雪豹食性研究结果来看, 除了中国新疆地区(家畜FO = 156.3%)及巴基斯坦北部Baltistan之外(家畜FO = 69.3%), 世界范围内雪豹食物以野生动物为主, 家畜为辅(FO = 0-38.5%) (表3)。在野生动物中, 又以有蹄类为主(种类因地区而异)。此外, 中小型哺乳类例如旱獭(Marmota spp.)和鼠兔也是雪豹较为重要的食物来源。本研究结果显示卧龙雪豹主要以野生动物为食, 野生动物中则又以岩羊为主(FO = 66.7%)。同时, 本研究显示鸟类在雪豹食性中也占据了少量比例, 据不完全统计, 卧龙保护区鸟类约16目318种, 其中地栖大型雉类12种(施小刚等, 2017)。在新疆地区的雪豹食性中鸟类(雉类)的出现频率(FO)甚至达到了81.3%, 这可能与当地鸟类物种多样性和种群数量有关。因此不同地区雪豹的食性结构可能与当地潜在食物类群的多样性与丰度密切相关。另外, 比较有意思的一个现象是雪豹粪便样品中偶尔会观察到有植物成分例如嫩枝等出现, 据推测植物可能为雪豹提供了无法从动物性食物中获取的矿物质或维生素等(Shehzad et al, 2012a), 也有研究认为植物可能在消化过程中起到了清理肠道的作用(Bocci et al, 2017)。本研究仅使用脊椎动物通用引物扩增食物成分, 因此无法鉴定雪豹食性中的植物成分, 后续研究可以通过使用植物特异引物扩增相应条形码区段, 以更全面地了解雪豹的食物构成。
在巴基斯坦Baltistan地区, 由于捕猎严重, 野生有蹄类已近灭绝, 家畜在雪豹食性中占主要地位(FO = 69.3%), 导致雪豹和人冲突较为严重(Anwar et al, 2011)。另外在新疆地区的研究也发现雪豹食性中包含大量家畜成分(家畜FO = 156.3%)。本研究发现卧龙雪豹食性虽然以野生动物为食, 但是家畜已经占到了较大的比例(FO = 33.3%), 低于新疆和Baltistan地区, 而远远高于我国青海地区(FO = 6.3-16.0%) (表3)。虽然基于食性分析无法判断雪豹食物中的家畜是来自于直接捕食家畜活体还是捡食家畜残骸(例如因病或自然死亡个体, 或被狼等其他食肉动物捕食后剩余的残骸), 但这一结果显
表3 不同地区雪豹食性研究结果比较
Table 3
| 研究地区 Study area | 样本数 No. of samples | 物种鉴定 方法 Species identification | 食性分析 方法 Diet analysis | 食物种类(出现频率) Prey taxa (FO) (%) | 参考文献 References | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 野生有 蹄类 Wild ungulates | 中小型 哺乳类 Small mammals | 鸟类 Birds | 家畜 Livestock | 植物 Plant | 未鉴定 Unidentified items | |||||
| 巴基斯坦 Baltistan, Pakistan | 49 | DNA | 形态学 Morphology | 24.5 | 6.1 | 4.2 | 69.3 | 59.2 | 32.6 | Anwar et al, 2011 |
| 蒙古 South Gobi, Mongolia | 81 | DNA | DNA | 79.0 | - | 1.2 | 19.8 | - | - | Shehzad et al, 2012a |
| 尼泊尔 Manang, Nepal | 213 | 形态学 Morphology | 形态学 Morphology | 51.6 | 53.5 | 1.4 | 17.8 | - | - | Oli et al, 1994 |
| 尼泊尔 PinValley, Nepal | 51 | 形态学 Morphology | 形态学 Morphology | 56.9 | 3.9 | - | 23.6 | 25.5 | 5.9 | Bagchi & Mishra, 2006 |
| 尼泊尔 Kibber, Nepal | 44 | 形态学 Morphology | 形态学 Morphology | 29.6 | 6.8 | 15.9 | 38.5 | 27.3 | 19.5 | Bagchi & Mishra, 2006 |
| 塔吉克斯坦 Murghab, Tajikistan | 36 | DNA | 形态学 Morphology | 78.0 | 45.0 | - | - | - | - | Kachel et al, 2016 |
| 塔吉克斯坦 Madiyan, Tajikistan | 20 | DNA | 形态学 Morphology | 50.0 | 55.0 | - | 2.0 | - | - | Kachel et al, 2016 |
| 印度 Ladakh, India | 173 | 形态学 Morphology | 形态学 Morphology | 35.2 | 25.5 | 4.6 | 22.1 | 60.7 | - | Chundawat & Rawat, 1994 |
| 中国青海 Qinghai, China | 206 | 形态学 Morphology | 形态学 Morphology | 37.6 | 44.3 | - | 6.3 | 8.0 | 1.4 | Schaller, 1988 |
| 中国青海 Qinghai, China | 307 | 形态学 Morphology | 形态学 Morphology | 36.9 | 58.9 | 2.3 | 9.8 | 2.9 | 34.2 | Liu et al, 2003 |
| 中国青海 Qinghai, China | 250 | 形态学 Morphology | 形态学 Morphology | 67.6 | 17.6 | - | 16.0 | 18.0 | 6.4 | Li, 2012 |
| 中国新疆 Xinjiang, China | 16 | DNA | 形态学 Morphology | 50.0 | 112.5 | 81.3 | 156.3 | 25.0 | - | Wang et al, 2014 |
| 中国四川 Sichuan, China | 18 | DNA | DNA | 66.7 | 11.1 | 11.1 | 33.3 | - | - | 本研究 This study |
示卧龙雪豹对区内散放家畜的依赖度较高, 可能在当地社区中已存在相当程度的人兽冲突。已有大量的研究结果显示, 人兽冲突带来的经济损失和安全威胁, 会导致当地社区对雪豹等“肇事”动物产生负面印象、容忍度下降; 如果缺乏适当的管理措施, 人兽冲突可逐渐激烈化, 进而导致对“肇事”动物的报复性猎杀或清除(Messmer, 2000)。因此, 在卧龙及周边的雪豹分布区内, 亟需对雪豹导致的人兽冲突现状进行系统评估, 以制定有针对性的保护和管理策略来缓解冲突, 包括逐步减少保护区内散放家畜的数量以及为家畜设立保险等, 避免冲突激化对野生雪豹种群带来严重负面影响。
3.3 DNA条形码分辨率
本研究使用脊椎动物线粒体12S rRNA基因的 V5区段作为识别食物物种的条形码, 共鉴定出6个食物类别, 其中4个类别鉴定到物种水平, 2个类别鉴定到科水平。鉴定到科的两个类别均为鸟类, 说明该条形码在鸟类中种间差异小, 物种分辨度有限, 这种情况在利用同一条形码在四川北部进行的豹猫食性研究中也有发现(Xiong et al, 2017)。如鉴定为鹡鸰科的序列在GenBank数据库中对应当地多种鹡鸰科的鸟类, 且最高一致度均为100%, 因此只能将该食物类别记为鹡鸰科。而鉴定为雉科的序列在GenBank数据库中最高一致度仅为97%, 且对应物种灰孔雀雉(Polyplectron bicalcaratum)在当地无分布, 因此将鉴定结果记为雉科。已知在当地雪豹栖息地内有分布的雉科物种包括血雉(Ithaginis cruentus)、绿尾虹雉(Lophophorus lhuysii)、藏雪鸡(Tetraogallus tibetanus)等, 是否为这些物种之一或其他近缘未知物种仍有待进一步验证。如需对食性中的鸟类进行更精确的判定, 可采用鸟类特异的引物扩增进行后续分析。此外, 条形码数据库中物种序列信息的全面程度, 也将影响鉴定结果的准确性。由于本研究使用的12SV5区段在GenBank中仅收录了高原鼠兔和红耳鼠兔序列, 其他卧龙保护区分布的鼠兔种例如藏鼠兔(Ochotona thibetana)、间颅鼠兔(O. cansus)等的相应序列并未收录, 因此虽然物种鉴定结果为高原鼠兔和红耳鼠兔, 但是不排除为其他鼠兔种的可能。
3.4 食性数据统计方法比较
我们的研究结果显示, 利用食物出现频率FO和相对序列丰度RRA得到的雪豹食性结果并未显示明显差异(表2)。唯一出现较大差异的是高原鼠兔这一食物类别, 其FO为5.5%, 而RRA为0.5%。这一差别可能是由于FO反映的是某一食物类别出现的频次, 因此会增加生物量贡献较低的食物类别的比例。而RRA利用食物扩增序列数与原始生物量组成相关的关系, 反映了相对生物量, 对高通量食性分析提供了更多信息, 具有潜在的应用价值。
表2 卧龙保护区雪豹粪便样品(N = 18)分子食性分析结果
Table 2
| 食物类别 Prey taxa | 出现样品数 No. of occurrence | 出现频率 FO (%) | 相对序列丰度 RRA (%) |
|---|---|---|---|
| 岩羊 Pseudois nayaur | 12 | 66.7 | 57.1 |
| 牦牛 Bos grunniens | 6 | 33.3 | 27.4 |
| 高原鼠兔Ochotona curzoniae | 1 | 5.5 | 0.5 |
| 红耳鼠兔Ochotona erythrotis | 1 | 5.5 | 5.6 |
| 鹡鸰科Motacillidae | 1 | 5.5 | 5.6 |
| 雉科 Phasianidae | 1 | 5.5 | 3.8 |
| 总计 Total | 100 |
3.5 个体食性分析
根据样品采集地点的覆盖面积及在该区域内红外相机的监测数据, 本研究的样品应来自于多只雪豹个体。由于研究条件所限, 本研究并未对每份粪便样品利用分子遗传学方法进行个体鉴定, 因此难以准确评估样品来源个体数量及不同个体的样品在总体样品中的份额。在今后研究中可以引入粪便样品个体识别技术, 针对不同个体进行食性分析, 从而更深入地了解雪豹的食物选择和捕食策略。
4 结论
本研究将基于粪便DNA的食肉动物物种鉴定与分子食性分析相结合, 对采自我国四川卧龙的雪豹粪便样品进行高通量分子食性分析, 表明野生猎物尤其是岩羊是卧龙雪豹的主要食物资源, 然而家畜(牦牛)在雪豹食物中也占了较高比例, 因此需要对该区域由雪豹捕食散放家畜引发的潜在人兽冲突问题给予关注, 制定相应的管理与保护措施, 以确保这一相对孤立的雪豹区域种群能够长期延续, 维持高山生态系统的结构与功能完整性。
致谢: 感谢卧龙自然保护区全体参与野外调查采样的工作人员的鼎力协助。
(责任编委: 李明 责任编辑: 时意专)
参考文献
Assessment of bias in morphological identification of carnivore scats confirmed with molecular scatology in north-eastern Himalayan region of Pakistan
Face value: Towards robust estimates of snow leopard densities
Food habits of the snow leopard Panthera uncia (Schreber, 1775) in Baltistan, Northern Pakistan
Living with large carnivores: Predation on livestock by the snow leopard (Uncia uncia)
Sympatric snow leopards and Tibetan wolves: Coexistence of large carnivores with human-driven potential competition
OBITOOLS: A unix-inspired software package for DNA metabarcoding
Evaluation on giant panda habitat fragmentation in Wolong Nature Reserve
卧龙自然保护区大熊猫生境破碎化研究
Estimating snow leopard density using fecal DNA in a large landscape in north-central Nepal
Food habits of the snow leopard in Ladakh. In: International Snow Leopard Trust and Northwest Plateau Institute of Biology (eds Fox JL, Jizeng D), pp.
OligoTag: A program for designing sets of tags for next-generation sequencing of multiplexed samples
Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data?
Population monitoring of snow leopards using noninvasive collection of scat samples: A pilot study
Large carnivores and low diversity of optimal prey: A comparison of the diets of snow leopards Panthera uncia and wolves Canis lupus in Sarychat-Ertash Reserve in Kyrgyzstan
Investigating the potential impact of trophy hunting of wild ungulates on snow leopard Panthera uncia conservation in Tajikistan
Ecology and Conservation Strategy of Snow Leopard (Panthera uncia) in Sanjiangyuan Area on the Tibetan Plateau
.
青藏高原三江源地区雪豹(Panthera uncia)的生态学研究及保护
.
a) Human-snow leopard conflicts in the Sanjiangyuan Region of the Tibetan Plateau
b) A communal sign post of snow leopards (Panthera uncia) and other species on the Tibetan Plateau, China
Challenges of snow leopard conservation in China
Cats living with pandas: The status of wild felids within giant panda range, China
Research on foods and foods source about snow leopard (Panthera uncia)
雪豹的食性与食源调查研究
The application of high-throughput sequencing technologies to wildlife diet analysis
高通量测序技术在野生动物食性分析中的应用
Restoring a keystone predator may endanger a prey species in a human-altered ecosystem: The return of the snow leopard to Sagarmatha National Park
Markings as indicator of snow leopard in field survey, in Xinjiang
新疆雪豹调查中的痕迹分析
A downlist is not a demotion: Red List status and reality
What is a snow leopard? Biogeography and status overview. In: Snow Leopards (eds McCarthy T, Mallon D), pp.
The emergence of human-wildlife conflict management: Turning challenges into opportunities
Biodiversity hotspots for conservation priorities
Snow leopard Panthera uncia predation of livestock: An assessment of local perceptions in the Annapurna Conservation Area, Nepal
DNA barcoding of life: A classification of uses according to function and scale after ten years of development
生物DNA条形码: 十年发展历程、研究尺度和功能
Who is eating what: Diet assessment using next generation sequencing
ecoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis
China: Current state of snow leopard conservation in China. In: Snow Leopards (eds McCarthy T, Mallon D), pp.
Status and ecological effects of the world’s largest carnivores
Status of the snow leopard Panthera uncia in Qinghai and Gansu provinces, China
a) Prey preference of snow leopard (Panthera uncia) in South Gobi, Mongolia
b) Carnivore diet analysis based on next-generation sequencing: Application to the leopard cat (Prionailurus bengalensis) in Pakistan
Camera-trapping surveys of the mammal and bird diversity in Wolong National Nature Reserve, Sichuan Province
利用红外相机调查四川卧龙国家级自然保护区鸟兽多样性
Diet and prey selection by snow leopards in the Nepalese Himalayas
Towards next-generation biodiversity assessment using DNA metabarcoding
Research on snow leopards (Panthera uncia) using camera-trapping in Wolong National Nature Reserve, China
基于红外相机技术对四川卧龙国家级自然保护区雪豹(Panthera uncia)的研究
Himalayan fossils of the oldest known pantherine establish ancient origin of big cats
DNA barcoding for ecologists
A new 2CTAB/PCI method improves DNA amplification success from faeces of Mediterranean (Barbary macaques) and tropical (lowland gorillas) primates
Dietary overlap of snow leopard and other carnivores in the Pamirs of Northwestern China
Molecular dietary analysis of two sympatric felids in the mountains of Southwest China biodiversity hotspot and conservation implications
Molecular analysis of vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) in southwest China
Population survey and genetic diversity of snow leopards Panthera uncia as revealed by fecal DNA
基于粪便DNA的青藏高原雪豹种群调查和遗传多样性分析

{kind=link}
{kind=link}