


功能性状(functional trait)是指在个体水平上能够影响植物生长、繁殖和存活能力, 且能对生态系统的功能产生影响以及对植物周围环境进行响应的核心属性(Díaz et al, 1998, 2010; Violle et al, 2007)。植物功能性状可有效表达在漫长的进化过程中植物对环境异质性的适应程度, 反映植物生长、繁殖及利用资源的对策(Garnier et al, 2004; Bu et al, 2017), 可在不同区域尺度内反映植物与环境因子的关系(Mcintyre et al, 1999)。植物的功能性状之间可通过相互影响、共同作用来体现对环境的适应程度(Osnas et al, 2018)。
竹类是世界植物资源和森林资源的重要组成部分, 是一类集经济、生态、观赏于一体的多功能典型类群, 在我国南方林业中占有举足轻重的地位。竹类植物按其地下茎结构及竹秆群集程度分为合轴丛生、单轴散生和复轴混生三大类, 这三大类在中国均有分布(李睿等, 2003)。竹类植物的系统演化是由合轴丛生型到复轴混生型, 再到单轴散生型(江泽慧, 2002), 这个过程反映了竹类植物通过构型的演化来适应复杂多变的环境并获取外界资源(高贵宾等, 2016)。而从丛生竹至混生竹, 再至散生竹的进化过程中, 哪些功能性状产生了分化值得关注。竹笋是竹子的营养繁殖体, 竹类植物的生长季节性较强, 故常有四季笋竹出现(毛丹和陈建华, 2005)。竹子在自然状态下的出笋期普遍只有1-2个月, 最多可延长至5个月(代凤贵, 2014)。不同竹类植物的出笋期各不相同, 出笋持续时间也有差异(钟远标等, 2014)。而不同出笋季节的竹类植物在功能性状上有何差异仍知之甚少。
特有现象是植物地理学的重要研究内容, 对特有现象的研究有利于对物种起源、种系分化和演化过程有更深入的理解(冯建孟和朱有勇, 2010)。中国特有属常常被认为是只分布在中国或者分布中心在中国, 且分布范围未超出国界太多的属(吴征镒 2003)。其中, 中国特有的竹类植物有371种, 隶属于23属, 占中国所有竹类植物的68.96%。而中国特有竹类植物与非中国特有竹类植物在生物学特性上的差异研究也很少。
竹类植物的生长和繁殖具有独特性(不受年龄因素的影响): 首先, 竹类植物是一种重要的克隆植物, 父代和子代间往往具有相同的基因型, 遗传结构相对稳定(宋明华和董鸣, 2002); 其次, 竹类植物没有次生生长, 虽然外形似树, 但其生长过程同树木有很大区别, 木本竹类的笋在短短几个月便可长成新竹, 完成秆的生长(汪阳东, 2001)。鉴于竹类植物独特的生长和繁殖特性, 其功能性状值较为稳定, 在Flora of China (Wu et al, 2006)中的描述也相对准确。因而, 本文选取中国竹类植物作为研究对象, 通过收集Flora of China中竹类植物的生物学特性描述, 主要探讨以下3个问题: (1)不同地下茎类型的竹类植物在系统演化进程中哪些功能性状优先进行分化; (2)中国特有种和非中国特有种在形态学上有何差异; (3)不同出笋季节的竹类植物在形态学上产生何种适应。
1 材料与方法
1.1 研究区域概况
中国是世界竹类植物的起源和分布中心之一, 是竹资源最丰富的国家, 拥有竹类植物共34属500多种, 竹林面积520万ha, 约占世界竹林总面积的1/4, 被誉为“竹子王国” (陈松河, 2009; 阳斐和辉朝茂, 2014)。中国竹类植物主要产于秦岭-淮河流域以南地区, 自然分布范围极广, 既有适宜生长在南方热带地区的合轴丛生竹, 也有在亚热带地区广泛分布的单轴散生竹, 亦有在高纬度高海拔地区生长良好的复轴混生竹(陈其兵, 2007)。竹类植物喜爱温暖潮湿的气候, 适宜生长在土壤呈微酸性的平原、盆地、山地、丘陵、高原等地带。
1.2 数据来源与方法
中国竹类植物的基本形态学功能性状数据主要来自Flora of China, 并参考了其他地方植物志(江苏省中国科学院植物研究所, 1977; 林泉, 1993; 易同培, 2008)和一些期刊论文(卢志军等, 2009; Huang et al, 2011; 冀琳珂等, 2016)。竹类植物属于禾本科竹亚科, Flora of China共收录了34属534种, 其中丛生竹245种, 混生竹108种, 散生竹181种; 中国特有种371种, 非中国特有种163种。由于有关竹类植物出笋季节的数据不全, 目前仅收集了359种具有出笋时段的竹类植物, 其中春季出笋的180种, 夏季出笋的130种, 秋季出笋的42种, 冬季出笋的7种。
研究选取的具体功能性状指标包括秆高、直径、节间长、竹壁厚、叶长和叶宽。其中秆高是指植物主要光合组织(不包括花序)的上边界与地面之间的最短距离, 与物种的生长形态、物种在植被垂直光梯度中的位置、竞争活力、生殖能力、全株繁殖力、潜在寿命息息相关(Perez-Harguindeguy et al, 2016)。竹秆的直径代表了竹类植物的粗细, 与其重量和材积息息相关; 节间长是指相邻两个竹秆节部之间的距离, 竹秆节部不仅具有机械支撑作用, 而且物质的横向交流也主要发生在节部, 由于遗传结构的作用, 同一种竹子的节数通常趋于一常数, 因而节间长也与遗传有关(周芳纯, 1998); 竹壁厚也是构成秆形结构的主要因子之一, 与竹子的重量和材积密切相关, 壁厚不同的竹材利用率大不相同(汪阳东, 2001); 叶面积是指植物叶片进行光合作用的最小面积单元, 叶面积主要由叶长与叶宽决定, 是衡量叶子大小的最常用指标, 植物会通过叶片性状的改变实现不同环境条件下正常的光合作用和基本的植物功能(唐青青等, 2016)。
此次研究通过收集Flora of China中秆高、直径、节间长、竹壁厚、叶长和叶宽的基本形态学功能性状指标的描述, 获取其取值范围, 包括最大值、最小值、均值等。若只描述了形态学指标的最大值和最小值, 则取最大值和最小值的平均值作为功能性状的取值, 本研究中所有功能性状取值均为该功能性状的均值。
1.3 数据分析
为了探讨不同功能群竹类植物在基本形态学性状上的差别, 通过地下茎类型(丛生、混生和散生)、特有性分布(中国特有种与否)和出笋季节(春、夏、秋和冬)划分不同功能群竹类植物, 利用单因素方差分析和多重比较探讨不同类型竹类植物的差异性(显著性检验采用多重比较Tukey法); 通过Pearson相关系数揭示各个功能性状之间的相关性。 为了揭示不同类群竹类植物功能性状变异的贡献率, 对不同类群竹类植物进行主成分分析(PCA), 提取各个性状指标的因子载荷值, 探讨不同功能群竹类植物功能性状的变异。所有数据分析与作图均使用R 3.1.1的基础包与vegan包完成。
2 结果
2.1 竹类植物的功能性状差异
通过534种竹类植物6个功能性状的描述性统计可知(表1), 变异系数最大的为直径(CV = 104%), 最小的为节间长(CV = 45%)。6个功能性状的变异程度依次为直径 > 竹壁厚 > 叶宽 > 秆高 > 叶长 > 节间长。
表1 竹类植物6个功能性状的描述性统计
Table 1
| 平均Mean | 最小值Minimum | 最大值Maximum | CV (%) | |
|---|---|---|---|---|
| 秆高 Culm height (m) | 6.36 | 0.3 | 40 | 75 |
| 直径 Diameter (cm) | 3.16 | 0.15 | 25 | 104 |
| 节间长 Internode length (cm) | 29.04 | 0.75 | 90 | 45 |
| 竹壁厚 Wall thickness (mm) | 5.22 | 0.16 | 32.5 | 95 |
| 叶长 Leaf length (cm) | 14.35 | 0.95 | 45 | 48 |
| 叶宽 Leaf width (cm) | 2.19 | 0.2 | 20.5 | 94 |
不同地下茎类型的竹类植物之间主要功能性状存在显著差异(图1)。从秆高、直径和节间长来说, 丛生的竹类植物最大, 散生的竹类植物次之, 而混生的竹类植物最小。丛生的竹类植物具有最大的竹壁厚, 与其他两类存在显著差异, 而混生和散生的竹类植物之间则无显著差异。混生的竹类植物具有最大的叶长, 丛生的竹类植物次之, 散生的竹类植物最小, 并且它们之间均存在显著差异。丛生和混生的竹类植物具有较大的叶宽, 但差异不显著, 而散生的竹类植物则具有较小的叶宽, 并与其他两类均存在显著差异。综合来看, 丛生的竹类植物具有最大的秆高、直径、节间长、竹壁厚, 较大的叶长和叶宽; 混生的竹类植物具有最小的秆高、直径、节间长、竹壁厚, 最大的叶长和叶宽; 而散生的竹类植物则具有中等的秆高、直径、节间长、竹壁厚和最小的叶长与叶宽。
图1
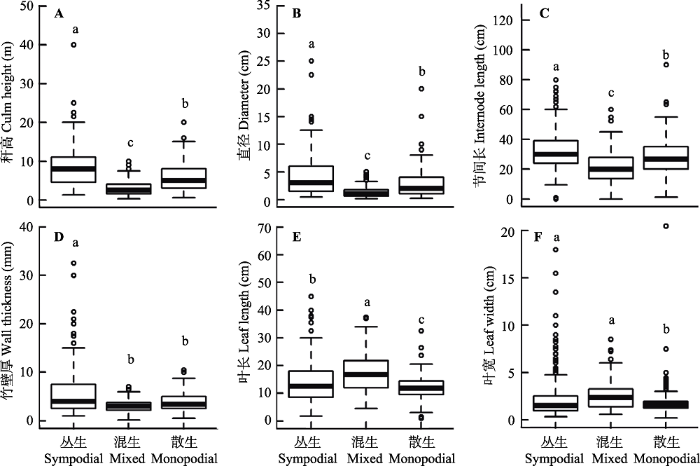
图1
不同地下茎类型竹类植物的形态学性状差异。不同字母表示差异显著(P < 0.05)。
Fig. 1
Variations in morphological traits of bamboo plants with different rhizome types. Boxes with different letters differ significantly at P < 0.05.
中国特有种与非中国特有种竹类植物的功能性状之间存在显著差异(图2)。相较于非中国特有种, 中国特有种具有较小的秆高、直径、竹壁厚和叶长。尽管在均值上中国特有种的节间长和叶宽均小于非中国特有种, 但无显著差异。总体说来, 与非中国特有种相比, 中国特有种往往具有较小的功能性状值。
图2
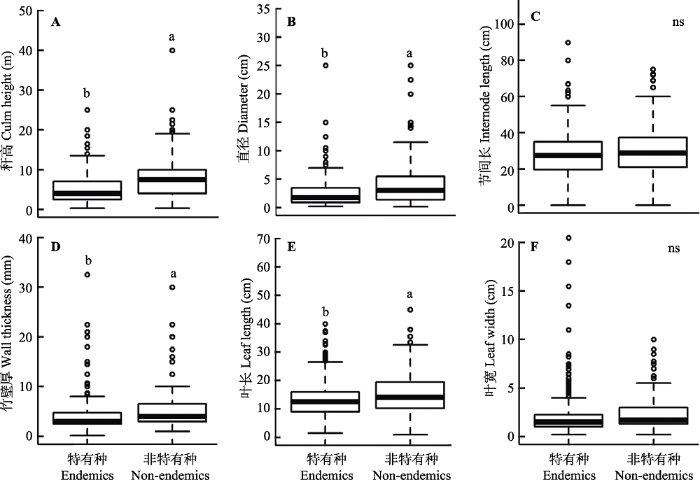
图2
不同特有性分布竹类植物的形态学性状差异。不同字母表示差异显著(P < 0.05)。
Fig. 2
Variations in morphological traits of bamboo plants with different endemism. Boxes with different letters differ significantly at P < 0.05.
从出笋季节来看, 各个出笋季节的竹类植物在秆高、直径、节间长、竹壁厚、叶长和叶宽方面均不存在显著差异(图3)。
图3
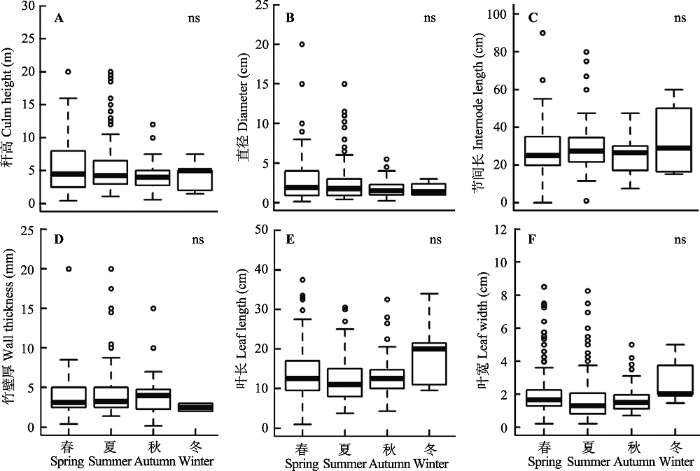
图3
不同出笋季节竹类植物的形态学性状差异。ns表示无显著性差异。
Fig. 3
Variations in morphological traits of bamboo plants with different shooting season. ns means no significant difference.
2.2 竹类植物功能性状变异的多样性
相关性分析结果(图4)表明: 竹类植物的秆高与直径存在很强的相关性(r > 0.9), 同时与竹壁厚、节间长也存在较大的相关性, 而与叶长、叶宽则不相关; 直径与节间长、竹壁厚、叶长存在较强的相关性, 而与叶宽不相关; 节间长与竹壁厚存在一定的相关性, 而与叶长、叶宽则不相关; 竹壁厚与叶长、叶宽均不相关; 叶长与叶宽存在显著的相关性。总体来说, 秆高、直径、节间长、竹壁厚之间均呈显著正相关, 相关系数r均大于0.29, 但它们与叶长、叶宽的相关性较小, 尽管叶长和叶宽之间存在显著的正相关。
图4
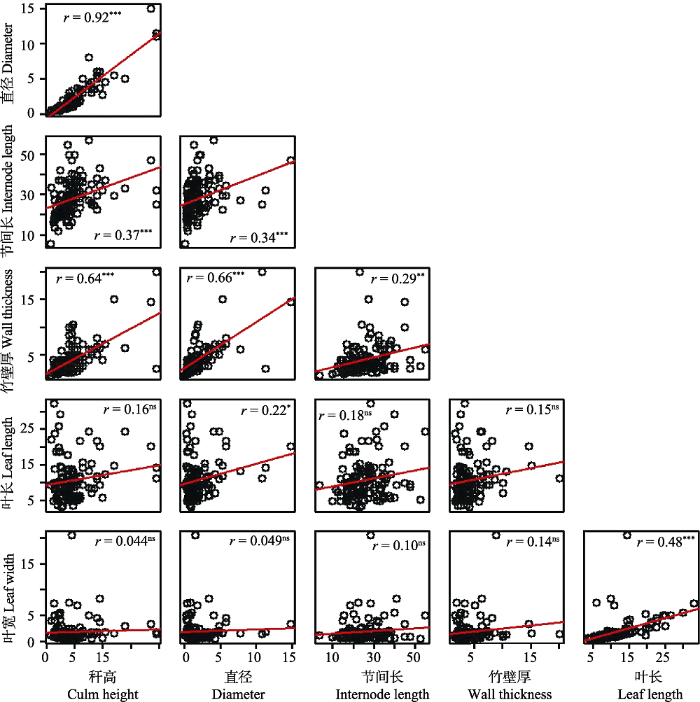
图4
竹类植物功能性状之间的相关性。横纵坐标代表对应性状的数值。* P < 0.05, ** P < 0.01, *** P < 0.001, ns表示无显著性差异。
Fig. 4
Correlations among functional traits of bamboo plants. The horizontal and vertical coordinates represent the values of traits. * P < 0.05, ** P < 0.01, *** P < 0.001, ns means no significant difference.
对不同类群竹类植物的功能性状指标进行主成分分析发现: 第一主轴PCA1能够反映功能性状变异的40%-73% (表2), 代表了竹类植物功能性状变异的绝大部分。对全部中国竹类植物的功能性状指标进行主成分分析发现(图5): 第一主成分(PCA1)的贡献率为52.56%, 第二主成分的贡献率为17.93%, 基本能够反映各个形态学指标的信息。其中第一主成分主要表征了“大小性状”, 包括竹类植物的直径、秆高和竹壁厚; 第二主轴主要是叶片性状, 包括叶长和叶宽。丛生竹的功能性状主要分布在第一主轴左侧, 具有最大的直径和秆高。混生竹的功能性状集中在第二主轴下侧, 故具有最大的叶长和叶宽。散生竹的功能性状一般集中在第一主轴和第二主轴的中心, 具有较为中等的性状值。
表2 不同类型竹类植物功能性状的主成分分析(PCA)表
Table 2
| 地下茎类型 Rhizome type | 特有种 Endemic species | 出笋季节 Shooting season | 综合 Overall | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 丛生 Sympodial | 混生 Mixed | 散生 Monopodial | 是 Yes | 否 No | 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | ||
| PCA1累计贡献率 PCA1 Cumulative variance | 60.75% | 46.59% | 40.19% | 52.01% | 55.25% | 42.26% | 45.58% | 49.37% | 73.69% | 52.56% |
| 排名前三的性状及因子载荷值 Top 3 traits and factor loadings | 直径 Diameter | 竹壁厚 Wall thickness | 秆高 Culm height | 秆高 Culm height | 直径 Diameter | 秆高 Culm height | 直径 Diameter | 直径 Diameter | 节间长 Internode length | 直径 Diameter |
| -1.93 | 1.26 | -1.87 | -2.03 | 1.73 | -1.85 | -1.64 | -1.21 | -0.76 | -2.21 | |
| 叶长 Leaf length | 直径 Diameter | 直径 Diameter | 直径 Diameter | 叶长 Leaf length | 直径 Diameter | 秆高 Culm height | 秆高 Culm height | 秆高 Culm height | 秆高 Culm height | |
| -1.87 | 1.24 | -1.79 | -2.02 | 1.65 | -1.78 | -1.62 | -1.18 | -0.74 | -2.11 | |
| 秆高 Culm height | 秆高 Culm height | 节间长 Internode length | 竹壁厚 Wall thickness | 秆高 Culm height | 节间长 Internode length | 叶长 Leaf length | 竹壁厚 Wall thickness | 直径 Diameter | 竹壁厚 Wall thickness | |
| -1.77 | 1.22 | -1.45 | -1.87 | 1.53 | -1.21 | -1.48 | -1.16 | -0.73 | -1.95 | |
图5
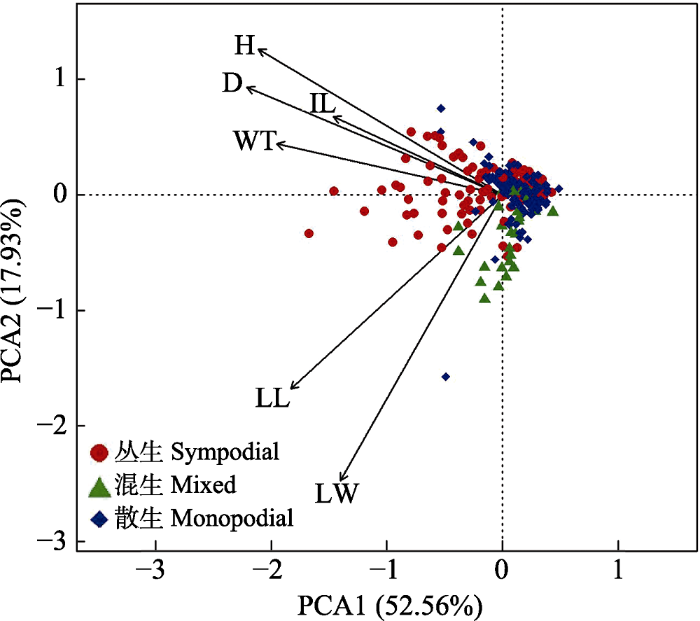
图5
中国竹类植物功能性状主成分分析。H: 秆高; D: 直径; IL: 节间长; WT: 竹壁厚; LL: 叶长; LW: 叶宽。
Fig. 5
PCA of functional traits of Chinese bamboo plants. H, Culm height; D, Diameter; IL, Internode length; WT, Wall thickness; LL, Leaf length; LW, Leaf width.
3 讨论
3.1 不同功能群竹类植物功能性状的变异
从地下茎类型来看, 丛生竹、散生竹和混生竹之间在功能性状上通常存在显著差异。丛生竹具有最大的秆高、直径、节间长和竹壁厚, 散生竹次之, 而混生竹最小。因而, 由合轴丛生型到复轴混生型的系统演化过程中, 大小性状趋于减小, 叶片性状趋于变大; 而由复轴混生型到单轴散生型的系统演化过程中, 大小性状变大, 叶片性状变小。竹类植物的秆高与竹子的节间数量和节间长度息息相关。一般来说, 丛生竹的根系发达, 有较高的生物量, 虽然成丛生长, 但是竹秆形态高大(高素萍等, 2002; 陈宝昆等, 2007)。丛生竹的节间数量较多。中小型散生竹具有较短的节间伸长区以及较少的节间数量, 故和丛生竹相比秆高较小。混生竹具有最大的叶长和叶宽, 散生竹最小, 丛生竹的叶长和叶宽介于两者之间。叶片大小可以反映植物对光能的截取以及对碳的获取能力(Milla & Reich, 2007)。光照强度越强, 植物叶片细胞分裂的速度越快。可能由于混生竹既有横向分布的地下茎, 又有密集分布的竹丛, 混生的竹类植物在进行幼竹高生长的过程中即抽枝展叶(Milla & Reich, 2007), 早于散生竹和丛生竹。与丛生竹和散生竹相比, 混生竹获得光照的时间更长, 故有最大的叶长和叶宽。丛生竹由于没有长距离的横向竹鞭, 竹秆分布密集, 多成丛生长, 因此较大的竹叶有利于对光源的捕捉与吸收, 从而减少光照限制对丛生竹生长的影响。散生的竹类植物竹鞭横向分布, 一年可生长2-5 m, 其出笋成竹较为分散, 叶片虽小但仍可以获取充足的光照。
从特有性分布来看, 中国特有种在秆高、直径、竹壁厚、叶长方面显著小于非特有种, 而在节间长及叶宽方面则无显著差异, 中国特有种的功能性状相对较小。因为植物特有现象的出现既受到物种自身遗传条件的显著影响, 还会受到周围自然条件的影响, 两者相互作用(张宏达, 1997)。植物的特有性现象只局限于特定区域内, 特有种植物的分布区较为狭窄(Anderson, 1994), 其适应的环境类型也有一定的局限性。植物功能性状与环境息息相关, 可以通过功能性状的改变来适应外部环境(Wright et al, 2007)。为了更好地生存, 植物体会将更多的能量分配给繁殖和生长, 从而形成一系列经济型的器官结构, 如叶片的光合速率较高且比叶面积较大、植株茎干生长迅速而木质密度较低等(刘晓娟和马克平, 2015)。与中国特有种相比, 非中国特有种的功能性状需要适应更为普遍的环境, 故具有较大的性状值。
3.2 竹类植物功能性状变异的多样性
本研究发现, 不同功能群竹类植物的秆高和直径都在第一主轴上具有较大的贡献率。竹秆的生长是通过竹笋节间自下而上逐节伸长形成的。居间分生组织通过细胞分化、伸长加大和老化成熟形成节间生长(周芳纯, 1993)。秆高是竹类植物重要的秆形指标之一。通过对毛竹(Phyllostachys edulis)、厚竹(P. edulis ‘Pachyloen’)等的研究发现, 秆高与竹类植物的直径有直接关系, 直径越大, 竹秆越高(周芳纯, 1981)。故竹秆在垂直方向获得了更多的能量。丛生竹的叶长在第一主轴有较大的因子载荷值。植物会通过增大叶面积、改变叶倾角来捕获更多的光能, 或者通过减小叶面积、改变叶形指数来降低蒸腾损失, 减少光器官损伤(黎莉等, 2010)。丛生竹的叶片可以在水平方向上截获更多的光能, 从而对功能性状的变异产生影响。春季出笋的竹类植物的节间长亦在第一主轴的因子载荷值较大。因为春季出笋的竹子在入冬之前已长成成竹, 新竹已老化, 有发育完整的地上部分。竹类植物的生长对水热条件有较高的要求。水分条件是影响竹类植物生长和分布的重要限制因子, 温度会对竹类植物的种子萌芽、笋芽分化、地下茎生长和光合作用产生极大的影响(顾大形等, 2010)。春季气温回暖, 与自然分布区的雨季同步, 适宜竹类植物的居间分生组织伸长, 节间生长进而发笋长竹, 最终长成秆高节疏的优良竹种。所以竹秆在垂直方向捕获光照的能力对春季出笋的竹类植物功能性状的变异影响较大。
所有物种的PCA分析表明: 竹类植物的直径、秆高和竹壁厚在第一主轴因子载荷值较大。说明在竹类植物功能性状的变异中, 大小性状尤其是直径起主要作用。树木的枝干(包括树高、胸径等)对自身的生长发育起到重要作用。树干的生长是通过固碳来加强形成层的分化进而改变形态结构, 具体表现为枝干高度增加、胸径增大、树皮变厚等(Meinzer et al, 2011)。竹类植物亦是如此。竹笋出土以后依靠节间伸长来完成长高, 秆高和竹壁厚度会随竹秆直径的增大而增加(周益权等, 2010; 张雷等, 2017)。因此竹类植物可在垂直方向获得较多的能量。叶长和叶宽在第二主轴的贡献率较大。竹类植物的生长主要取决于获取的光照(郭雯等, 2018)。叶片是吸收太阳能的主要光合器官, 竹叶与竹林同化光能的数量息息相关(郑维鹏等, 2002)。叶片性状可反映植物对外部环境的高度适应能力和在复杂生境下的自我调控能力(王常顺和汪诗平, 2015)。
综合来看, 不同类型的竹类植物在功能性状上存在显著差异, 但各个功能性状之间存在紧密联系。竹类植物垂直方向上的大小性状(包括直径、秆高和竹壁厚等)在第一主轴的贡献率最大, 主要反映竹类植物在垂直方向上对于光资源的竞争和捕获。随着秆性状值的增加, 竹类植物可以在垂直方向上截获更多的光能, 从而对大小相关的功能性状变异产生影响; 而竹类植物水平方向上的叶片功能性状包括叶长和叶宽在第二主轴具有较大的贡献率, 主要反映了竹类植物在水平方向上对光资源的捕获, 竹类植物可通过调节叶片长度来适应环境变化引起的光资源变化, 在水平方向上获取更多的光能, 以增强资源的获取, 进而影响功能性状的变异。因此竹类植物在水平和垂直方向上对光资源的捕获差异是驱动竹类植物系统演化、特有性分布和出笋季节差异的关键因素。
致谢:
感谢香港嘉道理农场暨植物园标本馆馆长张金龙博士在数据整理和分析方面提供的支持和帮助。
参考文献
Area and endemism
DOI:10.1086/418743 URL [本文引用: 1]
Interspecific and intraspecific variation in specific root length drives aboveground biodiversity effects in young experimental forest stands
DOI:10.1093/jpe/rtw096 URL [本文引用: 1]
A study on cultivation and integrated utilization of large-size cluster bamboo
大型丛生竹的培育技术及其综合利用研究
A study on development and utilization of bamboo shoots forests in four seasons
四季笋用竹林开发利用研究
Plant functional traits and environmental filters at a regional scale
DOI:10.2307/3237229 URL [本文引用: 1]
Plant functional traits, ecosystem structure and land-use history along a climatic gradient in central-western Argentina
On the genera of seed plants endemic to China in Yunnan
云南地区中国种子植物特有属的研究
Morphological plasticity of Indocalamus decorus Q.H.Dai in the heterogeneous environment
异质环境条件下美丽箬竹的形态可塑性
The selection of paper-pulp bamboo species and the development of bamboo paper sector in Sichuan
四川省优良纸浆竹种选择与竹纸产业化发展
Plant functional markers capture ecosystem properties during secondary succession
DOI:10.1890/03-0799 URL [本文引用: 1]
Review of the ecologicol adaptability of bamboo
竹子生态适应性研究综述
Research progress of photosynthetic characteristics and leaf functional traits of bamboo plants
竹类植物光合特性与叶片功能性状研究
Features and distribution patterns of Chinese endemic seed plant species
DOI:10.1111/j.1759-6831.2011.00119.x URL [本文引用: 1]
Organic carbon allocation pattern and changes regulation in various organs of Phyllostachys violascens clone system in shooting period
雷竹克隆系统出笋期有机碳分布变化规律
Relationship between photosynthetic characteristics and leaf functional traits of 12 plant species of marshes in Sanjiang Plain
三江平原12种湿地植物光合特性与叶功能性状的关系研究
Values of bamboo biodiversity and its protection in China
中国竹类植物生物多样性的价值及保护进展
Plant functional traits—Concepts, applications and future directions
植物功能性状研究进展
Spatial-temporal patterns of Bashania fargesii bamboo shoot emergence and giant panda herbivory
巴山木竹发笋和大熊猫取食的时空格局及相关性分析
The present situation and development trend of research on bamboo sprout
笋用竹研究现状及发展趋势
Disturbance response in vegetation: Towards a global perspective on functional traits
DOI:10.2307/3237077 URL [本文引用: 1]
Size- and Age-Related Changes in Tree Structure and Function
The scaling of leaf area and mass: The cost of light interception increases with leaf size
DOI:10.1098/rspb.2007.0417 URL [本文引用: 2]
Divergent drivers of leaf trait variation within species, among species, and among functional groups.
DOI:10.1073/pnas.1803989115 URL [本文引用: 1]
Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide
DOI:10.1071/BT12225_CO URL [本文引用: 1]
Importance of clonal plants in community
群落中克隆植物的重要性
Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests
亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异
Let the concept of trait be functional!
DOI:10.1111/oik.2007.116.issue-5 URL [本文引用: 1]
A review of research on responses of leaf traits to climate change
植物叶片性状对气候变化的响应研究进展
Current research on bamboo culm form
竹子秆形生长和变异的研究进展
Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests
DOI:10.1093/aob/mcl066 URL [本文引用: 1]
The areal-types of Chinese genera of seed plants
世界种子植物科的分布区类型系统
Flora of China: Poaceae Bambusoideae
Studies on bamboo diversity and its evaluation in Yunnan, China
云南竹类多样性及其评价研究现状
Plant endemism and biodiversity
植物的特有现象与生物多样性
Form and structure of bamboo culm of Phyllostachys edulis ‘Pachyloen’ growing in different habitats
不同产地厚竹秆形结构比较
The relationship between leaf area index and growth factor of Dendrocamopsis oldhami
绿竹叶面积指数与生长因子的关系
Research progress in growth rhythm of bamboos
竹类植物生长规律研究进展
Studies on the structure of culm form of Phyllostachys pubescens
毛竹秆形结构的研究
Bamboo forest cultivation
竹林培育学
Cultivation and Utilization on Bamboos
竹林培育和利用

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}