


探索生物多样性的分布格局及其影响机制一直是宏观生态学和生物地理学的基本问题和研究热点(Gaston, 2000; Green & Bohannan, 2006)。基于栅格法(张荣祖, 2011; 王健铭等, 2019)和行政区划法(冯建孟和朱有勇, 2010; 张宇和冯刚, 2018)进行不同尺度生物空间分布格局分析是最简单有效的方法。生物多样性的空间分布格局是各种生态因子梯度变化的综合反映, 已有许多学者提出水热动态假说(O’Brien, 2006)、环境能量假说(Turner, 2004)、寒冷忍耐假说(Hawkins, 2001)、种库假说(Zobel, 1997)以及生境异质性假说(Cox et al, 2016)等来解释物种多样性空间分布格局的形成机制, 亦有研究通过分析物种多样性的地理分异格局及其与区系分化强度之间的关系(冯建孟和朱有勇, 2010), 来解释物种多样性的地理分布格局及其成因。
关于昆虫地理分布格局及其形成机制的研究, 随着分类研究的深入正逐步开展, 涉及蝗虫(许升全等, 2004; 白义等, 2006)、蚜虫(乔格侠等, 2003; 郭昆和乔格侠, 2005)、飞虱(李红荣和陈祥盛, 2009)、瓢蜡蝉(张争光等, 2018)、蚂蚁(沈梦伟等, 2016)等类群, 并应用分子生物学数据探讨生物地理学分布(刘殿锋等, 2009; Dong et al, 2018)。基于昆虫物种的分布信息, 探讨昆虫的多样性及区系分化格局, 结合地质、地貌、气候及生态环境等特征进行空间分异研究, 有助于理解昆虫区系起源及其进化、并为生物多样性保护提供依据。研究表明, 在较小尺度下, 单一类群昆虫的分布格局主要受某类因子影响, 例如在山地森林中蚂蚁的丰富度主要受能量制约(Sanders et al, 2007); 在较大空间尺度, 降水量、年均温、最冷月温度、生境异质性等均可显著影响蚂蚁丰富度格局(Gotelli & Arnett, 2000; 沈梦伟等, 2016)。影响多类群昆虫分布格局的因子主要有植物多样性、海拔等生境异质性因素(Schuldt et al, 2010; Shah et al, 2015; 张宇和冯刚, 2018)。
贺兰山地处银川平原与阿拉善高原之间, 是北温带草原向荒漠过渡的地带, 是我国北方阿拉善-鄂尔多斯生物多样性中心的核心区域。贺兰山以西是腾格里沙漠, 以东是毛乌素沙漠, 以北是乌兰布和沙漠, 是干旱半干旱地区具有代表性的自然综合体和较完整的自然生态系统, 其孤立于荒漠之中, 是趋于中生生境的“岛屿”, 具有带谱比较完整的山地植被垂直带结构和丰富的生物资源(王小明, 2011; 梁存柱等, 2012)。关于该地区昆虫区系的研究集中于近些年的综合考察, 其中记录甲虫30科211属355种(王新谱和杨贵军, 2010; 白晓拴等, 2013)。甲虫是昆虫纲中最大的类群, 起源于三叠纪, 其生态功能十分重要(Diniz-Filho et al, 2010), 其分类信息及地理分布信息的缺乏, 是限制进行生物地理分布格局研究的主要原因。作者在近些年考察的基础上, 陆续采集并整理鉴定该地区甲虫31科252属469种, 并详细记录了物种分布信息(附录1)。本文依据贺兰山甲虫的分布信息及其相关的环境数据, 以期探究以下问题: (1)研究区甲虫物种丰富度及科属区系分化强度在水平方向上存在怎样的分布格局; (2)研究区甲虫物种丰富度的主导因素是什么, 不同优势科物种多样性的影响因素是否存在差异。
1 研究区域与方法
1.1 研究区概况
贺兰山位于105°41′-106°41′ E, 38°13′-39°30′ N, 山体呈西南-东北走向, 长约270 km, 宽约20-40 km, 山体东坡地势平缓, 西坡相对陡峭。山地主体海拔2,000-3,000 m, 最高峰鄂博疙瘩海拔3,556 m。该区域属中温带干旱气候区, 是典型的大陆性气候, 且山地气候明显。年均气温为-0.8℃, 年降水量为200-400 mm。受气候、土壤、地势等因素影响, 贺兰山保护区植被具有明显的垂直分异特征(王小明, 2011; 梁存柱等, 2012)。
1.2 物种分布数据
昆虫的物种分布信息主要通过以下途径获取: (1)作者2007-2018年的实地采集; (2)宁夏农林科学院昆虫馆保存标本; (3) 2007-2011年贺兰山自然保护区综合科学考察中的甲虫标本信息(王新谱和杨贵军, 2010; 白晓拴等, 2013)。共获取标本40,000余份, 隶属于31科252属469种, 其中步甲科(31属97种)、拟步甲科(24属63种)、金龟科(26属49种)、天牛科(31属42种)、象甲科(27属34种)和瓢虫科(16属27种)数量较多。
图1
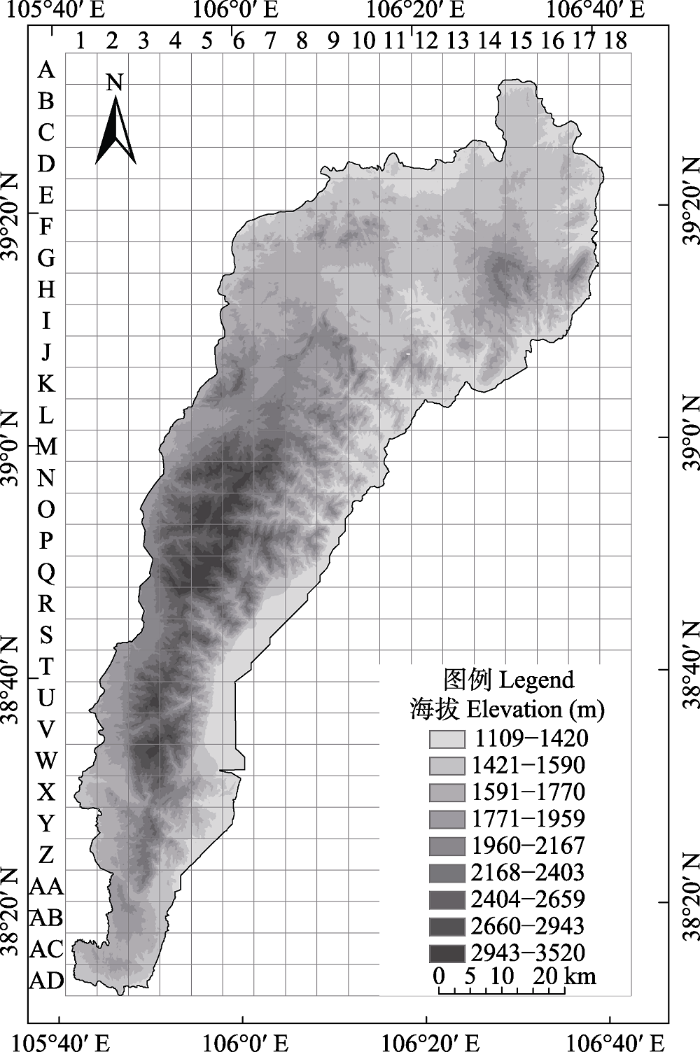
图1
贺兰山地区甲虫分布格局研究的地理单元
Fig. 1
Grids for study on geographic distribution patterns of beetle in the Helan Mountain area
1.3 环境数据
1.3.1 气候数据
选取影响物种丰富度分布格局的7个气候指标, 包括3个水分因子, 即年均降水量(mean annual precipitation, MAP)、年均实际蒸散量(mean annual actual evapotranspiration, AET)、年均潜在蒸散量(mean annual potential evapotranspiration, PET); 4个能量因子, 即年均温(mean annual temperature, MAT)、最冷月均温(mean temperature of coldest month, MTCM)、最热月均温(mean temperature of warmest month, MTWM)和年均太阳辐射(mean annual solar radiation,MASR)。其中, PET和AET数据来源于国际农业研究组织提供的1 km × 1 km分辨率的全球干旱指数和潜在蒸散气候数据库(http://www.cgiarcsi.org), 其余气候变量数据来自世界气候数据网(http://www.worldclim.org) 1 km × 1 km分辨率的气候数据, 以上数据均为1970-2000年的平均值。
1.3.2 生境异质性数据
用单位栅格内的海拔高差(altitude difference, AD)、植被类型数(植被多样性, vegetation diversity, VD)和植被归一化指数(normalized difference vegetation index, NDVI)表征生境异质性。海拔高差, 即各栅格内最高海拔与最低海拔之差, 根据贺兰山数字高程模型(DEM)计算得到。将等面积栅格系统与贺兰山植被1:250000植被图(梁存柱等, 2012)叠加, 统计每个网格内的植被类型数(VD)。植被归一化指数(NDVI)获取自地理国情监测云平台(http://www.dsac.cn) 30 m × 30 m分辨率的数据(2000-2016)。
1.4 物种丰富度、多样性、区系分化强度和相似性分析
(1)以每个栅格内昆虫物种数表征物种丰富度。采用G-F指数(蒋志刚和纪力强, 1999)衡量属(DG)与科(DF)水平上的物种多样性变化, DG是DF的次一级分类阶元多样性, 所以DG ≤ DF; 非单种科越多, 则DG/DF比值越小, G-F指数越高, 计算公式如下:
式中, pi = Ski/Sk, Sk为群落中k科的物种数, Ski为群落中k科i属的物种数。qj= Sj/S, S为群落物种总数, Sj为群落中j属的物种数。
(2)属(Dg)和科(Df)的区系分化强度计算公式如下:
其中, Nsp、Ng和Nf分别为栅格内的物种数、属数和科数(冯建孟和朱有勇, 2010)。
(3)基于栅格单元中的物种分布数据(二元数据, 有分布为1, 无分布为0), 运用Jaccard系数(Cj)计算相似性, 采用非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)对地理单元进行层次聚类分析。Cj = c/(a + b - c), 式中Cj是两个研究地理单元间的相似性系数, a和b分别是两单元的物种数, c是两个单元的共有物种数(Jaccard, 1912)。
1.5 数据分析
运用Origin 9.0软件, 采用一元非线性回归方法的高斯函数模型, 分析甲虫物种丰富度和区系分化强度在南北、东西方向上的变化, 以阐明物种丰富度的空间分布格局。用SPSS 23.0软件对区域内物种丰富度与气候因素、生境异质性进行一元线性回归分析, 确定物种丰富度与不同环境因素的关系。
利用物种丰富度与环境变量(水分因子、能量因子及生境异质性)做冗余分析(RDA), 并通过方差分解来探讨水分、能量和生境异质性3组变量对物种丰富度的独立解释率以及共同解释率。冗余分析(RDA)和方差分解用Canoco 5.12软件, 用前向选择(forward selection)分析各种环境因子对甲虫分布及多样性的贡献率和显著性, 分析前对地表甲虫类群个体数量及环境因子数据进行lg(x + 1)转换。
2 结果
2.1 甲虫物种丰富度及科属区系分化强度的空间分布格局
图2
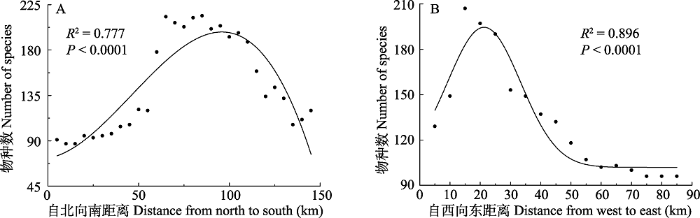
图2
贺兰山甲虫物种丰富度分布格局
Fig. 2
The distribution patterns of beetle species richness in Helan Mountain
图3
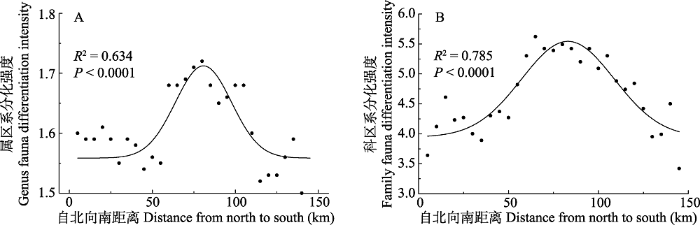
图3
贺兰山甲虫属、科的区系分化在南北方向变化
Fig. 3
The shift of fauna differentiation of the genus and the family of beetles from north to south in Helan Mountain
图4
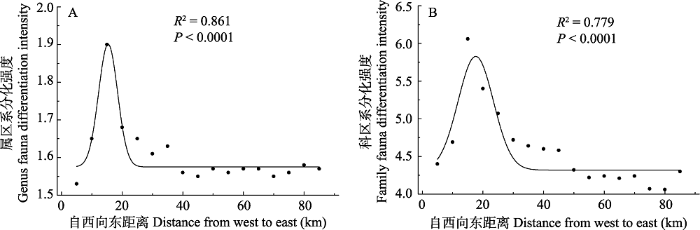
图4
贺兰山甲虫属、科的区系分化在东西方向变化
Fig. 4
The shift of fauna differentiation of the genus and the family of beetles from west to east in Helan Mountain
总体上, 贺兰山东麓甲虫区系分化强度低于西坡, 东坡106.05°E以东范围, 属和科的分化强度基本呈水平变化。因此, 区系分化强度较大区域主要集中在地形复杂、物种多样性丰富的中段区域, 而在北段与南段, 甲虫属和科的区系分化能力则相对较弱。
2.2 甲虫分布格局的相似性聚类分析
基于183个地理单元中469种昆虫的二元分布数据, 采用Jaccard相似性系数计算相似性矩阵, 运用非加权组平均法(UPGMA), 对地理单元进行聚类分析。结果表明, 在Jaccard相似性系数为0.36时, 贺兰山183个地理单元的甲虫聚为3个地理群(图5), 即北段强旱生景观甲虫地理群(I), 界线大致为汝箕沟(东坡)-油门沟(西坡)一线以北; 中西段半湿生景观甲虫地理群(II), 界线大致为汝箕沟(东坡)-油门沟(西坡)一线以南, 榆树沟(西坡-东坡)以北海拔1,800 m以上; 中东段及南段半旱生景观甲虫地理群(III), 界线大致为中段东部汝箕沟(东坡)和南段榆树沟以南海拔1,200-1,800 m范围的半旱生景观甲虫地理群。
图5
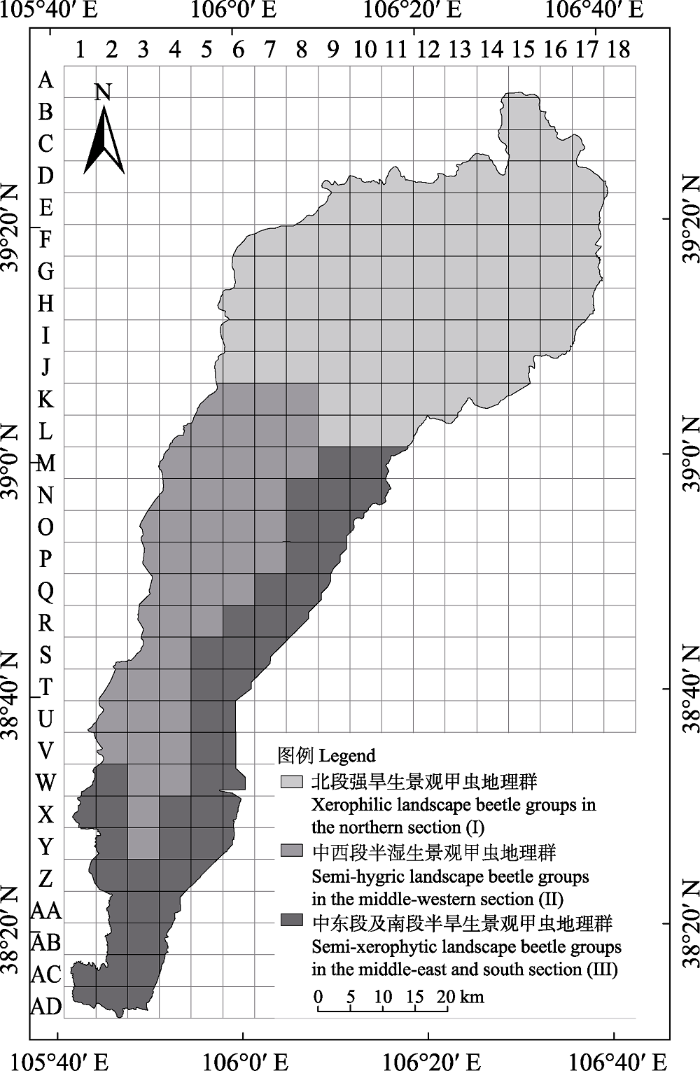
不同地理单元群甲虫多样性比较结果见表1, 地理群III的科、属数最多, 地理群II的物种丰富度最高, 从DG、DF指数看, 中东段及南段半旱生景观甲虫地理群(III)均最高, 其次是中西段半湿生景观甲虫地理群(II), 说明贺兰山中段和南段种类比较丰富。G-F指数方面, 3个地理群都非常接近, 可能是因为同属于一个山体, 地理位置相近, 总体上地理群II和III大于地理群I, 说明贺兰山中段和南段甲虫非单种科最多, 而北段区域最少。属和科的区系分化强度均以地理群II为最高。
表1 贺兰山不同地理单元甲虫群落多样性比较
Table 1
| 科数 No. of family | 属数 No. of genus | 种数 No. of species | G指数 (DG) | F指数 (DF) | G-F指数 (DG-F) | 属区系分化强度 Genus fauna differentiation intensity (Dg) | 科区系分化强度 Family fauna differentiation intensity (Df) | |
|---|---|---|---|---|---|---|---|---|
| 北段强旱生景观甲虫地理群 Xerophilic landscape beetle groups in the northern section (I) | 26 | 145 | 250 | 4.71 | 26.80 | 0.824 | 1.57 | 3.19 |
| 中西段半湿生景观甲虫地理群 Semi-hygric landscape beetle groups in the middle-western section (II) | 29 | 218 | 419 | 5.03 | 35.35 | 0.858 | 1.73 | 4.29 |
| 中东段及南段半旱生景观甲虫地理群Semi-xerophytic landscape beetle groups in the middle-east and south section (III) | 30 | 222 | 378 | 5.14 | 37.49 | 0.863 | 1.53 | 3.79 |
2.3 不同环境因子与甲虫物种丰富度及区系分化强度的关系
线性回归分析结果显示(表2), 贺兰山甲虫物种丰富度和区系分化强度与全部水分因子呈极显著的正相关(P < 0.001), 而与全部能量因子呈极显著的负相关(P < 0.001)。生境异质性可明显提高甲虫物种丰富度及科、属区系分化强度(P < 0.001)。
表2 贺兰山甲虫物种丰富度和区系分化强度与环境因子的一元回归相关系数
Table 2
| 环境因子 Environmental factor | 物种丰富度 Species richness (Nsp) | 属区系分化强度 Genus fauna differentiation intensity (Dg) | 科区系分化强度 Family fauna differentiation intensity (Df) |
|---|---|---|---|
| 海拔高差 Altitude difference (AD) | 0.54* | 0.27* | 0.46* |
| 植被类型数 Vegetation diversity (VD) | 0.55* | 0.49* | 0.27* |
| 植被归一化指数 Normalized difference vegetation index (NDVI) | 0.56* | 0.41* | 0.60* |
| 年均降水量 Mean annual precipitation (MAP) | 0.61* | 0.34* | 0.62* |
| 年均潜在蒸散量 Mean annual potential evapotranspiration (PET) | 0.31* | 0.50* | 0.25* |
| 年均实际蒸散量 Mean annual actual evapotranspiration (AET) | 0.49* | 0.39* | 0.36* |
| 年均温 Mean annual temperature (MAT) | -0.34* | -0.58* | -0.44* |
| 最冷月均温 Mean temperature of coldest month (MTCM) | -0.26* | -0.51* | -0.19* |
| 最热月均温 Mean temperature of warmest month (MTWM) | -0.41* | -0.58* | -0.34* |
| 年均太阳辐射 Mean annual solar radiation (MASR) | -0.56* | -0.32* | -0.57* |
*P < 0.001
2.4 水分、热量与生境异质性对甲虫物种丰富度的相对影响
运用Canoco软件对甲虫物种丰富度与环境因子进行冗余分析(RDA), 前项选择结果表明(表3), 年均温(34.8%, P < 0.01)、年均降水量(9.9%, P < 0.01)、海拔高差(3.0%, P < 0.01)分别是能量因子、水分因子和生境异质性因子中独立解释率最高的因子。
表3 环境因子对甲虫物种丰富度分布的相对贡献
Table 3
| 环境因子 Environmental factor | 解释率 Percentage of variance explained (%) | 贡献率 Contribution (%) | F | P |
|---|---|---|---|---|
| 年均温 Mean annual temperature (MAT) | 34.8 | 57.0 | 96.6 | 0.002 |
| 年均降水量 Mean annual precipitation (MAP) | 9.9 | 16.2 | 32.1 | 0.002 |
| 最冷月均温 Mean temperature of coldest month (MTCM) | 3.8 | 6.1 | 13 | 0.002 |
| 年均潜在蒸散量 Mean annual potential evapotranspiration (PET) | 3.6 | 5.9 | 13.3 | 0.002 |
| 海拔高差 Altitude difference (AD) | 3.0 | 4.9 | 11.7 | 0.002 |
| 最热月均温 Mean temperature of warmest month (MTWM) | 2.8 | 4.5 | 11.6 | 0.002 |
| 年均实际蒸散量 Mean annual actual evapotranspiration (AET) | 1.5 | 2.5 | 6.5 | 0.002 |
| 年均太阳辐射 Mean annual solar radiation (MASR) | 1.0 | 1.6 | 4.2 | 0.002 |
| 植被归一化指数 Normalized difference vegetation index (NDVI) | 0.6 | 0.9 | 2.5 | 0.002 |
| 植被类型数 Vegetation diversity (VD) | 0.3 | 0.5 | 1.4 | 0.048 |
冗余分析和方差分解分析的结果显示(图6), 水分、能量和生境异质性因子共同解释了全部物种丰富度58.9%的变异, 其中水分和能量共同解释率为57.1%, 单独解释率分别为5.9%和7.1%。生境异质性可以解释物种丰富度35.2%的变异, 单独解释率仅为1.8%。水分、能量和生境异质性因子三者公共解释率为25.0%。
图6
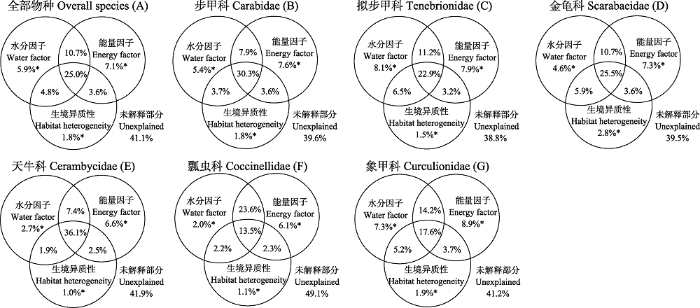
图6
水分、能量与生境异质性因子对甲虫全部物种和优势科丰富度分布的影响(A-G)。*P < 0.001。
Fig. 6
Relative influence of water, energy and habitat heterogeneity on distribution of beetle species richness of overall and dominant family. *P < 0.001.
水分、能量与生境异质性对不同优势科类群丰富度解释率不同。水分因子对拟步甲科丰富度具有最大解释率(48.7%), 能量因子对天牛科丰富度有着最大解释率(52.6%), 生境异质性对所有科的独立解释率均低于水分、能量因子的影响, 其中对天牛科的解释率最大(41.5%)。水分、能量和生境异质性三类因子中任意两个因子的共同影响均表现为对天牛科丰富度具有最大解释率。
图7

图7
水分、能量与生境异质性因子对不同地理单元甲虫丰富度分布格局的解释。*P < 0.001。
Fig. 7
Relative influence of water, energy and habitat heterogeneity on distribution of beetle species richness in different geographical units. *P < 0.001.
3 讨论
3.1 贺兰山甲虫物种丰富度及区系分化强度格局
贺兰山是我国北方阿拉善-鄂尔多斯生物多样性中心的核心区域, 是北温带草原向荒漠过渡的地带, 该区域生态环境脆弱, 但孕育了较为丰富的生物多样性, 植物群落物种丰富度存在着显著的空间异质性, 即中段以森林和中生灌丛为主, 南、北段荒漠化程度较高, 东坡比西坡植被覆盖度低(王炜等, 2004; 梁存柱等, 2012)。本研究结果表明, 贺兰山已知的31科252属469种甲虫也存在着明显的地理分布差异, 物种丰富度以贺兰山中段(自北段插旗口沟至南段小口子沟)较高, 西坡较东麓高, 说明贺兰山中段地区甲虫物种多样性最为丰富, 值得重点保护和关注, 而其他地区则相对贫乏。区系分化强度较大区域主要集中在物种多样性丰富的中段区域及西坡, 而在北段与南段, 甲虫属和科的区系分化能力则相对较弱, 说明贺兰山地区甲虫物种丰富度与区系分化强度之间存在密切的联系。甲虫物种丰富度和区系分化与植被空间分异有较好的对应关系, 说明植物多样性增加有助于甲虫多样性的提高, 为甲虫提供了更丰富的生态空间及食物资源(Zhang et al, 2016)。
属的分化通常被认为是近现代历史地质和环境的表现, 科的分化被认为表征比较久远的地质历史(李果等, 2009; 梁存柱等, 2012)。目前关于贺兰山昆虫的起源未见报道, 但植物与昆虫是协同进化的, 植物的区系变化、地质历史、气候变迁特征等均在一定程度上反映本地甲虫区系的起源历史非常悠久(王新谱和杨贵军, 2010)。研究表明贺兰山是滨太平洋和特提斯-喜马拉雅构造域联合作用的结果, 贺兰山经历了3次较大隆升, 致使大陆性气候增强, 山地垂直植被带于第三纪末形成, 中段抬升最高, 在南北方向和东西方向上具有不同的水热条件组合以及植被组成(赵红格等, 2007; 王小明, 2011)。生境异质性的增强为甲虫分布创造了条件, 地形相对复杂的中段地区甲虫区系强烈分化, 其物种丰富度显著增加, 南、北段则较低, 尤其是北段整体维持在较低水平。基于甲虫分布栅格的聚类分析表明, 中段的地理群II的物种丰富度、属和科的区系分化强度最大(表1), 推测是由于该区域海拔高差最大, 由此引起的生境异质性最大, 昆虫物种分化明显。因此, 该区域是甲虫物种多样性保护的核心区域。地理群III具有较高的科、属多样性, G-F指数较大, 可能是由于该区域有银川平原及湿地昆虫种类的渗透。
3.2 气候和生境异质性对甲虫物种丰富度分布格局的相对影响
大尺度上的水热条件被认为是地带性植被与生物物种多样性地理分布格局形成的决定性因素。贺兰山地处银川平原和阿拉善高原之间, 位于中温带干旱气候区, 是典型的大陆性气候, 山体陡峭、地形复杂, 山地气候明显。贺兰山三面环绕的沙漠景观对该山体水热空间分布有着强烈影响。因此, 较小尺度下, 山地生物多样性空间分布格局受自身地形地貌及周围景观引起的水热条件的影响。本研究表明, 甲虫物种丰富度与水分因子呈显著正相关, 与能量因子显著负相关, 说明能量对于地处干旱区的贺兰山甲虫多样性更多是抑制因素, 这可能是贺兰山地区日照强, 蒸发量大, 降低了昆虫直接或间接对水资源的利用(王健铭等, 2019)。方差分解结果显示, 水分和能量对甲虫全部物种丰富度变异的总解释率相当, 共同解释了贺兰山全部甲虫物种丰富度57.1%的变异, 且它们均存在显著的单独解释率(图6), 说明它们共同决定了贺兰山地区甲虫物种丰富度地理格局。冗余分析(RDA)方差分解结果显示(图7), 水分因子对地理群I和III甲虫物种丰富度影响大于能量因子, 地理群II中能量因子影响大于水分因子。很多研究证明生境异质性可以为更多物种提供更多样化的资源与生态位(Chen et al, 2011)。本研究显示, 贺兰山甲虫物种丰富度均与生境异质性存在显著的正相关, 这说明高生境异质性可以为甲虫物种共存与多样性维持提供更多的机会。但是, 生境异质性对全部物种单独解释率明显低于水分和能量因子。在地理群III中, 生境异质性对全部物种的解释率明显提高, 和水分因子成为贺兰山北段、南段甲虫物种丰富度的限制性因子。
水分、能量与生境异质性对不同优势科物种丰富度影响并不一致, 这可能是由于进化历史、食性、扩散能力与生活史策略的差异造成的(王志恒等, 2009)。本研究表明(图6), 水分因子对拟步甲科丰富度具有最大解释率, 该科昆虫在贺兰山的分布以海拔2,000 m以下的干旱生境为主, 说明水分因子是干旱地区拟步甲昆虫物种多样性维持最为关键的限制因子。步甲科广泛分布于各种生境, 且在中高海拔趋于向中生湿润的环境聚集, 能量因子的作用大于水分因子。天牛科和瓢虫科等其他科昆虫趋向分布于植被郁闭度较高的中生环境, 物种丰富度受能量因子的作用略大。生境异质性对所有科的独立解释率均低于水分、能量因子的单独影响, 在各自的模型中, 占比最高的是天牛科, 最低的是瓢虫科, 可能是由于瓢虫科几乎全部是肉食性, 生境异质性对该科的影响比对植食性的天牛科昆虫要小。
综上可见, 水分和能量因子是贺兰山地区甲虫物种丰富度空间分布格局的主导因子, 生境异质性有助于提高甲虫物种丰富度, 而在贺兰山的南、北段, 生境异质性和水分因子作用明显。在本研究中, 水分、能量和生境异质性并没有完全解释对贺兰山甲虫全部物种和优势科种类丰富度格局的所有变异。山地是具有一定的海拔高度、相对高度和坡度的生态复合系统, 地形通过地貌过程控制了光、热、水和土壤养分等资源因子的空间再分配, 对植被产生直接作用(Cantón et al, 2004)。已有研究表明, 地形因子对地表甲虫分布存在影响(杨贵军等, 2016; 杨益春等, 2017), 土壤理化性质对地表甲虫多样性也存在重要作用(Thomas & Marshall, 1999; 郭海滨等, 2009; 娄巧哲等, 2011; 李岳诚等, 2014)。地表甲虫在甲虫中占较大比例, 因此, 今后的研究应进一步引入土壤与地形等因素, 将有助于更全面探究该区域甲虫物种丰富度分布格局的潜在影响机制。
附录 Supplementary Material
附录1 贺兰山甲虫物种名录
Appendix 1 List of beetle species in Helan Mountain
参考文献
Cluster analysis on the distribution patterns of grasshopper in Shaanxi
陕西蝗虫地理分布格局的聚类分析
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain
Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China
Biogeography: An Ecological and Evolutionary Approach
Defying the curse of ignorance: Perspectives in insect macroecology and conservation biogeography
Biogeographic patterns and diversification dynamics of the genus Cardiodactylus Saussure (Orthoptera, Grylloidea, Eneopterinae) in Southeast Asia
Geographical patterns of diversity of Gymnosperms in northwest Yunnan and their correlation with flora differentiation
滇西北地区裸子植物多样性的地理分布格局及其与区系分化之间的关系
Biogeographic effects of red fire ant invasion
Spatial scaling of microbial biodiversity
Insect community structure in relation to environmental factors on bermudagrass lawn
狗牙根草坪昆虫群落组成与环境因子相关性研究
Study on geographical distributional pattern of the subfamily Hormaphidinae (Homoptera, Ormaphididae)
扁蚜亚科(同翅目, 扁蚜科)蚜虫地理分布格局初探
The distribution of the flora in the alpine zone
Fauna and Distribution of Tenebrionid Beetles in Alxa Plateau (Coleoptera: Tenebrionoidea)
阿拉善高原拟步甲区系与地理分布(鞘翅目: 拟步甲总科)
Avian-mammalian species diversity in nine representative sites in China
鸟兽物种多样性测度的G-F指数方法
The spatial pattern of species richness and diversity centers of gymnosperm in China
中国裸子植物物种丰富度空间格局与多样性中心
The fauna and biogeography of planhoppers (Hemiptera, Fulgoroidea, Delphacidae) in Guizhou
贵州飞虱科昆虫物种多样性及地理分布格局(半翅目, 蜡蝉总科, 飞虱科)
Species diversity of ground- dwelling beetles and its relationship with environmental factors in the artificial Caragana brushland of fixed sandy lands in Ningxia
荒漠景观固沙柠条林地地表甲虫多样性及其与环境因子的关系
Molecular biogeography of some species of the Acrididae in China
中国蝗科部分昆虫的分子生物地理学分析
Diversity of ground- dwelling beetles within the southern Gurbantunggut Desert and its relationship with environmental factors
古尔班通古特沙漠南缘地表甲虫物种多样性及其与环境的关系
Biological relativity to water-energy dynamics
Species diversity and geographical distribution patterns of Drepanosiphidae in China
中国斑蚜科物种多样性及地理分布格局
Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains
Tree diversity promotes insect herbivory in subtropical forests of south-east China
Latitudinal patterns and large-scale environmental determinants of stream insect richness across Europe
Relationships between geographic patterns of ant species richness and environmental factors in China
中国蚂蚁丰富度地理分布格局及其与环境因子的关系
Arthropod abundance and diversity in differently vegetated margins of arable fields. Agriculture,
Explaining the global biodiversity gradient: Energy, area, history and natural selection
Biogeographic patterns and environmental interpretation of plant regional species richness in Alxa Plateau of northern China
阿拉善高原植物区域物种丰富度格局及其环境解释
The diversity and spatial distribution of plant communities in the Helan Mountains
贺兰山植物群落类型多样性及其空间分异
The species-energy hypothesis as a mechanism for species richness patterns
物种多样性地理格局的能量假说
Cluster analysis on the distribution patterns of grasshopper in Ningxia
宁夏蝗虫地理分布格局的聚类分析
Distribution of darkling beetles and its relationships with topography in Helan Mountain, Ningxia
宁夏贺兰山拟步甲科昆虫分布与地形的关系
Effects of topographic factors on the distribution pattern of carabid species diversity in the Helan Mountains, northwestern China
地形对贺兰山步甲群落物种多样性分布格局的影响
Plant diversity accurately predicts insect diversity in two tropical landscapes
Distribution pattern and mechanism of insect species diversity in Inner Mongolia
内蒙古昆虫物种多样性分布格局及其机制
Geographical distribution pattern of Issidae (Hemiptera: Fulgromorpha) from China
中国瓢蜡蝉科(半翅目: 蜡蝉总科)昆虫的地理分布格局
Uplift time and its evolution of Helan Mountain
贺兰山隆升时限及其演化
The relative role of species pools in determining plant species richness: An alternative explanation of species coexistence?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}