Designing leaf marginal shapes: Regulatory mechanisms of leaf serration or dissection
Jinxiu Ke, Duo Chen, Yanping Guo*,
Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, and College of Life Sciences, Beijing Normal University, Beijing 100875
The mechanism of formation and evolution of phenotypic diversity is one of the key problems in biodiversity science because phenotype diversity is not only a marker of species diversity, but carries the designs adjusted to environments. Plant leaves exhibit a great deal of morphological variation. Such variation is attributed largely to changes of leaf marginal architecture. Leaf marginal shapes can be described as entire, serrate, lobed (varying in depth and patterns) and dissected (also referred to as compound leaf). The molecular mechanism controlling the development of leaf marginal shape has been intensively studied in Arabidopsis thaliana, Cardamine hirsuta, Solanum lycopersicum, and some other plants. Many important regulatory factors such as transcription factors, small RNAs and plant hormones have been found involved in the development of leaf serration or dissection. Among those factors, the transcription factor NAM/CUC, miR164 and auxin in the auxin efflux module play a central role through a feedback loop, and this regulatory module appears to be conserved across the eudicots; the transcription factors TCPs, SPLs and some other miRNAs also take part in the auxin efflux pathway. Transcription factors of the KNOX family play roles in the development of leaf lobes as well although most of the researches about KNOX genes have focused on their regulation of the morphogenesis of compound leaves. In addition, studies in Arabidopsis, Cardamine and other taxa of the Brassicaceae have shown that the gene RCO promotes the development of leaf dissection by repressing growth at the flanks of protrusions generated by CUC-auxin patterning. The present paper reviews the recent progress and integrate the major results of researches about the molecular mechanisms that underlie leaf serration or dissection. We hope this may provide reference for unraveling the morphogenetic origin of the spectacular diversity of leaf marginal shapes.
Keywords:leaf serration/dissection
;
morphogenesis
;
evolutionary development
;
regulatory network
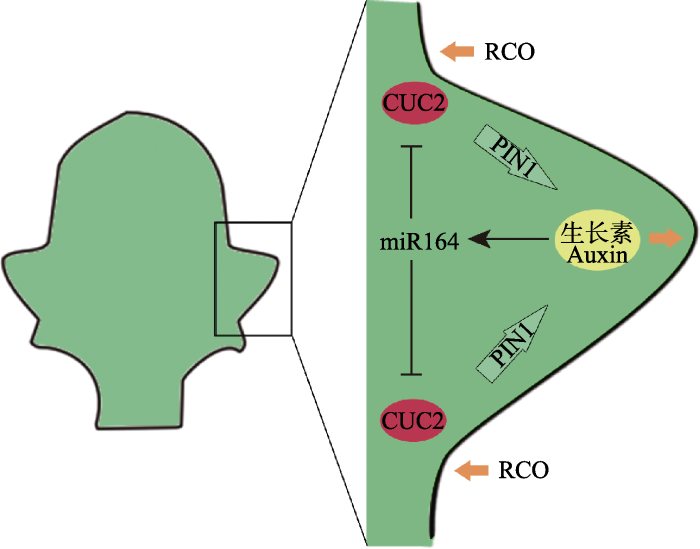
Fig. 1 A model for the molecular regulation of the development of leaf marginal serration according to studies on Arabidopsis thaliana and Cardamine hirsute (Barkoulas et al, 2007; Runions & Tsiantis, 2017). At the heart of the model is a feedback loop between CUC2 and auxin activities. CUC2 is required for PIN1-mediated auxin polar transport (horrow arrows); in turn, auxin activity maxima at the tip of the developing serration activates miR164 which represses CUC2 posttranscriptionally and generates an interspersed pattern of auxin maxima and CUC2 expression at the leaf margin. MIR164 and CUC2 are expressed in partially overlapping regions at the sinus of the serrations. Auxin enhances outgrowth of the serrations. There are additional growth regulators modulating leaf growth to shape the form of protrusions, for instance, RCO inhibits growth in indentations, producing more dissected forms.
1.2 真双子叶植物中NAM/CUC类基因对叶缘形态调控作用的保守性
Blein等(2008)比较研究了5个真双子叶植物中NAM (CUC1/2)和CUC3在叶边缘形态建成中的作用, 发现该类基因具有调控单叶叶齿和复叶上小叶形成的双重作用, 且这个作用在真双子叶植物中是保守的。研究表明, 降低番茄植株叶片中NAM/CUC基因的表达后, 小叶几乎融合(Berger et al, 2009), 数目变少, 小叶边缘变为全缘(Blein et al, 2008); 碎米荠中减少NAM/CUC基因的表达后, 小叶融合, 小叶和托叶的数目减少(Blein et al, 2008); 在豌豆的NAM/CUC基因缺失突变体中检测到这一基因的直接下游调控基因的表达量明显下降, 同时叶形态发生异常(Blein et al, 2008); 耧斗菜(Aquilegia caerulea)的NAM基因或CUC3基因被沉默后, 小叶叶裂减少, 而同时沉默NAM基因和CUC3基因后, 小叶变为全缘(Blein et al, 2008)。此后在苜蓿中的研究也发现, MtNAM基因缺失突变体的叶片也发生小叶融合或边缘锯齿减少的现象(Cheng et al, 2012)。
植物miRNA负调控内源性靶基因, 通过与靶基因转录物的部分或完全结合来识别和裂解这些转录物。在叶发育过程中, miRNAs与其靶向基因共同作用于生长素的输出通路, 从而调控叶缘齿/裂的发育。研究表明, 生长素最大积累瞬时诱导MIR164的表达, miR164通过对靶基因CUC2的转录后负调控来影响器官原基的边界建立。利用T-DNA插入方法获得的拟南芥mir164突变体, MIR164表达量降低导致CUC2表达量升高, 叶片裂刻加深(Nikovics et al, 2006), 而当miR164过表达时, CUC2表达降低, 导致拟南芥叶变全缘、碎米荠小叶变少(Rubio-Somoza et al, 2014)。CUC2的旁系同源基因CUC3缺乏miR164的结合位点而不受其调控, 但CUC3与CUC2之间存在相互作用, CUC2能促进CUC3活性, 两者形成的异源二聚体在叶片发育过程中调节叶齿/裂的发育和叶缘复杂性的建成(Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014)。
另一组能调控叶缘复杂性的转录因子是受miR319负调控的TCPs (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF)家族成员。MIR319A和MIR319B单突变一定程度上抑制拟南芥叶缘齿裂的形成, 双突变使得拟南芥叶变全缘(Koyama et al, 2017)。过表达MIR319, TCP水平降低, 导致拟南芥叶齿增多、碎米荠小叶增多(Palatnik et al, 2003; Daniel et al, 2004)。Rubio-Somoza和他的团队将miR164-CUC与miR319-TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低。其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017)。
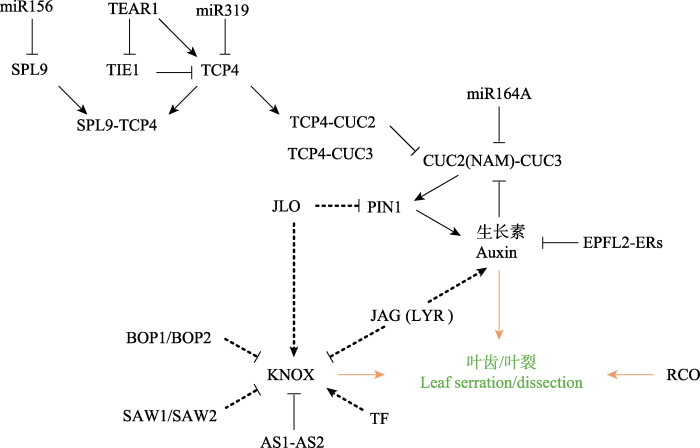
Fig. 2 The molecular regulating network underlining the development leaf serration/dissection. The solid lines indicate the relatively clear interactions between molecules, while the dotted lines suggest unsure interactions reported by a few studies. The orange arrows suggest promotion of leaf serration/dissection.
6 展望
植物叶的发育是一个非常有趣的过程。首先, 叶发育的过程具有很强的可塑性, 不同的环境决定着不同的叶片大小和形态; 而另一方面, 植物叶片的发育又遵循着一个基本模式, 是一个比较保守的有序过程。叶边缘发育是叶形态建成的最后步骤(Tang et al, 2016), 很大程度上决定了植物叶形的复杂性。
Transcriptomic analysis suggests a key role for SQUAMOSA PROMOTER BINDING PROTEIN LIKE, NAC and YUCCA genes in the heteroblastic development of the temperate rainforest tree Gevuina avellana (Proteaceae)
Arabidopsis type II Phosphatidylinositol 4-Kinase PI4Kγ5 regulates auxin biosynthesis and leaf margin development through interacting with membrane-bound transcription factor ANAC078
The TIE1 transcriptional repressor links TCP transcription factors with TOPLESS/ TOPLESS-RELATED corepressors and modulates leaf development in Arabidopsis
... A model for the molecular regulation of the development of leaf marginal serration according to studies on Arabidopsis thaliana and Cardamine hirsute (Barkoulas et al, 2007; Runions & Tsiantis, 2017). At the heart of the model is a feedback loop between CUC2 and auxin activities. CUC2 is required for PIN1-mediated auxin polar transport (horrow arrows); in turn, auxin activity maxima at the tip of the developing serration activates miR164 which represses CUC2 posttranscriptionally and generates an interspersed pattern of auxin maxima and CUC2 expression at the leaf margin. MIR164 and CUC2 are expressed in partially overlapping regions at the sinus of the serrations. Auxin enhances outgrowth of the serrations. There are additional growth regulators modulating leaf growth to shape the form of protrusions, for instance, RCO inhibits growth in indentations, producing more dissected forms. ...
A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta
1
2008
... 围绕叶边缘齿或裂刻的生长素调控途径, 有关生长素输出途径中PIN蛋白及诸多调控因子的研究较为深入.研究表明, 拟南芥的齿裂叶中, 每个叶齿突出处都是生长素的高浓度汇聚点, 这与PIN1蛋白在细胞膜上的极性定位有关.PIN1的动态表达量数据显示, 叶边缘的PIN1蛋白在表皮细胞中形成一个极性汇聚点, 生长素的流动方向与PIN1在细胞中的极性定位方向一致, 从而建立了生长素的最大积累, 促使此处的叶齿形成.在pin1突变体中, 生长素的梯度分布被打乱, 沿着整个叶边缘均匀分布, 叶缘裂刻消失, 形成边缘光滑的全缘叶, 说明依赖于PIN1的生长素浓度梯度分布决定了叶边缘裂刻的形成和分布(Benkova et al, 2003; Hay et al, 2006; Barkoulas et al, 2008; Bilsborough et al, 2011). ...
Local, efflux-dependent auxin gradients as a common module for plant organ formation
... 生物体表型的形成是大量基因表达(包括转录和翻译)的集合产物(Bonn & Furlong, 2008), 这些基因共同构成复杂的调控网络(gene regulatory network, GRN), 上、下游基因以及相关的小RNA联结成通路(Carroll, 2008), 通过表达的时空模式和/或表达水平的改变, 影响着形态特征的发育和建成.关于叶边缘形态建成的发育调控机制, 在拟南芥(Arabidopsis thaliana)、碎米荠(Cardamine hirsuta)、烟草(Nicotiana tabacum)、番茄(Solanum lycopersicum)、苜蓿(Medicago truncatula)、豌豆(Pisum sativum)等模式植物中已有大量的研究报道(Barkoulas et al, 2007; Blein et al, 2008, 2010; Efroni et al, 2010; Chitwood & Sinha, 2014; Bar & Ori, 2015).叶是茎端分生组织(shoot apical meristem, SAM)产生的第一类侧生器官, 其发生和形态建成受茎顶端分生组织生长和分化的影响.KNOX类基因(Class I KNOTTED1-like homeobox)主要在茎端分生组织中表达, 调控细胞的分裂和生长, 决定叶原基的形成和复叶形态的发育, 是迄今为止研究最多的与叶形成和发育密切相关的调控基因(Hay et al, 2006; Hay & Tsiantis, 2010; Bar & Ori, 2015).叶原基起始后叶向两侧延展, 叶缘形态随即建成.叶边缘形态多样性也是在复杂的基因调控网络作用下形成的, 涉及众多的因子, 如特异基因、转录因子、激素、小RNA等.本文综述此方面近20年的研究进展, 探讨叶片边缘形态发育的分子调控机理. ...
... 植物miRNA负调控内源性靶基因, 通过与靶基因转录物的部分或完全结合来识别和裂解这些转录物.在叶发育过程中, miRNAs与其靶向基因共同作用于生长素的输出通路, 从而调控叶缘齿/裂的发育.研究表明, 生长素最大积累瞬时诱导MIR164的表达, miR164通过对靶基因CUC2的转录后负调控来影响器官原基的边界建立.利用T-DNA插入方法获得的拟南芥mir164突变体, MIR164表达量降低导致CUC2表达量升高, 叶片裂刻加深(Nikovics et al, 2006), 而当miR164过表达时, CUC2表达降低, 导致拟南芥叶变全缘、碎米荠小叶变少(Rubio-Somoza et al, 2014).CUC2的旁系同源基因CUC3缺乏miR164的结合位点而不受其调控, 但CUC3与CUC2之间存在相互作用, CUC2能促进CUC3活性, 两者形成的异源二聚体在叶片发育过程中调节叶齿/裂的发育和叶缘复杂性的建成(Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014). ...
... 另一组能调控叶缘复杂性的转录因子是受miR319负调控的TCPs (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF)家族成员.MIR319A和MIR319B单突变一定程度上抑制拟南芥叶缘齿裂的形成, 双突变使得拟南芥叶变全缘(Koyama et al, 2017).过表达MIR319, TCP水平降低, 导致拟南芥叶齿增多、碎米荠小叶增多(Palatnik et al, 2003; Daniel et al, 2004).Rubio-Somoza和他的团队将miR164-CUC与miR319-TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低.其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017). ...
... JAGGED (JAG)编码具有C2H2锌指结构的转录因子.JAG功能缺失能引起拟南芥的叶产生叶裂; 番茄中的JAG同源基因是LYRATE (LYR), lyr突变株产生更多小叶, 而LYR的过表达则引起小叶融合.可见, JAG/LYR可能通过与KNOX以及生长素途径中的因子相互作用, 共同参与叶裂深度和小叶数目的调控(Dinneny et al, 2004; David-Schwartz et al, 2009; Bar & Ori, 2014). ...
Morphogenesis of simple and compound leaves: A critical review
4
2010
... 生物体表型的形成是大量基因表达(包括转录和翻译)的集合产物(Bonn & Furlong, 2008), 这些基因共同构成复杂的调控网络(gene regulatory network, GRN), 上、下游基因以及相关的小RNA联结成通路(Carroll, 2008), 通过表达的时空模式和/或表达水平的改变, 影响着形态特征的发育和建成.关于叶边缘形态建成的发育调控机制, 在拟南芥(Arabidopsis thaliana)、碎米荠(Cardamine hirsuta)、烟草(Nicotiana tabacum)、番茄(Solanum lycopersicum)、苜蓿(Medicago truncatula)、豌豆(Pisum sativum)等模式植物中已有大量的研究报道(Barkoulas et al, 2007; Blein et al, 2008, 2010; Efroni et al, 2010; Chitwood & Sinha, 2014; Bar & Ori, 2015).叶是茎端分生组织(shoot apical meristem, SAM)产生的第一类侧生器官, 其发生和形态建成受茎顶端分生组织生长和分化的影响.KNOX类基因(Class I KNOTTED1-like homeobox)主要在茎端分生组织中表达, 调控细胞的分裂和生长, 决定叶原基的形成和复叶形态的发育, 是迄今为止研究最多的与叶形成和发育密切相关的调控基因(Hay et al, 2006; Hay & Tsiantis, 2010; Bar & Ori, 2015).叶原基起始后叶向两侧延展, 叶缘形态随即建成.叶边缘形态多样性也是在复杂的基因调控网络作用下形成的, 涉及众多的因子, 如特异基因、转录因子、激素、小RNA等.本文综述此方面近20年的研究进展, 探讨叶片边缘形态发育的分子调控机理. ...
... KNOX家族成员在植物顶端分生组织中表达, 其调控作用对叶和花的发生、发育都是必不可少且是保守的(Bharathan et al, 2002; Piazza et al, 2010).KNOX基因在叶原基形成的起始时期表达下调或没有表达, 但随后在叶片发育过程中其表达存在物种特异性, 这与叶缘形态的多样性有关(Piazza et al, 2010).在绝大多数双子叶植物中, KNOX的表达活性与叶缘的复杂性高度相关, 在很多具羽状复叶或叶缘有裂刻的物种中可检测到KNOX在叶片中表达 (Bharathan et al, 2002; Piazza et al, 2010), 如在碎米荠、番茄的复叶里, KNOX基因在叶片发育过程中有表达.拟南芥中存在全缘叶的植株类型, Piazza等(2010)在用15个基因位点的DNA序列信息对拟南芥属及十字花科其他支系的物种进行系统发生分析基础上, 推断拟南芥中的齿裂叶为祖征, 全缘叶为衍生, 而齿裂的消失与叶片中STM (KNOX家族成员)的表达缺失有关, 因为在其他叶缘分裂的物种的发育叶片中均检测到STM的表达, 且敲减其表达后叶裂消失; 进一步的分析显示该表达缺失可能与STM启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... ; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... 通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
Control of Arabidopsis leaf morphogenesis through regulation of the YABBY and KNOX families of transcription factors
2
2010
... BLADE ON PETIOLE1 (BOP1)和BOP2属于BTB蛋白家族, 两者能形成二聚物, 发挥转录激活因子的功能.BOP1和BOP2在侧生器官基部表达, 在叶基部表达的BOP能直接正调控AS2, 抑制该区域KNOX基因的表达(Jun et al, 2010).bop1和bop2双突体的叶片变大, 在叶柄处长出叶齿、叶翅等(Ha et al, 2003, 2010; Norberg et al, 2005).因此, BOP通过抑制叶基部和叶柄处KNOX的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
... 的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis
1
2003
... BLADE ON PETIOLE1 (BOP1)和BOP2属于BTB蛋白家族, 两者能形成二聚物, 发挥转录激活因子的功能.BOP1和BOP2在侧生器官基部表达, 在叶基部表达的BOP能直接正调控AS2, 抑制该区域KNOX基因的表达(Jun et al, 2010).bop1和bop2双突体的叶片变大, 在叶柄处长出叶齿、叶翅等(Ha et al, 2003, 2010; Norberg et al, 2005).因此, BOP通过抑制叶基部和叶柄处KNOX的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
BLADE-ON-PETIOLE1 coordinates organ determinacy and axial polarity in Arabidopsis by directly activating ASYMMETRIC LEAVES2
1
2010
... BLADE ON PETIOLE1 (BOP1)和BOP2属于BTB蛋白家族, 两者能形成二聚物, 发挥转录激活因子的功能.BOP1和BOP2在侧生器官基部表达, 在叶基部表达的BOP能直接正调控AS2, 抑制该区域KNOX基因的表达(Jun et al, 2010).bop1和bop2双突体的叶片变大, 在叶柄处长出叶齿、叶翅等(Ha et al, 2003, 2010; Norberg et al, 2005).因此, BOP通过抑制叶基部和叶柄处KNOX的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
Auxin influx importers modulate serration along the leaf margin
Roles of miR319 and TCP transcription factors in leaf development
2
2017
... 另一组能调控叶缘复杂性的转录因子是受miR319负调控的TCPs (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF)家族成员.MIR319A和MIR319B单突变一定程度上抑制拟南芥叶缘齿裂的形成, 双突变使得拟南芥叶变全缘(Koyama et al, 2017).过表达MIR319, TCP水平降低, 导致拟南芥叶齿增多、碎米荠小叶增多(Palatnik et al, 2003; Daniel et al, 2004).Rubio-Somoza和他的团队将miR164-CUC与miR319-TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低.其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017). ...
... ; Koyama et al, 2017). ...
PIN and AUX/LAX proteins: Their role in auxin accumulation
The balance between the MIR164A and CUC2 genes controls leaf margin serration in Arabidopsis
3
2006
... PIN1的极性定位受一类转录因子NAM (CUC)的调控.NAM (NO APICAL MERISTEM)首先在矮牵牛(Petunia hybrida)中发现, 其作用主要是决定植物边缘器官和形态的建成(Souer et al, 1996).拟南芥等十字花科植物中的NAM直系同源基因有两个, CUC1和CUC2 (CUP-SHAPED COTYLEDON1, 2), 其中cuc2突变体表现为两枚子叶融合为杯状, 相邻的花萼、雄蕊等边缘也相互融合(Aida et al, 1997).NAM (CUC1 2)的旁系同源拷贝为CUC3, 该基因在功能上与前者存在部分冗余(Vroemen et al, 2003; Hasson et al, 2011).一系列实验研究表明, CUC/NAM转录因子对叶缘齿/裂的形成起着重要调控作用: 拟南芥的CUC2和CUC3在叶齿之间凹陷处均有表达信号, cuc2突变体的叶片裂刻消失变为全缘, cuc3的叶边缘裂齿程度有所降低; 而cuc1对叶缘形态基本没有影响.CUC2的作用是在叶片发育早期决定齿的形成, 而CUC3则是维持齿的生长(Nikovics et al, 2006; Kawamura et al, 2010; Bilsborough et al, 2011; Hasson et al, 2011).另外, 在具有复叶的植物中, 沉默NAM/CUC类基因对小叶的形成也有影响, 其结果使碎米荠和番茄等的小叶融合、小叶数目和裂齿程度下降(Blein et al, 2008). ...
... 关于NAM/CUC类基因对叶边缘形态发育的调控, 实验数据结合计算机模拟形成了一个基本模型, 即: 叶齿(serration)是一个空间上特异分布的调控模块的形态学产物, 该模块的核心是CUC2和生长素的最大作用位点沿发育中的叶边缘间隔分布.具体的调控模型如图1所示: CUC2的表达随叶的生长在叶边缘呈不连续分布, 受其调控, PIN1蛋白在细胞中重新定位, 将生长素以相对的方向逆浓度梯度运输到一个汇聚点, 此处即长出叶齿; 而逐渐积累的生长素又通过激活一类miRNA——miR164对CUC2进行负调控.因此, CUC2和生长素之间形成一个反馈作用环, CUC2调控生长素的极性汇聚, 而生长素的汇聚又抑制CUC2的表达, 致使CUC2的活跃区与生长素的最大浓度区相互间隔, 映射到表型上则是: 叶齿突出部位是生长素的最大浓度区, 而凹陷处是CUC2的活跃表达区.在此调控模块中, CUC2和MIR164A在共同区域表达, 其作用呈动态平衡; 生长素及其输出载体蛋白PIN1既是叶齿发育的直接作用者, 又是调控因子间相互作用的介导者(Nikovics et al, 2006; Barkoulas et al, 2007; Bilsborough et al, 2011; Bar & Ori, 2014). ...
... 植物miRNA负调控内源性靶基因, 通过与靶基因转录物的部分或完全结合来识别和裂解这些转录物.在叶发育过程中, miRNAs与其靶向基因共同作用于生长素的输出通路, 从而调控叶缘齿/裂的发育.研究表明, 生长素最大积累瞬时诱导MIR164的表达, miR164通过对靶基因CUC2的转录后负调控来影响器官原基的边界建立.利用T-DNA插入方法获得的拟南芥mir164突变体, MIR164表达量降低导致CUC2表达量升高, 叶片裂刻加深(Nikovics et al, 2006), 而当miR164过表达时, CUC2表达降低, 导致拟南芥叶变全缘、碎米荠小叶变少(Rubio-Somoza et al, 2014).CUC2的旁系同源基因CUC3缺乏miR164的结合位点而不受其调控, 但CUC3与CUC2之间存在相互作用, CUC2能促进CUC3活性, 两者形成的异源二聚体在叶片发育过程中调节叶齿/裂的发育和叶缘复杂性的建成(Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014). ...
The BLADE ON PETIOLE genes act redundantly to control the growth and development of lateral organs
1
2005
... BLADE ON PETIOLE1 (BOP1)和BOP2属于BTB蛋白家族, 两者能形成二聚物, 发挥转录激活因子的功能.BOP1和BOP2在侧生器官基部表达, 在叶基部表达的BOP能直接正调控AS2, 抑制该区域KNOX基因的表达(Jun et al, 2010).bop1和bop2双突体的叶片变大, 在叶柄处长出叶齿、叶翅等(Ha et al, 2003, 2010; Norberg et al, 2005).因此, BOP通过抑制叶基部和叶柄处KNOX的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
Transcriptomic analysis suggests a key role for SQUAMOSA PROMOTER BINDING PROTEIN LIKE, NAC and YUCCA genes in the heteroblastic development of the temperate rainforest tree Gevuina avellana (Proteaceae)
... 另一组能调控叶缘复杂性的转录因子是受miR319负调控的TCPs (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF)家族成员.MIR319A和MIR319B单突变一定程度上抑制拟南芥叶缘齿裂的形成, 双突变使得拟南芥叶变全缘(Koyama et al, 2017).过表达MIR319, TCP水平降低, 导致拟南芥叶齿增多、碎米荠小叶增多(Palatnik et al, 2003; Daniel et al, 2004).Rubio-Somoza和他的团队将miR164-CUC与miR319-TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低.其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017). ...
... -TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低.其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017). ...
PIN proteins perform a rate-limiting function in cellular auxin efflux
Arabidopsis thaliana leaf form evolved via loss of KNOX expression in leaves in association with a selective sweep
6
2010
... KNOX家族成员在植物顶端分生组织中表达, 其调控作用对叶和花的发生、发育都是必不可少且是保守的(Bharathan et al, 2002; Piazza et al, 2010).KNOX基因在叶原基形成的起始时期表达下调或没有表达, 但随后在叶片发育过程中其表达存在物种特异性, 这与叶缘形态的多样性有关(Piazza et al, 2010).在绝大多数双子叶植物中, KNOX的表达活性与叶缘的复杂性高度相关, 在很多具羽状复叶或叶缘有裂刻的物种中可检测到KNOX在叶片中表达 (Bharathan et al, 2002; Piazza et al, 2010), 如在碎米荠、番茄的复叶里, KNOX基因在叶片发育过程中有表达.拟南芥中存在全缘叶的植株类型, Piazza等(2010)在用15个基因位点的DNA序列信息对拟南芥属及十字花科其他支系的物种进行系统发生分析基础上, 推断拟南芥中的齿裂叶为祖征, 全缘叶为衍生, 而齿裂的消失与叶片中STM (KNOX家族成员)的表达缺失有关, 因为在其他叶缘分裂的物种的发育叶片中均检测到STM的表达, 且敲减其表达后叶裂消失; 进一步的分析显示该表达缺失可能与STM启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... 基因在叶原基形成的起始时期表达下调或没有表达, 但随后在叶片发育过程中其表达存在物种特异性, 这与叶缘形态的多样性有关(Piazza et al, 2010).在绝大多数双子叶植物中, KNOX的表达活性与叶缘的复杂性高度相关, 在很多具羽状复叶或叶缘有裂刻的物种中可检测到KNOX在叶片中表达 (Bharathan et al, 2002; Piazza et al, 2010), 如在碎米荠、番茄的复叶里, KNOX基因在叶片发育过程中有表达.拟南芥中存在全缘叶的植株类型, Piazza等(2010)在用15个基因位点的DNA序列信息对拟南芥属及十字花科其他支系的物种进行系统发生分析基础上, 推断拟南芥中的齿裂叶为祖征, 全缘叶为衍生, 而齿裂的消失与叶片中STM (KNOX家族成员)的表达缺失有关, 因为在其他叶缘分裂的物种的发育叶片中均检测到STM的表达, 且敲减其表达后叶裂消失; 进一步的分析显示该表达缺失可能与STM启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... ; Piazza et al, 2010), 如在碎米荠、番茄的复叶里, KNOX基因在叶片发育过程中有表达.拟南芥中存在全缘叶的植株类型, Piazza等(2010)在用15个基因位点的DNA序列信息对拟南芥属及十字花科其他支系的物种进行系统发生分析基础上, 推断拟南芥中的齿裂叶为祖征, 全缘叶为衍生, 而齿裂的消失与叶片中STM (KNOX家族成员)的表达缺失有关, 因为在其他叶缘分裂的物种的发育叶片中均检测到STM的表达, 且敲减其表达后叶裂消失; 进一步的分析显示该表达缺失可能与STM启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... 基因在叶片发育过程中有表达.拟南芥中存在全缘叶的植株类型, Piazza等(2010)在用15个基因位点的DNA序列信息对拟南芥属及十字花科其他支系的物种进行系统发生分析基础上, 推断拟南芥中的齿裂叶为祖征, 全缘叶为衍生, 而齿裂的消失与叶片中STM (KNOX家族成员)的表达缺失有关, 因为在其他叶缘分裂的物种的发育叶片中均检测到STM的表达, 且敲减其表达后叶裂消失; 进一步的分析显示该表达缺失可能与STM启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... 启动子区域序列的变异有关(Piazza et al, 2010).另外, 在叶片中异常表达KNOX基因可引起叶缘形态的强烈反应: 过表达KNOX基因时, 拟南芥、烟草以及番茄的La突变体(为单叶)出现增多的叶裂; 降低碎米荠中STM的表达时, 小叶发育被严重抑制, 甚至形成单叶, 而过表达KNOX则引起小叶增加(Hay & Tsiantis, 2006; Shani et al, 2009; Efroni et al, 2010); 在野生番茄叶中过表达玉米的KN1, 产生超级复叶, 小叶数目剧增(Hareven et al, 1996; Efroni et al, 2010).可见, 无论是小叶还是叶缘裂刻的发育都受到KNOX家族基因的调控, 其作用机制可能是KNOX通过负调控赤霉素的生物合成基因或促进细胞分裂素的生物合成而改变激素之间的平衡, 从而控制叶缘形态的发育(Efroni et al, 2010; Moon & Hake, 2011).然而也有研究表明, 具多裂叶的豆类植物叶片上并没有KNOX基因的表达(Piazza et al, 2010). ...
... 基因的表达(Piazza et al, 2010). ...
Temporal control of leaf complexity by miRNA-regulated licensing of protein complexes
5
2014
... 植物miRNA负调控内源性靶基因, 通过与靶基因转录物的部分或完全结合来识别和裂解这些转录物.在叶发育过程中, miRNAs与其靶向基因共同作用于生长素的输出通路, 从而调控叶缘齿/裂的发育.研究表明, 生长素最大积累瞬时诱导MIR164的表达, miR164通过对靶基因CUC2的转录后负调控来影响器官原基的边界建立.利用T-DNA插入方法获得的拟南芥mir164突变体, MIR164表达量降低导致CUC2表达量升高, 叶片裂刻加深(Nikovics et al, 2006), 而当miR164过表达时, CUC2表达降低, 导致拟南芥叶变全缘、碎米荠小叶变少(Rubio-Somoza et al, 2014).CUC2的旁系同源基因CUC3缺乏miR164的结合位点而不受其调控, 但CUC3与CUC2之间存在相互作用, CUC2能促进CUC3活性, 两者形成的异源二聚体在叶片发育过程中调节叶齿/裂的发育和叶缘复杂性的建成(Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014). ...
... ; Rubio-Somoza et al, 2014). ...
... 另一组能调控叶缘复杂性的转录因子是受miR319负调控的TCPs (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF)家族成员.MIR319A和MIR319B单突变一定程度上抑制拟南芥叶缘齿裂的形成, 双突变使得拟南芥叶变全缘(Koyama et al, 2017).过表达MIR319, TCP水平降低, 导致拟南芥叶齿增多、碎米荠小叶增多(Palatnik et al, 2003; Daniel et al, 2004).Rubio-Somoza和他的团队将miR164-CUC与miR319-TCP两条miRNA调节通路在蛋白水平联系起来, 表明TCP4能分别与CUC2和CUC3形成二聚体, 从而阻止CUC2-CUC3二聚体的形成, 导致叶齿/裂程度降低.其他研究者的相关研究也都表明, TCP类转录因子可负调控CUC类基因而发挥对叶边缘形态的调控作用(Palatnik et al, 2003; Daniel et al, 2004; Chitwood & Sinha, 2014; Rubio-Somoza et al, 2014; Koyama et al, 2017). ...
... A model for the molecular regulation of the development of leaf marginal serration according to studies on Arabidopsis thaliana and Cardamine hirsute (Barkoulas et al, 2007; Runions & Tsiantis, 2017). At the heart of the model is a feedback loop between CUC2 and auxin activities. CUC2 is required for PIN1-mediated auxin polar transport (horrow arrows); in turn, auxin activity maxima at the tip of the developing serration activates miR164 which represses CUC2 posttranscriptionally and generates an interspersed pattern of auxin maxima and CUC2 expression at the leaf margin. MIR164 and CUC2 are expressed in partially overlapping regions at the sinus of the serrations. Auxin enhances outgrowth of the serrations. There are additional growth regulators modulating leaf growth to shape the form of protrusions, for instance, RCO inhibits growth in indentations, producing more dissected forms. ...
... RCO调控叶裂的机制在于能确保小叶/裂片形成位置之间的叶边缘区域细胞的增殖和生长受到阻遏, 其功能仅限于对叶缘部分区域的生长起抑制作用.在碎米荠中, RCO基因缺失并不会产生除叶形变化以外的任何其他可见变化(Vlad et al, 2014).另外, 在十字花科中, RCO并没有对生长素的运输产生影响(Vlad et al, 2014), 但它能在CUC-auxin调控模块的基础上抑制叶齿两侧细胞的生长, 从而改变齿裂的深度, 产生更多的齿裂形态式样(Runions & Tsiantis, 2017) (图1). ...
To be serrate or pinnate: Diverse leaf forms of yarrows (Achillea) are linked to differential expression patterns of NAM genes
The No Apical Meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries
1
1996
... PIN1的极性定位受一类转录因子NAM (CUC)的调控.NAM (NO APICAL MERISTEM)首先在矮牵牛(Petunia hybrida)中发现, 其作用主要是决定植物边缘器官和形态的建成(Souer et al, 1996).拟南芥等十字花科植物中的NAM直系同源基因有两个, CUC1和CUC2 (CUP-SHAPED COTYLEDON1, 2), 其中cuc2突变体表现为两枚子叶融合为杯状, 相邻的花萼、雄蕊等边缘也相互融合(Aida et al, 1997).NAM (CUC1 2)的旁系同源拷贝为CUC3, 该基因在功能上与前者存在部分冗余(Vroemen et al, 2003; Hasson et al, 2011).一系列实验研究表明, CUC/NAM转录因子对叶缘齿/裂的形成起着重要调控作用: 拟南芥的CUC2和CUC3在叶齿之间凹陷处均有表达信号, cuc2突变体的叶片裂刻消失变为全缘, cuc3的叶边缘裂齿程度有所降低; 而cuc1对叶缘形态基本没有影响.CUC2的作用是在叶片发育早期决定齿的形成, 而CUC3则是维持齿的生长(Nikovics et al, 2006; Kawamura et al, 2010; Bilsborough et al, 2011; Hasson et al, 2011).另外, 在具有复叶的植物中, 沉默NAM/CUC类基因对小叶的形成也有影响, 其结果使碎米荠和番茄等的小叶融合、小叶数目和裂齿程度下降(Blein et al, 2008). ...
A secreted peptide and its receptors shape the auxin response pattern and leaf margin morphogenesis
2
2016
... 植物中一种名为EPFL2 (EPIDERMAL PATTERNING FACTOR-LIKE)的肽和ERECTA (ER)家族成员也参与叶边缘锯齿的形态发生过程(Tameshige et al, 2016).当拟南芥中的EPFL2或ER发生突变时, 叶片边缘会变得圆滑, 并检测到生长素在整个叶缘区分布(在具有齿的野生型叶片中, 叶边缘的生长素只在齿的位置累积).EPFL2与ERECTA形成配体-受体对, 从而抑制生长素在叶齿处的响应, 而生长素又反过来抑制EPFL2的表达, 形成负反馈环.该反馈系统在叶缘生长过程中维持着生长素的响应式样(Tameshige et al, 2016). ...
... 的表达, 形成负反馈环.该反馈系统在叶缘生长过程中维持着生长素的响应式样(Tameshige et al, 2016). ...
Arabidopsis type II Phosphatidylinositol 4-Kinase PI4Kγ5 regulates auxin biosynthesis and leaf margin development through interacting with membrane-bound transcription factor ANAC078
The TIE1 transcriptional repressor links TCP transcription factors with TOPLESS/ TOPLESS-RELATED corepressors and modulates leaf development in Arabidopsis
2
2013
... 上述调控模块中的TCP转录因子的活性又受到TIE1 (TCP INTERACTOR-CONTAINING EAR MOTIF PROTEIN1)转录抑制因子的作用: 在叶缘形态发育中TIE1能招募共抑制因子TOPLESS (TPL)/TOPLESS-RELATED (TPR)在蛋白水平上抑制TCP的活性, 因此, 过表达TIE1能引起叶边缘裂刻的加剧(Tao et al, 2013); 而TIE1又与TEAR1 (TIE1-ASSOCIATED RING-TYPE E3 LIGASE1)有相互作用, TEAR1及其同源基因发生突变可引起叶片边缘锯齿增多(Zhang et al, 2017).秦跟基课题组(Tao et al, 2013; Zhang et al, 2017)根据其实验结果提出一种新的精确调控TCP活性的叶缘形态发育调控机制: TIE1招募TPL/TPRs共同抑制TCP的活性, 而TEAR1通过介导TIE1的降解来解除TIE1对TCP的抑制, 从而正调控TCP的活性.可见, TIE1和TEAR1通过对TCP的调控而间接影响了叶缘的形态发育. ...
... ).秦跟基课题组(Tao et al, 2013; Zhang et al, 2017)根据其实验结果提出一种新的精确调控TCP活性的叶缘形态发育调控机制: TIE1招募TPL/TPRs共同抑制TCP的活性, 而TEAR1通过介导TIE1的降解来解除TIE1对TCP的抑制, 从而正调控TCP的活性.可见, TIE1和TEAR1通过对TCP的调控而间接影响了叶缘的形态发育. ...
The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings
Divide et impera: Boundaries shape the plant body and initiate new meristems
2
2015
... BLADE ON PETIOLE1 (BOP1)和BOP2属于BTB蛋白家族, 两者能形成二聚物, 发挥转录激活因子的功能.BOP1和BOP2在侧生器官基部表达, 在叶基部表达的BOP能直接正调控AS2, 抑制该区域KNOX基因的表达(Jun et al, 2010).bop1和bop2双突体的叶片变大, 在叶柄处长出叶齿、叶翅等(Ha et al, 2003, 2010; Norberg et al, 2005).因此, BOP通过抑制叶基部和叶柄处KNOX的表达来保证正常的叶片形态建成(Ha et al, 2010; Wang et al, 2015). ...
... JAGGED LATERAL ORGANS (JLO)被认为是拟南芥中器官边界建立的“调节器” (Wang et al, 2015), 属于LATERAL ORGAN BOUNDARY DOMAIN (LBD) 基因家族成员, 在分生组织和器官原基的边界特异表达.JLO能诱导叶片中KNOX家族STM和KNAT1的表达, 并抑制PIN的基因表达, 当JLO在拟南芥叶片异位表达时可产生深裂、叶柄减短的叶片(Borghi et al, 2007). ...
The Arabidopsis RING-type E3 ligase TEAR1 controls leaf development by targeting the TIE1 transcriptional repressor for degradation
2
2017
... 上述调控模块中的TCP转录因子的活性又受到TIE1 (TCP INTERACTOR-CONTAINING EAR MOTIF PROTEIN1)转录抑制因子的作用: 在叶缘形态发育中TIE1能招募共抑制因子TOPLESS (TPL)/TOPLESS-RELATED (TPR)在蛋白水平上抑制TCP的活性, 因此, 过表达TIE1能引起叶边缘裂刻的加剧(Tao et al, 2013); 而TIE1又与TEAR1 (TIE1-ASSOCIATED RING-TYPE E3 LIGASE1)有相互作用, TEAR1及其同源基因发生突变可引起叶片边缘锯齿增多(Zhang et al, 2017).秦跟基课题组(Tao et al, 2013; Zhang et al, 2017)根据其实验结果提出一种新的精确调控TCP活性的叶缘形态发育调控机制: TIE1招募TPL/TPRs共同抑制TCP的活性, 而TEAR1通过介导TIE1的降解来解除TIE1对TCP的抑制, 从而正调控TCP的活性.可见, TIE1和TEAR1通过对TCP的调控而间接影响了叶缘的形态发育. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}