亚热带常绿阔叶林在我国分布面积最广, 类型最为复杂多样, 是全球常绿阔叶林的主体(宋永昌, 2005 )。我国的中亚热带是常绿阔叶林的典型分布区(吴征镒,1980 ), 中亚热带常绿阔叶林有着丰富的生物多样性资源和巨大的生态效益, 越来越引起人们广泛的重视。
古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分。长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林。但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 )。
古田山四季分明, 降水充沛, 年均温度15.3℃, 最高气温为38.1℃ (7月), 最低气温为-6.8℃ (1月), 年均降雨量1,963.7 mm, 每年的3-6月为第一雨季, 7-8月为伏旱期, 8月底至9月受台风影响形成第二雨季, 10月到翌年2月为干旱期(开化林业志编写组, 1988 )。该区域地形复杂, 山势陡峭, 山体呈东北-西南走向, 最高海拔1,258 m。该区域物种多样性丰富, 自然分布的种子植物有149科648属1,426种(包括种下分类等级), 分别占浙江省种子植物科、属、种总数的81.9%, 51.7%和41.6%, 其中国家二、三级重点保护植物18种, 并且还有一些珍稀群落, 如分布集中且面积较大的香果树(Emmenopterys benriyi )、野含笑(Michelia skinneriana )和紫茎(Stewartia sinensis )等植物群落(楼炉焕和金水虎, 2000 )。
许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 )。虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识。本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征。
1 材料与方法
1.1 样地设置与调查
古田山24 ha样地位于古田山保护区内, 东西长600 m, 南北长400 m, 样地内地形复杂多变, 地势较陡, 坡度范围13°-62°, 最高海拔为714.9 m, 最低海拔为446.3 m, 平均海拔为580.6 m, 最大高差为268.6 m, 植被为典型中亚热带低海拔常绿阔叶原始林(祝燕等, 2008 )。2005年按照CTFS (Centre for Tropical Forest Science)样地的建设方法(Condit, 1998 ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 )。本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种。
1.2 数据处理
将24 ha样地划分成24个1 ha小样地, 分别计算: (1) 1 ha小样地所有物种的重要值, 分析群落优势物种在空间上的变化; (2) 1 ha小样地α多样性, 包括物种个体数、物种丰富度、Shannon-Wiener指数及Pielou均匀度指数; (3) 1 ha小样地内β多样性, 表征1 ha小样地内物种的替代速率; (4)任意两个相邻样地的β多样性, 分析样地间物种替代速率的空间分布。采用的多样性测度指标如下(钱迎倩和马克平, 1994 ):
物种丰富度(S ): 1 ha小样地内胸径 ≥ 1 cm木本植物个体所属的物种数量。
Shannon-Wiener指数(H ):
$H=—\sum\limits_{i=1}^{S}P_{i}log(P_{i})$ (1)
Pi = Ni /N 0 , Ni 为第i 个物种的多度, N 0 为所有物种的多度总和, 即小样地内所有胸径 ≥ 1 cm的木本植物个体数。
Pielou均匀度指数(Jsw ):
$J_{sw}=H/logS$ (2)
样地内β 多样性Whittaker指数(βw ):
$\beta_{w}=\frac{S}{\alpha}-1$ (3)
α为1 ha小样地内所有20 m × 20 m小样方物种数的平均数。
样地间β多样性Jaccard指数($\beta_{J}$):
$\beta_{J}=1-\frac{S_{12}}{S_{1}+S_{2}-S_{12}}$ (4)
$S_{1}$为相邻样地1的物种数, $S_{2}$为相邻样地2的物种数, $S_{12}$为样地1、2共有的物种数。
采用R软件(Version 3.4.3)执行所有分析、计算和作图。
2 结果
2.1 小样地(1 ha)间α多样性的变异特征
24个1 ha小样地第一优势物种以甜槠(Castanopsis eyrei )为主, 仅有两个小样地为木荷(Schima superba ); 第二优势物种以木荷为主, 仅有2个小样地为甜槠和马尾松(Pinus massoniana ); 但第三优势物种变化较大, 其中6个小样地为马尾松, 5个小样地为浙江新木姜子(Neolitsea aurata var. chekiangensis ), 5个小样地为马银花(Rhododendron ovatum ), 3个为浙江红花油茶(Camellia chekiangoleosa ), 其余5个1 ha小样地的优势种分别为杨梅叶蚊母树(Distylium myricoides )、甜槠、柳叶蜡梅(Chimonanthus salicifolius )、檵木(Loropetalum chinense )和短柄枹(Quercus serrata )(附录1)。
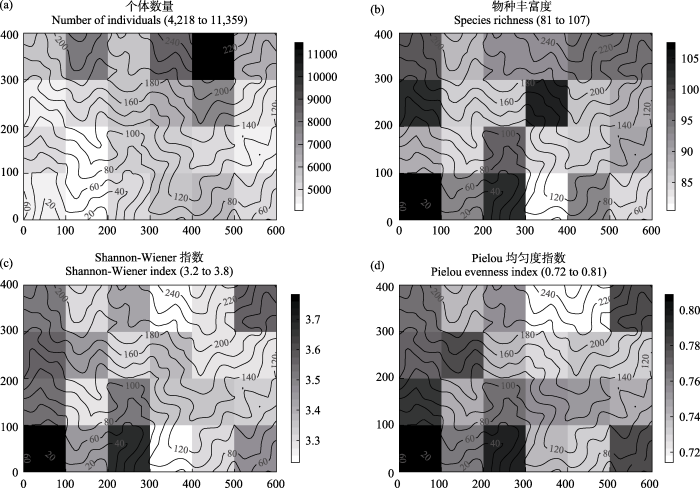
24个1 ha小样地中, 不同样地所包含的个体数差异最大, 样地木本植物个体数平均值为5,862.5± 1,599 (SD), 变异系数为27.3%; 最大值为11,359株, 位于24 ha样地最高处山脊, 相对于24 ha大样地最低点的高差(相对高差)200-260 m; 最小值为4,218, 位于相对高差80-120 m山脊(图1 a)。不同样地物种丰富度的平均值为91.8$\pm$7 (SD), 变异系数为7.8%,物种最丰富的样地位于最低位置山谷(107个物种),物种最少的样地位于相对高差80-140 m的山脊(81个物种)(图1 b)。Shannon-Wiener指数平均值为3.41 ± 0.14 (SD), 变异系数为4.1%, 最大值为3.8, 位于样地最低位置山谷, 与物种数最多的样地重合, 最小值为3.2, 位于样地相对高差180-260 m的较高位置(图1 c)。Pielou均匀度指数变异最小, 平均值为0.76, 变异系数为3.3%, 最大值为0.81, 与物种数最丰富的样地重合, 最小值为0.72, 位于样地最高位置, 与Shannon-Wiener指数最小值重叠(图1 d)。
图1 古田山24 ha森林大样地内1 ha小样地间α多样性的变异特征。(a) 1 ha小样地内总的木本植物个体数量; (b)物种丰富度; (c) Shannon-Wiener指数; (d) Pielou均匀度指数。背景曲线为等高线, 数字为相对于24 ha样地最低点的高差。横坐标、纵坐标分别指24 ha大样地的长度和宽度(单位: m)
Fig. 1 Spatial variation of α diversity among 24 subplots of 1-ha each within the 24 ha forest dynamics plot in Gutianshan. (a) Total number of individuals within 1-ha subplot; (b) Species richness; (c) Shannon-Wiener index; (d) Pielou evenness index. The background curves are contours and the numbers are elevations relative to the lowest point in the whole plot of 24 ha. X- and Y-axes are length and width of 24 ha plot (unit: m).
样地所包含的木本植物个体数与物种丰富度没有显著相关性(r = 0.04, P = 0.85), 与Shannon-Wiener指数和Pielou均匀度指数均呈显著负相关(r = -0.41, P = 0.04; r = -0.55, P = 0.006)。物种丰富度与Shannon-Wiener指数呈显著正相关(r = 0.66, P = 0.0005), 而与Pielou均匀度指数没有相关性。Shannon-Wiener指数与Pielou均匀度指数呈极强的正相关关系(r = 0.92, P << 0.0001)。
2.2 小样地内和小样地间β多样性的变异特征
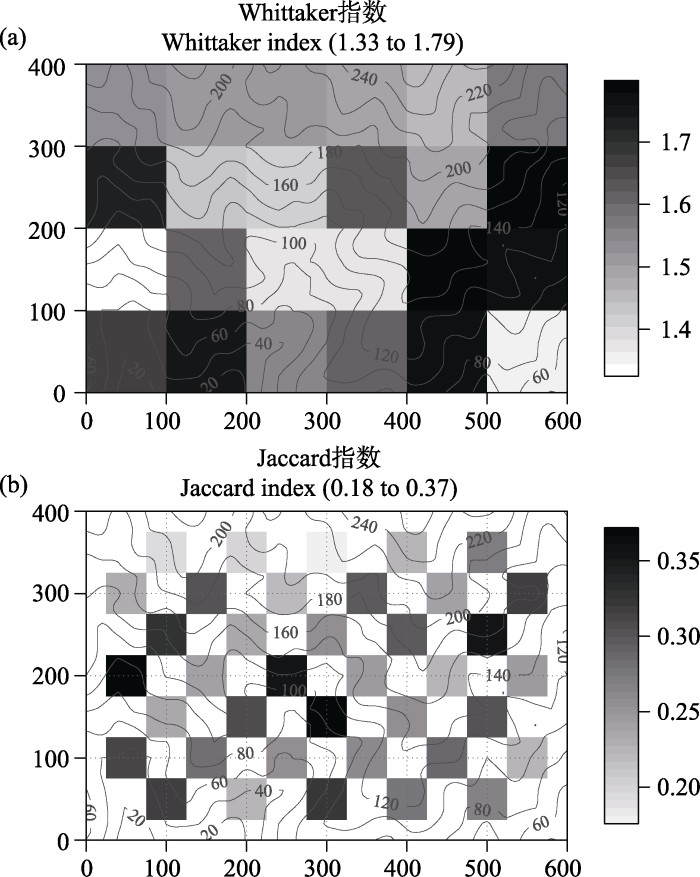
样地内物种的替代速率空间变异较大, Whittaker指数的平均值为1.56 ± 0.15 (SD), 变异系数为9.5%, 最大值为1.79, 位于相对高差约100-180 m的小样地, 最小值为1.33, 位于相对高差40-120 m的小样地(图2 a); 小样地内物种替代速率与小样地α多样性无显著相关性。相邻小样地间物种替代速率空间变化很大, Jaccard指数平均值为0.27 ± 0.05 (SD), 变异系数为18%, 最大值为0.37, 位于环境差异较大的相邻小样地间, 如中、低海拔山脊和沟谷样地之间物种替代速率最大; 最小值为0.18, 位于高海拔且地势相对平坦的两小样地之间(图2 b)。小样地间物种替代速率与物种丰富度格局呈现明显的差异(图2 b)。
图2 古田山24 ha森林大样地内1 ha小样地内β多样性(a)及样地间β多样性(b)的空间变异特征。背景曲线为等高线, 数字为相对于24 ha样地最低点的高差。图b灰白斑块背景为物种丰富度的空间变异(见图1 b)。
Fig. 2 Variation of β-diversity within a 1-ha plot (a) and between-nearest-neighbor-plots (b). The background curves are contours and the numbers are elevations relative to the lowest point in the whole 24 ha plot. The gray patchy ground refers to the variation in species richness among plots, the same as Fig. 1 b.
3 讨论
本研究揭示了物种多样性存在很明显的空间变异特征, 多样性测度指标在空间上的变化特征有很大差别, 这反映了森林群落组成结构在空间上有很大的变异性。传统群落生态学研究中, 通常将群落定义为特定空间或特定生境不同种群的集合(Odum & Barrette, 2009 )。除了生境空间变化, 其他生态学过程也可以使群落的空间特征发生变化, 如扩散、竞争、干扰等(Wright, 2002 ), 从而导致物种组成和群落结构的空间变化。本研究表明相邻小样地之间物种替代速率空间变异较大, 虽然各小样地的第一、二优势种基本上都是甜槠和木荷, 十分相似, 但第三优势种差别较大, 24个1 ha小样地有多达10个不同的优势物种。这意味着中亚热带常绿阔叶林群落即使在面积为1 ha时(远大于传统的亚热带常绿阔叶林最小群落面积), 继续增大面积还会增加很多物种, 或者说群落的组成和结构会随面积(尺度)的增大而改变。群落的生态学过程具有尺度依赖性并决定群落的结构和组成(Levin, 1992 ), 因此, 对群落生态学而言, 在一定的空间尺度比较群落的组成和结构,对正确认识不同群落的差异, 研究推断群落变化的生态学过程, 制定科学的保育措施有重要意义。
另外, 由于物种多样性存在空间变异, 并呈现出一定的空间结构特点, 比如邻近样地之间的物种替代速率在相对中、低海拔处较大, 在相对高海拔处较低, 并且与物种丰富度的空间格局变异有很大的差异, 这意味着随着样地面积的扩大和空间结构的变化, 物种丰富度的格局将可能会发生变化。因此, 在生物多样性的保护和管理实践中, 应该根据保护和管理的目标, 确定科学合理的空间尺度。
物种多样性的空间格局也反映了生态学过程。古田山24个1 ha小样地木本植物的个体数量与物种丰富度没有显著的相关性, 与植物个体数越多物种数量越多的假说不同(Evans et al, 2005 ), 表明植物的个体数量并不一定和物种数量的增加相同步, 这可能与本研究小样地(1 ha)的尺度较大有关。当取样尺度较小时, 生境异质性相对较弱, 取样的植物个体数量越多, 发现不同物种数量的概率就越大(Evans et al, 2005 ); 但当取样尺度增加, 不同的生境类型会增加, 物种数量的增加将主要由生境异质性或扩散限制起作用(Harms et al, 2001 ; Wright, 2002 )。个体数量和Shannon-Wiener指数及物种均匀度呈显著的负相关, 且物种均匀度和Shannon-Wiener指数高的样地都位于低海拔沟谷地带, 这可能是由于在低海拔沟谷生境中群落内各物种的种内竞争较强, 导致种群数量下降, 均匀度和多样性增加。
本研究揭示了古田山中亚热带常绿阔叶林群落物种多样性的空间变异特征, 这种变化特征说明给定空间尺度对认识和比较群落的特征具有重要意义。同时, 生态过程是多尺度变化的(Levin, 1992 ), 因此, 这种物种多样性明显的空间变化意味着: 较大的样地可能更有利于研究亚热带常绿阔叶林群落组成和结构的维持机制。
致谢: 感谢浙江大学于明坚教授、古田山国家级自然保护区管理局陈声文、方腾等对样地建设的贡献, 感谢相关单位研究生、本科生及当地合作伙伴在样地建设过程中的辛勤付出。感谢责任编委和审稿专家对本文提出的修改意见。
附录 Supplementary Material
附录1 古田山国家级自然保护区24个1 ha小样地物种信息表
Appendix 1 Species information of 24 1-ha subplots in Gutianshan National Nature Reserve, Zhejiang Province
http://www.biodiversity-science.net/fileup/PDF/2018138-1.pdf
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Chen JH Feng ZJ 2002 ) Study on geographical compositions of seed plant flora in Gutian Mountain of Zhejiang Province
. Journal of East China Normal University (Natural Science) , (1 ), 104 -111 . (in Chinese with English abstract)
[本文引用: 1]
[陈建华 , 冯志坚 (2002 ) 浙江古田山种子植物区系的地理成分研究
. 华东师范大学学报(自然科学版) , (1 ), 104 -111 .]
[本文引用: 1]
[2]
Condit R 1998 ) Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
. Springer Science & Business Media, Berlin .
[本文引用: 1]
[3]
Evans KL Warren PH Gaston KJ 2005 ) Species-energy relationships at the macroecological scale: A review of the mechanisms
. Biological Reviews , 80 , 1 -25 .
[本文引用: 2]
[4]
Harms KE Condit R Hubbell SP Foster RB 2001 ) Habitat associations of trees and shrubs in a 50-ha Neotropical forest plot
. Journal of Ecology , 89 , 947 -959 .
[本文引用: 1]
[5]
Hu ZH Yu MJ Ding BY Fang T Qian HY Chen QC 2003 ) Types of evergreen broad-leaved forests and their species diversity in Gutian Mountain National Nature Reserve
. Chinese Journal of Applied and Environmental Biology , 9 , 341 -345 . (in Chinese with English abstract)
[本文引用: 1]
[胡正华 , 于明坚 , 丁炳扬 , 方腾 , 钱海源 , 陈启瑺 (2003 ) 古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究
. 应用与环境生物学报 , 9 , 341 -345 .]
[本文引用: 1]
[6]
Hu ZH Yu MJ Fang T Yu JP Ding BY 2002 ) Study on characters of forest community in Gutianshan Nature Reserve of Zhejiang
. Journal of Zhejiang Forestry Science and Technology , 22 (6 ), 1 -4, 51 . (in Chinese with English abstract)
[本文引用: 1]
[胡正华 , 于明坚 , 方腾 , 余建平 , 丁炳扬 (2002 ) 浙江古田山自然保护区森林群落特征研究
. 浙江林业科技 , 22 (6 ), 1 -4, 51 .]
[本文引用: 1]
[7]
Hu ZH Yu MJ Suo FX 2005 ) The plant species diversity of the evergreen broad-leaved forest in Gutian Mountain National Nature Reserve
. Chinese Agricultural Science Bulletin , 21 (3 ), 134 -137 . (in Chinese with English abstract)
[本文引用: 1]
[胡正华 , 于明坚 , 索福喜 (2005 ) 古田山国家自然保护区常绿阔叶林植物物种多样性研究
. 中国农学通报 , 21 (3 ), 134 -137 .]
[本文引用: 1]
[8]
Huang JH Chen B Liu CR Lai JS Zhang JL Ma KP 2012 ) Identifying hotspots of endemic woody seed plant diversity in China
. Diversity and Distributions , 18 , 673 -688 .
[本文引用: 1]
[9]
Lai JS Mi XC Ren HB Ma KP 2010 ) Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees―a case study of 24 hm2 Gutianshan forest plot in China
. Chinese Journal of Plant Ecology , 34 , 761 -769 . (in Chinese with English abstract)
[本文引用: 1]
[赖江山 , 米湘成 , 任海保 , 马克平 (2010 ) 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
. 植物生态学报 , 34 , 761 -769 .]
[本文引用: 1]
[10]
Legendre P Mi XC Ren HB Ma KP Yu MJ Sun YF He FL 2009 ) Partitioning beta diversity in a subtropical broad-leaved forest of China
. Ecology , 90 , 663 -674 .
[本文引用: 1]
[11]
Levin SA 1992 ) The problem of pattern and scale in ecology
. Ecology , 73 , 1943 -1967 .
[本文引用: 2]
[12]
Lou LH Jin SH 2000 ) Spermatophyta flora of Gutianshan Nature Reserve in Zhejiang
. Journal of Beijing Forestry University , 22 (5 ), 33 -39 . (in Chinese with English abstract)
[本文引用: 1]
[楼炉焕 , 金水虎 (2000 ) 浙江古田山自然保护区种子植物区系分析
. 北京林业大学学报 , 22 (5 ), 33 -39 .]
[本文引用: 1]
[13]
Odum EP Barrette GW Lu JJ Wang W Wang TH He WS Li XZ 2009 ) Fundamentals of Ecology . Higher Education Press , Beijing .
[本文引用: 1]
(in Chinese) [Odum EP Barrette GW 陆健健 , 王伟 , 王天慧 , 何文珊 , 李秀珍译 ) (2009 ) 生态学基础 . 高等教育出版社 , 北京 .]
[本文引用: 1]
[14]
Qian YQ Ma KP 1994 ) The Principles and Methods of Biodiversity Research . Chinese Science and Technology Press , Beijing . (in Chinese)
[本文引用: 1]
[钱迎倩 , 马克平 (1994 ) 生物多样性研究的原理与方法 . 中国科学技术出版社 , 北京 .]
[本文引用: 1]
[15]
Song YC Chen XY Wang XH 2005 ) Studies on evergreen broad-leaved forests of China: A retrospect and prospect
. Journal of East China Normal University (Natural Science) , (1 ), 1 -8 . (in Chinese with English abstract)
[本文引用: 1]
[宋永昌 , 陈小勇 , 王希华 (2005 ) 中国常绿阔叶林研究的回顾与展望
. 华东师范大学学报(自然科学版) , (1 ), 1 -8 .]
[本文引用: 1]
[16]
Song YF Chen SW Wang W Yu JP Qian HY Wang YQ Chen L Mi XC Ren HB Ye D Chen JH Ma KP 2017 ) Effects of negative density dependence and habitat filtering on the functional diversity of seedlings in the subtropical forest of Gutianshan
. Biodiversity Science , 25 , 959 -965 . (in Chinese with English abstract)
[本文引用: 1]
[宋云峰 , 陈声文 , 王薇 , 余建平 , 钱海源 , 王云泉 , 陈磊 , 米湘成 , 任海保 , 叶铎 , 陈建华 , 马克平 (2017 ) 负密度制约和生境过滤对古田山幼苗功能多样性年际变化的影响
. 生物多样性 , 25 , 959 -965 .]
[本文引用: 1]
[17]
The Compilation Group of Kaihua Forestry Records (1988 ) Kaihua Forestry Records . Zhejiang People’s Publishing House , Hangzhou . (in Chinese)
[本文引用: 1]
[开化林业志编写组 (1988 ) 开化林业志 . 浙江人民出版社 , 杭州 .]
[本文引用: 1]
[18]
Wright JS 2002 ) Plant diversity in tropical forests: A review of mechanisms of species coexistence
. Oecologia , 130 , 1 -14 .
[本文引用: 2]
[19]
Wu ZY 1980 ) Vegetation of China . Science Press , Beijing . (in Chinese)
[本文引用: 2]
[吴征镒 (1980 ) 中国植被 . 科学出版社 , 北京 .]
[本文引用: 2]
[20]
Yu MJ Hu ZH Yu JP Ding BY Fang T 2001 ) Forest vegetation types in Gutianshan Natural Reserve in Zhejiang
. Journal of Zhejiang University (Agriculture and Life Sciences) , 27 , 375 -380 . (in Chinese with English abstract)
[本文引用: 1]
[于明坚 , 胡正华 , 余建平 , 丁炳扬 , 方腾 (2001 ) 浙江古田山自然保护区森林植被类型
. 浙江大学学报(农业与生命科学版) , 27 , 375 -380 .]
[本文引用: 1]
[21]
Zhu Y Zhao GF Zhang LW Shen GC Mi XC Ren HB Yu MJ Chen JH Chen SW Fang T Ma KP 2008 ) Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, East China
. Journal of Plant Ecology (Chinese Version) , 32 , 262 -273 . (in Chinese with English abstract)
[本文引用: 3]
[祝燕 , 赵谷风 , 张俪文 , 沈国春 , 米湘成 , 任海保 , 于明坚 , 陈建华 , 陈声文 , 方腾 , 马克平 (2008 ) 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
. 植物生态学报 , 32 , 262 -273 .]
[本文引用: 3]
浙江古田山种子植物区系的地理成分研究
1
2002
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
浙江古田山种子植物区系的地理成分研究
1
2002
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
1
1998
... 古田山24 ha样地位于古田山保护区内, 东西长600 m, 南北长400 m, 样地内地形复杂多变, 地势较陡, 坡度范围13°-62°, 最高海拔为714.9 m, 最低海拔为446.3 m, 平均海拔为580.6 m, 最大高差为268.6 m, 植被为典型中亚热带低海拔常绿阔叶原始林(祝燕等, 2008 ).2005年按照CTFS (Centre for Tropical Forest Science)样地的建设方法(Condit, 1998 ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 ).本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种. ...
Species-energy relationships at the macroecological scale: A review of the mechanisms
2
2005
... 物种多样性的空间格局也反映了生态学过程.古田山24个1 ha小样地木本植物的个体数量与物种丰富度没有显著的相关性, 与植物个体数越多物种数量越多的假说不同(Evans et al, 2005 ), 表明植物的个体数量并不一定和物种数量的增加相同步, 这可能与本研究小样地(1 ha)的尺度较大有关.当取样尺度较小时, 生境异质性相对较弱, 取样的植物个体数量越多, 发现不同物种数量的概率就越大(Evans et al, 2005 ); 但当取样尺度增加, 不同的生境类型会增加, 物种数量的增加将主要由生境异质性或扩散限制起作用(Harms et al, 2001 ; Wright, 2002 ).个体数量和Shannon-Wiener指数及物种均匀度呈显著的负相关, 且物种均匀度和Shannon-Wiener指数高的样地都位于低海拔沟谷地带, 这可能是由于在低海拔沟谷生境中群落内各物种的种内竞争较强, 导致种群数量下降, 均匀度和多样性增加. ...
... ), 表明植物的个体数量并不一定和物种数量的增加相同步, 这可能与本研究小样地(1 ha)的尺度较大有关.当取样尺度较小时, 生境异质性相对较弱, 取样的植物个体数量越多, 发现不同物种数量的概率就越大(Evans et al, 2005 ); 但当取样尺度增加, 不同的生境类型会增加, 物种数量的增加将主要由生境异质性或扩散限制起作用(Harms et al, 2001 ; Wright, 2002 ).个体数量和Shannon-Wiener指数及物种均匀度呈显著的负相关, 且物种均匀度和Shannon-Wiener指数高的样地都位于低海拔沟谷地带, 这可能是由于在低海拔沟谷生境中群落内各物种的种内竞争较强, 导致种群数量下降, 均匀度和多样性增加. ...
Habitat associations of trees and shrubs in a 50-ha Neotropical forest plot
1
2001
... 物种多样性的空间格局也反映了生态学过程.古田山24个1 ha小样地木本植物的个体数量与物种丰富度没有显著的相关性, 与植物个体数越多物种数量越多的假说不同(Evans et al, 2005 ), 表明植物的个体数量并不一定和物种数量的增加相同步, 这可能与本研究小样地(1 ha)的尺度较大有关.当取样尺度较小时, 生境异质性相对较弱, 取样的植物个体数量越多, 发现不同物种数量的概率就越大(Evans et al, 2005 ); 但当取样尺度增加, 不同的生境类型会增加, 物种数量的增加将主要由生境异质性或扩散限制起作用(Harms et al, 2001 ; Wright, 2002 ).个体数量和Shannon-Wiener指数及物种均匀度呈显著的负相关, 且物种均匀度和Shannon-Wiener指数高的样地都位于低海拔沟谷地带, 这可能是由于在低海拔沟谷生境中群落内各物种的种内竞争较强, 导致种群数量下降, 均匀度和多样性增加. ...
古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究
1
2003
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究
1
2003
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
浙江古田山自然保护区森林群落特征研究
1
2002
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
浙江古田山自然保护区森林群落特征研究
1
2002
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
古田山国家自然保护区常绿阔叶林植物物种多样性研究
1
2005
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
古田山国家自然保护区常绿阔叶林植物物种多样性研究
1
2005
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
Identifying hotspots of endemic woody seed plant diversity in China
1
2012
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
Partitioning beta diversity in a subtropical broad-leaved forest of China
1
2009
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
The problem of pattern and scale in ecology
2
1992
... 本研究揭示了物种多样性存在很明显的空间变异特征, 多样性测度指标在空间上的变化特征有很大差别, 这反映了森林群落组成结构在空间上有很大的变异性.传统群落生态学研究中, 通常将群落定义为特定空间或特定生境不同种群的集合(Odum & Barrette, 2009 ).除了生境空间变化, 其他生态学过程也可以使群落的空间特征发生变化, 如扩散、竞争、干扰等(Wright, 2002 ), 从而导致物种组成和群落结构的空间变化.本研究表明相邻小样地之间物种替代速率空间变异较大, 虽然各小样地的第一、二优势种基本上都是甜槠和木荷, 十分相似, 但第三优势种差别较大, 24个1 ha小样地有多达10个不同的优势物种.这意味着中亚热带常绿阔叶林群落即使在面积为1 ha时(远大于传统的亚热带常绿阔叶林最小群落面积), 继续增大面积还会增加很多物种, 或者说群落的组成和结构会随面积(尺度)的增大而改变.群落的生态学过程具有尺度依赖性并决定群落的结构和组成(Levin, 1992 ), 因此, 对群落生态学而言, 在一定的空间尺度比较群落的组成和结构,对正确认识不同群落的差异, 研究推断群落变化的生态学过程, 制定科学的保育措施有重要意义. ...
... 本研究揭示了古田山中亚热带常绿阔叶林群落物种多样性的空间变异特征, 这种变化特征说明给定空间尺度对认识和比较群落的特征具有重要意义.同时, 生态过程是多尺度变化的(Levin, 1992 ), 因此, 这种物种多样性明显的空间变化意味着: 较大的样地可能更有利于研究亚热带常绿阔叶林群落组成和结构的维持机制. ...
浙江古田山自然保护区种子植物区系分析
1
2000
... 古田山四季分明, 降水充沛, 年均温度15.3℃, 最高气温为38.1℃ (7月), 最低气温为-6.8℃ (1月), 年均降雨量1,963.7 mm, 每年的3-6月为第一雨季, 7-8月为伏旱期, 8月底至9月受台风影响形成第二雨季, 10月到翌年2月为干旱期(开化林业志编写组, 1988 ).该区域地形复杂, 山势陡峭, 山体呈东北-西南走向, 最高海拔1,258 m.该区域物种多样性丰富, 自然分布的种子植物有149科648属1,426种(包括种下分类等级), 分别占浙江省种子植物科、属、种总数的81.9%, 51.7%和41.6%, 其中国家二、三级重点保护植物18种, 并且还有一些珍稀群落, 如分布集中且面积较大的香果树(Emmenopterys benriyi )、野含笑(Michelia skinneriana )和紫茎(Stewartia sinensis )等植物群落(楼炉焕和金水虎, 2000 ). ...
浙江古田山自然保护区种子植物区系分析
1
2000
... 古田山四季分明, 降水充沛, 年均温度15.3℃, 最高气温为38.1℃ (7月), 最低气温为-6.8℃ (1月), 年均降雨量1,963.7 mm, 每年的3-6月为第一雨季, 7-8月为伏旱期, 8月底至9月受台风影响形成第二雨季, 10月到翌年2月为干旱期(开化林业志编写组, 1988 ).该区域地形复杂, 山势陡峭, 山体呈东北-西南走向, 最高海拔1,258 m.该区域物种多样性丰富, 自然分布的种子植物有149科648属1,426种(包括种下分类等级), 分别占浙江省种子植物科、属、种总数的81.9%, 51.7%和41.6%, 其中国家二、三级重点保护植物18种, 并且还有一些珍稀群落, 如分布集中且面积较大的香果树(Emmenopterys benriyi )、野含笑(Michelia skinneriana )和紫茎(Stewartia sinensis )等植物群落(楼炉焕和金水虎, 2000 ). ...
1
2009
... 本研究揭示了物种多样性存在很明显的空间变异特征, 多样性测度指标在空间上的变化特征有很大差别, 这反映了森林群落组成结构在空间上有很大的变异性.传统群落生态学研究中, 通常将群落定义为特定空间或特定生境不同种群的集合(Odum & Barrette, 2009 ).除了生境空间变化, 其他生态学过程也可以使群落的空间特征发生变化, 如扩散、竞争、干扰等(Wright, 2002 ), 从而导致物种组成和群落结构的空间变化.本研究表明相邻小样地之间物种替代速率空间变异较大, 虽然各小样地的第一、二优势种基本上都是甜槠和木荷, 十分相似, 但第三优势种差别较大, 24个1 ha小样地有多达10个不同的优势物种.这意味着中亚热带常绿阔叶林群落即使在面积为1 ha时(远大于传统的亚热带常绿阔叶林最小群落面积), 继续增大面积还会增加很多物种, 或者说群落的组成和结构会随面积(尺度)的增大而改变.群落的生态学过程具有尺度依赖性并决定群落的结构和组成(Levin, 1992 ), 因此, 对群落生态学而言, 在一定的空间尺度比较群落的组成和结构,对正确认识不同群落的差异, 研究推断群落变化的生态学过程, 制定科学的保育措施有重要意义. ...
1
2009
... 本研究揭示了物种多样性存在很明显的空间变异特征, 多样性测度指标在空间上的变化特征有很大差别, 这反映了森林群落组成结构在空间上有很大的变异性.传统群落生态学研究中, 通常将群落定义为特定空间或特定生境不同种群的集合(Odum & Barrette, 2009 ).除了生境空间变化, 其他生态学过程也可以使群落的空间特征发生变化, 如扩散、竞争、干扰等(Wright, 2002 ), 从而导致物种组成和群落结构的空间变化.本研究表明相邻小样地之间物种替代速率空间变异较大, 虽然各小样地的第一、二优势种基本上都是甜槠和木荷, 十分相似, 但第三优势种差别较大, 24个1 ha小样地有多达10个不同的优势物种.这意味着中亚热带常绿阔叶林群落即使在面积为1 ha时(远大于传统的亚热带常绿阔叶林最小群落面积), 继续增大面积还会增加很多物种, 或者说群落的组成和结构会随面积(尺度)的增大而改变.群落的生态学过程具有尺度依赖性并决定群落的结构和组成(Levin, 1992 ), 因此, 对群落生态学而言, 在一定的空间尺度比较群落的组成和结构,对正确认识不同群落的差异, 研究推断群落变化的生态学过程, 制定科学的保育措施有重要意义. ...
1
1994
... 将24 ha样地划分成24个1 ha小样地, 分别计算: (1) 1 ha小样地所有物种的重要值, 分析群落优势物种在空间上的变化; (2) 1 ha小样地α多样性, 包括物种个体数、物种丰富度、Shannon-Wiener指数及Pielou均匀度指数; (3) 1 ha小样地内β多样性, 表征1 ha小样地内物种的替代速率; (4)任意两个相邻样地的β多样性, 分析样地间物种替代速率的空间分布.采用的多样性测度指标如下(钱迎倩和马克平, 1994 ): ...
1
1994
... 将24 ha样地划分成24个1 ha小样地, 分别计算: (1) 1 ha小样地所有物种的重要值, 分析群落优势物种在空间上的变化; (2) 1 ha小样地α多样性, 包括物种个体数、物种丰富度、Shannon-Wiener指数及Pielou均匀度指数; (3) 1 ha小样地内β多样性, 表征1 ha小样地内物种的替代速率; (4)任意两个相邻样地的β多样性, 分析样地间物种替代速率的空间分布.采用的多样性测度指标如下(钱迎倩和马克平, 1994 ): ...
中国常绿阔叶林研究的回顾与展望
1
2005
... 亚热带常绿阔叶林在我国分布面积最广, 类型最为复杂多样, 是全球常绿阔叶林的主体(宋永昌, 2005 ).我国的中亚热带是常绿阔叶林的典型分布区(吴征镒,1980 ), 中亚热带常绿阔叶林有着丰富的生物多样性资源和巨大的生态效益, 越来越引起人们广泛的重视. ...
中国常绿阔叶林研究的回顾与展望
1
2005
... 亚热带常绿阔叶林在我国分布面积最广, 类型最为复杂多样, 是全球常绿阔叶林的主体(宋永昌, 2005 ).我国的中亚热带是常绿阔叶林的典型分布区(吴征镒,1980 ), 中亚热带常绿阔叶林有着丰富的生物多样性资源和巨大的生态效益, 越来越引起人们广泛的重视. ...
负密度制约和生境过滤对古田山幼苗功能多样性年际变化的影响
1
2017
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
负密度制约和生境过滤对古田山幼苗功能多样性年际变化的影响
1
2017
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
1
1988
... 古田山四季分明, 降水充沛, 年均温度15.3℃, 最高气温为38.1℃ (7月), 最低气温为-6.8℃ (1月), 年均降雨量1,963.7 mm, 每年的3-6月为第一雨季, 7-8月为伏旱期, 8月底至9月受台风影响形成第二雨季, 10月到翌年2月为干旱期(开化林业志编写组, 1988 ).该区域地形复杂, 山势陡峭, 山体呈东北-西南走向, 最高海拔1,258 m.该区域物种多样性丰富, 自然分布的种子植物有149科648属1,426种(包括种下分类等级), 分别占浙江省种子植物科、属、种总数的81.9%, 51.7%和41.6%, 其中国家二、三级重点保护植物18种, 并且还有一些珍稀群落, 如分布集中且面积较大的香果树(Emmenopterys benriyi )、野含笑(Michelia skinneriana )和紫茎(Stewartia sinensis )等植物群落(楼炉焕和金水虎, 2000 ). ...
1
1988
... 古田山四季分明, 降水充沛, 年均温度15.3℃, 最高气温为38.1℃ (7月), 最低气温为-6.8℃ (1月), 年均降雨量1,963.7 mm, 每年的3-6月为第一雨季, 7-8月为伏旱期, 8月底至9月受台风影响形成第二雨季, 10月到翌年2月为干旱期(开化林业志编写组, 1988 ).该区域地形复杂, 山势陡峭, 山体呈东北-西南走向, 最高海拔1,258 m.该区域物种多样性丰富, 自然分布的种子植物有149科648属1,426种(包括种下分类等级), 分别占浙江省种子植物科、属、种总数的81.9%, 51.7%和41.6%, 其中国家二、三级重点保护植物18种, 并且还有一些珍稀群落, 如分布集中且面积较大的香果树(Emmenopterys benriyi )、野含笑(Michelia skinneriana )和紫茎(Stewartia sinensis )等植物群落(楼炉焕和金水虎, 2000 ). ...
Plant diversity in tropical forests: A review of mechanisms of species coexistence
2
2002
... 本研究揭示了物种多样性存在很明显的空间变异特征, 多样性测度指标在空间上的变化特征有很大差别, 这反映了森林群落组成结构在空间上有很大的变异性.传统群落生态学研究中, 通常将群落定义为特定空间或特定生境不同种群的集合(Odum & Barrette, 2009 ).除了生境空间变化, 其他生态学过程也可以使群落的空间特征发生变化, 如扩散、竞争、干扰等(Wright, 2002 ), 从而导致物种组成和群落结构的空间变化.本研究表明相邻小样地之间物种替代速率空间变异较大, 虽然各小样地的第一、二优势种基本上都是甜槠和木荷, 十分相似, 但第三优势种差别较大, 24个1 ha小样地有多达10个不同的优势物种.这意味着中亚热带常绿阔叶林群落即使在面积为1 ha时(远大于传统的亚热带常绿阔叶林最小群落面积), 继续增大面积还会增加很多物种, 或者说群落的组成和结构会随面积(尺度)的增大而改变.群落的生态学过程具有尺度依赖性并决定群落的结构和组成(Levin, 1992 ), 因此, 对群落生态学而言, 在一定的空间尺度比较群落的组成和结构,对正确认识不同群落的差异, 研究推断群落变化的生态学过程, 制定科学的保育措施有重要意义. ...
... 物种多样性的空间格局也反映了生态学过程.古田山24个1 ha小样地木本植物的个体数量与物种丰富度没有显著的相关性, 与植物个体数越多物种数量越多的假说不同(Evans et al, 2005 ), 表明植物的个体数量并不一定和物种数量的增加相同步, 这可能与本研究小样地(1 ha)的尺度较大有关.当取样尺度较小时, 生境异质性相对较弱, 取样的植物个体数量越多, 发现不同物种数量的概率就越大(Evans et al, 2005 ); 但当取样尺度增加, 不同的生境类型会增加, 物种数量的增加将主要由生境异质性或扩散限制起作用(Harms et al, 2001 ; Wright, 2002 ).个体数量和Shannon-Wiener指数及物种均匀度呈显著的负相关, 且物种均匀度和Shannon-Wiener指数高的样地都位于低海拔沟谷地带, 这可能是由于在低海拔沟谷生境中群落内各物种的种内竞争较强, 导致种群数量下降, 均匀度和多样性增加. ...
2
1980
... 亚热带常绿阔叶林在我国分布面积最广, 类型最为复杂多样, 是全球常绿阔叶林的主体(宋永昌, 2005 ).我国的中亚热带是常绿阔叶林的典型分布区(吴征镒,1980 ), 中亚热带常绿阔叶林有着丰富的生物多样性资源和巨大的生态效益, 越来越引起人们广泛的重视. ...
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
2
1980
... 亚热带常绿阔叶林在我国分布面积最广, 类型最为复杂多样, 是全球常绿阔叶林的主体(宋永昌, 2005 ).我国的中亚热带是常绿阔叶林的典型分布区(吴征镒,1980 ), 中亚热带常绿阔叶林有着丰富的生物多样性资源和巨大的生态效益, 越来越引起人们广泛的重视. ...
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
浙江古田山自然保护区森林植被类型
1
2001
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
浙江古田山自然保护区森林植被类型
1
2001
... 古田山国家级自然保护区(29°10°19.4- 29°17°41.4 N, 118°03°49.7-118°11°12.2 E)位于浙江省开化县西部的苏庄镇境内, 总面积8,107 ha, 属于中亚热带常绿阔叶林地带(吴征镒, 1980 ), 也是国际生物多样性保护的热点地区(Huang et al, 2012 ), 是钱江源国家公园试点区的核心部分.长期以来, 亚热带地区由于严重的人为干扰, 大部分常绿阔叶林原始植被已经被严重破坏, 退化为荒地、人工林或次生林.但是, 古田山保护区仍保留着大面积未受人为干扰的典型的低海拔亚热带常绿阔叶林(350-800 m), 这在中亚热带东部地区十分少见(于明坚等, 2001 ; 胡正华等, 2003 ). ...
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
3
2008
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
... 古田山24 ha样地位于古田山保护区内, 东西长600 m, 南北长400 m, 样地内地形复杂多变, 地势较陡, 坡度范围13°-62°, 最高海拔为714.9 m, 最低海拔为446.3 m, 平均海拔为580.6 m, 最大高差为268.6 m, 植被为典型中亚热带低海拔常绿阔叶原始林(祝燕等, 2008 ).2005年按照CTFS (Centre for Tropical Forest Science)样地的建设方法(Condit, 1998 ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 ).本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种. ...
... ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 ).本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种. ...
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
3
2008
... 许多学者已经对古田山的植被和生物多样性进行了研究(陈建华和冯志坚, 2002 ; 胡正华等, 2002 , 2005 ), 特别是自2005年古田山24 ha长期森林动态监测样地建立以来, 国内外学者对古田山常绿阔叶林进行了广泛而深入的研究(祝燕等, 2008 ; Legendre et al, 2009 ; 赖江山等, 2010 ; 宋云峰等, 2017 ).虽然对古田山常绿阔叶林的群落组成及结构的维持机制已有一定的认识, 但是目前还缺乏对其生物多样性空间变异特征的认识.本研究以古田山24 ha大样地数据为基础,尝试揭示古田山常绿阔叶林群落物种多样性的空间变化特征. ...
... 古田山24 ha样地位于古田山保护区内, 东西长600 m, 南北长400 m, 样地内地形复杂多变, 地势较陡, 坡度范围13°-62°, 最高海拔为714.9 m, 最低海拔为446.3 m, 平均海拔为580.6 m, 最大高差为268.6 m, 植被为典型中亚热带低海拔常绿阔叶原始林(祝燕等, 2008 ).2005年按照CTFS (Centre for Tropical Forest Science)样地的建设方法(Condit, 1998 ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 ).本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种. ...
... ), 用全站仪将整个样地进行标定, 挂牌标记样地内所有胸径(DBH) ≥ 1 cm的木本植物个体, 测量其胸径、坐标并鉴定到种(祝燕等, 2008 ).本研究使用2005年首次完成的调查数据进行分析, 共140,700株个体, 隶属于49科104属159个物种. ...
 , 米湘成
, 米湘成

{kind=link}
{kind=link}
{kind=link}
{kind=link}