Distribution pattern and mechanism of insect species diversity in Inner Mongolia
Yu Zhang, Gang Feng*,
Ministry of Education Key Laboratory of Ecology and Resource Use of the Mongolian Plateau & Inner Mongolia Key Laboratory of Grassland Ecology, School of Ecology and Environment, Inner Mongolia University, Hohhot 010021
How species are distributed geographically and what drives these distributions remain core issues in macroecology and biogeography. Both regional and local scale factors such as temperature, precipitation, altitudinal range, habitat filtering, predation, competition and reciprocity affect the large-scale distribution pattern of insect species diversity. However, few studies have simultaneously discussed the effects of these multi-scale drivers on the geographical distribution of insect diversity. Using insect diversity data from 86 counties in Inner Mongolia Autonomous Region, we assessed the distribution patterns and main drivers of insect species diversity. We included mean annual temperature, mean annual precipitation, paleoclimate change, altitudinal range and plant diversity as predictors. Plant diversity and altitudinal range influenced insect diversity in Inner Mongolia while climatic factors had less influence. Our results show that interspecific relationships (food diversity) and habitat heterogeneity may play a crucial role in shaping distribution patterns of insect diversity in Inner Mongolia.
Keywords:insect species diversity
;
habitat heterogeneity
;
interspecific interactions
;
climatic factor
;
altitudinal range
;
plant diversity
YuZhang, GangFeng. Distribution pattern and mechanism of insect species diversity in Inner Mongolia[J]. Biodiversity Science, 2018, 26(7): 701-706 https://doi.org/10.17520/biods.2018144
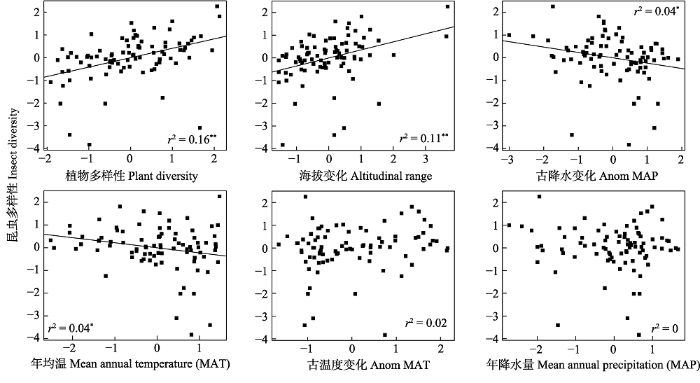
图1 昆虫多样性及其相关变量散点图。古降水变化与古温度变化是末次最大冰期到现代的降水和温度的变化。* P < 0.05, ** P < 0.01。
Fig. 1 Scatter plots of insect species diversity and the related variables. Anom MAP and Anom MAT are the change of MAP/MAT between LGM and contemporary time.
Table 1
表1
表1 最小二乘法(OLS)和空间自回归(SAR)模型单变量分析结果
Table 1 Results of single-variable analysis by ordinary least squares (OLS) and simultaneous autoregressive (SAR) models
coefOLS
r2OLS
coefSAR
r2SAR
年平均气温 Mean annual temperature (MAT)
-0.22
0.04*
-0.24
0.08**
年降水量 Mean annual precipitation (MAP)
-0.06
0
-0.05
0
古温度变化 Anom MAT
0.17
0.02
0.2
0.06**
古降水变化 Anom MAP
-0.24
0.04*
-0.23
0.07*
植物多样性 Plant diversity
0.35
0.16**
0.36
0.13**
海拔变化 Altitudinal range
0.42
0.11**
0.41
0.18**
Anom MAT and Anom MAP are the change of MAT/MAP between LGM and contemporary time. * P < 0.05, ** P < 0.01.
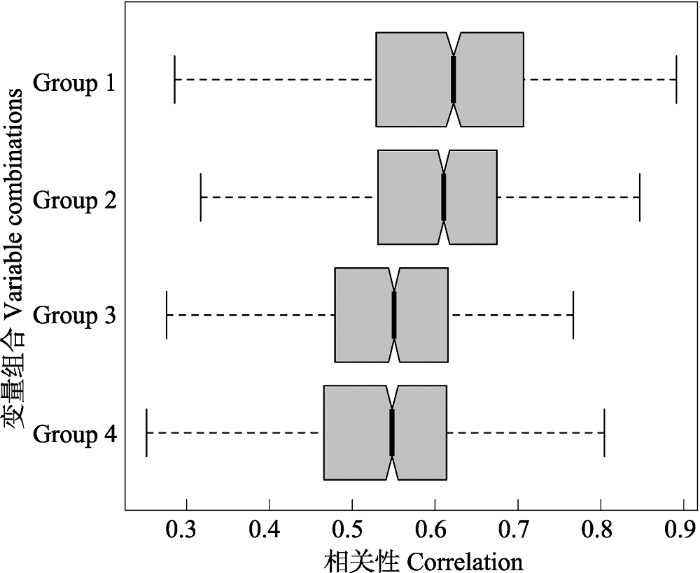
图2 随机森林模型得到的与昆虫物种多样性相关性最高的4个组合。Group 1为Anom MAP + Plant diversity, Group 2为Anom MAP + Altitudinal range + Plant diversity, Group 3为MAT + Anom MAP + Altitudinal range + Plant diversity, Group 4为MAP + Anom MAP + Altitudinal range + Plant diversity。Anom MAP表示古降水变化。
Fig. 2 Four combinations of variables most associated with insect species diversity obtained from the random forest model. Group 1 are Anom MAP and Plant diversity, Group 2 are Anom MAP, Altitudinal range and Plant diversity, Group 3 are MAT, Anom MAP, Altitudinal range and Plant diversity, Group 4 are MAP, Anom MAP, Altitudinal range and Plant diversity. Anom MAP is the change of MAP between LGM and contemporary time.
Historical refugia have shaped biogeographical patterns of species richness and phylogenetic diversity in mite harvestmen (Arachnida, Opiliones, Cyphophthalmi) endemic to the Australian Wet Tropics
Agricultural management, vegetation traits and landscape drive orthopteran and butterfly diversity in a grassland-forest mosaic: A multi-scale approach
Spatial temporal distribution of the Hymenoptera in the Brazilian Savanna and the effects of habitat heterogeneity on these patterns
1
2015
... 生境异质性假说同样是生态学研究中的一个基本问题.较高的生境异质性可以为生物提供更多的生态位、避难所及物种形成的机会, 从而进一步促进较高的物种多样性的产生(Stein et al, 2014).Aranda和Graciolli (2015)对巴西热带稀树草原的研究发现, 生境异质性即植被结构影响膜翅目昆虫群落组成.Marini等(2010)对意大利阿尔卑斯山区域的研究发现, 木本植被的比例影响直翅目及蝴蝶的多样性. ...
The geographic scaling of biotic interactions
1
2014
... 生物之间的相互作用是影响生物多样性地理分布格局的重要因子之一(Wisz et al, 2013; Araújo & Rozenfeld, 2014).植物多样性与昆虫多样性的关系是生态学研究中的基本问题之一(朱慧等, 2008; Zhang et al, 2016).较高的植物多样性可以为昆虫提供更多、更优质的食物, 从而直接影响昆虫多样性; 与此同时, 较高的植物多样性可以提供更多的生态位空间、影响局域生境条件(土壤及空气温湿度), 从而间接地影响昆虫多样性(朱慧等, 2008; Zhang et al, 2016). ...
Arthropod diversity in a tropical forest
1
2012
... 我们的研究结果发现内蒙古昆虫多样性与植物多样性的相关性要高于与气候因素、海拔变化的相关性, 表明植物多样性对昆虫多样性分布格局有重要的影响.这与前人的多数研究结果一致, 比如, 美国爱达荷州草原的植物多样性与昆虫多样性呈正相关关系(Wenninger & Inouye, 2008).在中国热带地区的云南西双版纳布龙自然保护区与海南鹦哥岭自然保护区, 根据植物多样性可以准确地预测昆虫多样性(Zhang et al, 2016).在巴拿马热带雨林地区, 植物多样性同样可以较好地估计昆虫多样性的分布(Basset et al, 2012).已有的研究还发现内蒙古植物多样性能够较好地解释其鸟类多样性的分布格局(Liang et al, 2018).结合已有研究, 我们的结果进一步证明生物之间相互作用或生境多样性可能对昆虫多样性的地理分布格局有重要作用. ...
Historical refugia have shaped biogeographical patterns of species richness and phylogenetic diversity in mite harvestmen (Arachnida, Opiliones, Cyphophthalmi) endemic to the Australian Wet Tropics
1
2016
... 气候因子, 包括当代气候及历史气候变化, 是影响大尺度生物多样性分布格局的重要因素(Fine, 2015; Svenning et al, 2015).我们的研究发现古降水变化显著影响内蒙古昆虫多样性分布格局, 尤其是随机森林分析结果显示古降水变化同植物多样性一样, 出现在4个相关性最高的模型组合.并且古降水变化越小的区域具有越高的昆虫多样性.这个发现也与前人的研究相似, 比如, 澳大利亚热带地区的历史冰期避难所拥有较高的昆虫物种多样性与系统发育多样性(Boyer et al, 2016).我们的结果进一步证明了冰期避难所对生物多样性的保护作用. ...
... 物种多样性的地理分布格局及其机制是生态学及生物地理学的核心问题之一(Currie, 1991; Brown, 2014; Fine, 2015).其中, 区域尺度的物种形成、灭绝及迁徙, 与局域水平的生境过滤、种间关系及扩散限制共同影响物种多样性的地理分布格局(Fine, 2015; Qu et al, 2015; Svenning et al, 2015).已有研究发现中国亚热带森林中树木多样性显著促进昆虫多样性(Schuldt et al, 2010).中国蚂蚁丰富度与最冷月温度、年降水量和年温度变化显著相关(沈梦伟等, 2016).然而, 较少有研究同时比较不同尺度影响因素对大尺度昆虫多样性分布格局的影响. ...
Energy and large scale patterns of animal and plant species richness
1
1991
... 物种多样性的地理分布格局及其机制是生态学及生物地理学的核心问题之一(Currie, 1991; Brown, 2014; Fine, 2015).其中, 区域尺度的物种形成、灭绝及迁徙, 与局域水平的生境过滤、种间关系及扩散限制共同影响物种多样性的地理分布格局(Fine, 2015; Qu et al, 2015; Svenning et al, 2015).已有研究发现中国亚热带森林中树木多样性显著促进昆虫多样性(Schuldt et al, 2010).中国蚂蚁丰富度与最冷月温度、年降水量和年温度变化显著相关(沈梦伟等, 2016).然而, 较少有研究同时比较不同尺度影响因素对大尺度昆虫多样性分布格局的影响. ...
Defying the curse of ignorance: Perspectives in insect macroecology and conservation biogeography
3
2010
... 昆虫是地球上较早出现的动物类群之一, 距今约有4亿8千万年(Misof et al, 2014).作为地球上多样性最为丰富的多细胞生物类群, 迄今已经发现100多万种, 并且仍有更多未描述的物种(Diniz-Filho et al, 2010).与此同时, 昆虫的生物多样性保护对于陆地生态系统多样性维持的生态学及进化学过程有重要作用(Thomas et al, 2008; Diniz-Filho et al, 2010).然而, 由于分类信息及地理分布信息的缺乏, 昆虫生物地理学分布格局的相关研究一直以来较为匮乏(Diniz-Filho et al, 2010). ...
... ; Diniz-Filho et al, 2010).然而, 由于分类信息及地理分布信息的缺乏, 昆虫生物地理学分布格局的相关研究一直以来较为匮乏(Diniz-Filho et al, 2010). ...
... ).然而, 由于分类信息及地理分布信息的缺乏, 昆虫生物地理学分布格局的相关研究一直以来较为匮乏(Diniz-Filho et al, 2010). ...
Ecological and evolutionary drivers of geographic variation in species diversity
4
2015
... 物种多样性的地理分布格局及其机制是生态学及生物地理学的核心问题之一(Currie, 1991; Brown, 2014; Fine, 2015).其中, 区域尺度的物种形成、灭绝及迁徙, 与局域水平的生境过滤、种间关系及扩散限制共同影响物种多样性的地理分布格局(Fine, 2015; Qu et al, 2015; Svenning et al, 2015).已有研究发现中国亚热带森林中树木多样性显著促进昆虫多样性(Schuldt et al, 2010).中国蚂蚁丰富度与最冷月温度、年降水量和年温度变化显著相关(沈梦伟等, 2016).然而, 较少有研究同时比较不同尺度影响因素对大尺度昆虫多样性分布格局的影响. ...
... ).其中, 区域尺度的物种形成、灭绝及迁徙, 与局域水平的生境过滤、种间关系及扩散限制共同影响物种多样性的地理分布格局(Fine, 2015; Qu et al, 2015; Svenning et al, 2015).已有研究发现中国亚热带森林中树木多样性显著促进昆虫多样性(Schuldt et al, 2010).中国蚂蚁丰富度与最冷月温度、年降水量和年温度变化显著相关(沈梦伟等, 2016).然而, 较少有研究同时比较不同尺度影响因素对大尺度昆虫多样性分布格局的影响. ...
Very high resolution interpolated climate surfaces for global land areas
1
2005
... 本研究选择的昆虫物种多样性影响因子有现代气候(年均温及年降水)、古气候变化(末次最大冰期到现在的温度变化及降水变化)、海拔变化与植物多样性.从WorldClim收集了海拔变化信息及气候变量, 即年平均温度(mean annual temperature, MAT)、年平均降水量(mean annual precipitation, MAP)、末次最大冰期温度(MATLGM)、末次最大冰期降水量(MAPLGM) (Hijmans et al, 2005).以海拔变化梯度作为环境异质性的代表(Stein et al, 2014).MATLGM和MAPLGM来自于CCSM3 (Otto-Bliesner et al, 2006)和MIROC3.2 (Hasumi & Emori, 2004)两个模型的平均值, 然后用现代气候变量值减去末次最大冰期气候变量值来表示温度及降水的变化(Sandel et al, 2011).气候数据及海拔数据的精度为0.0417°, 其中MAT和MAP是1960-1990年的平均值.计算了每个旗县的平均值来代表该旗县的气候变量值, 同时计算了每个旗县的海拔变化范围. ...
Bird species richness is associated with phylogenetic relatedness, plant species richness, and altitudinal range in Inner Mongolia
2
2018
... 我们的研究结果发现内蒙古昆虫多样性与植物多样性的相关性要高于与气候因素、海拔变化的相关性, 表明植物多样性对昆虫多样性分布格局有重要的影响.这与前人的多数研究结果一致, 比如, 美国爱达荷州草原的植物多样性与昆虫多样性呈正相关关系(Wenninger & Inouye, 2008).在中国热带地区的云南西双版纳布龙自然保护区与海南鹦哥岭自然保护区, 根据植物多样性可以准确地预测昆虫多样性(Zhang et al, 2016).在巴拿马热带雨林地区, 植物多样性同样可以较好地估计昆虫多样性的分布(Basset et al, 2012).已有的研究还发现内蒙古植物多样性能够较好地解释其鸟类多样性的分布格局(Liang et al, 2018).结合已有研究, 我们的结果进一步证明生物之间相互作用或生境多样性可能对昆虫多样性的地理分布格局有重要作用. ...
... 作为生境异质性的一个重要又易于测量的指标, 海拔变化被广泛用于研究生境异质性与多样性分布格局的关系(Tews et al, 2004).南非蝗虫的多样性分布显著地受到地形异质性的影响(Gebeyehu & Samways, 2006).坡度变化与意大利阿尔卑斯山区域的昆虫与植物多样性显著正相关(Marini et al, 2009).海拔变化同样显著影响内蒙古鸟类多样性的分布格局(Liang et al, 2018).与前人的研究相似, 我们的结果表明海拔变化能够较好地预测内蒙古昆虫多样性分布格局, 从而验证了生境异质性假说对内蒙古昆虫多样性地理分布格局的影响. ...
Agricultural management, vegetation traits and landscape drive orthopteran and butterfly diversity in a grassland-forest mosaic: A multi-scale approach
1
2010
... 生境异质性假说同样是生态学研究中的一个基本问题.较高的生境异质性可以为生物提供更多的生态位、避难所及物种形成的机会, 从而进一步促进较高的物种多样性的产生(Stein et al, 2014).Aranda和Graciolli (2015)对巴西热带稀树草原的研究发现, 生境异质性即植被结构影响膜翅目昆虫群落组成.Marini等(2010)对意大利阿尔卑斯山区域的研究发现, 木本植被的比例影响直翅目及蝴蝶的多样性. ...
Impact of farm size and topography on plant and insect diversity of managed grasslands in the Alps
1
2009
... 作为生境异质性的一个重要又易于测量的指标, 海拔变化被广泛用于研究生境异质性与多样性分布格局的关系(Tews et al, 2004).南非蝗虫的多样性分布显著地受到地形异质性的影响(Gebeyehu & Samways, 2006).坡度变化与意大利阿尔卑斯山区域的昆虫与植物多样性显著正相关(Marini et al, 2009).海拔变化同样显著影响内蒙古鸟类多样性的分布格局(Liang et al, 2018).与前人的研究相似, 我们的结果表明海拔变化能够较好地预测内蒙古昆虫多样性分布格局, 从而验证了生境异质性假说对内蒙古昆虫多样性地理分布格局的影响. ...
Phylogenomics resolves the timing and pattern of insect evolution
1
2014
... 昆虫是地球上较早出现的动物类群之一, 距今约有4亿8千万年(Misof et al, 2014).作为地球上多样性最为丰富的多细胞生物类群, 迄今已经发现100多万种, 并且仍有更多未描述的物种(Diniz-Filho et al, 2010).与此同时, 昆虫的生物多样性保护对于陆地生态系统多样性维持的生态学及进化学过程有重要作用(Thomas et al, 2008; Diniz-Filho et al, 2010).然而, 由于分类信息及地理分布信息的缺乏, 昆虫生物地理学分布格局的相关研究一直以来较为匮乏(Diniz-Filho et al, 2010). ...
... 气候因子, 包括当代气候及历史气候变化, 是影响大尺度生物多样性分布格局的重要因素(Fine, 2015; Svenning et al, 2015).我们的研究发现古降水变化显著影响内蒙古昆虫多样性分布格局, 尤其是随机森林分析结果显示古降水变化同植物多样性一样, 出现在4个相关性最高的模型组合.并且古降水变化越小的区域具有越高的昆虫多样性.这个发现也与前人的研究相似, 比如, 澳大利亚热带地区的历史冰期避难所拥有较高的昆虫物种多样性与系统发育多样性(Boyer et al, 2016).我们的结果进一步证明了冰期避难所对生物多样性的保护作用. ...
Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures
1
2004
... 作为生境异质性的一个重要又易于测量的指标, 海拔变化被广泛用于研究生境异质性与多样性分布格局的关系(Tews et al, 2004).南非蝗虫的多样性分布显著地受到地形异质性的影响(Gebeyehu & Samways, 2006).坡度变化与意大利阿尔卑斯山区域的昆虫与植物多样性显著正相关(Marini et al, 2009).海拔变化同样显著影响内蒙古鸟类多样性的分布格局(Liang et al, 2018).与前人的研究相似, 我们的结果表明海拔变化能够较好地预测内蒙古昆虫多样性分布格局, 从而验证了生境异质性假说对内蒙古昆虫多样性地理分布格局的影响. ...
Where within a geographical range do species survive best? A matter of scale
1
2008
... 昆虫是地球上较早出现的动物类群之一, 距今约有4亿8千万年(Misof et al, 2014).作为地球上多样性最为丰富的多细胞生物类群, 迄今已经发现100多万种, 并且仍有更多未描述的物种(Diniz-Filho et al, 2010).与此同时, 昆虫的生物多样性保护对于陆地生态系统多样性维持的生态学及进化学过程有重要作用(Thomas et al, 2008; Diniz-Filho et al, 2010).然而, 由于分类信息及地理分布信息的缺乏, 昆虫生物地理学分布格局的相关研究一直以来较为匮乏(Diniz-Filho et al, 2010). ...
Insect community response to plant diversity and productivity in a sagebrush steppe ecosystem
1
2008
... 我们的研究结果发现内蒙古昆虫多样性与植物多样性的相关性要高于与气候因素、海拔变化的相关性, 表明植物多样性对昆虫多样性分布格局有重要的影响.这与前人的多数研究结果一致, 比如, 美国爱达荷州草原的植物多样性与昆虫多样性呈正相关关系(Wenninger & Inouye, 2008).在中国热带地区的云南西双版纳布龙自然保护区与海南鹦哥岭自然保护区, 根据植物多样性可以准确地预测昆虫多样性(Zhang et al, 2016).在巴拿马热带雨林地区, 植物多样性同样可以较好地估计昆虫多样性的分布(Basset et al, 2012).已有的研究还发现内蒙古植物多样性能够较好地解释其鸟类多样性的分布格局(Liang et al, 2018).结合已有研究, 我们的结果进一步证明生物之间相互作用或生境多样性可能对昆虫多样性的地理分布格局有重要作用. ...
The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling
1
2013
... 生物之间的相互作用是影响生物多样性地理分布格局的重要因子之一(Wisz et al, 2013; Araújo & Rozenfeld, 2014).植物多样性与昆虫多样性的关系是生态学研究中的基本问题之一(朱慧等, 2008; Zhang et al, 2016).较高的植物多样性可以为昆虫提供更多、更优质的食物, 从而直接影响昆虫多样性; 与此同时, 较高的植物多样性可以提供更多的生态位空间、影响局域生境条件(土壤及空气温湿度), 从而间接地影响昆虫多样性(朱慧等, 2008; Zhang et al, 2016). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}