植物的驯化与利用催生了农耕文明。农耕文明的起源与发展, 促进了工具的运用与不断改良, 转变了人类的生活方式, 促使人类由迁徙转变为定居, 出现聚落、城邦及相应的社会组织结构, 为人类文明的发展奠定了基础。中华文明(或华夏文明)是世界上唯一一直传承的古老文明。考古证据显示, 长江流域(总体位于24-35° N之间)有着和黄河流域一样辉煌的史前文明, 两者都是中华文明之源; 长江文明以稻作文明为内核, 黄河文明则以粟作文明为其鲜明特征(Barton et al, 2009 ; Zhao, 2011 )。
水稻(即亚洲栽培稻Oryza sativa )是世界上最重要的粮食作物之一, 目前全球有超过半数以上人口以稻米为食。水稻起源于野生稻。先人从对野生稻的原始照看到有意识的栽培管理, 再到有目的的选育, 最终驯化出适应栽培环境的栽培稻, 并广泛栽培而形成稻作农业, 经过了长达数千年的发展历程(赵志军, 2009 )。在此过程中, 一方面, 野生稻原有的一些性状经过人工选择, 逐渐发生适应于栽培环境的改变, 形成驯化性状(domesticated traits), 最终演变为栽培稻; 另一方面, 因适应水稻栽培管理的需要, 人们不断改进生产工具, 变革生产方式, 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业, 逐步确立了长江流域以稻作为主要生产方式的农耕文明。
水稻驯化及稻作农业为长江文明的发展和繁荣提供了重要的物质和文化基础, 也是长江文明为人类做出的巨大的开创性贡献(Larson et al, 2014 )。关于水稻是何时、何地、在什么环境下开始驯化等问题一直是国际学术界关注的热点。得益于新技术手段的不断运用, 越来越多的考古学和遗传学研究结果显示, 中国最早开始利用水稻, 长江流域及以南地区是水稻的驯化起源地(Cross & Zhao, 2014 )。本文简要评述了近年来有关水稻起源的考古学和遗传学研究进展, 特别关注长江流域的相关研究, 分析了各种研究的结论不一致的可能原因, 并讨论了未来值得深入研究的科学问题。
1 水稻关键驯化性状及相关基因
稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b )。我们通常所称的水稻是指亚洲栽培稻。与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 )。普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a )。这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议。目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 )。
因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 )。粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区。水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离。
理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地。水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 )。然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异。野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求。普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 )。谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 )。普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 )。新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 )。随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 )。此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 )。因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 )。
在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 )。从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻。落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 )。
不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 )。普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 )。因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 )。小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 )。落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 )。其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 )。sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 )。sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 )。分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 )。qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 )。
粒形是另一个重要驯化性状。古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择。水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 )。一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 )。粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 )。水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 )。水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 )。
图1 普通野生稻、籼稻、粳稻的谷粒及种子形态。a: 籼稻9311; b: 粳稻日本晴(Nipponbare); c: 普通野生稻(来自江西东乡)
Fig. 1 Grain and seed of Oryza rufipogon , O. sativa ssp. indica and japonica . a, indica variety 9311; b, japonica variety Nipponbare; c, O. rufipogon from Dongxiang population of Jiangxi Province.
水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 )。普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型。目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 )。该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 )。另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 )。
普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 )。普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 )。通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 )。rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 )。
2 水稻起源的分子生物学证据
分子检测技术的进步极大推动了栽培稻驯化起源研究。中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论)。较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 )。类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源。随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群。进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区。粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持。然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点。Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域。Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻。通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持。但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 )。Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区。最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来。
理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧。关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 )。但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源。Singh等(2017) 持类似观点。Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model)。
纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用。值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 )。比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 )。这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰。稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判。充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果。
3 水稻起源于长江流域的考古证据
据Silva等(2015) 统计, 世界各地已发现的涉及亚洲栽培稻的水稻遗址共有470多处, 其中约1/3分布在中国。Gong等(2007) 认为中国共有280处水稻文物遗址。公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处。通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 )。这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程。
图2 中国水稻遗址时空分布图
Fig. 2 The temporal and spatial distribution of ancient rice relics in China
在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N)。这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 )。仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 )。植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间。不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 )。玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 )。在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶。玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 )。
接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N)。上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块。这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 )。对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 )。Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系。
从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等。八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等。这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 )。最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 )。宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 )。彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕。张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化。同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 )。以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 )。
随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等。跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等。该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 )。跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升。到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合。河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 )。田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具。Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植。此外, 田螺山还发现了距今约7,000-4,500年的稻田。以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 )。
到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域。这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景。这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻。城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名。该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸。稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 )。这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 )。
到了新石器时代后期(5,000-4,000 BP), 长江流域的水稻考古遗址数量剧增, 反映出稻作农业生产体系的发展与传播与当时人口大幅度增长的密切关系。例如, 在长江下游的杭州湾地区和太湖流域, 距今5,200-4,300年间的良渚文化时期的遗址的分布异常密集(图2 )。同样, 在长江中游地区也发现了大量距今5,300-4,600年间的屈家岭(113.12° E, 31.02° N)文化时期和距今4,600-4,000年间的石家河(113.09° E, 30.76° N)文化时期的稻作遗址群。这些遗址出土的文物和动植物遗存充分说明, 稻作农业已经取代采集狩猎成为当地经济的主体。同一时期, 在邻近长江流域的淮河流域、黄河流域甚至更北地区也涌现出大量的水稻遗址(图2 ), 反映了水稻自驯化起源之地——长江流域向外辐射与传播, 标志着稻作文明的快速发展。
4,000 BP以后, 古稻遗址以长江中下游为中心, 向北、向西、向南辐射, 展示了稻作文明繁荣与传播的图景。这些遗址古稻(包括来自长江流域的)基本上属于粳型, 而且与现代粳稻类型基本一致(公婷婷, 2017 )。
4 稻作的起源与发展催生了长江文明
遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域)。同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 )。长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型)。伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 )。这个发展过程体现在诸多方面。首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级。从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 )。其次, 生活方式变迁。居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 )。第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 )。第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 )。制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物。因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心。
5 研究展望
尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议。第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 )。第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 )。未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化。
当前, 长江流域的野生稻分布点很少, 但大量的古稻遗址位于长江流域及以北地区。这种格局暗示, 历史气候变化和长期的生境利用引起野生稻分布区向南收缩。当前, 长江流域是人类活动强度最大的地区之一。这种高强度的人类活动如何影响以野生稻为代表的重要资源植物的进化过程, 依然知之甚少。进行古气候、古植被、土壤地层学和考古学等资料的整合分析, 将有助于探讨此问题, 其结果对于植物遗传资源的保护也有启示意义。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Asano K Yamasaki M Takuno S Miura K Katagiri S Ito T Doi K , Wu JZ, Ebana K, Matsumoto T, Innan H, Kitano H, Ashikari M, Matsuoka M (2011 ) Artificial selection for a green revolution gene during japonica rice domestication
. Proceedings of the National Academy of Sciences, USA , 108 , 11034 -11039 .
[本文引用: 1]
[2]
Barton L Newsome SD Che FH Wang H Guilderson TP Bettinger RL 2009 ) Agricultural origins and the isotopic identity of domestication in northern China
. Agricultural origins and the isotopic identity of domestication in northern China, Proceedings of the National Academy of Sciences , 106 , 5523 -5528 .
[本文引用: 1]
[3]
Caicedo AL Williamson SH Hernandez RD Boyko A Fledel-Alon A York TL Polato NR Olsen KM Nielsen R McCouch SR 2007 ) Genome-wide patterns of nucleotide polymorphism in domesticated rice
. PLoS Genetics , 3 , 1745 -1756 .
[本文引用: 2]
[4]
Castillo CC Tanaka K Sato YI Ishikawa R Bellina B Higham C Chang N Mohanty R Kajale M Fuller DQ 2016 ) Archaeogenetic study of prehistoric rice remains from Thailand and India: Evidence of early japonica in South and Southeast Asia
. Archaeological and Anthropological Science , 8 , 523 -543 .
[本文引用: 2]
[5]
Choi JY Platts AE Fuller DQ Hsing YL Wing RA Purugganan MD 2017 ) The rice paradox: Multiple origins but single domestication in Asian rice
. Molecular Biology and Evolution . 34 , 969 -979 .
[本文引用: 5]
[6]
Choi JY Purugganan MD 2018 ) Multiple origin but single domestication led to Oryza sativa. G3-Genes Genomes Genetics
, 8 , 797 -803 .
[本文引用: 2]
[7]
Civáň P Brown TA 2017 ) Origin of rice (Oryza sativa L.) domestication genes
. Genetic Resources and Crop Evolution , 64 , 1125 -1132 .
[本文引用: 1]
[8]
Civáň P Craig H Cox CJ Brown TA 2015 ) Three geographically separate domestications of Asian rice
. Nature Plants , 1 , 15164 .
[本文引用: 1]
[9]
Cohn MA Butera DL Hughes JA 1983 ) Seed dormancy in red rice. III. Response to nitrite, nitrate, and ammonium ions
. Plant Physiology , 73 , 381 -384 .
[本文引用: 1]
[10]
Cross BL Zhao ZJ 2014 ) Archaeological and genetic insights into the origins of domesticated rice
. Archaeological and genetic insights into the origins of domesticated rice, Proceedings of the National Academy of Sciences , 111 , 6190 -6197 .
[本文引用: 3]
[11]
Fuller DQ 2007 ) Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World
. Annals of Botany (London) , 100 , 903 -924 .
[本文引用: 3]
[12]
Fuller DQ 2011 ) Pathways to Asian civilizations: Tracing the origins and spread of rice and rice cultures
. Rice , 4 , 78 -92 .
[本文引用: 1]
[13]
Fuller DQ Qin L Harvey E 2008 ) Rice archaeobotany revisted
. Antiquity , 82 , 315 .
[本文引用: 1]
[14]
Fuller DQ Qin L Zheng YF Zhao ZJ Chen XG Hosoya LA Sun GP 2009 ) The domestication process and domestication rate in rice: Spikelet bases from the Lower Yangtze
. Science , 323 , 1607 -1610 .
[本文引用: 2]
[15]
Fuller DQ Sato YI Castillo C Qin L Weisskopf AR Kingwell-Banham EJ Song JX Ahn SM van Etten J 2010 ) Consilience of genetics and archaeobotany in the entangled history of rice
. Archaeological and Anthropological Science , 2 , 115 -131 .
[本文引用: 5]
[16]
Gao LZ Innan H 2008 ) Nonindependent domestication of the two rice subspecies, Oryza sativa ssp. indica and ssp. japonica , demonstrated by multilocus microsatellites
. Genetics , 179 , 965 -976 .
[本文引用: 1]
[17]
Garris AJ Tai TH Coburn J Kresovich S McCouch S 2005 ) Genetic structure and diversity in Oryza sativa L
. Genetics , 169 , 1631 -1638 .
[本文引用: 2]
[18]
Ge S Sang T 2011 ) In appropriate model rejects independent domestications of indica and japonica rice
. Proceedings of the National Academy of Sciences,USA , 108 , E755 .
[本文引用: 1]
[19]
Gong TT 2017 ) Origin, domestication and dispersal of Chinese Cultivated Rice . PhD dissertation, Minzu University of China , Beijing . (in Chinese with English abstract)
[本文引用: 5]
[公婷婷 (2017 ) 中国水稻的起源、驯化及传播研究 . 博士学位论文, 中央民族大学 , 北京 .]
[本文引用: 5]
[20]
Gong ZT Chen HZ Yuan DG Zhao YG Wu YJ Zhang GL 2007 ) The temporal and spatial distribution of ancient rice in China and its implications
. Chinese Science Bulletin , 52 , 1071 -1079 .
[21]
Gross BL Zhao ZJ 2014 ) Archaeological and genetic insights into the origins of domesticated rice
. Archaeological and genetic insights into the origins of domesticated rice, Proceedings of the National Academy of Sciences , 111 , 6190 -6197 .
[本文引用: 1]
[22]
Gu XY Foley ME Horvath DP Anderson JV Feng JH Zhang LH Mowry CR Ye H Suttle JC Kadowaki K Chen ZX 2011 ) Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice
. Genetics , 189 , 1515 -1524 .
[本文引用: 1]
[23]
Guo JY Guo LX 2014 ) The origin and formation of rice cultivation: Spatio-temporal parameters
. Agricultural History of China , 5 , 3 -13 . (in Chinese)
[本文引用: 2]
[郭静云 , 郭立新 (2014 ) 论稻作萌生与成熟的时空问题
. 中国农史 , 5 , 3 -13 .]
[本文引用: 2]
[24]
Hammer K 1984 ) The domestication syndrome
. Genetic Resources and Crop Evolution , 32 , 11 -34 .
[本文引用: 1]
[25]
He ZW Zhai WW Wen HJ Tang T Wang Y Lu XM Greenberg AJ Hudson RR 2011 ) Two evolutionary histories in the genome of rice: The roles of domestication genes
. PLoS Genetics , 7 , e1002100 .
[本文引用: 1]
[26]
Hu DK Clift PD Böning P Hannigan R Hillier S Blusztajn J Wan SM Fuller DQ 2013 ) Holocene evolution in weathering and erosion patterns in the Pearl River delta
. Geochemistry Geophysics Geosystems , 14 , 2349 -2368 .
[本文引用: 1]
[27]
Huang P Molina J Flowers JM Rubinstein S Jackson SA Purugganan MD Schaal BA 2012 a) Phylogeography of Asian wild rice, Oryza rufipogon : A genome-wide view
. Molecular Ecology , 21 , 4593 -4604 .
[本文引用: 1]
[28]
Huang P Schaal BA 2012 ) Association between the geographic distribution during the last glacial maximum of Asian wild rice, Oryza rufipogon (Poaceae), and its current genetic variation
. American Journal of Botany , 99 , 1866 -1874 .
[本文引用: 4]
[29]
Huang XH Kurata N Wei X Wang ZX Wang A Zhao Q Zhao Y Liu KY Lu HY Li WJ 2012b ) A map of rice genome variation reveals the origin of cultivated rice
. Nature , 490 , 497 -501 .
[30]
Jiang LP Liu L 2006 ) New evidence for the origins of sedentism and rice domestication in the Lower Yangtze River, China
. Antiquity , 80 , 355 -361 .
[本文引用: 1]
[31]
Jin J Huang W Gao JP Yang J Shi M Zhu MZ Luo D Lin HX 2008 ) Genetic control of rice plant architecture under domestication
. Nature Genetics , 40 , 1365 -1369 .
[本文引用: 2]
[32]
Jones MK Liu XY 2009 ) Archaeology: Origins of agriculture in East Asia
. Science , 324 , 730 -731 .
[本文引用: 2]
[33]
Khush GS 1997 ) Origin, dispersal, cultivation and variation of rice
. Plant Molecular Biology , 35 , 25 -34 .
[本文引用: 1]
[34]
Konishi S Izawa T Lin SY Ebana K Fukuta Y Sasaki T Yano M 2006 ) An SNP caused loss of seed shattering during rice domestication
. Science , 312 , 1392 -1396 .
[本文引用: 2]
[35]
Kovach MJ Sweeney MT McCouch SR 2007 ) New insights into the history of rice domestication
. Trends in Genetics , 23 , 578 -587 .
[本文引用: 2]
[36]
Larson G Piperno DR Allaby RG Purugganan MD Andersson L Arroyo-Kalin M Barton L Vigueira CC Denham T Dobney K 2014 ) Current perspectives and the future of domestication studies
. Current perspectives and the future of domestication studies, Proceedings of the National Academy of Sciences , 111 , 6139 -6146 .
[本文引用: 1]
[37]
Li CB Zhou AL Sang T 2006 ) Rice domestication by reducing shattering
. Science , 311 , 1936 -1939 .
[本文引用: 3]
[38]
Lin ZW Griffith ME Li XR Zhu ZF Tan LB Fu YC Zhang WX Wang XK Xie DX Sun CQ 2007 ) Origin of seed shattering in rice (Oryza sativa L.)
. Planta , 226 , 11 -20 .
[本文引用: 1]
[39]
Liu J 2006 ) Hemudu Culture . Cultural Relics Press , Beijing (in Chinese)
[本文引用: 2]
[刘军 (2006 ) 河姆渡文化 . 文物出版社 , 北京 .]
[本文引用: 2]
[40]
Liu R Zheng XM Zhou L Zhou HF Ge S 2015 ) Population genetic structure of Oryza rufipogon and Oryza nivara : Implications for the origin of O. nivara
. Molecular Ecology , 24 , 5211 -5228 .
[本文引用: 1]
[41]
Liu T Liu Y Sun QL Zong YQ Finlayson B Chen ZY 2017 ) Early Holocene groundwater table fluctuations in relation to rice domestication in the middle Yangtze River basin, China
. Quaternary Science Reviews , 155 , 79 -85 .
[本文引用: 1]
[42]
Londo JP Chiang YC Hung KH Chiang TY Schaal BA 2006 ) Phylogeography of Asian wild rice, Oryza rufipogon , reveals multiple independent domestications of cultivated rice, Oryza sativa
. Proceedings of the National Academy of Sciences, USA , 103 , 9578 -9583 .
[本文引用: 2]
[43]
McNally KL Childs KL Bohnert R Davidson RM Zhao K Ulat VJ Zeller G Clark RM Hoen DR Bureau TE 2009 ) Genomewide SNP variation reveals relationships among landraces and modern varieties of rice
. Genomewide SNP variation reveals relationships among landraces and modern varieties of rice, Proceedings of the National Academy of Sciences , 106 , 12273 -12278 .
[本文引用: 1]
[44]
Molina J Sikora M Garud N Flowers JM Rubinstein S Reynolds A Huang P Jackson S Schaal BA Bustamante CD Boyko AR Purugganan MD 2011 ) Molecular evidence for a single evolutionary origin of domesticated rice
. Proceedings of the National Academy of Sciences,USA , 108 , 8351 -8356 .
[本文引用: 2]
[45]
Ou SJ Wang HR Chu CC 2012 ) Major domestication traits in Asian rice
. Hereditas (Beijing) , 34 , 1379 -1389 . (in Chinese with English abstract)
[本文引用: 3]
[区树俊 , 汪鸿儒 , 储成才 (2012 ) 亚洲栽培稻主要驯化性状研究进展
. 遗传 , 34 , 1379 -1389 .]
[本文引用: 3]
[46]
Paterson AH Li ZK 2011 ) Paleo-green revolution for rice
. Paleo-green revolution for rice, Proceedings of the National Academy of Sciences , 108 , 10931 -10932 .
[本文引用: 1]
[47]
Sakuma S Salomon B Komatsuda T 2011 ) The domestication syndrome genes responsible for the major changes in plant form in the Triticeae crops
. Plant Cell Physiology , 52 , 738 -749 .
[本文引用: 1]
[48]
Sang T Ge S 2007 ) Genetics and phylogenetics of rice domestication
. Current Opinion in Genetics & Development , 17 , 533 -538 .
[本文引用: 1]
[49]
Second G 1982 ) Origin of the genetic diversity of cultivated (Oryza spp.) study of the polymorphism scored at 40 isozyme loci
. Japan Journal of Genetics , 57 , 25 -37 .
[本文引用: 1]
[50]
Seshu DV Dadlani M 1991 ) Mechanism of seed dormancy in rice
. Seed Scicene Research , 1 , 187 -194 .
[本文引用: 1]
[51]
Shomura A Izawa T Ebana K Ebitani T Kanegae H Konishi S Yano M 2008 ) Deletion in a gene associated with grain size increased yields during rice domestication
. Nature Genetics , 40 , 1023 -1028 .
[本文引用: 1]
[52]
Silva F Stevens CJ Weisskopf A Castillo C Qin L Bevan A Fuller DQ 2015 ) Modelling the geographical origin of rice cultivation in Asia using the rice archaeological database
. PLoS ONE , 10 , e0137024 .
[本文引用: 2]
[53]
Singh N Singh B Rai V Sidhu S Singh AK Singh NC 2017 ) Evolutionary insights based on SNP haplotypes of red pericarp, grain size and starch synthase genes in wild and cultivated rice
. Frontiers in Plant Science , 8 , 972 .
[本文引用: 1]
[54]
Song ZP Zhu WY Rong J Xu X Chen JK Lu BR 2006 ) Evidences of introgression from cultivated rice to Oryza rufipogon (Poaceae) populations based on SSR fingerprinting: Implications for wild rice differentiation and conservation
. Evolutionary Ecology , 20 , 501 -522 .
[本文引用: 1]
[55]
Sun CQ Wang XK Yoshimura A Iwata N 1997 ) RFLP analysis of nuclear DNA in common wild rice (Oryza rufipogon Griff.) and cultivated rice (Oryza sativa L.)
. Scientia Agricultura Sinica , 30 , 37 -44 .
[本文引用: 1]
[56]
Sweeney M McCouch S 2007 ) The complex history of the domestication of rice
. Annals of Botany , 100 , 951 -957 .
[本文引用: 3]
[57]
Sweeney MT Thomson MJ Cho YG Park YJ Williamson SH Bustamante CD McCouch SR 2007 ) Global dissemination of a single mutation conferring white pericarp in rice
. PLoS Genetics , 3 , e133 .
[58]
Tan LB Li XR Liu FX Sun XY Li CG Zhu ZF Fu CY Cai HW Wang XK Xie DX Sun CQ 2008 ) Control of a key transition from prostrate to erect growth in rice domestication
. Nature Genetics , 40 , 1360 -1364 .
[本文引用: 1]
[59]
Tang T Lu J Huang JZ He JH McCouch SR Shen Y Kai Z Purugganan MD Shi SH Wu CI 2006 ) Genomic variation in rice: genesis of highly polymorphic linkage blocks during domestication
. PLoS Genetics , 2 , 1824 -1833 .
[本文引用: 1]
[60]
Vaughan DA Ge S Kaga A Tomooka N 2008 a) Phylogeny and biogeography of the genus Oryza. In: Rice Biology in the Genomics Era (eds Hirano HY, Hirai A, Sano Y, Sasaki T) , pp 219 -231 . Springer-Verlag, Berlin , Heidelberg .
[本文引用: 2]
[61]
Vaughan DA Lu BR Tomooka N 2008 b) Was Asian rice (Oryza sativa ) domesticated more than once?
Rice , 1 , 16 -24 .
[本文引用: 3]
[62]
Vaughan DA Morishima H Kadowaki K 2003 ) Diversity in the Oryza genus
. Current Opinion in Plant Biology , 6 , 139 -146 .
[本文引用: 1]
[63]
Vitte C Ishii T Lamy F Brar D Panaud O 2004 ) Genomic paleontology provides evidence for two distinct origins of Asian rice (Oryza sativa L.)
. Molecular Genetics and Genomics , 272 , 504 -511 .
[本文引用: 1]
[64]
Wang H Vieira FG Crawford JE Chu CC Nielsen R 2017 ) Asian wild rice is a 477 hybrid swarm with extensive gene flow and feralization from domesticated rice
. Genome Research , 27 , 1029 -1038 .
[本文引用: 1]
[65]
Wang XK 1996 ) New research progresses relating to several main problems about the origin of rice cultivation in China. In: Origin and Differentiation of Chinese Cultivated Rice (eds Wang XK, Sun CQ) , pp. 2 -7. China Agricultural University Press , Beijing . (in Chinese with English abstract)
[本文引用: 1]
[王象坤 (1996 ) 中国稻作起源研究中几个主要问题的研究新进展. 见: 人类历史发展的必然选择 . 重庆出版社 , 重庆 .]
[本文引用: 1]
[66]
Wang YH Li JY 2005 ) The plant architecture of rice (Oryza sativa )
. Plant Molecular Biology , 59 , 75 -84 .
[本文引用: 1]
[67]
Wei X Qiao WH Chen YT Wang RS Cao LR Zhang WX Yuan NN Li ZC Zeng HL Yang QW 2012 ) Domestication and geographic origin of Oryza sativa in China: Insights from multilocus analysis of nucleotide variation of O. sativa and O. rufipogon
. Molecular Ecology , 21 , 5073 -5087 .
[本文引用: 3]
[68]
Wu XH Zhang C Goldberg P Cohen D Pan Y Arpin T Bar-Yosef O 2012 ) Early pottery at 20,000 years ago in Xianrendong Cave, China
. Science , 336 , 1696 -1700 .
[本文引用: 1]
[69]
Yang XY Fuller DQ Huan XJ Perry L Li Q Li Z Zhang JP Ma ZK Zhuang YJ Jiang LP Ge Y Lu HY 2015 ) Barnyard grasses were processed with rice around 10000 years ago
. Scientific Reports , 5 , 16251 .
[本文引用: 1]
[70]
Zhang LB Zhu QH Wu ZQ Ross-Ibarra J Gaut BS Ge S Sang T 2009 ) Selection on grain shattering genes and rates of rice domestication
. New Phytologist , 184 , 708 -720 .
[本文引用: 3]
[71]
Zhang WX Gu HB 2005 ) Study of ancient rice from Chengtoushan site in Lixian County, Hunan Province
. Acta Agronomica Sinica , 31 , 736 -741 . (in Chinese with English abstract)
[本文引用: 2]
[张文绪 , 顾海滨 (2005 ) 湖南澧县城头山遗址古稻研究
. 作物学报 , 31 , 736 -741 .]
[本文引用: 2]
[72]
Zhang WX Pei AP Mao TL 2003 ) Studies on the printings of bi-peaked tubercles on lemmas of rice kernels in broken pottery pieces unearthed from Pengtoushan excavation site in Li County, Hunan
. Acta Agronomica Sinica , 29 , 263 -267 . (in Chinese with English abstract)
[本文引用: 1]
[张文绪 , 裴安平 , 毛同林 (2003 ) 湖南澧县彭头山遗址陶片中水稻稃壳双峰乳突印痕的研究
. 作物学报 , 29 , 263 -267 .]
[本文引用: 1]
[73]
Zhang WX Yuan JR 1998 ) Primary study of ancient rice from Yuchanyan site in Daoxian County, Hunan Province
. Acta Agronomica Sinica , 24 , 416 -420 . (in Chinese with English abstract)
[本文引用: 1]
[张文续 , 袁家荣 (1998 ) 湖南道县玉蟾岩古栽培稻的初步研究
. 作物学报 , 24 , 416 -420 .]
[本文引用: 1]
[74]
Zhao DL Pei AP Zhang WX 2000 ) Restudy of ancient cultivated rice from Bashidang site in Lixian County, Hunan Province
. Chinese Journal of Rice Sicence , 14 (3 ), 139 -143 . (in Chinese with English abstract)
[本文引用: 1]
[赵笃乐 , 裴安平 , 张文绪 (2000 ) 湖南澧县八十垱遗址古栽培稻的再研究
. 中国水稻科学 , 14 (3 ), 139 -143 .]
[本文引用: 1]
[75]
Zhao KY Wright M Kimball J Eizenga G McClung A Kovach M Tyagi W Ali ML Tung CW Reynolds A Bustamante CD McCouch SR 2010 ) Genomic diversity and introgression in O. sativa reveal the impact of domestication and breeding on the rice genome
. PLoS ONE , 5 , e10780 .
[本文引用: 1]
[76]
Zhao Y Vrieling K Liao H Xiao MQ Zhu YQ Rong J Zhang WJ Wang YG Yang J Chen JK Song ZP 2013 ) Are habitat fragmentation, local adaptation and isolation-by-distance driving population divergence in wild rice Oryza rufipogon ?
Molecular Ecology , 22 , 5531 -5547 .
[本文引用: 2]
[77]
Zhao ZJ 1998 ) The middle Yangtze region in China is one place where rice was domesticated: Phytolith evidence from the Diaotonghuan cave, northern Jiangxi
. Antiquity , 278 , 885 -897 .
[本文引用: 1]
[78]
Zhao ZJ 2009 ) New informations and research advances about the origin of cultivated rice and rice agriculture
. Southern Relics , 3 , 59 -63 . (in Chinese)
[本文引用: 9]
[赵志军 (2009 ) 栽培稻与稻作农业起源研究的新资料和新进展
. 南方文物 , 3 , 59 -63 .]
[本文引用: 9]
[79]
Zhao ZJ 2011 ) New archaeobotanic data for the study of the origins of agriculture in China
. Current Anthropology , 52 (S4 ), S295 -S306 .
[本文引用: 1]
[80]
Zhao ZJ Jiang LP 2016 ) Analysis of plant relics from Shangshan site in Pujiang of Zhejiang
. Southern Relics , 3 , 109 -116 . (in Chinese with English abstract)
[本文引用: 1]
[赵志军 , 蒋乐平 (2016 ) 浙江浦江上山遗址浮选出土植物遗存分析
. 南方文物 , 3 , 109 -116 .]
[本文引用: 1]
[81]
Zheng YF Chen XG Wang HM 2013 ) Rice cultivation from Xiaohuangshan site in Shengzhou of Zhejiang Province, based on phytoliths analysis
. Agricultural Archaeology , 4 , 11 -17 . (in Chinese with English abstract)
[本文引用: 1]
[郑云飞 , 陈旭高 , 王海明 (2013 ) 浙江嵊州小黄山遗址的稻作生产——来自植物硅酸体的证据
. 农业考古 , 4 , 11 -17 .]
[本文引用: 1]
[82]
Zheng YF Jiang LP 2007 ) Rice relics from Shangshan site and its implications
. Archaeology , 9 , 19 -25 . (in Chinese with English abstract)
[本文引用: 2]
[郑云飞 , 蒋乐平 (2007 ) 上山遗址出土的古稻遗存及其意义
. 考古 , 9 , 19 -25 .]
[本文引用: 2]
[83]
Zheng YF Jiang LP Zheng JM 2004 ) Study on the remains of ancient rice from Kuahuqiao site in Zhejiang Province
. Chinese Journal of Rice Science , 18 (2 ), 119 -124 . (in Chinese with English abstract)
[本文引用: 1]
[郑云飞 , 蒋乐平 , 郑建明 (2004 ) 浙江跨湖桥遗址的古稻遗存研究
. 中国水稻科学 , 18 (2 ), 119 -124 .]
[本文引用: 1]
[84]
Zhu QH Ge S 2005 ) Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes
. New Phytologist , 167 , 249 -265 .
[本文引用: 1]
[85]
Zong Y Chen Z Innes JB Chen C Wang Z Wang H 2007 ) Fire and flood management of coastal swamp enabled first rice paddy cultivation in East China
. Nature , 449 , 459 -462 .
[本文引用: 1]
[86]
Zuo XX Lu HY Jiang LP Zhang JP Yang XY Huan XJ He KY Wang W Wu NQ 2017 ) Dating rice remains through phytolith carbon-14 study reveals domestication at the beginning of the Holocene
. Proceedings of the National Academy of Sciences,USA , 114 , 6486 -6491 .
[本文引用: 3]
Artificial selection for a green revolution gene during japonica rice domestication
1
2011
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
Agricultural origins and the isotopic identity of domestication in northern China
1
2009
... 植物的驯化与利用催生了农耕文明.农耕文明的起源与发展, 促进了工具的运用与不断改良, 转变了人类的生活方式, 促使人类由迁徙转变为定居, 出现聚落、城邦及相应的社会组织结构, 为人类文明的发展奠定了基础.中华文明(或华夏文明)是世界上唯一一直传承的古老文明.考古证据显示, 长江流域(总体位于24-35° N之间)有着和黄河流域一样辉煌的史前文明, 两者都是中华文明之源; 长江文明以稻作文明为内核, 黄河文明则以粟作文明为其鲜明特征(Barton et al, 2009 ; Zhao, 2011 ). ...
Genome-wide patterns of nucleotide polymorphism in domesticated rice
2
2007
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Archaeogenetic study of prehistoric rice remains from Thailand and India: Evidence of early japonica in South and Southeast Asia
2
2016
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
The rice paradox: Multiple origins but single domestication in Asian rice
5
2017
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
... 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
Multiple origin but single domestication led to Oryza sativa. G3-Genes Genomes Genetics
2
2018
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
Origin of rice (Oryza sativa L.) domestication genes
1
2017
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
Three geographically separate domestications of Asian rice
1
2015
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Seed dormancy in red rice. III. Response to nitrite, nitrate, and ammonium ions
1
1983
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
Archaeological and genetic insights into the origins of domesticated rice
3
2014
... 水稻驯化及稻作农业为长江文明的发展和繁荣提供了重要的物质和文化基础, 也是长江文明为人类做出的巨大的开创性贡献(Larson et al, 2014 ).关于水稻是何时、何地、在什么环境下开始驯化等问题一直是国际学术界关注的热点.得益于新技术手段的不断运用, 越来越多的考古学和遗传学研究结果显示, 中国最早开始利用水稻, 长江流域及以南地区是水稻的驯化起源地(Cross & Zhao, 2014 ).本文简要评述了近年来有关水稻起源的考古学和遗传学研究进展, 特别关注长江流域的相关研究, 分析了各种研究的结论不一致的可能原因, 并讨论了未来值得深入研究的科学问题. ...
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World
3
2007
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 粒形是另一个重要驯化性状.古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择.水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 ).一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 ).粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 ).水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 ).水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 ). ...
Pathways to Asian civilizations: Tracing the origins and spread of rice and rice cultures
1
2011
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
Rice archaeobotany revisted
1
2008
... 粒形是另一个重要驯化性状.古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择.水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 ).一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 ).粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 ).水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 ).水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 ). ...
The domestication process and domestication rate in rice: Spikelet bases from the Lower Yangtze
2
2009
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
Consilience of genetics and archaeobotany in the entangled history of rice
5
2010
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 ).从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻.落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 ). ...
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
Nonindependent domestication of the two rice subspecies, Oryza sativa ssp. indica and ssp. japonica , demonstrated by multilocus microsatellites
1
2008
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Genetic structure and diversity in Oryza sativa L
2
2005
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
In appropriate model rejects independent domestications of indica and japonica rice
1
2011
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
5
2017
... 粒形是另一个重要驯化性状.古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择.水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 ).一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 ).粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 ).水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 ).水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 ). ...
... 据Silva等(2015) 统计, 世界各地已发现的涉及亚洲栽培稻的水稻遗址共有470多处, 其中约1/3分布在中国.Gong等(2007) 认为中国共有280处水稻文物遗址.公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处.通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 ).这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程. ...
... 认为中国共有280处水稻文物遗址.公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处.通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 ).这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程. ...
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... 4,000 BP以后, 古稻遗址以长江中下游为中心, 向北、向西、向南辐射, 展示了稻作文明繁荣与传播的图景.这些遗址古稻(包括来自长江流域的)基本上属于粳型, 而且与现代粳稻类型基本一致(公婷婷, 2017 ). ...
5
2017
... 粒形是另一个重要驯化性状.古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择.水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 ).一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 ).粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 ).水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 ).水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 ). ...
... 据Silva等(2015) 统计, 世界各地已发现的涉及亚洲栽培稻的水稻遗址共有470多处, 其中约1/3分布在中国.Gong等(2007) 认为中国共有280处水稻文物遗址.公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处.通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 ).这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程. ...
... 认为中国共有280处水稻文物遗址.公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处.通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 ).这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程. ...
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... 4,000 BP以后, 古稻遗址以长江中下游为中心, 向北、向西、向南辐射, 展示了稻作文明繁荣与传播的图景.这些遗址古稻(包括来自长江流域的)基本上属于粳型, 而且与现代粳稻类型基本一致(公婷婷, 2017 ). ...
The temporal and spatial distribution of ancient rice in China and its implications
2007
Archaeological and genetic insights into the origins of domesticated rice
1
2014
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice
1
2011
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
论稻作萌生与成熟的时空问题
2
2014
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
... ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
论稻作萌生与成熟的时空问题
2
2014
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
... ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
The domestication syndrome
1
1984
... 在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 ).从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻.落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 ). ...
Two evolutionary histories in the genome of rice: The roles of domestication genes
1
2011
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Holocene evolution in weathering and erosion patterns in the Pearl River delta
1
2013
... 尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议.第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 ).第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 ).未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化. ...
Phylogeography of Asian wild rice, Oryza rufipogon : A genome-wide view
1
2012
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Association between the geographic distribution during the last glacial maximum of Asian wild rice, Oryza rufipogon (Poaceae), and its current genetic variation
4
2012
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议.第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 ).第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 ).未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化. ...
A map of rice genome variation reveals the origin of cultivated rice
2012b
New evidence for the origins of sedentism and rice domestication in the Lower Yangtze River, China
1
2006
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
Genetic control of rice plant architecture under domestication
2
2008
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
... ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
Archaeology: Origins of agriculture in East Asia
2
2009
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议.第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 ).第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 ).未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化. ...
Origin, dispersal, cultivation and variation of rice
1
1997
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
An SNP caused loss of seed shattering during rice domestication
2
2006
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
New insights into the history of rice domestication
2
2007
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
Current perspectives and the future of domestication studies
1
2014
... 水稻驯化及稻作农业为长江文明的发展和繁荣提供了重要的物质和文化基础, 也是长江文明为人类做出的巨大的开创性贡献(Larson et al, 2014 ).关于水稻是何时、何地、在什么环境下开始驯化等问题一直是国际学术界关注的热点.得益于新技术手段的不断运用, 越来越多的考古学和遗传学研究结果显示, 中国最早开始利用水稻, 长江流域及以南地区是水稻的驯化起源地(Cross & Zhao, 2014 ).本文简要评述了近年来有关水稻起源的考古学和遗传学研究进展, 特别关注长江流域的相关研究, 分析了各种研究的结论不一致的可能原因, 并讨论了未来值得深入研究的科学问题. ...
Rice domestication by reducing shattering
3
2006
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
Origin of seed shattering in rice (Oryza sativa L.)
1
2007
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
2
2006
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
2
2006
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
Population genetic structure of Oryza rufipogon and Oryza nivara : Implications for the origin of O. nivara
1
2015
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
Early Holocene groundwater table fluctuations in relation to rice domestication in the middle Yangtze River basin, China
1
2017
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
Phylogeography of Asian wild rice, Oryza rufipogon , reveals multiple independent domestications of cultivated rice, Oryza sativa
2
2006
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Genomewide SNP variation reveals relationships among landraces and modern varieties of rice
1
2009
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
Molecular evidence for a single evolutionary origin of domesticated rice
2
2011
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
亚洲栽培稻主要驯化性状研究进展
3
2012
... 在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 ).从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻.落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 ). ...
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
亚洲栽培稻主要驯化性状研究进展
3
2012
... 在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 ).从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻.落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 ). ...
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
Paleo-green revolution for rice
1
2011
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
The domestication syndrome genes responsible for the major changes in plant form in the Triticeae crops
1
2011
... 在人类栽培过程中, 植物逐渐发生关键的形态(和遗传)改变, 以适应栽培环境, 在性状上表现出驯化适应性综合征(adaptive syndrome of domestication), 包括更加紧凑的株型、花序增大且小花数目增多、果实或种子变大、落粒性减弱, 以及种子休眠性降低等(Hammer, 1984 ).从普通野生稻到栽培稻的转变过程中, 野生稻逐渐失去了多年生、落粒性、长芒、匍匐生长等特征, 成为依赖人工种植的栽培稻.落粒性、休眠性和株型等性状是水稻驯化的关键性状, 种皮颜色、籽粒形态等性状也与水稻驯化密切相关(Fuller et al, 2010 ; Sakuma et al, 2011 ; 区树俊等, 2012 ). ...
Genetics and phylogenetics of rice domestication
1
2007
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
Origin of the genetic diversity of cultivated (Oryza spp.) study of the polymorphism scored at 40 isozyme loci
1
1982
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Mechanism of seed dormancy in rice
1
1991
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
Deletion in a gene associated with grain size increased yields during rice domestication
1
2008
... 粒形是另一个重要驯化性状.古人在栽培植物过程中, 逐渐意识到粒形是产量性状, 开始对大粒形种子进行有意识的选择.水稻粒形(谷粒大小)是重要的产量和品质农艺性状, 是水稻驯化过程中最突出的生物学特征变化之一(Fuller et al, 2008 ).一般情况下, 普通野生稻的种子粒形为细长形且附有长芒, 籼稻长圆形或细长形, 而粳稻则为阔卵形或短圆形(图1 ).粒形(包括颖壳)可以较好地区别野生种与栽培种, 由于稻谷相对容易保存, 出土稻谷遗存较多, 粒形分析成为研究栽培稻驯化的重要手段(Fuller, 2007 ).水稻粒形也是数量遗传性状, 包含粒长、粒宽和粒厚等指标, 其中粒宽更引人注意, 其驯化时间早于粒长(公婷婷, 2017 ).水稻全基因组测序的完成促进了粒宽性状的控制基因的发掘, 已有多个粒宽主效基因被克隆, 例如qSW 5, 该基因在粳稻群体中受到强烈选择(Shomura et al, 2008 ). ...
Modelling the geographical origin of rice cultivation in Asia using the rice archaeological database
2
2015
... 据Silva等(2015) 统计, 世界各地已发现的涉及亚洲栽培稻的水稻遗址共有470多处, 其中约1/3分布在中国.Gong等(2007) 认为中国共有280处水稻文物遗址.公婷婷(2017) 对我国水稻考古遗址重新进行了较全面的梳理, 认为有明确的实物遗存的水稻遗址为219处.通过对上述资料进行比对和重新统计相关资料, 我们共获得了328处中国的水稻考古遗址信息, 按照年代进行等级划分, 制作了中国古稻分布图(图2 ).这些水稻遗址空间分布格局、出土的植物遗存和其他文物类型的变迁, 大致反映了水稻驯化起源与扩散的历史脉络, 以及稻作农业的形成与稻作文明的发展与传播历程. ...
... 尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议.第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 ).第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 ).未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化. ...
Evolutionary insights based on SNP haplotypes of red pericarp, grain size and starch synthase genes in wild and cultivated rice
1
2017
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
Evidences of introgression from cultivated rice to Oryza rufipogon (Poaceae) populations based on SSR fingerprinting: Implications for wild rice differentiation and conservation
1
2006
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
RFLP analysis of nuclear DNA in common wild rice (Oryza rufipogon Griff.) and cultivated rice (Oryza sativa L.)
1
1997
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
The complex history of the domestication of rice
3
2007
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
... 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
Global dissemination of a single mutation conferring white pericarp in rice
2007
Control of a key transition from prostrate to erect growth in rice domestication
1
2008
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
Genomic variation in rice: genesis of highly polymorphic linkage blocks during domestication
1
2006
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
2
2008
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
... ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
Was Asian rice (Oryza sativa ) domesticated more than once?
3
2008
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
... 普通野生稻种子具有较强休眠性, 这有利于其度过不利的生长季节, 栽培稻则没有明显的休眠性, 这适应于生产上同步发芽的需要(Cohn et al, 1983 ; Sweeney & McCouch, 2007 ).普通野生稻的种皮通常是红色, 而栽培稻多为白色; 野生稻颖壳黑色或褐黑色, 栽培稻多为浅黄色(图1 ), 这些都与种子休眠性的差异有关(Seshu & Dadlani, 1991 ).通过分子遗传学研究, 人们已发现了多个与水稻种子休眠相关的QTL位点(区树俊等, 2012 ), 其中qSD 7-1位点通过母体组织(包括颖壳、果皮和种皮)来控制水稻种子休眠(Gu et al, 2011 ).rc 基因导致水稻白种皮(Sweeney et al, 2007 ), 该基因与sh 4类似, 起源于粳稻而后扩散到籼稻(Vaughan et al, 2008b ; Cross & Zhao, 2014 ). ...
Diversity in the Oryza genus
1
2003
... 稻属包含2个栽培种和20多个野生种, 栽培种分别为亚洲栽培稻(O. sativa )和非洲栽培稻(O. glaberrima ) (Vaughan et al, 2003 , 2008b ).我们通常所称的水稻是指亚洲栽培稻.与水稻遗传关系最近的是普通野生稻(O. rufipogon )和尼瓦拉野生稻(O. nivara ) (Vaughan et al, 2008a ), 它们广泛分布在南亚和东南亚以及中国部分地区的永久性或季节性湿地中(Fuller et al, 2010 ).普通野生稻为多年生, 对光周期敏感, 异交率较高; 尼瓦拉野生稻则是一年生, 对光周期不敏感, 自交率比较高(Vaughan et al, 2008a ).这两种野生稻虽然有明显的遗传分化(Liu et al, 2015 ), 但它们遗传上都与水稻关系密切, 以至于究竟谁是水稻的直接祖先种存在争议.目前广泛支持普通野生稻是水稻的祖先种(Khush, 1997 ; Castillo et al, 2016 ). ...
Genomic paleontology provides evidence for two distinct origins of Asian rice (Oryza sativa L.)
1
2004
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Asian wild rice is a 477 hybrid swarm with extensive gene flow and feralization from domesticated rice
1
2017
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
1
1996
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
1
1996
... 因适应不同栽培区的气候和环境条件, 水稻分化形成了不同的亚种或生态型, 其中最重要的两个亚种是粳稻(O. sativa ssp. japonica )和籼稻(O. sativa ssp. indica ), 其次还有aus稻和香稻(aromatic) (Garris et al, 2005 ; Caicedo et al, 2007 ; Vaughan et al, 2008b ; McNally et al, 2009 ).粳稻主要分布于东亚的亚热带地区、东南亚的高原和南亚高地, 而籼稻则主要集中在亚洲的热带地区.水稻驯化起源问题实际上还牵涉到不同栽培亚种, 特别是籼稻和粳稻的起源分化(indica-japonica differentiation)问题(王象坤, 1996 ; Kovach et al, 2007 ), 因而愈发扑朔迷离. ...
The plant architecture of rice (Oryza sativa )
1
2005
... 水稻株型主要由分蘖数、分蘖角度、株高和穗型所决定(Wang & Li, 2005 ).普通野生稻通常分蘖多, 分蘖角度大而匍匐生长, 散穗型; 栽培稻则分蘖少, 分蘖角度小而直立生长, 紧穗型.目前已经发现多个与株型性状相关的基因(区树俊等, 2012 ), 其中最引人注意的是prog 1基因(Jin et al, 2008 ; Tan et al, 2008 ).该基因位于水稻第7号染色体上, 编码具有转录活性的锌指蛋白类转录因子, 在叶腋分生组织(分蘖芽的形成部位)、芽茎尖分生组织和幼叶中高表达, 控制分蘖角度和分蘖数, 受到了人工选择, 使水稻的株型由野生型转变成驯化型(Jin et al, 2008 ).另外, “绿色革命”基因(矮杆基因) sd 1在水稻驯化过程中也受到人为驯化选择(Asano et al, 2011 ), 相对于普通野生稻, 栽培稻中sd 1基因序列的遗传多样性减少了98% (Paterson & Li, 2011 ). ...
Domestication and geographic origin of Oryza sativa in China: Insights from multilocus analysis of nucleotide variation of O. sativa and O. rufipogon
3
2012
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
... 尽管关于水稻起源问题, 考古学和遗传学研究得出的结论总体趋于一致, 但仍有争议.第一, 考古学证据明确显示长江流域是水稻的最早起源地(Silva et al, 2015 ), 而且考古学研究结果还显示, 农业大约在2,700 BP才开始进入珠江流域(Hu et al, 2013 ), 但部分遗传学研究结果则倾向于水稻起源于珠江流域(如Huang et al, 2012b ; Wei et al, 2012 ).第二, 考古学估计的水稻起源时间(~10,000 BP)明显近于遗传学研究基于分子钟估算的时间(一般>13,000 BP) (Jones & Liu, 2009 ).未来采用更加灵敏的分析技术, 比如建立水稻遗存物的古DNA高效提取与分析技术体系, 或者是单粒古孢粉测序技术, 有望消除栽培稻基因流对分析结果的干扰, 从而追溯水稻驯化过程中的遗传变化. ...
Early pottery at 20,000 years ago in Xianrendong Cave, China
1
2012
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
Barnyard grasses were processed with rice around 10000 years ago
1
2015
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
Selection on grain shattering genes and rates of rice domestication
3
2009
... 不落粒作为农作物驯化的关键性状, 是区分栽培种及其野生祖先的最主要依据之一(Jones & Liu, 2009 ).普通野生稻的籽粒成熟时, 护颖和小穗轴的接合处(小穗基盘)一般会形成离层, 这使得种子易于脱落与传播, 但不利于种子的收获; 栽培稻小穗基盘的离层发育不完全, 成熟种子需借助外力才能脱粒, 从而有利于稻谷的收获(Li et al, 2006 ).因此, 普通野生稻小穗基盘光滑, 而栽培稻粗糙(Fuller, 2007 ).小穗轴是植物考古中通过浮选法相对容易获得的水稻实物遗存之一, 因而小穗基盘的形态业已成为判断古稻类型的重要依据之一(如Fuller et al, 2009 ).落粒性是数量遗传性状(QTL), 遗传学研究先后确定了多个与此性状有关的QTL位点, 如 sh- h、sh 4、qsh 1等(Konishi et al, 2006 ; Li et al, 2006 ).其中sh 4和qsh 1是控制水稻落粒的主要基因(Zhang et al, 2009 ).sh 4为一个隐性突变, 是广泛存在于所有栽培稻(粳稻、籼稻、香稻和aus稻)的4号染色体上的一个基因的SNP, 会导致离层发育不完全(Li et al, 2006 ).sh 4是与栽培稻驯化起源关联的主要突变(Sang & Ge, 2007 ; Fuller et al, 2009 ).分子系统学研究显示, 粳稻和籼稻的sh 4基因位于同一进化支上, 暗示该基因是通过一次驯化保留到基因组上, 这被认为是水稻单次起源或者籼粳分化出现在水稻驯化之后的最有力证据(Lin et al, 2007 ; Molina et al, 2011 ).qsh 1主要存在于温带粳稻中(Konishi et al, 2006 ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... ), 是在促使东亚水稻向温带粳稻分化的遗传瓶颈过程中或者之后被固定下来的(Zhang et al, 2009 ). ...
... 理论上, 对驯化相关基因的进化分析可以解决栽培物种的驯化起源问题, 然而相关研究结果却存在分歧.关键驯化基因, 如落粒性基因sh 4, 红种皮基因rc , 株型基因prog 1的系统重建都指向栽培稻为中国单起源(Sweeney et al, 2007 ; Zhang et al, 2009 ; Choi et al, 2017 ; Choi & Purugganan, 2018 ).但是, Civáň和Brown (2017) 对这些基因(还包括无芒基因LABA 1)及其在基因组上邻近序列进行分析后, 认为这些驯化基因在野生稻中出现的时间远早于栽培稻的驯化, 暗示不同栽培稻亚种有不同起源.Singh等(2017) 持类似观点.Choi等(2017) 以及Choi和Purugganan (2018)也发现驯化基因起源早于栽培稻的驯化, 并因此提出了“多起源/单驯化”假设(the multiple origin/single domestication model). ...
湖南澧县城头山遗址古稻研究
2
2005
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
湖南澧县城头山遗址古稻研究
2
2005
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
湖南澧县彭头山遗址陶片中水稻稃壳双峰乳突印痕的研究
1
2003
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
湖南澧县彭头山遗址陶片中水稻稃壳双峰乳突印痕的研究
1
2003
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
湖南道县玉蟾岩古栽培稻的初步研究
1
1998
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
湖南道县玉蟾岩古栽培稻的初步研究
1
1998
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
湖南澧县八十垱遗址古栽培稻的再研究
1
2000
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
湖南澧县八十垱遗址古栽培稻的再研究
1
2000
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
Genomic diversity and introgression in O. sativa reveal the impact of domestication and breeding on the rice genome
1
2010
... 纵观有关栽培稻驯化的遗传学和分子进化生物学研究, 我们初步可以总结如下几点: (1)普通野生稻O. rufipogon 是栽培稻的直接祖先, 一年生野生稻O. nivara 也是栽培稻的祖先种; (2)栽培稻亚种粳稻和籼稻遗传分化明显, 分别于不同的时间起源于不同的地区(多起源); (3)栽培稻最早起源于中国长江流域及以南地区(包括珠江流域), 粳稻驯化早于籼稻; (4)关键驯化基因的起源时间早于栽培稻的驯化; (5)杂交及基因流在栽培稻(尤其是籼稻)的驯化起源以及驯化基因的扩散中发挥了重要作用.值得注意的是, 栽培稻自被驯化以来, 就一直与野生稻频繁发生基因交流, 存在共进化关系(Song et al, 2006 ; Zhao et al, 2010 ; Ge & Sang, 2011 ; Choi et al, 2017 ).比如, 最近的大规模基因组分析结果表明, 普通野生稻种群广泛受到栽培稻的基因流的影响, 现存普通野生稻实际上是野-栽杂种群(hybrid swarm) (Wang et al, 2017 ).这意味着, 用现存普通野生稻为参照进行遗传关系分析时, 很难排除栽培稻基因流的干扰.稻作强度越大地区的野生稻种群受到栽培稻基因渐渗(introgression)的影响越大, 遗传上与栽培稻的关系也更近, 导致可能出现起源地的误判.充分利用考古发现的水稻实物遗存, 结合古DNA分析(如Castillo et al, 2016 ), 能够降低栽培稻基因流的干扰, 从而获得更加可信的结果. ...
Are habitat fragmentation, local adaptation and isolation-by-distance driving population divergence in wild rice Oryza rufipogon ?
2
2013
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
The middle Yangtze region in China is one place where rice was domesticated: Phytolith evidence from the Diaotonghuan cave, northern Jiangxi
1
1998
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
栽培稻与稻作农业起源研究的新资料和新进展
9
2009
... 水稻(即亚洲栽培稻Oryza sativa )是世界上最重要的粮食作物之一, 目前全球有超过半数以上人口以稻米为食.水稻起源于野生稻.先人从对野生稻的原始照看到有意识的栽培管理, 再到有目的的选育, 最终驯化出适应栽培环境的栽培稻, 并广泛栽培而形成稻作农业, 经过了长达数千年的发展历程(赵志军, 2009 ).在此过程中, 一方面, 野生稻原有的一些性状经过人工选择, 逐渐发生适应于栽培环境的改变, 形成驯化性状(domesticated traits), 最终演变为栽培稻; 另一方面, 因适应水稻栽培管理的需要, 人们不断改进生产工具, 变革生产方式, 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业, 逐步确立了长江流域以稻作为主要生产方式的农耕文明. ...
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
... ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
栽培稻与稻作农业起源研究的新资料和新进展
9
2009
... 水稻(即亚洲栽培稻Oryza sativa )是世界上最重要的粮食作物之一, 目前全球有超过半数以上人口以稻米为食.水稻起源于野生稻.先人从对野生稻的原始照看到有意识的栽培管理, 再到有目的的选育, 最终驯化出适应栽培环境的栽培稻, 并广泛栽培而形成稻作农业, 经过了长达数千年的发展历程(赵志军, 2009 ).在此过程中, 一方面, 野生稻原有的一些性状经过人工选择, 逐渐发生适应于栽培环境的改变, 形成驯化性状(domesticated traits), 最终演变为栽培稻; 另一方面, 因适应水稻栽培管理的需要, 人们不断改进生产工具, 变革生产方式, 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业, 逐步确立了长江流域以稻作为主要生产方式的农耕文明. ...
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
... ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
... 到了6,000-4,000 BP, 水稻遗址进一步增多, 共有178处, 其中近80处在长江流域.这些遗址出土了大量的水稻实物遗存, 以及生产、生活用具等文物, 充分反映了当时水稻广泛栽培与稻作农业快速发展的情景.这一时期, 长江流域的各遗址古稻基本上都表现为偏粳型, 比如城头山古稻.城头山遗址(6,000-5,000 BP) (111.60° E, 29.82° N)位于湖南省常德市澧县, 以出土时代最早、保存最完整的古城和灌溉设施完备的稻田而闻名.该遗址出土了稻、瓜等170多种人工种植和野生植物的籽粒, 以及猪、羊、狗、鹿等20多种家养和野生动物骨骸.稻谷粒形分析显示, 城头山古稻种群中含有类似野生稻粒形的个体仅占5.59%, 94%以上为栽培稻粒形(张文绪和顾海滨, 2005 ).这些证据表明, 在当时水稻驯化业已完成, 稻作农业体系已经建立, 稻作文明已经形成(赵志军, 2009 ; Fuller et al, 2010 ; Gross & Zhao, 2015 ). ...
... 遗传学研究关于水稻(粳稻)的最早起源的具体地点是长江流域还是珠江流域尚存在争议, 但可以概括为: 水稻最先起源于长江流域及以南地区(珠江流域).同时, 大量的考古学证据表明水稻于10,000-8,000 BP在长江流域开始驯化(Cross & Zhao, 2014 ).长江流域考古遗址中的古稻类型随年代下降逐渐转变为栽培稻类型(粳稻类型).伴随着此过程, 长江流域出现了一脉相承但有其特色的史前文明, 尤其是长江中游的农耕发展, 自彭头山以降, 经过皂市下层(111.40° E, 29.65° N)、汤家岗(112.17° E, 29.41° N)、大溪(109.63° E, 30.98° N)、屈家岭、石家河等连续发展出来的文化, 表现出了完整的进步过程, 到新石器中期的汤家岗文化, 已表现出相当成熟的农业文明形态(郭静云和郭立新, 2014 ).这个发展过程体现在诸多方面.首先, 出土文物中盛物用具和耕作工具逐渐增多, 而且越来越高级.从玉蟾岩遗址的最原始的夹炭陶, 到河姆渡文化时期(罗家角遗址)的白陶, 再到大溪文化(7,500-5,000 BP)与屈家岭文化(5,000-4,000 BP)时期的黑陶; 从原始粗糙的旧石器, 到精小石器, 发展到石质、骨质与木质器具(如河姆渡文化时期), 再到耒耜出现, 最后出现铜器、铁器; 从最早的刀耕水耨农业到耜耕农业、再到犁耕农业(刘军, 2006 ; Fuller, 2011 ).其次, 生活方式变迁.居住点从洞穴, 到台地, 再到聚落, 发展至城邦; 活动方式从迁徙, 到季节性迁徙, 到定居(郭静云和郭立新, 2014 ).第三, 生业经济从狩猎-采集, 到照顾植物与狩猎采集并重, 再到栽培植物为主, 进而养殖动植物的农业(赵志军, 2009 ).第四, 从直接利用湿地到雨浇地, 发展到具有排灌设施的稻田系统, 如城头山遗址(张文绪和顾海滨, 2005 ).制陶技艺、犁耕农业、排灌系统、城邦、古老的宗教与艺术(如高庙文化, 7,800-6,800 BP)等都是长江文明的重要符号, 它们的发展或出现无疑都是稻作农业发展的必然产物.因此, 可以说是稻作的起源与发展催生了长江文明, 长江文明以稻作文明为核心. ...
New archaeobotanic data for the study of the origins of agriculture in China
1
2011
... 植物的驯化与利用催生了农耕文明.农耕文明的起源与发展, 促进了工具的运用与不断改良, 转变了人类的生活方式, 促使人类由迁徙转变为定居, 出现聚落、城邦及相应的社会组织结构, 为人类文明的发展奠定了基础.中华文明(或华夏文明)是世界上唯一一直传承的古老文明.考古证据显示, 长江流域(总体位于24-35° N之间)有着和黄河流域一样辉煌的史前文明, 两者都是中华文明之源; 长江文明以稻作文明为内核, 黄河文明则以粟作文明为其鲜明特征(Barton et al, 2009 ; Zhao, 2011 ). ...
浙江浦江上山遗址浮选出土植物遗存分析
1
2016
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
浙江浦江上山遗址浮选出土植物遗存分析
1
2016
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
浙江嵊州小黄山遗址的稻作生产——来自植物硅酸体的证据
1
2013
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
浙江嵊州小黄山遗址的稻作生产——来自植物硅酸体的证据
1
2013
... 从距今9,000年开始, 稻作遗址逐渐増多(图2 ), 遗址中稻作遗存所处年代也更为确切, 比如长江中游地区的属于彭头山文化(9,000-8,300 BP)的八十垱(111.85° E, 29.76° N)、彭头山(111.65° E, 29.66° N)和宋家岗遗址(111.78° E, 29.72° N), 长江下游地区的小黄山遗址(120.73° E, 29.55° N)等.八十垱遗址出土了距今8,500-7,500年的数万粒完整形态的炭化稻谷、稻米, 还有环绕聚落的围墙、挡水坝, 以及大量的墓葬和以杆栏式建筑为主的居住房址, 还有大量的陶器、石器、骨器、木器、百余种植物杆茎与果核等.这些古稻从形态上具有原始性和过渡性的特征, 属于未经人工选择的原始栽培稻类型(赵笃乐等, 2000 ).最近的植物考古分析显示, 在人类定居之前(大约8,600年前), 八十垱所在地区为泛滥平原, 随后因全新世季风降水增多, 水位上升, 逐渐变成湿地, 辅以火烧和简单清理, 这些都有利于水稻栽培, 使得水稻驯化在该地区持续兴盛了数百年(Liu et al, 2017 ).宋家岗遗址出土了距今约9,000年的100多粒炭化稻米, 这些稻米集中分布在一处面积约4-5 m2 的生活区, 紧邻一处灶坑, 同时伴出大量夹有谷粒与稻壳的陶片, 少量石制品等生活用器, 人类食用水稻的痕迹明显(赵志军, 2009 ).彭头山遗址出土的陶器制造古朴简单, 全部为原始的贴塑法制成, 大部分陶器的胎泥中夹有炭屑, 部分含有稻壳与谷粒或稃壳印痕.张文绪等(2003) 用扫描电镜对这些陶片中的水稻稃壳印痕进行分析后认为, 这些古稻形态上已向粳稻演化.同样, 植硅石和颖壳双峰乳突的形态分析结果表明, 小黄山古稻属于原始栽培稻, 而且有向粳稻演化的趋势(郑云飞等, 2013 ).以上这些考古证据表明, 一方面我国长江流域在更新世晚期至全新世早期有野生稻分布, 另一方面栽培稻在10,000 BP左右开始出现, 尽管当时的生计形态仍处在采集狩猎阶段(赵志军, 2009 ; 公婷婷, 2017 ). ...
上山遗址出土的古稻遗存及其意义
2
2007
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
上山遗址出土的古稻遗存及其意义
2
2007
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
浙江跨湖桥遗址的古稻遗存研究
1
2004
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
浙江跨湖桥遗址的古稻遗存研究
1
2004
... 随着时间推移, 中国的古稻遗址迅速增多, 8,000-6,000 BP的水稻遗址有62处, 其中35处位于长江流域, 例如, 8,000-7,000 BP的位于长江下游的跨湖桥遗址(120.26° E, 30.18° N)等.跨湖桥遗址位于浙江省萧山, 出土的植物遗存包括水稻、菱角、芡实等.该遗址出土的稻谷经过鉴定, 60%为野生稻, 40%属于栽培稻(郑云飞等, 2004 ; 郑云飞和蒋乐平, 2007 ).跨湖桥遗址还出现了专门用于煮米饭的蒸锅“罾”, 显示米饭在食物中的地位开始提升.到了7,000-6,000 BP, 遗址古稻以栽培型为主, 既有粳型或向粳稻方向发展, 又有偏粳、偏籼混合或者偏粳、偏籼和中间型混合.河姆渡遗址(121.30° E, 29.98° N)距今7,000-5,800年间, 位于浙江余姚市, 出土了丰富的植物遗存, 包括稻谷、栎果、菱角、芡实等可食用植物遗存, 特别是大量的稻谷遗存, 显示河姆渡文化已经发展到了较发达的稻作农业生产阶段(刘军, 2006 ; 赵志军, 2009 ).田螺山遗址(121.40° E, 30.04° N)也位于浙江省余姚市, 距今年代约为7,000-5,500年间, 出土的食用植物包括稻谷、菱角、芡实、橡实, 还出土了木质和骨质的锄和锹等农业用具.Fuller等(2009)分析了田螺山6,900-6,600年前的土层中的植物遗存类型及水稻小穗基盘形态, 发现在此300年间, 植物遗存中水稻遗存的比例从8%上升到24%, 不落粒的栽培稻小穗基盘的比例从27%上升到39%, 稻田杂草如一年生的禾草、莎草和其他草本植物呈现聚集趋势, 这些都说明水稻的消费比例上升, 稻谷驯化正在不断加强, 出现了水稻种植.此外, 田螺山还发现了距今约7,000-4,500年的稻田.以上这些考古发现充分说明, 即使当时种植水稻的水平可能很低, 食物还大量依赖于采集和狩猎, 但稻作农业开始形成(赵志军, 2009 ). ...
Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes
1
2005
... 分子检测技术的进步极大推动了栽培稻驯化起源研究.中性标记检测结果显示, 粳稻和籼稻的遗传分化显著(Garris et al, 2005 ), 加之它们在形态上也有明显差异(图1 ), 这导致通常认为粳稻和籼稻有不同的起源(多源论).较早的等位酶分析结果将普通野生稻分为中国和东南亚两个遗传谱系, 因而推测粳稻和籼稻分别起源于不同地区(Second, 1982 ).类似地, Sun等(1997) 的RFLP检测结果显示, 中国普通野生稻种群偏粳稻, 而东南亚的普通野生稻种群偏籼稻, 普通野生稻已发生了籼粳分化, 粳稻和籼稻应是独立起源.随后的分子系统学研究将粳稻和籼稻的遗传分歧时间估计在0.4-0.2百万年前(Vitte et al, 2004 ; Zhu & Ge, 2005 ; Tang et al, 2006 ), 这明显早于考古学研究所估计的水稻驯化起源时间(大约10,000 BP), 暗示粳、籼稻分别起源于不同祖先群.进而, Londo等(2006) 基于两个核基因和一个叶绿体基因的序列分析, 认为粳稻大约在10,000年前起源于中国南方(泛指长江流域及以南地区), 而籼稻稍晚起源于东印度地区.粳、籼稻分别起源的观点得到了Caicedo等(2007) 的111个基因片段分析以及He等(2011) 基因组分析的结果的支持.然而, Gao和Innan (2008) 的SSR分析结果不支持粳、籼稻分别起源的观点.Molina等(2011) 同样支持单起源的观点, 认为栽培稻大约在13,500-8,200 BP起源于中国长江流域.Huang等(2012a) 的谱系地理学研究也支持栽培稻起源于中国的单起源假说, 但认为首先起源的是籼稻而非粳稻.通过对大样本(446份普通野生稻和1,083份栽培稻)的基因组分析, Huang等(2012b) 也支持单起源的观点, 并认为粳稻最先起源于中国的珠江流域, 这一观点得到Wei等(2012) 的研究结果的支持.但后者一方面支持中国珠江流域是栽培稻的驯化中心, 另一方面又认为粳、籼稻为独立起源(Wei et al, 2012 ).Civáň等(2015) 重新分析Huang等(2012b) 的基因组数据后认为, 水稻驯化为多起源, 粳稻、籼稻与aus稻分别起源于不同地区.最近, Choi等(2017) 以及Choi和Purugganan (2018) 的基因组研究结果也支持多起源的观点, 认为粳稻、籼稻、aus稻分别来自不同祖先群O. rufipogon 或O. nivara , 而且粳稻最先于18.3 ka (24.1-13.1 ka)前从分布于中国长江流域的O. rufipogon 中驯化出来. ...
Fire and flood management of coastal swamp enabled first rice paddy cultivation in East China
1
2007
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
Dating rice remains through phytolith carbon-14 study reveals domestication at the beginning of the Holocene
3
2017
... 理论上, 通过分析野生稻的地理和生态分布格局、历史气候变化对野生稻分布格局的影响以及野生种群与栽培稻的遗传关系, 就可以推测栽培稻起源地.水稻最初的驯化事件必然发生在先民能够直接利用野生种群的地区, 具有野生稻分布是确定水稻起源地的先决条件(Fuller et al, 2010 ).然而, 野生稻的现代分布格局是历史气候变化和人类土地利用活动共同影响的结果, 与历史分布存在差异.野生稻的自然分布一般集中在热带和亚热带, 对温度、湿度均有较高要求.普通野生稻的现代最北缘分布为我国江西省东乡县(28°14′ N), 但基于古气候、地形学和花粉数据重建古植被的研究结果显示, 野生稻可能曾在长江流域和华东地区具有广泛分布, 最北到达山东(Huang & Schaal, 2012 ; Zhao et al, 2013 ).谱系地理学研究结果提示, 历史上, 在更新世冰期干旱气候期间, 普通野生稻种群退缩到湿热的避难所, 如印度东部、云南和东南亚等区域(Londo et al, 2006 ).普通野生稻种群最近的地理扩张发生在大约16,500-15,000 BP的冰后期, 当时的气候变得湿暖, 而且CO2 浓度迅速从18 Pa (atmospheric partial pressure)上升到27 Pa, 这对植物特别是C3 植物水稻的光合效率和生物量生产有重要影响(Fuller et al, 2010 ).新仙女木时代(Younger Dryas, 13,000-11,500 BP)气候急剧变冷, 迫使普通野生稻向南退缩(Kovach et al, 2007 ).随后, 随着栽培稻的驯化与推广种植, 以及唐宋时期(1,200-1,000 BP)的气候又一次变冷, 普通野生稻的原生生境进一步减少和向南退缩, 最终形成现在的分布格局(Zhao et al, 2013 ).此外, 考古资料还证实, 距今10,000年前(栽培稻开始驯化时间), 普通野生稻是长江流域的习见物种(Fuller, 2007 ; Zong et al, 2007 ; Zuo et al, 2017 ).因此, 虽然当前长江流域仅在江西东乡县、湖南茶陵县和江永县等地有少量普通野生稻种群, 但栽培稻依然可能起源于此区域(Choi et al, 2017 ). ...
... 在中国, 早于8,000年的水稻遗址共有16处, 除了广东的牛栏洞遗址(113.45° E, 24.33° N)和河南的贾湖遗址(113.70° E, 33.62° N)外, 其余14处遗址全部位于长江流域(图2 ), 其中包括了世界上最古老的三个水稻遗址, 即江西省万年县的仙人洞遗址(117.179° E, 28.721° N)和吊桶环遗址(117.178° E, 28.721° N)、湖南省道县的玉蟾岩遗址(115.00° E, 25.50° N).这三处遗址都处于长江中游, 均是洞穴遗址, 年代为旧石器时代晚期向新石器时代早期的过渡时期(约13,000-11,000 BP) (赵志军, 2009 ; Wu et al, 2012 ).仙人洞与吊桶环相邻, 遗址内旧石器时代晚期的地层中发现了野生稻的植硅石(phytolith), 而在新石器时代初期的地层中则发现了栽培稻的植硅石, 年代距今1万年以前(Zhao, 1998 ).植硅石是指细胞组织中的蛋白石体(SiO2 •nH2 O), 可以在土壤中保留相当长的一段时间.不同植物的植硅石形态上存在差异, 比如粳稻的扇形植硅石为“厚而尖”的β型, 籼稻则是“薄而圆”的α型, 而普通野生稻植硅石的特征介于粳稻、籼稻之间, 因而常常被用于水稻考古研究(Zuo et al, 2017 ).玉蟾岩遗址不仅出土了一粒极为珍贵的、形态与普通野生稻相似但无芒的稻壳, 而且还出土了掺有稻秆和谷壳的贴塑法成型的夹炭陶(张文续和袁家荣, 1998 ).在稻作农业地区, 夹炭陶常常与古稻伴生出土, 而不从事稻作农业生产的地方往往只有夹砂陶.玉蟾岩的贴塑法成型工艺, 被认为是后代贴塑法工艺的开山鼻祖, 也是稻作农业发祥的重要佐证材料之一(赵志军, 2009 ). ...
... 接下来较古老的水稻遗址是上山遗址(11,000-9,000 BP) (119.89° E, 29.52° N)和荷花山遗址(10,000-9,000 BP) (119.30° E, 29.06° N).上山遗址出土了结构比较完整的木构建筑基址、灰坑、石器、陶器、骨器等文物, 特别引人注目的是出土了几十粒炭化稻米和少量稻穗基盘, 以及含有稻谷颖壳的碎陶片和烧制土残块.这些水稻遗存从稻谷的大小、小穗基盘的形态和植硅石类型上来看, 属于早期的栽培稻(Jiang & Liu, 2006 ; 郑云飞和蒋乐平, 2007 ; 赵志军和蒋乐平, 2016 ).对上山考古点出土的植物遗存的植硅石和淀粉微化石的进一步分析显示, 除了水稻之外, 稗草在当地大量出现, 其次还有橡实和菱角, 这说明当时的先人最初管理湿地环境是为了收获包括水稻和稗子在内的禾草植物的籽粒, 同时也暗示水稻是比较容易栽培的禾草而逐渐被驯化利用(Yang et al, 2015 ).Zuo等(2017) 对上山遗址和荷花山遗址的古稻的植硅石进行了比较分析, 发现从9,400-8,600 BP, 古稻的驯化程度在逐步加强, 但驯化速度缓慢, 进一步确认水稻开始驯化的年代大致在距今约1万年前后的全新世早期, 也表明我国长江流域水稻驯化与世界上其他主要农作物(西亚的小麦、中美洲的玉米)驯化基本上是同步的, 都发生在更新世末向全新世初过渡的时期, 这与当时逐渐变暖、变湿的全球气候格局转变有密切的关系. ...
 , 陈家宽
, 陈家宽


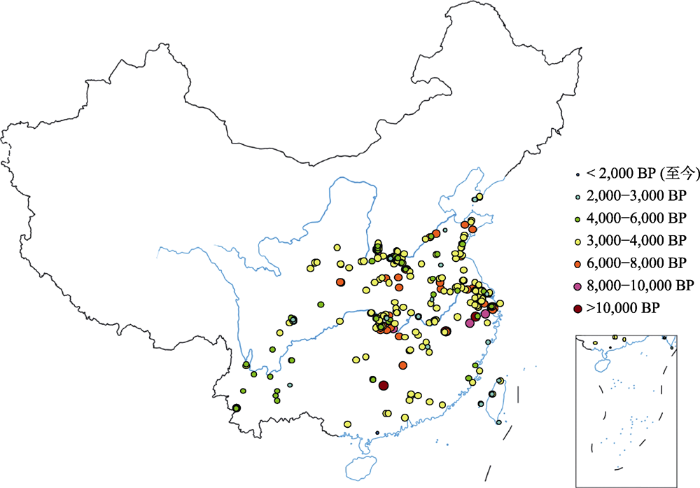
{kind=link}
{kind=link}
{kind=link}
{kind=link}