岩溶洞穴是一类特殊的自然景观, 极具观赏和游览价值。近年来, 岩溶洞穴资源被不断开发, 人类活动的频繁加剧了洞穴环境的恶化。由于岩溶洞穴的脆弱性和低自净能力, 一旦遭受污染就很难恢复(魏兴萍等, 2014 )。其中, 重金属污染是洞穴环境恶化的主要原因之一, 它不仅会危害环境, 而且会积累在人体中, 阻碍大脑和神经系统发育(Toth et al, 2016 )。通常, 岩溶洞穴与地下水是相通的, 污染物会通过渗透、淋溶等方式危及到地下水。同时, 洞穴重金属污染也会对洞穴生物群落构成威胁, 如: (1)洞穴物种多样性减少; (2)物种繁殖受阻; (3)群落稳定性降低等。因此, 对洞穴重金属进行监测对保护洞穴生物群落具有重要意义。
目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道。相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷。一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 )。不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料。
本文以贵州织金洞入口处为研究地点, 同时结合前人对岩溶洞穴环境的研究, 选取洞内可能存在的重金属污染元素Zn、Cr和Hg为监测对象。通过野外调查和室内研究相结合, 一方面可弥补织金洞世界地质公园苔藓植物类群数据的空白; 另一方面, 通过研究苔藓群落结构对重金属污染的响应特征, 探索苔藓群落对洞穴重金属污染的指示作用及其在重金属监测和评价方面的价值, 为环保监测部门积累基础资料。
1 材料与方法
1.1 区域概况
织金洞位于贵州省织金县官寨苗族乡内, 地处26°38°31°°-26°52°35°° N, 105°44°42°°-106°11°38°° E, 是典型的暗河型溶洞。洞内岩溶地下水丰富, 主要发育有织金洞古暗河系统和现代打铁洞暗河系统(李景阳等, 1991 )。全洞长12.1 km, 总面积70多万m2 。洞内地质构造复杂, 平均气温15℃, 相对湿度93%, 植被覆盖率低, 仅入口分布有大量的苔藓植物、蕨类植物和少量的草本、灌木植物。在织金洞旅游局的管理下, 洞内植物很少受到人为破坏。洞内苔藓植物主要分布于从入口至洞内82 m范围内, 出口门坎带也有少量分布。本研究主要在苔藓分布较为集中的入口处开展。
1.2 样品采集
于2017年9月, 在织金洞距入口不同深度(80 m、60 m、40 m、20 m、0 m)设置5个5 m × 5 m样点, 每个样点按照洞内环境特征随机设定10 cm × 10 cm重复样方数个, 样点信息见表1 。此外, 按照实际情况在出口分布有苔藓植物的地方设置2个样点共10个样方, 以补充织金洞苔藓物种数据。采集样方内全部苔藓和部分基质, 将每个样方内的全部苔藓记为一个群落, 利用优势种原则为群落命名(张朝晖和Pentecost, 2002 )。采集时, 每个样点均用数字照度计(PM6612L, 华谊联表)测定光照强度; 手持式空气温湿度计(HT-635, 广州宏诚科技)测定洞内空气温湿度; 手持式激光测距仪(SW-1500A, 深达威)测量距洞口距离。样方内每项指标测定3次, 取平均值。详细记录每个样地的海拔、坡度、坡向、盖度等详细生境信息, 共采集苔藓及基质样品各50份。
1.3 样品处理
将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定。使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定。苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测。对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解。
基质pH值用2.5:1水土悬液磁力搅拌, 采用玻璃电极法测定(上海虹益仪器仪表有限公司)。苔藓基质含量Cr、Zn和Ca采用HNO3 -HCLO4 -HF混酸法消解, 电感耦合等离子发射光谱(美国Perkin Elmer公司)测定。基质Hg含量用王水(50%)水浴消解法浸提, 苔藓Hg含量用高压密闭罐消解, AFS-230E双道原子荧光光度计(北京海光仪器公司)测定。实验所用硝酸、盐酸、氢氟酸、高氯酸均为优级纯, 30%过氧化氢和硫脲为分析纯, 分析用水均为超纯水。苔藓植物及基质含量在测定过程中分别采用柑橘叶标准物质(GSB-11)和国家标准物质(GSS-5、GSS-8、GSS-25、GSS-11)进行质量控制。同时设定全程序样品空白和20%平行双样, 检测误差要求在10%以内。所有测试工作在贵州师范大学分析测试中心完成。
1.4 数据分析
以盖度代替个体数分别计算研究区域各样点苔藓群落多样性指标(Mario & Bitetti, 2000 ; Amaral et al, 2013 ):
Patrick丰富度指数: D = Q (1)
Shannon-Wiener多样性指数:
${H}'=-\sum\limits_{i=\text{1}}^{m}{\text{(}{{p}_{i}}\times \ln {{p}_{i}}\text{)}}$ (2)
式中, Q 表示样地中的物种/群落数目, pi = Ni /N , Ni 为第i 个物种的个体数, N 为全部种的个体总数。
以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 )。重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 )。
重要值: IV = (相对盖度 + 相对频度)/200 (3)
生态优势度: $C={{\sum\limits_{i=1}^{s}{\left( \frac{I{{V}_{i}}}{IV} \right)}}^{2}}$ (4)
生态位宽度(Levins): ${{B}_{i}}=1/\sum\limits_{j=1}^{n}{p_{ij}^{2}}$ (5)
式中, 相对盖度或频度指某一种的盖度或频度占全部种盖度或频度的百分比, s 为样地内群落个数, IVi 为群落i 的重要值, IV 为群落中所有群落的重要值之和, Pij 为群落i 在j 个资源状态下的盖度占该群落所有盖度的比例, Bi 的范围为1-n 。
以实测基质重金属含量计算距洞口不同深度的各重金属元素污染超标倍数, 计算公式为:
${{P}_{i}}=\left( {{C}_{i}}-{{S}_{i}} \right)/{{S}_{i}}$ (6)
式中, Ci 为苔藓基质重金属含量实测值, Si 为该重金属的背景标准值。本研究背景标准值采用贵州省土壤背景值(国家环境保护局, 1990 )。
数据的差异性检验在SPSS 21.0里完成; 多样性数据的统计分析用R语言的Vegan包进行(郭水良等, 2015 ); 图表的绘制采用Excel 2016和Origin 9.0; RDA分析过程在CANOCO 5.0软件中进行。
2 结果
2.1 苔藓植物群落特征
对织金洞内苔藓群落进行统计分析, 共发现苔藓群落类型34种。为进一步解释洞内苔藓群落分布特征, 我们分别计算了各群落类型的重要值、生态优势度和生态位宽度(表2 )。其中尖叶美喙藓群落在这3项指标中均占据最大值(分别为1.62、0.0975、3.56), 说明尖叶美喙藓群落在织金洞内占有重要的生态地位, 对洞穴环境具有较大的生态适应范围。这3项指标较高的还有鳞叶凤尾藓群落(Fissidens taxifolius Com.) (分别为1.06、0.0635、2.56)、小凤尾藓群落(F. bryoides Com.) (分别为0.86、0.0515、2.42)、蛇苔群落(Conocephalum conicum Com.) (分别为0.71、0.0425、2.57)和粗肋凤尾藓群落(F. laxus Com.) (分别为1.03、0.0620、2.49)。
对洞内苔藓物种进行统计, 共发现苔藓植物种类20科33属49种, 其中苔类8科8属12种, 藓类12科25属37种。优势科为丛藓科、凤尾藓科和提灯藓科, 物种所占比例分别为16%、14%、14%。此外, 实地考察发现, 凤尾藓科群落对生长环境的要求具有很大的相似性, 该科物种在洞内石壁上具有较为广泛的分布, 并且相比生长于洞穴外部的凤尾藓科群落, 生长于洞穴内部的群落具有十分明显的向光性。这与洞穴内部长期受单侧光源照射有关(Christie et al, 1999 ), 洞穴内部环境阴暗、潮湿, 一些低矮的丛集型苔藓植物群落如凤尾藓在洞穴底部无法接触到光照从而退居到养分少但是相对干燥的洞穴石壁上, 以光为环境信号来调整自身的代谢、器官发生和形态建成。
2.2 苔藓群落基质重金属含量特征
重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 )。本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 。从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05)。其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重。距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大。从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致。Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 )。而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 )。因此, 应重视Hg的污染状况。
2.3 苔藓群落特征对重金属污染的生态监测
对表3 中基质重金属含量与相应的苔藓群落特征进行线性拟合。为消除数量级上的差异, 将拟合数据进行以10为底的对数转换处理, 其拟合结果见图1 。从图1 可以看出, 苔藓群落指标随着重金属元素含量的上升总体呈下降趋势, 这种趋势随着基质污染程度的加重愈加明显。所测定的重金属元素中基质Hg含量与苔藓群落指标的拟合程度最好, 与其对应的苔藓群落指标均呈显著负相关关系(P < 0.05), 说明苔藓群落特征能够很好地反映洞内Hg污染的变化趋势。其中Hg含量与苔藓群落丰富度指标的拟合方程为y = -0.3237x + 0.9367, r 2 = 0.81, P < 0.01, 与物种丰富度指标的拟合方程为y = -0.2561x + 1.2605, r 2 = 0.63, P < 0.05, 与物种多样性指标的拟合方程为y = -0.1707x + 0.7136, r 2 = 0.64, P < 0.05。基质Cr含量与苔藓物种丰富度指标和物种多样性指标呈显著负相关关系, 与群落丰富度指标无显著相关性。其中, Cr含量与物种丰富度指标的拟合方程为y = -0.1.021x + 3.2202, r 2 = 0.83, P < 0.01, 与物种多样性指标的拟合方程为y = -0.6308x + 1.9201, r 2 = 0.73, P < 0.01。Cr是自然界中广泛存在的重金属污染物之一, 对植物的胁迫主要表现为抑制植物的新陈代谢, 干扰其生物量的累积(龚林, 2014)。织金洞内Cr污染尚轻, 苔藓群落指标对Cr含量变化的响应说明其对环境变化的敏感程度较高。基质Zn含量与苔藓群落指标拟合度较差, 与其对应的群落指标均无显著相关性。
图1 织金洞基质重金属含量与苔藓群落指标的关系(数据进行以10为底的对数转换处理)
Fig. 1 Relationship between heavy metal content in substrate and community indexes of bryophytes in Zhijin Cave. Logarithmic conversion processing based on 10 is carried out on the data.
2.4 苔藓植物群落与基质重金属污染指数和洞穴环境因子的相互关系
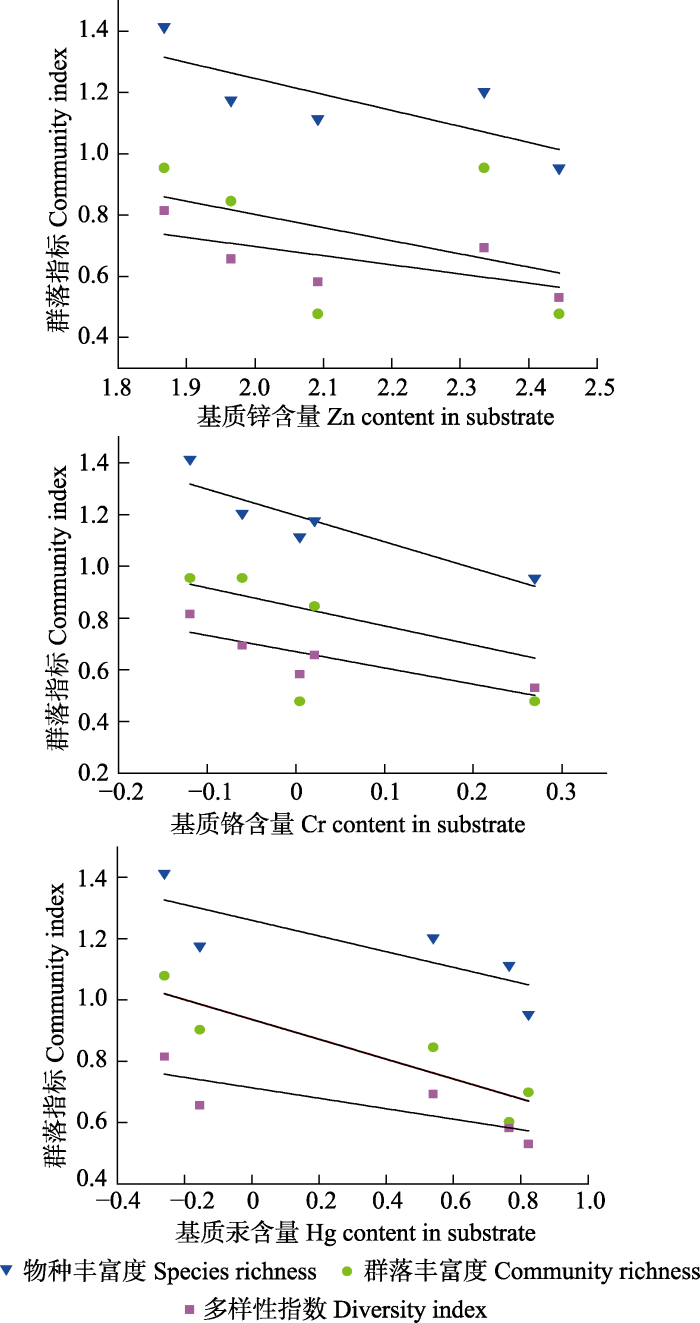
通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 )。第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息。RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量。耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴。光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求。在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 )。在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致。但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处。同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落。这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性。另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性。Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 )。
图2 苔藓优势群落与环境因子的冗余分析排序图。优势群落缩写见表2 。图中, 实心箭头代表重金属, 空心箭头代表其他环境因子, 星号代表苔藓优势群落; 箭头长短代表该变量对环境因子影响的大小, 各箭头之间的夹角代表变量之间的相关性, 夹角余弦值代表两变量之间的相关性系数; 苔藓群落与线段之间的连线长短代表群落与环境变量之间的对应关系。
Figs.2 RDA ordination diagram between dominant communityof bryophytes and environmental factors. Abbreviations of dominant communities are shown in Table 2 . In the figure, solid arrows represent heavy metals, hollow arrows represent other environmental factors, and asterisks represent dominant bryophyte communities. The length of the arrow represents the magnitude of the influence of the variable on the environmental factors, the included angle between the arrows represents the correlation between the variables, and the cosine value of the included angle represents the correlation coefficient between the two variables;The length of the connection line between bryophyte communities and line segments represents the corresponding relationship between communities and environmental variables.
2.5 苔藓Hg含量及其与基质Hg含量的相关性比较
上述结果表明, 织金洞内Hg污染较为严重, 并且对洞内苔藓群落有明显的胁迫作用。为了进一步说明织金洞内苔藓群落与Hg污染之间的关系, 我们从洞内优势群落中筛选出4种生物量较大的苔藓植物, 对其体内的Hg含量进行测定, 结果见表4 。从表4 中可以看出不同苔藓植物体中的平均Hg含量差异较大。其中, 粗肋凤尾藓(Fissidens laxus )的平均Hg含量明显低于其他3种, 这可能主要是因为它属于丛集型植物, 受洞内生长基质条件的限制, 植物体表面积较小, 对重金属的吸收能力较弱。植物体中平均Hg含量最高的是蛇苔(Conocephalum conicum ), 蛇苔属于平铺型植物, 体表面积较大, 能够更大程度上接触空气、水分并滞留大量的污染物颗粒, 从而增强了对重金属的吸附能力。
对4种苔藓植物及相应基质Hg含量进行相关性分析(表5 ), 结果发现仅蛇苔植物体内的Hg含量与其基质Hg含量具有显著相关性(P < 0.05), 说明蛇苔受基质Hg污染影响较大。在今后的治理过程中, 可以考虑将蛇苔作为洞内环境的生物监测材料之一。
3 讨论
苔藓群落具有很好的生物监测价值(Denayer et al, 1999 )。群落物种多样性特征可以反应环境污染状况(杨再超等, 2010)。环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 )。刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测。本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况。
目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况。由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测。而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广。首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集。其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小。第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采。因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 )。
蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 )。蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求。Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 )。在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等。然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属。当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落。这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 )。因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测。
致谢:织金洞管理局旅游服务中心人员在样品收集方面提供了大力帮助和支持, 吴启美老师对此文稿提出宝贵意见, 李芳同学参与野外采集标本工作。 在此致以衷心感谢!
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Amaral LDP Ferreira RA Lisboa GDS Longhi SJ Watzlawick LF 2013 ) Variabilidade espacial do Índice de Diversidade de Shannon-Wiener em Floresta Ombrófila Mista
. Emilio Montero Cartelle , 41 , 1961 -1972 .
[本文引用: 1]
[2]
An L Cao T Yu YH 2006 ) Heavy metals contents in Haplocladium and their relationships with Shanghai City environment
. Chinese Journal of Applied Ecology , 17 , 1490 -1494 . (in Chinese with English abstract)
[本文引用: 1]
[安丽 , 曹同 , 俞鹰浩 (2006 ) 上海市小羽藓属植物重金属含量及其与环境的关系
. 应用生态学报 , 17 , 1490 -1494 .]
[本文引用: 1]
[3]
Brown DH Buck GW 2013 ) Cation contents of acrocarpous and pleurocarpous mosses growing on a strontium-rich substratum
. Transactions of the British Bryological Society , 10 , 199 -209 .
[本文引用: 2]
[4]
Chen SF Wu DF Wu JN 1989 ) Using lichen communities as SO2 pollution monitors
. Journal of Nanjing Normal University (Natural Science) , 12 (4 ), 77 -82 . (in Chinese with English abstract)
[本文引用: 1]
[陈舒泛 , 吴鼎福 , 吴继农 (1989 ) 利用地衣群落监测SO2 的污染
. 南京师大学报(自然科学版) , 12 (4 ), 77 -82 .]
[本文引用: 1]
[5]
Christie JM Alomon MS Nozue K Wada M Briggs WR 1999 ) LOV (light, oxygen, or voltage) domains of the blue-light photoreceptor phototropin (nph1): Binding sites for the chromophore flavin mononucleotide
. Proceedings of the National Academy of Sciences, USA , 96 , 8779 -8783 .
[本文引用: 1]
[6]
Cui RP Ma XL Man HS Yao BJ Gu WX Xia QF 1983 ) Hazard of mercury-stained food chains
. Environmental Science , 4 (2 ), 16 -19 . (in Chinese)
[本文引用: 1]
[崔瑞平 , 马秀兰 , 满洪升 , 姚本基 , 顾文兴 , 夏其芳 (1983 ) 汞染食物链的危害
. 环境科学 , 4 (2 ), 16 -19 .]
[本文引用: 1]
[7]
Denayer FO Haluwyn CV Foucault BD Schumacker R Colein P 1999 ) Use of bryological communities as a diagnostic tool of heavy metal soil contamination (Cd, Pb, Zn) in northern France
. Plant Ecology , 140 , 191 -201 .
[本文引用: 1]
[8]
Foan L Domercq M Bermejo R Santamaría JM Simon V 2015 ) Mosses as an integrating tool for monitoring PAH atmospheric deposition: Comparison with total deposition and evaluation of bioconcentration factors. A year-long case-study
. Chemosphere , 119 , 452 -458 .
[本文引用: 1]
[9]
Gao Q 1996 ) Flora Bryophytorum Sinicorum, Vol. 2 . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[高谦 (1996 ) 中国苔藓志第二卷 . 科学出版社 , 北京 .]
[本文引用: 1]
[10]
Gao Q Wu YH 2010 ) Genera Hepaticopsida et Anthocerotopsida Sinicorum . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[高谦 , 吴玉环 (2010 ) 中国苔纲和角苔纲植物属志 . 科学出版社 , 北京 .]
[本文引用: 1]
[11]
Giordano S Adamo P Spagnuolo V Tretiach M Bargagli R 2013 ) Accumulation of airborne trace elements in mosses, lichens and synthetic materials exposed at urban monitoring stations: Towards a harmonisation of the moss-bag technique
. Chemosphere , 90 , 292 -299 .
[本文引用: 1]
[12]
Guo SL Yu J Chen GQ 2015 ) Ecological Data Analyses . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[郭水良 , 于晶 , 陈国奇 (2015 ) 生态学数据分析 . 科学出版社 , 北京 .]
[本文引用: 1]
[13]
Guo Y Wang ZH Zhang ZH 2017 ) Ability of brachymenium exile to monitor metal elements at an electrolytic aluminium factory in karst mountainous area
. Environmental Science & Technology , 40 (3 ), 148 -152 . (in Chinese with English abstract)
[本文引用: 1]
[郭云 , 王智慧 , 张朝晖 (2017 ) 岩溶山区电解铝厂纤枝短月藓对金属元素的监测能力
. 环境科学与技术 , 40 (3 ), 148 -152 .]
[本文引用: 1]
[14]
Hu RL Wang YF 2005 ) Flora Bryophytorum Sinicorum , Vol. 7. Science Press , Beijing . (in Chinese)
[本文引用: 1]
[胡人亮 , 王幼芳 (2005 ) 中国苔藓志第七卷 . 科学出版社 , 北京 .]
[本文引用: 1]
[15]
Li DH 2006 ) Study on correlation between fauna structure and some environmental factors in Shuiba Cave, Guizhou Province
. Carsologica Sinica , 25 , 256 -262 . (in Chinese with English abstract)
[本文引用: 1]
[黎道洪 (2006 ) 贵州水拔洞内动物群落结构与部分环境因子的相关性研究
. 中国岩溶 , 25 , 256 -262 .]
[本文引用: 1]
[16]
Li JY An YG Rong KF 1991 ) Forming and developing process of underground stream caves—Zhijin Cave, etc. as examples
. Journal of Guizhou Institute of Technology , 20 (3 ), 1 -9 . (in Chinese with English abstract)
[本文引用: 1]
[李景阳 , 安裕国 , 戎昆方 (1991 ) 暗河型溶洞的形成和演化过程——以贵州织金洞等为例
. 贵州工学院学报 , 20 (3 ), 1 -9 .]
[本文引用: 1]
[17]
Li XJ 2006 ) Flora Bryophytorum Sinicorum , Vol. 4. Science Press , Beijing . (in Chinese)
[本文引用: 1]
[黎兴江 (2006 ) 中国苔藓志第四卷 . 科学出版社 , 北京 .]
[本文引用: 1]
[18]
Liu R Zhang ZH Shen JC Wang ZH 2018 ) Monitoring of heavy metal concentrations and source apportionment in “24-Curve” Highway region using epiphytic mosses
. Chinese Journal of Ecology , 37 , 1723 -1729 . (in Chinese with English abstract)
[本文引用: 1]
[刘润 , 张朝晖 , 申家琛 , 王智慧 (2018 ) 应用树附生藓类监测晴隆二十四道拐地区重金属污染及来源
. 生态学杂志 , 37 , 1723 -1729 .]
[本文引用: 1]
[19]
Liu RX Wang ZH Zhang ZH 2009 ) Ecological characteristics of bryophyte communities from karst rock desertification peak cluster in Zhenfeng of Guizhou Province
. Bulletin of Botanical Research , 29 , 734 -741 . (in Chinese with English abstract)
[本文引用: 1]
[刘荣相 , 王智慧 , 张朝晖 (2009 ) 贵州贞丰喀斯特石漠峰丛苔藓植物群落生态特征
. 植物研究 , 29 , 734 -741 .]
[本文引用: 1]
[20]
Liu RX Wang ZH Zhang ZH 2011 ) Ecological monitoring of bryophytes for mercury pollution in Danzhai mercury mine area, Guizhou Province, China
. Acta Ecologica Sinica , 31 , 1558 -1566 . (in Chinese with English abstract)
[本文引用: 1]
[刘荣相 , 王智慧 , 张朝晖 (2011 ) 苔藓植物对贵州丹寨汞矿区汞污染的生态监测
. 生态学报 , 31 , 1558 -1566 .]
[本文引用: 1]
[21]
Long CB Zhang ZH 2016 ) Diversity of the bryophytes and their function in monitoring metal contamination in the karst bauxite area in Guangxi
. Journal of Safety & Environment , 16 , 358 -363 . (in Chinese with English abstract)
[本文引用: 1]
[龙朝波 , 张朝晖 (2016 ) 平果岩溶型铝土矿区苔藓多样性及对金属污染的监测指示
. 安全与环境学报 , 16 , 358 -363 .]
[本文引用: 1]
[22]
Mario S Bitetti D 2000 ) The distribution of grooming among female primates: Testing hypotheses with the Shannon- Wiener diversity index
. Behaviour , 137 , 1517 -1540 .
[本文引用: 1]
[23]
Mazur Z Radziemska M Maczuga O Makuch A 2013 ) Heavy metal concentrations in soil and moss (Pleurozium schreberi ) near railroad lines in Olsztyn (Poland)
. Fresenius Environmental Bulletin , 22 , 955 -961 .
[本文引用: 1]
[24]
Peng SL 1987 ) Ecological dominance of the subtropical forest communities in Guangdong
. Acta Ecologica Sinica , 7 , 38 -44 . (in Chinese with English abstract)
[本文引用: 1]
[彭少麟 (1987 ) 广东亚热带森林群落的生态优势度
. 生态学报 , 7 , 38 -44 .]
[本文引用: 1]
[25]
Samecka-Cymerman A Kempers AJ 1998 ) Bioindication of gold by aquatic bryophytes
. Acta Hydrochimica et Hydrobiologica , 26 , 90 -94 .
[本文引用: 1]
[26]
Shah TA Ali A 2010 ) Development of mercury resistant transgenic Nicotiana plants and their environmental impacts
. International Journal of Plant Sciences , 5 , 448 -451 .
[本文引用: 1]
[27]
Shen JC Zhang ZH Wang HH Huang H Wang ZH 2017 ) Water retention capacity of autumn mosses in south stone forest of Guiyang Karst park
. Journal of Ecology & Rural Environment , 33 , 907 -912 . (in Chinese with English abstract)
[本文引用: 1]
[申家琛 , 张朝晖 , 王慧慧 , 黄欢 , 王智慧 (2017 ) 贵阳喀斯特公园南石林秋季藓类植物的持水特性
. 生态与农村环境学报 , 33 , 907 -912 .]
[本文引用: 1]
[28]
State Environmental Protection Bureau 1990 ) Background Value of Soil Environment in China . China Environmental Science Press , Beijing (in Chinese)
[本文引用: 1]
[国家环境保护局 (1990 ) 中国土壤环境背景值 . 中国环境科学出版社 , 北京 .]
[本文引用: 1]
[29]
Tarkowska-Kukuryk M Mieczan T 2014 ) Distribution and environmental determinants of Chironomids (Diptera, Chironomidae) in Sphagnum microhabitats
. Polish Journal of Environmental Studies , 23 , 483 -490 .
[本文引用: 1]
[30]
Toth T Kopernicka M Sabo R Kopernicka T 2016 ) The evaluation of mercury in honey bees and their products from Eastern Slovakia
. Scientific Papers Animal Science & Biotechnologies , 49 , 257 -260 .
[本文引用: 1]
[31]
Wang AY Huang SS Zhong GF Xu GB L ZX Shen XB 2012 ) Effect of cr (VI) stress on growth of three herbaceous plants and their cr uptake
. Environmental Science , 33 , 2028 -2037 . (in Chinese with English abstract)
[王爱云 , 黄姗姗 , 钟国锋 , 徐刚标 , 刘志祥 , 申响保 (2012 ) 铬胁迫对3种草本植物生长及铬积累的影响
. 环境科学 , 33 , 2028 -2037 .]
[32]
Wang CX Li YY Xu SQ 2010 ) Biological monitoring and its application in environmental monitoring
. Asian Journal of Ecotoxicology , 5 , 628 -638 . (in Chinese with English abstract)
[本文引用: 1]
[王春香 , 李媛媛 , 徐顺清 (2010 ) 生物监测及其在环境监测中的应用
. 生态毒理学报 , 5 , 628 -638 .]
[本文引用: 1]
[33]
Wang SQ Zhang ZH Wang ZH 2015 ) Bryophyte communities as biomonitors of environmental factors in the Goujiang karst bauxite, southwestern China
. Science of the Total Environment , 538 , 270 -278 .
[本文引用: 2]
[34]
Wei XP Pu JB Zhao CY 2014 ) Assessment of karst groundwater vulnerability in Chongqing based on revised RISKE model
. Acta Ecologica Sinica , 34 , 589 -596 . (in Chinese with English abstract)
[本文引用: 1]
[魏兴萍 , 蒲俊兵 , 赵纯勇 (2014 ) 基于修正RISKE模型的重庆岩溶地区地下水脆弱性评价
. 生态学报 , 34 , 589 -596 .]
[本文引用: 1]
[35]
Wu PC 2002 ) Flora Bryophytorum Sinicorum , Vol. 6. Science Press , Beijing . (in Chinese)
[本文引用: 1]
[吴鹏程 (2002 ) 中国苔藓志第六卷 . 科学出版社 , 北京 .]
[本文引用: 1]
[36]
Wu PC Jia Y 2004 ) Flora Bryophytorum Sinicorum , Vol. 8. Science Press , Beijing . (in Chinese)
[本文引用: 1]
[吴鹏程 , 贾渝 (2004 ) 中国苔藓志第八卷 . 科学出版社 , 北京 .]
[本文引用: 1]
[37]
Xu CX Li ZZ Li DH 2013 ) Relationships between the diversity of animal communities and the lighting environment and content of heavy metals in soils in Guizhou Zhijin Cave
. Biodiversity Science , 21 , 62 -70 . (in Chinese with English abstract)
[本文引用: 1]
[徐承香 , 李子忠 , 黎道洪 (2013 ) 贵州织金洞洞穴动物群落多样性与光照强度及土壤重金属含量的关系
. 生物多样性 , 21 , 62 -70 .]
[本文引用: 1]
[38]
Xu MQ Cao H 2004 ) Ecological basis and application of PFU protozoan community in bio-monitoring with relation to water quality
. Acta Ecologica Sinica , 24 , 1540 -1547 . (in Chinese with English abstract)
[本文引用: 1]
[许木启 , 曹宏 (2004 ) PFU原生动物群落生物监测的生态学原理与应用
. 生态学报 , 24 , 1540 -1547 .]
[本文引用: 1]
[39]
Xu SC Cao T Yu J Chen Y Song GY 2006 ) Niche of epiphytic bryophytes on trees in Shanghai City
. Chinese Journal of Ecology , 25 , 1338 -1343 . (in Chinese with English abstract)
[本文引用: 1]
[徐晟翀 , 曹同 , 于晶 , 陈怡 , 宋国元 (2006 ) 上海市树附生苔藓植物生态位
. 生态学杂志 , 25 , 1338 -1343 .]
[本文引用: 1]
[40]
Xu YN Ke HL Zhao AN Liu RP Zhang JH 2007 ) Assessment of heavy metals contamination of farmland soils in some gold mining area of Xiaoqinling
. Chinese Journal of Soil Science , 38 , 732 -736 . (in Chinese with English abstract)
[本文引用: 1]
[徐友宁 , 柯海玲 , 赵阿宁 , 刘瑞平 , 张江华 (2007 ) 小秦岭某金矿区农田土壤重金属污染评价
. 土壤通报 , 38 , 732 -736 .]
[本文引用: 1]
[41]
Yang WC Li DH 2010 ) Correlations of heavy metal contents in cave animals and in environment in Dayan Cave of Guangxi
. Chinese Journal of Ecology , 29 , 1554 -1558 . (in Chinese with English abstract)
[本文引用: 1]
[杨卫诚 , 黎道洪 (2010 ) 广西大岩洞洞穴动物体与环境中重金属含量关系
. 生态学杂志 , 29 , 1554 -1558 .]
[本文引用: 1]
[42]
Zhang BK Fu Z Wang QS Yin CL Zhu QQ 1983 ) Soil zinc supply and zinc fertilizer yield increase in Nantong City
. Soils , 15 , 94 -101 . (in Chinese)
[本文引用: 1]
[张炳奎 , 傅棻 , 王岐山 , 尹楚良 , 朱其清 (1983 ) 南通市土壤锌的供给情况及锌肥的增产作用
. 土壤 , 15 , 94 -101 .]
[本文引用: 1]
[43]
Zhang YY 2015 ) Experimental Study on the Effects of Different Light and Iron Concentrations on the Growth of Phytoplankton in Xiaoxingkai Lake . PhD dissertation, Northeast Forestry University , Harbin . (in Chinese with English abstract)
[本文引用: 1]
[张莹莹 (2015 ) 不同光照和铁浓度对小兴凯湖浮游植物生长影响的实验研究 . 博士学位论文, 东北林业大学 , 哈尔滨 .]
[本文引用: 1]
[44]
Zhang ZH Pentecost A 2002 ) The floristic characteristics of the bryophytes and karst depostion in karst caves, England
. Acta Botanica Boreali-Occidentalia Sinica , 22 , 359 -367 . (in Chinese with English abstract)
[本文引用: 2]
[张朝晖 , Pentecost A 2002 ) 英格兰洞穴苔藓植物区系特征及其岩溶沉积研究
. 西北植物学报 , 22 , 359 -367 .]
[本文引用: 2]
[45]
Zhang ZH Wang ZH Zhu A 1996 ) A preliminary study on bryokarst of caves in Huangguoshu area
. Carsologica Sinica , 15 , 224 -232 . (in Chinese with English abstract)
[本文引用: 1]
[张朝晖 , 王智慧 , 祝安 (1996 ) 黄果树喀斯特洞穴群苔藓植物岩溶的初步研究
. 中国岩溶 , 15 , 224 -232 .]
[本文引用: 1]
Variabilidade espacial do Índice de Diversidade de Shannon-Wiener em Floresta Ombrófila Mista
1
2013
... 以盖度代替个体数分别计算研究区域各样点苔藓群落多样性指标(Mario & Bitetti, 2000 ; Amaral et al, 2013 ): ...
上海市小羽藓属植物重金属含量及其与环境的关系
1
2006
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
上海市小羽藓属植物重金属含量及其与环境的关系
1
2006
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
Cation contents of acrocarpous and pleurocarpous mosses growing on a strontium-rich substratum
2
2013
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
... 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
利用地衣群落监测SO2 的污染
1
1989
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
利用地衣群落监测SO2 的污染
1
1989
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
LOV (light, oxygen, or voltage) domains of the blue-light photoreceptor phototropin (nph1): Binding sites for the chromophore flavin mononucleotide
1
1999
... 对洞内苔藓物种进行统计, 共发现苔藓植物种类20科33属49种, 其中苔类8科8属12种, 藓类12科25属37种.优势科为丛藓科、凤尾藓科和提灯藓科, 物种所占比例分别为16%、14%、14%.此外, 实地考察发现, 凤尾藓科群落对生长环境的要求具有很大的相似性, 该科物种在洞内石壁上具有较为广泛的分布, 并且相比生长于洞穴外部的凤尾藓科群落, 生长于洞穴内部的群落具有十分明显的向光性.这与洞穴内部长期受单侧光源照射有关(Christie et al, 1999 ), 洞穴内部环境阴暗、潮湿, 一些低矮的丛集型苔藓植物群落如凤尾藓在洞穴底部无法接触到光照从而退居到养分少但是相对干燥的洞穴石壁上, 以光为环境信号来调整自身的代谢、器官发生和形态建成. ...
汞染食物链的危害
1
1983
... 重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 ).本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 .从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05).其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重.距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大.从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致.Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 ).而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 ).因此, 应重视Hg的污染状况. ...
汞染食物链的危害
1
1983
... 重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 ).本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 .从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05).其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重.距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大.从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致.Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 ).而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 ).因此, 应重视Hg的污染状况. ...
Use of bryological communities as a diagnostic tool of heavy metal soil contamination (Cd, Pb, Zn) in northern France
1
1999
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
Mosses as an integrating tool for monitoring PAH atmospheric deposition: Comparison with total deposition and evaluation of bioconcentration factors. A year-long case-study
1
2015
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
1
1996
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
1996
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2010
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2010
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
Accumulation of airborne trace elements in mosses, lichens and synthetic materials exposed at urban monitoring stations: Towards a harmonisation of the moss-bag technique
1
2013
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
1
2015
... 数据的差异性检验在SPSS 21.0里完成; 多样性数据的统计分析用R语言的Vegan包进行(郭水良等, 2015 ); 图表的绘制采用Excel 2016和Origin 9.0; RDA分析过程在CANOCO 5.0软件中进行. ...
1
2015
... 数据的差异性检验在SPSS 21.0里完成; 多样性数据的统计分析用R语言的Vegan包进行(郭水良等, 2015 ); 图表的绘制采用Excel 2016和Origin 9.0; RDA分析过程在CANOCO 5.0软件中进行. ...
岩溶山区电解铝厂纤枝短月藓对金属元素的监测能力
1
2017
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
岩溶山区电解铝厂纤枝短月藓对金属元素的监测能力
1
2017
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
1
2005
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2005
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
贵州水拔洞内动物群落结构与部分环境因子的相关性研究
1
2006
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
贵州水拔洞内动物群落结构与部分环境因子的相关性研究
1
2006
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
暗河型溶洞的形成和演化过程——以贵州织金洞等为例
1
1991
... 织金洞位于贵州省织金县官寨苗族乡内, 地处26°38°31°°-26°52°35°° N, 105°44°42°°-106°11°38°° E, 是典型的暗河型溶洞.洞内岩溶地下水丰富, 主要发育有织金洞古暗河系统和现代打铁洞暗河系统(李景阳等, 1991 ).全洞长12.1 km, 总面积70多万m2 .洞内地质构造复杂, 平均气温15℃, 相对湿度93%, 植被覆盖率低, 仅入口分布有大量的苔藓植物、蕨类植物和少量的草本、灌木植物.在织金洞旅游局的管理下, 洞内植物很少受到人为破坏.洞内苔藓植物主要分布于从入口至洞内82 m范围内, 出口门坎带也有少量分布.本研究主要在苔藓分布较为集中的入口处开展. ...
暗河型溶洞的形成和演化过程——以贵州织金洞等为例
1
1991
... 织金洞位于贵州省织金县官寨苗族乡内, 地处26°38°31°°-26°52°35°° N, 105°44°42°°-106°11°38°° E, 是典型的暗河型溶洞.洞内岩溶地下水丰富, 主要发育有织金洞古暗河系统和现代打铁洞暗河系统(李景阳等, 1991 ).全洞长12.1 km, 总面积70多万m2 .洞内地质构造复杂, 平均气温15℃, 相对湿度93%, 植被覆盖率低, 仅入口分布有大量的苔藓植物、蕨类植物和少量的草本、灌木植物.在织金洞旅游局的管理下, 洞内植物很少受到人为破坏.洞内苔藓植物主要分布于从入口至洞内82 m范围内, 出口门坎带也有少量分布.本研究主要在苔藓分布较为集中的入口处开展. ...
1
2006
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2006
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
应用树附生藓类监测晴隆二十四道拐地区重金属污染及来源
1
2018
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
应用树附生藓类监测晴隆二十四道拐地区重金属污染及来源
1
2018
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
贵州贞丰喀斯特石漠峰丛苔藓植物群落生态特征
1
2009
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
贵州贞丰喀斯特石漠峰丛苔藓植物群落生态特征
1
2009
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
苔藓植物对贵州丹寨汞矿区汞污染的生态监测
1
2011
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
苔藓植物对贵州丹寨汞矿区汞污染的生态监测
1
2011
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
平果岩溶型铝土矿区苔藓多样性及对金属污染的监测指示
1
2016
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
平果岩溶型铝土矿区苔藓多样性及对金属污染的监测指示
1
2016
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
The distribution of grooming among female primates: Testing hypotheses with the Shannon- Wiener diversity index
1
2000
... 以盖度代替个体数分别计算研究区域各样点苔藓群落多样性指标(Mario & Bitetti, 2000 ; Amaral et al, 2013 ): ...
Heavy metal concentrations in soil and moss (Pleurozium schreberi ) near railroad lines in Olsztyn (Poland)
1
2013
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
广东亚热带森林群落的生态优势度
1
1987
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
广东亚热带森林群落的生态优势度
1
1987
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
Bioindication of gold by aquatic bryophytes
1
1998
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
Development of mercury resistant transgenic Nicotiana plants and their environmental impacts
1
2010
... 重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 ).本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 .从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05).其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重.距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大.从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致.Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 ).而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 ).因此, 应重视Hg的污染状况. ...
贵阳喀斯特公园南石林秋季藓类植物的持水特性
1
2017
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
贵阳喀斯特公园南石林秋季藓类植物的持水特性
1
2017
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
1
1990
... 式中, Ci 为苔藓基质重金属含量实测值, Si 为该重金属的背景标准值.本研究背景标准值采用贵州省土壤背景值(国家环境保护局, 1990 ). ...
1
1990
... 式中, Ci 为苔藓基质重金属含量实测值, Si 为该重金属的背景标准值.本研究背景标准值采用贵州省土壤背景值(国家环境保护局, 1990 ). ...
Distribution and environmental determinants of Chironomids (Diptera, Chironomidae) in Sphagnum microhabitats
1
2014
... 通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 ).第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息.RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量.耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴.光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求.在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 ).在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致.但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处.同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落.这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性.另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性.Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 ). ...
The evaluation of mercury in honey bees and their products from Eastern Slovakia
1
2016
... 岩溶洞穴是一类特殊的自然景观, 极具观赏和游览价值.近年来, 岩溶洞穴资源被不断开发, 人类活动的频繁加剧了洞穴环境的恶化.由于岩溶洞穴的脆弱性和低自净能力, 一旦遭受污染就很难恢复(魏兴萍等, 2014 ).其中, 重金属污染是洞穴环境恶化的主要原因之一, 它不仅会危害环境, 而且会积累在人体中, 阻碍大脑和神经系统发育(Toth et al, 2016 ).通常, 岩溶洞穴与地下水是相通的, 污染物会通过渗透、淋溶等方式危及到地下水.同时, 洞穴重金属污染也会对洞穴生物群落构成威胁, 如: (1)洞穴物种多样性减少; (2)物种繁殖受阻; (3)群落稳定性降低等.因此, 对洞穴重金属进行监测对保护洞穴生物群落具有重要意义. ...
生物监测及其在环境监测中的应用
1
2010
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
生物监测及其在环境监测中的应用
1
2010
... 目前, 利用苔藓植物进行区域监测的研究多集中于测定苔藓植物体内的重金属含量, 从而确定相应环境的污染状况.由于不同苔藓物种对污染的富集能力差异较大, 所以这种监测方法的理想条件是找到同种(郭云等, 2017 ; 刘润等, 2018 )或者同属(安丽等, 2006 )苔藓进行定点监测.而对于矿山、洞穴、石漠化地区这类生存环境较差的小尺度区域而言, 这种监测方式难以推广.首先, 这些地区的苔藓植物生物量较少, 特别是在阴暗潮湿的洞穴环境中, 苔藓植物大多为丛集型, 体型较小且附生于洞穴石壁上, 标本难以采集.其次, 洞穴内部小生境复杂, 多数种类仅在一处或者几处有分布, 使用同一种苔藓监测的可能性较小.第三, 岩溶洞穴具有较高的景观价值, 不易于大范围的开采.因此, 使用苔藓群落学特征对环境污染或变化的反应对环境质量进行监测和评价具有更大的可行性(王春香等, 2010 ). ...
Bryophyte communities as biomonitors of environmental factors in the Goujiang karst bauxite, southwestern China
2
2015
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
基于修正RISKE模型的重庆岩溶地区地下水脆弱性评价
1
2014
... 岩溶洞穴是一类特殊的自然景观, 极具观赏和游览价值.近年来, 岩溶洞穴资源被不断开发, 人类活动的频繁加剧了洞穴环境的恶化.由于岩溶洞穴的脆弱性和低自净能力, 一旦遭受污染就很难恢复(魏兴萍等, 2014 ).其中, 重金属污染是洞穴环境恶化的主要原因之一, 它不仅会危害环境, 而且会积累在人体中, 阻碍大脑和神经系统发育(Toth et al, 2016 ).通常, 岩溶洞穴与地下水是相通的, 污染物会通过渗透、淋溶等方式危及到地下水.同时, 洞穴重金属污染也会对洞穴生物群落构成威胁, 如: (1)洞穴物种多样性减少; (2)物种繁殖受阻; (3)群落稳定性降低等.因此, 对洞穴重金属进行监测对保护洞穴生物群落具有重要意义. ...
基于修正RISKE模型的重庆岩溶地区地下水脆弱性评价
1
2014
... 岩溶洞穴是一类特殊的自然景观, 极具观赏和游览价值.近年来, 岩溶洞穴资源被不断开发, 人类活动的频繁加剧了洞穴环境的恶化.由于岩溶洞穴的脆弱性和低自净能力, 一旦遭受污染就很难恢复(魏兴萍等, 2014 ).其中, 重金属污染是洞穴环境恶化的主要原因之一, 它不仅会危害环境, 而且会积累在人体中, 阻碍大脑和神经系统发育(Toth et al, 2016 ).通常, 岩溶洞穴与地下水是相通的, 污染物会通过渗透、淋溶等方式危及到地下水.同时, 洞穴重金属污染也会对洞穴生物群落构成威胁, 如: (1)洞穴物种多样性减少; (2)物种繁殖受阻; (3)群落稳定性降低等.因此, 对洞穴重金属进行监测对保护洞穴生物群落具有重要意义. ...
1
2002
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2002
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2004
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
1
2004
... 将采回的标本分类整理, 并根据经典形态学分类方法进行鉴定.使用HWG-1型双筒解剖镜以及SMARTe·320型光学显微镜结合《中国苔藓志》第2卷、第4卷、第6-8卷(高谦, 1996 ; 吴鹏程, 2002 ; 吴鹏程和贾渝, 2004 ; 胡人亮和王幼芳, 2005 ; 黎兴江, 2006 )及《中国苔纲和角苔纲植物属志》(高谦和吴玉环, 2010 )等对苔藓植物进行鉴定.苔藓及其基质以样点内所采集样方按四分法混合均匀后自然烘干、磨细、过筛待测.对于采集的苔藓植物样品, 除去多余杂物后用去离子水洗净后烘干、研磨, 每个样品准确称取0.3 g用于消解. ...
贵州织金洞洞穴动物群落多样性与光照强度及土壤重金属含量的关系
1
2013
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
贵州织金洞洞穴动物群落多样性与光照强度及土壤重金属含量的关系
1
2013
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
PFU原生动物群落生物监测的生态学原理与应用
1
2004
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
PFU原生动物群落生物监测的生态学原理与应用
1
2004
... 苔藓群落具有很好的生物监测价值(Denayer et al, 1999 ).群落物种多样性特征可以反应环境污染状况(杨再超等, 2010).环境条件变化会改变动植物群落的结构与功能特征, 污染胁迫使群落的稳定性降低, 物种多样性下降(陈舒泛等, 1989 ; 许木启和曹宏, 2004 ).刘荣相等(2011) 发现苔藓群落结构组成的复杂程度可作为研究区Hg污染监测的指标之一; Wang等(2015) 通过分析苔藓群落和具芽孢苔藓群落的分布规律实现了对岩溶型铝土矿区重金属污染的监测.本文通过对织金洞入口不同距离苔藓群落及其基质重金属含量的研究发现, 苔藓群落特征与其基质Hg含量之间存在良好的线性关系, 通过对苔藓群落丰富度、物种丰富度和物种多样性的数量统计分析可以大致了解对应基质的Hg污染情况. ...
上海市树附生苔藓植物生态位
1
2006
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
上海市树附生苔藓植物生态位
1
2006
... 以相对盖度和相对频度计算苔藓群落的重要值、生态优势度以及生态位宽度(刘荣相等, 2009 ).重要值用于反映群落的地位和作用; 生态优势度是综合群落中各群落重要性, 用以反映群落结构特征的度量值(彭少麟, 1987 ); 生态位宽度是评价群落对资源获取和利用状况的尺度, 生态位宽度越大, 表明对环境的适应能力越强(徐晟翀等, 2006 ). ...
小秦岭某金矿区农田土壤重金属污染评价
1
2007
... 重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 ).本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 .从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05).其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重.距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大.从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致.Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 ).而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 ).因此, 应重视Hg的污染状况. ...
小秦岭某金矿区农田土壤重金属污染评价
1
2007
... 重金属污染超标倍数能够直接反映基质的污染程度(徐友宁等, 2007 ).本研究以同一样点的平均值表示织金洞距入口不同深度处基质重金属含量, 结果见表3 .从表中可以看出, 3种重金属含量在距入口不同距离均表现出显著性差异(P < 0.05).其中, 距洞口深度40 m处重金属Cr、Hg和Zn的污染超标倍数都达到了最大值, 分别为1.80、59.45和0.86, 说明该段重金属污染较为严重.距洞口深度40 m处位于织金洞入口大厅中段, 坡度最小(表1 ), 上述元素受风化、淋溶的影响逐渐向坡度平缓地段聚集的可能性较大.从表3 中可以看出Hg污染超标倍数在距洞口不同深度均远高于其他重金属, 说明洞内Hg污染较为严重, 与徐承香等(2013)的研究结果一致.Hg是毒性最强的重金属之一, 对水体和植物生长有着十分不利的影响(Shah & Ali, 2010 ).而食物链的转移和富集, 会大大增加Hg对人体健康的危害(崔瑞平等, 1983 ).因此, 应重视Hg的污染状况. ...
广西大岩洞洞穴动物体与环境中重金属含量关系
1
2010
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
广西大岩洞洞穴动物体与环境中重金属含量关系
1
2010
... 目前, 已有一些关于用动物群落监测洞穴重金属污染的研究(杨卫诚和黎道洪, 2010 ; 徐承香等, 2013 ), 而用植物群落监测洞穴环境的研究在国内还鲜有报道.相比于动物群落, 植物群落具有更加稳定的优势, 可以对环境变化实行定点监测, 采样和监测工作也更加简单、便捷.一些植物类群如苔藓, 对环境变化具有较高的敏感性, 常常被作为环境监测的生物指标(Giordano et al, 2013 ; Foan et al, 2015 ).不仅如此, 苔藓植物还对恶劣环境有很强的适应性, 在一些植物覆盖率低、污染严重的区域如矿区(龙朝波和张朝晖, 2016 )、石漠化地区(申家琛等, 2017 )、道路(Mazur et al, 2013 )等环境中也常用苔藓植物作为生态监测材料. ...
南通市土壤锌的供给情况及锌肥的增产作用
1
1983
... 通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 ).第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息.RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量.耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴.光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求.在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 ).在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致.但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处.同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落.这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性.另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性.Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 ). ...
南通市土壤锌的供给情况及锌肥的增产作用
1
1983
... 通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 ).第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息.RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量.耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴.光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求.在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 ).在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致.但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处.同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落.这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性.另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性.Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 ). ...
1
2015
... 通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 ).第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息.RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量.耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴.光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求.在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 ).在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致.但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处.同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落.这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性.另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性.Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 ). ...
1
2015
... 通过环境变量筛选出具有显著解释量的环境因子与苔藓优势群落进行冗余分析(RDA), 环境因子的显著性通过999次置换检验来确定(Tarkowska- Kukuryk & Mieczan, 2014 ).第一排序轴的特征值为0.78, 解释了78%的苔藓群落对基质重金属污染和环境适应特征, 反映了环境-基质重金属-群落的大部分信息.RDA结果显示, 光照、矿质元素Ca、坡度和重金属Hg是影响织金洞内苔藓群落分布的主要变量.耐重金属污染的群落位于第一排序轴正半轴, 适钙苔藓群落位于负半轴.光照因子对苔藓群落的影响主要表现为适钙苔藓群落对低光度的适应性, 以及耐重金属污染群落对光照的需求.在低光照的条件下可能会激发某些植物对重金属元素的需求(张莹莹, 2015 ).在排序图中, 大部分苔藓群落位于重金属Hg连线的相反方向, 说明苔藓群落分布总体上与其基质Hg含量呈负相关关系, 与线性拟合结果一致.但是, 与其他苔藓群落不同的是蛇苔群落(缩写Con.con ), 它与污染较为严重的Hg元素表现出了极大的相关性, 且其主要分布在污染严重超标的距入口40-20 m处.同时, 我们对织金洞出口分布有苔藓群落的基质重金属进行测定后发现, 在Hg含量高出背景标准值22倍的地段(距出口32 m)仅分布有由单一物种组成的蛇苔群落.这在一定程度上说明了蛇苔群落对Hg污染有良好的耐受性.另外, 对洞内环境具有较强生态适应范围的尖叶美喙藓群落(缩写Eur.eus )与元素Zn的连线最短, 相关性最大, 说明基质Zn含量与尖叶美喙藓群落在洞内的广泛分布可能存在一定相关性.Zn是植物生长发育所必需的微量营养元素之一, 适度的锌对植物生长有促进作用(张炳奎等, 1983 ). ...
英格兰洞穴苔藓植物区系特征及其岩溶沉积研究
2
2002
... 于2017年9月, 在织金洞距入口不同深度(80 m、60 m、40 m、20 m、0 m)设置5个5 m × 5 m样点, 每个样点按照洞内环境特征随机设定10 cm × 10 cm重复样方数个, 样点信息见表1 .此外, 按照实际情况在出口分布有苔藓植物的地方设置2个样点共10个样方, 以补充织金洞苔藓物种数据.采集样方内全部苔藓和部分基质, 将每个样方内的全部苔藓记为一个群落, 利用优势种原则为群落命名(张朝晖和Pentecost, 2002 ).采集时, 每个样点均用数字照度计(PM6612L, 华谊联表)测定光照强度; 手持式空气温湿度计(HT-635, 广州宏诚科技)测定洞内空气温湿度; 手持式激光测距仪(SW-1500A, 深达威)测量距洞口距离.样方内每项指标测定3次, 取平均值.详细记录每个样地的海拔、坡度、坡向、盖度等详细生境信息, 共采集苔藓及基质样品各50份. ...
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
英格兰洞穴苔藓植物区系特征及其岩溶沉积研究
2
2002
... 于2017年9月, 在织金洞距入口不同深度(80 m、60 m、40 m、20 m、0 m)设置5个5 m × 5 m样点, 每个样点按照洞内环境特征随机设定10 cm × 10 cm重复样方数个, 样点信息见表1 .此外, 按照实际情况在出口分布有苔藓植物的地方设置2个样点共10个样方, 以补充织金洞苔藓物种数据.采集样方内全部苔藓和部分基质, 将每个样方内的全部苔藓记为一个群落, 利用优势种原则为群落命名(张朝晖和Pentecost, 2002 ).采集时, 每个样点均用数字照度计(PM6612L, 华谊联表)测定光照强度; 手持式空气温湿度计(HT-635, 广州宏诚科技)测定洞内空气温湿度; 手持式激光测距仪(SW-1500A, 深达威)测量距洞口距离.样方内每项指标测定3次, 取平均值.详细记录每个样地的海拔、坡度、坡向、盖度等详细生境信息, 共采集苔藓及基质样品各50份. ...
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
黄果树喀斯特洞穴群苔藓植物岩溶的初步研究
1
1996
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
黄果树喀斯特洞穴群苔藓植物岩溶的初步研究
1
1996
... 蛇苔是岩溶洞穴中一类常见的苔类植物, 多生于潮湿碎石或土上, 阴湿的洞穴环境是其主要栖息地, 在很多相关研究中均有报道(张朝晖等, 1996 ; 张朝晖和Pentecost, 2002 ; 黎道洪, 2006 ).蛇苔与其基质Hg污染具有一定相关性, 一方面与它对汞的耐受性有关, 另一方面, 其自身生长可能对基质汞含量有特殊的要求.Brown和Buck (2013) 的研究表明, 一种管口苔属苔类Solenostoma crenulatum 在基质富含铜的情况下能够受到更好的被刺激从而提高光合作用效率(Brown & Buck, 2013 ).在光照很弱的洞穴环境中, 植物通常无法进行正常的光合作用, 为了生存, 某些苔藓群落可能会进化出与环境相匹配的独特生理特征如向光性运动、喜钙生基质、耐重金属生理等.然而, 并不是所有的苔藓群落都能很好地适应像Hg这类高毒性系数的重金属.当基质重金属含量富集到一定量时, 一些群落就很难再从别的生境迁入, 最后只剩下对基质含量有较高耐受性的群落.这也是为什么能够通过水生苔藓对Au的富集找到金矿, 通过铜藓(Meilichoferia elongata )找到铜矿(Samecka-Cymerman & Kempers, 1998 ).因此, 苔藓群落在不同环境梯度下呈现出的特有甚至专一的分布规律对于环境因子有重要的指示作用(Wang et al, 2015 ), 利用这种指示作用可以更加直观、长期、有效地对区域环境进行监测. ...
 , 申家琛
, 申家琛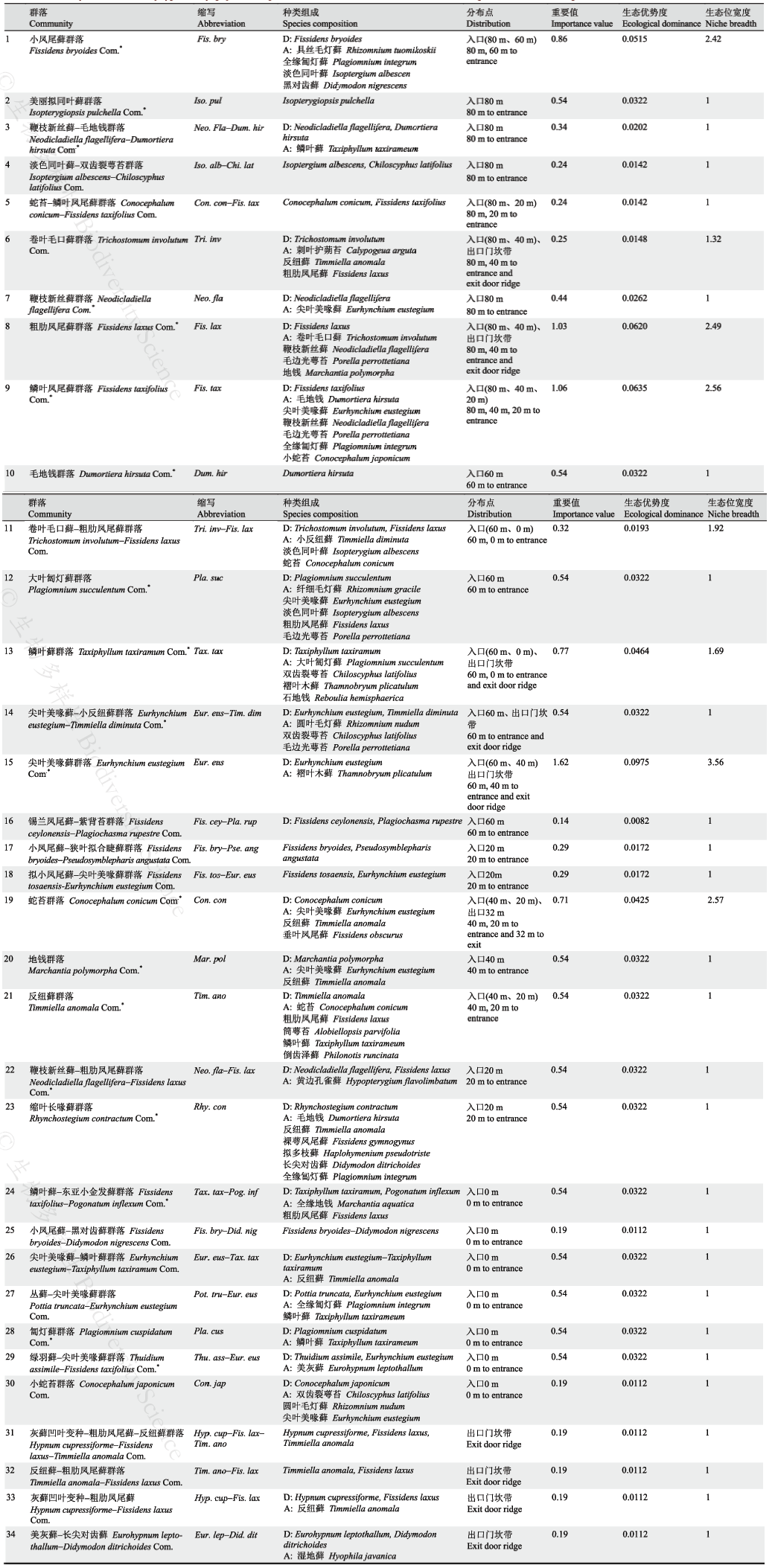


{kind=link}
{kind=link}
{kind=link}
{kind=link}