郭志文, 郑景明
北京林业大学林学院, 北京 100083
Zhiwen Guo, Jingming Zheng
通讯作者:
责任编辑:
收稿日期: 2017-01-17
接受日期: 2017-06-9
网络出版日期: 2017-09-20
版权声明: 2017 《生物多样性》编辑部 版权所有.《生物多样性》编辑部
基金资助:
展开
摘要
种子扩散方式对植物物种分布、种群动态及群落组成都有重要影响, 但目前有关种子扩散方式的数据还很欠缺。植物的生活史性状与种子扩散方式联系密切, 通过植物生活史性状预测种子的扩散方式是一种有效的研究手段。本文基于我国360种植物的生长型、株高、种子质量和果实类型以及种子扩散方式的数据集, 随机抽取288个物种数据(80%)作为训练样本, 采用神经网络、决策树、费舍尔线性判别和支持向量机算法, 分别建立种子扩散方式的预测模型, 将其余72个物种数据(20%)用于模型检验。以1,000次随机抽样后的平均判别正确率作为模型预测效果的评价指标。结果表明: 用生长型、株高、种子质量及果实类型作为主要预测变量, 构建的神经网络、决策树、费希尔线性判别和支持向量机模型均能达到较好的预测效果, 准确率分别为78.90%、77.09%、77.81%和78.14%, 其中以神经网络模型的预测效果最好。进一步研究发现, 神经网络模型对动物扩散、无助力扩散和风扩散的预测效果分别为81.32%、74.90%和81.45%。本研究为植物种子扩散方式预测提供了一种新的思路。
关键词:
Abstract
Mode of seed dispersal is an important trait for understanding geographical distributions, population dynamics, and community composition of plant species. However, data of dispersal modes are scarce for Chinese plant species. Previous studies have shown that growth form, plant height, fruit type, and seed mass have strong correlations with seed dispersal modes, thus predictions using modelling could be an alternative to gain this information. We collected information on growth forms, plant height, fruit types, seed mass, and dispersal modes from 360 kinds of Chinese angiosperm plants, and built a neural network model (NNET), decision tree (TREE), Fisher linear discriminant model (LDA), and support vector machine model (SVM) to predict seed syndromes from these four traits. For each model, an 80% sample (288 species) was randomly drawn from dataset as the training sample, with remaining 20% of data was used as a test sample. Results showed that all four models achieved rather good predictions, and the average total correctness rate for the NNET, TREE, LDA, and SVM was 78.90%, 77.09%, 77.81%, 78.14%, respectively. The neural network model had the highest correctness rates for different dispersal modes, i.e., zoochory (81.32%), autochory (74.90%), and anemochory (81.45%). This paper establishes the basis for the prediction of seed dispersal modes.
Keywords:
种子扩散是植物生活史的重要阶段, 营固着生活的植物都要通过种子扩散来扩大种群和提高幼苗成活率(Willson, 1993; Jordano et al, 2011)。种子扩散的时空格局决定了植物群落的分布格局(Clark et al, 1998; 于顺利等, 2007; 姚蓓等, 2015)及地理分布范围(Cain et al, 1998, 2000; Neubert & Caswell, 2000; Hewitt & Kellman, 2002; 郑景明等, 2004; Thuiller et al, 2008), 且与种子扩散方式关系密切, 但目前国内外的种子扩散方式数据还明显不足。建立种子扩散方式预测模型能够获得大量的植物种子扩散方式数据, 有助于今后开展相关生态学研究。
种子扩散方式与植物的生长型、株高、果实类型和种子质量等生活史性状密切相关(Westoby et al, 1992; Thomson et al, 2010)。在以木本植物占优势(尤其是乔木居多)的森林中, 动物扩散的比例通常较高(李娟等, 2013)。种子大小与扩散方式也密切相关, 动物协助扩散的种子质量比风扩散和无助力扩散的种子大(Westoby et al, 1990, 1996; Leishman & Westoby, 1994; 张大勇, 2004), 扩散方式能够解释种子质量变异的20-29% (Leishman et al, 1995; Moles et al, 2005)。株高对于无助力扩散和风扩散种子的扩散效率有显著影响, 高大植物更有利于无助力扩散和风扩散种子形成有效的扩散距离(Thompson & Rabinowitz, 1989; 于顺利等, 2007; Thomson et al, 2011; Li et al, 2012)。
以上研究表明通过植物生活史性状来预测种子的扩散方式是可行的(Hughes et al, 1994; Thomson et al, 2010)。本文利用360种植物的生长型、株高、果实类型和种子质量4个生活史性状以及扩散方式数据(附录1), 在分析物种生活史性状与种子扩散方式相关性的基础上, 以这4个生活史性状数据为主要预测变量, 采用神经网络(neural network)、决策树(decision tree)、费舍尔线性判别(Fisher linear discriminant)和支持向量机(support vector machine)算法分别建立种子扩散方式预测模型, 试图找到一种较为有效的判别方法, 为植物种子扩散方式研究提供一定的参考。
种子扩散方式数据来源于国内已发表的文献(王桔红等, 2009; 李娟等, 2013; 雷霄等, 2015)及英国邱园皇家植物园种子数据库(http://data.kew.org/ sid/), 生长型、株高、果实类型数据来源于《中国植物志》(http://frps.eflora.cn/), 种子质量数据来源于中国西南野生生物种质资源库(http://www.genobank.org/)。我们建立了一个具有500个物种的初始数据集, 由于其中水扩散的物种所占比例较小, 在本研究中未予以考虑。为了确保在随机抽样时每种扩散方式的物种被抽取的概率相同, 需保证用于建模的动物扩散、无助力扩散和风扩散方式的样本数相同。由于在500个物种中风扩散的仅有120种, 因此在建模时我们对其他2种传播方式也各随机抽取了120种, 最后得到一个包括360个物种的数据集, 这些物种涵盖了55科160属, 乔、灌、草、藤所占比例分别是26%、16%、54%和4%。
由于生长型、果实类型、种子扩散方式都是质量性状, 而株高和种子质量为数量性状, 为了将数据类型统一化, 我们将每个生活史性状划分成若干个分类水平(表1)。统一化后的5个植物生活史性状为分类变量, 用Gamma等级相关方法(用两个变量之间同序对和异序对数量的差异反映变量间的相关程度)进行相关分析,并检验生长型、株高、果实类型、种子质量与扩散方式这5个变量之间的相关性, 为下一步的模型建立提供依据。
表1 5个植物生活史性状的分类水平及划分方法
Table 1 The protocol of classification level in five life history traits of plant
| 生活史性状 Life history trait | 分类水平 Class level | 参考文献 Reference |
|---|---|---|
| 生长型 Growth form | 草本; 灌木; 藤本; 乔木 Herb; Shrub; Vine; Tree | Flora Reipublicae Popularis Sinicae, 1979 |
| 株高 Plant height | < 0.5 m; 0.5-2.0 m; 2.0-5.0 m; > 5.0 m | Westoby et al, 1990 |
| 果实类型 Fruit type | 裂果; 闭果; 肉质果 Dehisce; Achene; Fruit | Flora Reipublicae Popularis Sinicae, 1979 |
| 种子质量 Seed mass | < 0.1 mg; 0.1-100 mg; > 100 mg | Westoby et al, 1996 |
| 扩散方式 Dispersal mode | 无助力扩散; 风扩散; 动物扩散 Autochory; Anemochory; Zoochory | Baskin & Baskin, 1998,2014 |
为了建立有效的预测模型, 本研究分别采用了神经网络(刘明同, 2001)、决策树(刘明同, 2001; 唐华松和姚耀文, 2001)、费希尔判别( 赵丽娜 (2013) Fisher判别法的研究及应用. 硕士学位论文, 东北林业大学, 哈尔滨.)和支持向量机( Cortes, 1995; 刘明同等, 2001) 4种算法进行建模。将360个物种的5个生活史性状进行[0, 1]二进制赋值后, 从中随机抽取288个物种数据(80%)作为训练样本建立判别模型, 其中生长型、株高、果实类型和种子质量4个植物生活史性状作为自变量, 种子扩散方式为因变量, 再用剩余的72个物种(20%)作为检测样本对模型进行检验, 并分析各判别模型对扩散方式的预测能力。以1,000次随机抽样建模和检验的总体平均正确率作为各模型的预测效果评价指标, 得到最佳模型后进一步分析该模型对各扩散方式的预测正确率。
统计分析均在R 3.2.3中进行, 神经网络模型用nnet软件包, 决策树模型用tree软件包, 费希尔线性判别模型用MASS软件包, 支持向量机模型用e1071软件包。
通过分析生长型、株高、果实类型、种子质量及扩散方式间的相关性, 发现这5个生活史性状间均有显著的相关关系(P < 0.01) (表2)。其中生长型、株高、果实类型、种子质量与扩散方式间都显著相关(P < 0.01), 说明这4个生活史性状能够用于预测植物种子的扩散方式。
表2 5个植物生活史性状间的相关性
Table 2 Correlations between five life history traits of plants
| Gamma 系数 Gamma index | 生长型 Growth form | 株高 Plant height | 果实类型 Fruit type | 种子质量 Seed mass |
|---|---|---|---|---|
| 株高 Plant height | 0.96** | |||
| 果实类型 Fruit type | 0.55** | 0.47** | ||
| 种子质量 Seed mass | 0.69** | 0.64** | 0.49** | |
| 扩散方式 Dispersal mode | 0.70** | 0.63** | 0.87** | 0.50** |
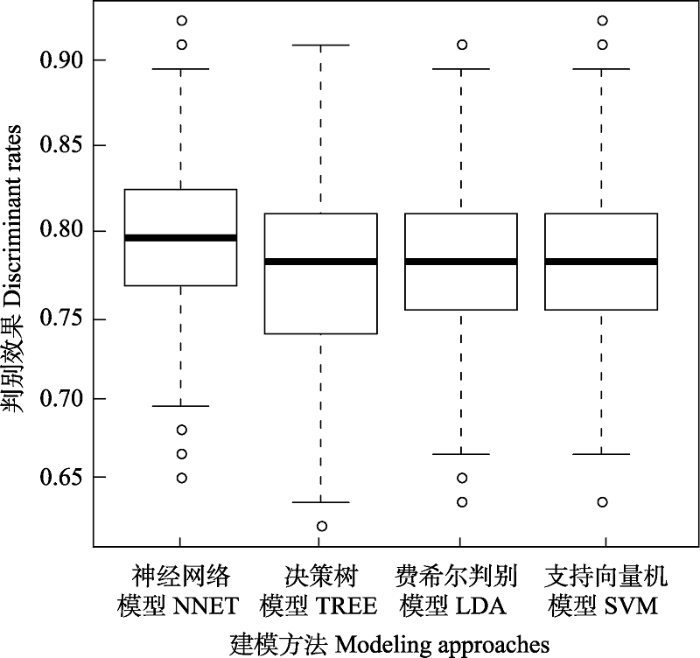
通过1,000次随机抽样建模和检验发现, 4种模型对种子扩散方式的预测效果分别是78.90%、77.09%、77.81%和78.14%, 且以神经网络模型的预测效果最好(变异系数为5.37%)。虽然决策树、费舍尔线性判别和支持向量机模型也能达到较好的预测效果, 但预测效果的变异系数均高于神经网络模型(图1)。
图1 4种模型对种子扩散方式的判别效果
Fig. 1 The discriminant rates of four models to seed dispersal modes. NNET, Neural network model; TREE, Decision tree model; LDA, Fisher linear discriminant model; SVM, Support vector machine model.
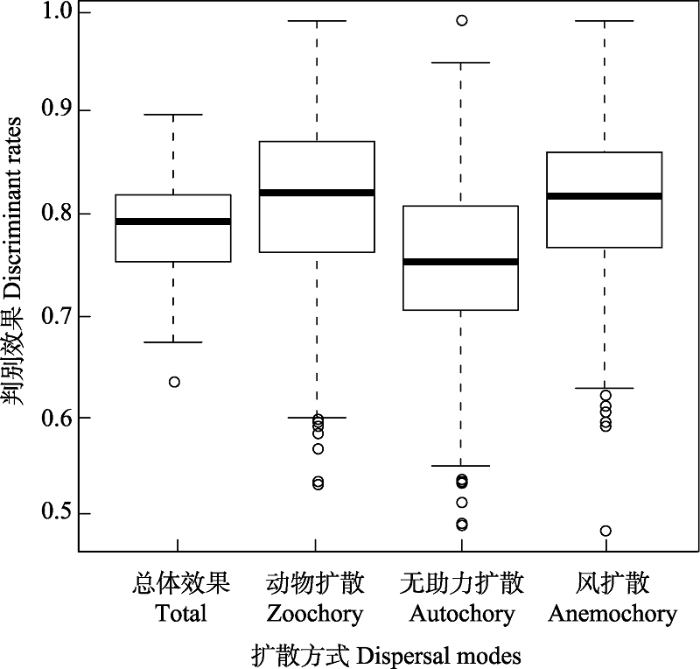
对神经网络模型的预测结果进行进一步分析发现, 该模型对动物扩散、无助力扩散和风扩散的预测效果都较好(图2), 分别是81.32%、74.90%和81.45%。
图2 神经网络模型对3种扩散方式的判别效果
Fig. 2 The discriminant rates of neural network model to three dispersal modes
本研究表明, 采用植物性状预测种子的传播方式是可行的, 但采用合适的方法建立通用预测模型存在一定的挑战。Hughes等(1994)在对有关种子扩散方式的假说进行验证时指出, 用植物生活史性状预测种子的扩散方式是种子生态学研究的重要手段。植物各性状之间存在高度的相关性, 传统的线性模型预测效果不佳, 而非线性的判别方法能够很好地解决高度相关变量的分类问题。
本研究以生长型、株高、果实类型和种子质量4个性状为预测变量, 采用神经网络、决策树、费希尔线性判别和支持向量机4种算法构建模型, 并对模型的预测效果进行了检验, 结果显示这4种模型对种子扩散方式的预测都有很好的效果。其中神经网络模型的预测效果最好, 也最稳定, 可以作为今后种子扩散方式预测的重要方法。4种模型的预测效果差异主要决定于各模型的算法特点, 决策树模型虽然可以处理非数值型的数据, 但由于本研究中各变量有多个属性值, 导致该模型的预测能力较差。支持向量机模型和费希尔线性判别模型的预测效果稍好, 其算法原理都是将多维变量映射到新的空间中, 以实现样本分类的目的, 但它们都只对二类分类问题有较好的效果, 对多类分类的预测效果也受到影响。而神经网络模型在建模过程中能够根据输出值与实际值的误差对模型各参数进行调整, 模型的学习反馈能力较强, 所以预测效果较好。
本文所建立的预测模型对有单一扩散方式的物种具有较强的预测能力, 而对有多种扩散方式的物种的预测效果稍差。以神经网络模型为例, 虽然该模型总体预测效果较好, 但对无助力扩散的预测效果低于平均水平(图2)。这主要是因为种子质量小于10 mg、果实为闭果的物种通常被认为通过无助力扩散方式散布种子, 但由于其果实或种子具有特殊结构, 它们也能够进行风扩散(Willson et al, 1990), 这些情况主要出现在菊科、罂粟科、毛茛科、苋科和石竹科植物中。一些没有明显附属结构的种子通常被认为是无助力扩散, 但事实上这些物种可能是通过风扩散、动物扩散或者水扩散的(Thomson et al, 2010)。如萹蓄(Polygonum aviculare)可进行无助力扩散和风扩散(van der Pijl, 1982), 橡胶树(Hevea brasiliensis)可进行无助力扩散和水扩散(van der Pijl, 1982; Turner, 2001; 周会平等, 2014), 但模型对这些能够通过多种方式扩散种子的物种预测能力较低。
除了对无助力扩散方式存在较高的误判率之外, 神经网络模型对风扩散和动物扩散方式的预测也存在一定的误判, 部分原因也是单个物种的种子具有多种扩散方式。一些能够进行动物扩散的植物种子也能进行风扩散, 如大狼杷草(Bidens frondosa)和鬼针草(B. pilosa)等物种在模型中容易被判定为风扩散(Mori & Brown, 1998)。而有些依赖动物扩散的物种, 如落葵(Basella alba)和木通(Akebia quinata)等也能进行无助力扩散。将这些对模型预测效果具有显著影响的物种剔除后建立的判别模型, 预测准确率可达89.87%, 证实了用于建模的数据质量会影响模型预测效果。
为了提高每一种扩散方式的预测准确率, 可以对每种扩散方式分别建立预测模型。如Thomson等(2010)以澳大利亚悉尼周边区域的1,176种植物为研究对象, 采用植被类型、生长型、株高、种子库类型、种子质量、生活周期和生物类群等植物生活史性状和生态学属性作为变量, 分别对每类种子扩散方式构建线性混合效应模型, 然后用其他地区的数据对模型进行检验, 结果显示这些模型预测的最高正确率为70%。但Thomson等(2010)也指出, 针对单一传播方式的预测模型会受到植被类型的影响, 其结果在外推到其他植被类型时会受到制约。在本研究中我们同时对3种扩散方式进行了预测, 后续将针对每一种传播方式作进一步的预测模型研究。
此外, 建立模型时引入更多的预测变量也可以改善预测效果, 如种子库类型、植被类型、冠幅等(Hughes et al, 1994)。本研究中我们仅选用了4个容易获取的植物生活史性状, 对于其他与种子扩散方式相关的性状和生态学属性, 由于数据的匮乏而在构建模型时并未包括在内, 使得预测变量所含信息存在一定程度的不足, 可能对模型的预测效果有一定影响。在今后的研究中可将种子库类型、生活史周期等植物生活史性状及生态属性考虑在内。
总之, 种子扩散方式是一个重要的植物生活史性状, 通过已有的生活史性状对其进行预测是今后生态学研究的一个重要方向。本文用神经网络、决策树、费希尔线性判别和支持向量机模型对种子扩散方式的预测取得了较好效果, 可以作为今后种子扩散方式预测的新方法。
Appendix 1 Checklist of 360 species used in modeling in this study
http://www.biodiversity-science.net/fileup/PDF/2017019-1.pdf
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
Seed dispersal and the Holocene migration of woodland herbs .https://doi.org/10.1890/0012-9615(1998)068[0325:SDATHM]2.0.CO;2 URL [本文引用: 1] 摘要
The distribution of many woodland herbs extends 1000-2000 km in a north-south direction, yet the majority of these species grow clonally, have little recruitment by seed, and possess no obvious mechanism for long-distance seed dispersal. Although aware that woodland herbs disperse poorly, ecologists have tacitly assumed that, given long periods of time, even small dispersal distances would allow woodland herbs to colonize the vast geographic region they now occupy. We examined this assumption for the understory herb Asarum canadense. To estimate long-term rates of spread by seed, we calibrated seed-dispersal diffusion models with life history data and with data on seed carries by ants. We supplemented our field observations and modeling results for A. canadense with a literature survey on the dispersal capabilities of other plant species. Ants transported A. canadense seeds up to 35 m, the largest distance ants are known to move the seeds of any woodland herb. Empirically calibrated diffusion models indicated that over the last 16 000 yr A. canadense should only have traveled 10-11 km from its glacial refugia. In actuality, A. canadense moved hundreds of kilometers during this time. Models that examined the tail of A. canadense's seed-dispersal curve indicated that occasional dispersal events had to have a high frequency ($\ge$0.001 on a per seed basis) and a large magnitude (dispersal distance >1 km) for A. canadense to have traveled over 200 km in 16 000 yr. The literature survey showed that most woodland herbs and many other forest, desert, coastal, and open-habitat plant species have limited seed-dispersal capabilities, similar to those in A. canadense. We conclude that woodland herbs, as well as many other plants, disperse so slowly that there is no documented mechanism by which most of these species could have reached their present geographical range since the last glacial maximum. This suggests that occasional events leading to long-distance dispersal dominate the Holocene colonization of northern temperate forest by woodland herbs, and this, in turn, has implications for issues ranging from the importance of genetic analyses to the structure of metapopulation models.
|
| [4] |
Long-distance seed dispersal in plant populations .https://doi.org/10.2307/2656714 URL PMID: 10991892 [本文引用: 1] 摘要
Long-distance seed dispersal influences many key aspects of the biology of plants, including spread of invasive species, metapopulation dynamics, and diversity and dynamics in plant communities. However, because long-distance seed dispersal is inherently hard to measure, there are few data sets that characterize the tails of seed dispersal curves. This paper is structured around two lines of argument. First, we argue that long-distance seed dispersal is of critical importance and, hence, that we must collect better data from the tails of seed dispersal curves. To make the case for the importance of long-distance seed dispersal, we review existing data and models of long-distance seed dispersal, focusing on situations in which seeds that travel long distances have a critical impact (colonization of islands, Holocene migrations, response to global change, metapopulation biology). Second, we argue that genetic methods provide a broadly applicable way to monitor long-distance seed dispersal; to place this argument in context, we review genetic estimates of plant migration rates. At present, several promising genetic approaches for estimating long-distance seed dispersal are under active development, including assignment methods, likelihood methods, genealogical methods, and genealogical/demographic methods. We close the paper by discussing important but as yet largely unexplored areas for future research.
|
| [5] |
Stages and spatial scales of recruitment limitation in southern Appalachian forests . |
| [6] |
Support-vector networks . |
| [7] |
|
| [8] |
Tree seed dispersal among forest fragments. II. Dispersal abilities and biogeographical controls .https://doi.org/10.1046/j.1365-2699.2002.00679.x URL [本文引用: 1] 摘要
Abstract Aim To investigate the medium to long-distance dispersal abilities of temperate hardwood tree species and the ecological controls on dispersal, including distance to and connectivity with seed sources, seed source strength, and species dispersal mechanism and seed size. Location A fragmented forest system in the Long Point region, Southern Ontario. Methods Pine plantations were the `seed traps' in which seedlings of hardwood species were enumerated to indicate past dispersal events. The influence of distance to seed sources, dispersal mechanism and seed mass on the probability of dispersal to plantations were evaluated using logistic analysis. Regression analysis was used to determine the effect of seed source strength (mature tree abundance within 150m of plantations) on seedling density in plantations. Connectivity was assessed by comparing the strength of correlations between seedling abundance in plantations and the abundance of mature trees around plantations in connected vs. unconnected source areas. Results Seedling presence in plantations decreased significantly with distance from the nearest potential seed source for species grouped according to dispersal mechanism. Probabilities of seedling presence were $\ge$0.8 at 25m distances, decreasing to under 0.3 at 175m distances. While twenty-seven of twenty-nine species were present in at least one plantation with a seed source within 25m, only thirteen of twenty-seven species occurred in plantations with a seed source $\ge$100m away, and only nine of twenty-four species in plantations $\ge$150m from seed sources, indicating limited potentials for interfragment migration. Seed source strength was significantly related to seedling density in plantations for twelve of fifteen species tested, indicating the importance of species commonness to interfragment migration. Connectivity was not related to dispersal frequency in the system, but this finding applies to a relatively well-connected system of forests and plantations and is expected to differ for systems with greater patch isolation. In addition to these general controls, dispersal was related to species dispersal mechanism. Seed size was negatively related to dispersal frequency, but only within the rodent dispersal spectra. Bird-dispersed species appeared to have superior interfragment dispersal abilities, closely followed by lighter seeded rodent- and wind-dispersed species. Large-seeded rodent-dispersed species ( Juglans spp., Quercus macrocarpa ) and species lacking well-developed adaptations for dispersers were infrequent in plantations generally, and with the exception of Q. macrocarpa , were absent from plantations more than 50m from seed sources. Species dispersal abilities were ranked according to dispersal mechanism and seed size such that: bird > lighter-seeded rodent = wind > larger-seeded rodent = unspecialized. Main conclusions Distance to seed sources appears to be a key determinant of patch colonization. Infrequent dispersal over distances of >100-150m for most species in this system raises concerns about the abilities of tree species to be sustained in fragmented forests. For some species, distances of as little as 50m appear to be isolating, and these, in particular, risk regional extinctions over time scales depending on their local population persistence. Artificial introductions may be needed to maintain fragmented tree populations, particularly for rare species and those with poor dispersal indicated in this study.
|
| [9] |
Predicting dispersal spectra: a minimal set of hypotheses based on plant attributes .https://doi.org/10.2307/2261456 URL [本文引用: 2] |
| [10] |
Frugivores and seed dispersal: mechanisms and consequences for biodiversity of a key ecological interaction .https://doi.org/10.1098/rsbl.2010.0986 URL PMID: 3097856 [本文引用: 1] 摘要
The 5th Symposium on Frugivores and Seed Dispersal, held in Montpellier (France), 13-18 June 2010, brought together more than 220 researchers exemplifying a wide diversity of approaches to the study of frugivory and dispersal of seeds. Following Ted Fleming and Alejandro Estrada's initiative in 1985, this event was a celebration of the 25th anniversary of the first meeting in Veracruz, Mexico. Frugivory and seed dispersal are active research areas that have diversified in multiple directions since 1985 to include evolution (e.g. phylogenetic diversity and dispersal adaptations), physiology (e.g. sensory cues and digestion), landscape ecology (movement patterns), molecular ecology (e.g. gene flow, genetic diversity and structure), community ecology (e.g. mutualistic interaction networks) and conservation biology (effects of hunting, fragmentation, invasion and extinction), among others. This meeting provided an opportunity to assess conceptual and methodological progress, to present ever more sophisticated insights into frugivory in animals and dispersal patterns in plants, and to report the advances made in examining the mechanisms and consequences of seed dispersal for plants and frugivores.
|
| [11] |
Character of seed rain of species with different dispersal modes in Tiantong evergreen broad-leaved forest, Zhejiang Province .https://doi.org/10.3969/j.issn.1000-5641.2015.02.015 URL [本文引用: 1] 摘要
为了研究不同传播方式物种种子雨的基本特征,以浙江天童常绿阔叶林20 hm2动态监测样地的种子雨为研究对象,分析了种子雨的密度、物种组成以及种子限制的强度.结果表明:(1)种子雨收集器共收集到种子136 38粒,隶属于28科66种,种子雨密度为146粒/m2;(2)样地内有56%的物种未收集到种子,因此种子雨与可繁殖个体物种组成的相似性较低且受不同传播方式影响,风力传播的相似性系数极值(Rmax)最大且距种子收集器距离(d)最远(风力传播:Rmax=0.226,d=20 m;重力传播:Rmax=0.077,d=9 m;鸟类传播:Rmax=0.094,d=13 m),证明通过风力传播的种子在远距离传播上更有优势;(3)样地内存在一定程度的种子限制,有43个物种的种子只在很少的(〈10)收集器中出现.对于较为优势的物种而言(IV≥1),传播方式可能是影响种子限制的主要因素.另外,群落中通过风力传播的物种种子限制很可能仅仅是由于可繁殖个体的缺少造成的,而通过重力和鸟类传播的物种存在很高的种子限制可能是因为种子的低产量以及种子扩散能力的不足.
浙江天童常绿阔叶林不同扩散方式物种种子雨的基本特征 .https://doi.org/10.3969/j.issn.1000-5641.2015.02.015 URL [本文引用: 1] 摘要
为了研究不同传播方式物种种子雨的基本特征,以浙江天童常绿阔叶林20 hm2动态监测样地的种子雨为研究对象,分析了种子雨的密度、物种组成以及种子限制的强度.结果表明:(1)种子雨收集器共收集到种子136 38粒,隶属于28科66种,种子雨密度为146粒/m2;(2)样地内有56%的物种未收集到种子,因此种子雨与可繁殖个体物种组成的相似性较低且受不同传播方式影响,风力传播的相似性系数极值(Rmax)最大且距种子收集器距离(d)最远(风力传播:Rmax=0.226,d=20 m;重力传播:Rmax=0.077,d=9 m;鸟类传播:Rmax=0.094,d=13 m),证明通过风力传播的种子在远距离传播上更有优势;(3)样地内存在一定程度的种子限制,有43个物种的种子只在很少的(〈10)收集器中出现.对于较为优势的物种而言(IV≥1),传播方式可能是影响种子限制的主要因素.另外,群落中通过风力传播的物种种子限制很可能仅仅是由于可繁殖个体的缺少造成的,而通过重力和鸟类传播的物种存在很高的种子限制可能是因为种子的低产量以及种子扩散能力的不足.
|
| [12] |
Hypotheses on seed size: tests using the semi-arid flora of western New South Wales, Australia .https://doi.org/10.1086/285638 URL [本文引用: 1] 摘要
Seed size varies over several orders of magnitude in any one community. We outline a number of hypotheses that could account for this variation, briefly discuss the reasoning and evidence underlying each of these hypotheses, and then test each hypothesis with a database of 248 species from the semiarid woodlands of western New South Wales, Australia. Information on seed weight, growth form, plant longevity, height, dispersal mode, dormancy, and germination season was used. We considered not only pairwise relationships between seed weight and each other variable, but also alternative hypotheses whereby relationships arose as a result of indirect correlations through other variables. The strongest associations of seed size were with plant height and growth form. The seed-size variation accounted for by growth form largely overlapped with that accounted for by plant height, but each also accounted for some further variation independently of the other. Of the five hypotheses tested, the correlative patterns were inconsistent for two. Two others showed the predicted pattern, but these patterns could alternatively be interpreted as arising from secondary correlation via the combination of plant height and growth form. Only plant height, growth form, and dispersal mode had significant associations with seed size independent of the other attributes measured.
|
| [13] |
Correlates of seed size variation: a comparison among five temperate floras .https://doi.org/10.2307/2261604 URL [本文引用: 1] 摘要
1 Five temperate floras were studied to assess to what extent seed size correlations with other plant attributes are consistent across floras. The floras were from three continents: Australia (semiarid woodlands of western New South Wales, arid woodlands of Central Australia, and the Sydney region), North America (Indiana Dunes) and Europe (Sheffield region, UK). The plant attributes used were growth form, plant height, perenniality and dispersal mode. We used general linear models to consider not only the primary correlations between seed size and each other attribute, but also the overlap patterns among correlations to determine if each correlation could be interpreted as a secondary effect via a third variable.
|
| [14] |
Seed rain dynamics reveals strong dispersal limitation, different reproductive strategies and responses to climate in a temperate forest in Northeast China .https://doi.org/10.1111/j.1654-1103.2011.01344.x URL [本文引用: 1] 摘要
QuestionsSpecies co-existence may be achieved by limited dispersal due to the reduced chance of inter-specific competition. But the timing of seed release also provides clues for species co-existence because asynchronous reproduction can alleviate inter-specific competition for dispersal agents, while synchronous reproduction can facilitate overall seed dispersal. How strong is seed dispersal limitation, do co-existing species release seeds synchronously or asynchronously, and what is the relationship between seed production and main meteorological measures?LocationA 25-ha plot in a temperate forest, Changbai Mountain, Northeast China.MethodsWe calculated Jaccard coefficients between seed rain composition and neighbouring adult tree composition, analysed long-term seed rain dynamics for both the whole community and different species, and regressed seed rain density with meteorological measures using autoregressive models.ResultsThe Jaccard coefficient dropped sharply as neighbourhood radius increased to about 10 m, indicating severe dispersal limitation. Both synchrony and asynchrony in seed release were observed: most species released mature seeds synchronously from August to November, with small segregations in time; one species released seeds in Jun. Bimodal and unimodal seasonal dynamics of seed rain were observed and some species released seeds beyond the main fruiting seasons. The seasonal dynamics of seed release might be driven by different strategies of seed dispersal. Seed rain density is significantly positively related to temperature and precipitation, with a 2-mo time lag.ConclusionsBoth dispersal limitation and timing of seed release by co-existing species may contribute to maintenance of diversity of this forest, but variations in temperature and precipitation considerably alter seed rain density.
|
| [15] |
Fruit composition and seed dispersal strategies of woody plants in a Dujiangyan subtropical forest, Southwest China .都江堰亚热带森林常见木本植物果实组成与种子扩散策略 . |
| [16] |
|
| [17] |
Factors that shape seed mass evolution .https://doi.org/10.1073/pnas.0501473102 URL PMID: 16030149 [本文引用: 1] 摘要
Abstract We used correlated divergence analysis to determine which factors have been most closely associated with changes in seed mass during seed plant evolution. We found that divergences in seed mass have been more consistently associated with divergences in growth form than with divergences in any other variable. This finding is consistent with the strong relationship between seed mass and growth form across present-day species and with the available data from the paleobotanical literature. Divergences in seed mass have also been associated with divergences in latitude, net primary productivity, temperature, precipitation, and leaf area index. However, these environmental variables had much less explanatory power than did plant traits such as seed dispersal syndrome and plant growth form.
|
| [18] |
Epizoochorous dispersal by barbs, hooks, and spines in a lowland moist forest in central French Guiana .https://doi.org/10.2307/2807846 URL [本文引用: 1] 摘要
An analysis of the flowering plant flora of a lowland moist forest in central French Guiana reveals 25 species with adaptations for epizoochorous dispersal by barbs, hooks, or spines (stick-tights). This represents 1.5% of the flowering plant flora. Stick-tights are represented among the monocotyledons of the flora of central French Guiana by five species of Poaceae. The Fabaceae and Asteraceae, each with six species with stick-tight dispersal, are the richest families of flowering plants in terms of stick-tight dispersal in central French Guiana. This region possesses numerous potential mammal and bird dispersal agents, therefore it is a puzzle why stick-tight dispersal, especially among species of the canopy, is not more frequent.
|
| [19] |
Demography and dispersal: calculation and sensitivity analysis of invasion speed for structured populations .https://doi.org/10.1890/0012-9658(2000)081[1613:DADCAS]2.0.CO;2 URL [本文引用: 1] |
| [20] |
Research on decision tree in data mining .https://doi.org/10.3969/j.issn.1001-3695.2001.08.006 URL [本文引用: 1] 摘要
决策树算法是DM的一个活跃的研究领域.首先给出了DM中决策树算法的基本思想,然后讨论了决策树算法中的难点问题,提出了利用熵与加权和的思想来选择取值的算法.
数据挖掘中决策树算法的探讨 .https://doi.org/10.3969/j.issn.1001-3695.2001.08.006 URL [本文引用: 1] 摘要
决策树算法是DM的一个活跃的研究领域.首先给出了DM中决策树算法的基本思想,然后讨论了决策树算法中的难点问题,提出了利用熵与加权和的思想来选择取值的算法.
|
| [21] |
Do big plants have big seeds? https://doi.org/10.1086/284947 URL [本文引用: 1] 摘要
ABSTRACT Big plants do tend to have big seeds, and small plants small seeds (in over half the families examined). The existence of this correlation ought, therefore, to be borne in mind by anyone seeking an adaptive explanation of seed size in herbaceous flowering plants. Three possible explanations of this correlation are proposed, although only one hypothesis proposes an adaptive connection between seed size and plant size. -from Authors
|
| [22] |
Chasing the unknown: predicting seed dispersal .https://doi.org/10.1111/j.1365-2745.2010.01724.x URL [本文引用: 3] 摘要
Summary 1. 68The dispersal capabilities of most plant species remain unknown. However, gaining basic dispersal information is a critical step for understanding species’ geographical distributions and for predicting the likely impacts of future climate change. Dispersal mechanisms can indicate short- or long-distance dispersers, and highlight important biological interactions. 2. 68To predict dispersal mechanisms for species where information is limited, we used generalized linear mixed models with basic life-history and ecological traits. Sets of models were created (using Australian species) for six dispersal categories: wind, unassisted, water, ant, vertebrate-ingestion and vertebrate-attachment dispersal mechanisms. We validated our models on the dispersal mechanisms of 50 Australian, 30 Californian, 30 Swiss plant species and a global compilation of 70 species. 3. 68Growth form, seed mass and vegetation type were the main predictor variables. Our models predicted dispersal mechanisms for Australian and Californian plant species equally well ( c.70% correct) and to a lesser extent for the Swiss flora ( c.50% correct). Our models predicted observed dispersal mechanisms ( c.50% correct) equally well to inferred dispersal mechanisms (based on seed morphology). 4.68 Synthesis. Our approach of using easily obtainable traits for predicting dispersal mechanisms of species allows dispersal information to be predicted for species where little is known. From here, the application of realistic dispersal curves to the predicted dispersal mechanisms will further understanding on the dispersal capabilities of species.
|
| [23] |
Seed dispersal distance is more strongly correlated with plant height than with seed mass .https://doi.org/10.1111/j.1365-2745.2011.01867.x URL [本文引用: 1] 摘要
Summary 1. 68It is often assumed that there is a trade-off between maternal provisioning and dispersal capacity, leading small-seeded species to disperse further than large-seeded species. However, this relationship between dispersal distance and seed mass has only been quantified for species from particular sites or with particular dispersal syndromes. 2. 68We provided the first large-scale, cross-species quantification of the correlations between dispersal distance and both seed mass and plant height. Seed mass was positively related to mean dispersal distance, with a 100-fold increase in seed mass being associated with a 4.5-fold increase in mean dispersal distance ( R 2 =0.16; n = 210 species; P < 0.001). However, plant height had substantially stronger explanatory power than did seed mass, and we found a 5-fold increase in height was associated with a 4.6-fold increase in mean dispersal distance ( R 2 =0.54; n = 211 species; P < 0.001). 3. 68Once plant height was accounted for, we found that small-seeded species dispersed further than did large-seeded species ( R 2 =0.54; n = 181 species; slope=610.130; P < 0.001); however, seed mass only added 2% to the R 2 of the model. Within dispersal syndromes, tall species dispersed further than did short species, while seed mass had little influence on dispersal distance. 4. 68 Synthesis. These findings enhance our understanding of plant life-history strategies and improve our ability to predict which species are best at colonizing new environments.
|
| [24] |
Predicting global change impacts on plant species’ distributions: future challenges .https://doi.org/10.1016/j.ppees.2007.09.004 URL [本文引用: 1] 摘要
In this review, we propose two main avenues to progress the understanding and prediction of the different processes occurring on the leading and trailing edge of the species’ distribution in response to any global change phenomena. Deliberately focusing on plant species, we first explore the different ways to incorporate species’ migration in the existing modelling approaches, given data and knowledge limitations and the dual effects of climate and land-use factors. Secondly, we explore the mechanisms and processes happening at the trailing edge of a shifting species’ distribution and how to implement them into a modelling approach. We finally conclude this review with clear guidelines on how such modelling improvements will benefit conservation strategies in a changing world.
|
| [25] |
|
| [26] |
Principles of Dispersal in Higher Plants. Springer-Verlag, New York . |
| [27] |
Germination characteristics of 61 common woody species from the eastern Qinghai-Tibet Plateau of China and their life history correlates .
为了评价亚高山地带木本植物种子萌发对策及其与生活史关联, 该文对青藏高原东缘61种常见木本植物的种子萌发特征以及种子萌发与种子大小、扩散方式、海拔及生境的关系进行了调查分析。结果表明: 1) 61种植物的种子萌发率呈现偏斜、双峰分布。6种植物(9.8%)的种子萌发率大于80%, 13种(21.3%)的萌发率在60%~80%之间, 9种(14.8%)的萌发率在40%~60%之间, 8种(13.1%)的萌发率在20%~40%之间, 25种(41%)的萌发率小于20%, 其中8种植物的种子萌发率为零, 显示了该生境中木本植物的种子以休眠、低萌发率或连续稳定的萌发对策占优势, 以避免不利的环境因素。2)种子扩散方式是决定61种木本植物种间萌发率变异(R2 = 24.1%, p < 0.001) 和萌发开始时间变异(R2 = 21.3%, p< 0.01) 的主要因素; 种子大小、母体植株的海拔和生境对种子萌发率和萌发开始时间几乎没有影响(p >0.05); 风扩散的种子比脊椎动物扩散和无助扩散的种子有更高的萌发率(F = 9.219, p<0.001) 和较早的萌发开始时间(F = 6.772, p<0.01)。说明植物生活史特征如种子萌发与扩散方式之间存在固有而紧密的联系, 扩散能力强的种子(如风扩散)由于能散布到较远而空旷的生境, 可能避免了各种有害因素或个体竞争, 因而种子以较强的萌发能力进行拓殖; 扩散能力弱的种子(如无助扩散)由于其散布距离较近, 种子以休眠或降低萌发的方式来避免个体或同胞竞争以及各种有害因素, 以获得最大的生态利益并确保物种延续。
青藏高原东缘61种常见木本植物种子萌发特性及其与生活史的关联 .
为了评价亚高山地带木本植物种子萌发对策及其与生活史关联, 该文对青藏高原东缘61种常见木本植物的种子萌发特征以及种子萌发与种子大小、扩散方式、海拔及生境的关系进行了调查分析。结果表明: 1) 61种植物的种子萌发率呈现偏斜、双峰分布。6种植物(9.8%)的种子萌发率大于80%, 13种(21.3%)的萌发率在60%~80%之间, 9种(14.8%)的萌发率在40%~60%之间, 8种(13.1%)的萌发率在20%~40%之间, 25种(41%)的萌发率小于20%, 其中8种植物的种子萌发率为零, 显示了该生境中木本植物的种子以休眠、低萌发率或连续稳定的萌发对策占优势, 以避免不利的环境因素。2)种子扩散方式是决定61种木本植物种间萌发率变异(R2 = 24.1%, p < 0.001) 和萌发开始时间变异(R2 = 21.3%, p< 0.01) 的主要因素; 种子大小、母体植株的海拔和生境对种子萌发率和萌发开始时间几乎没有影响(p >0.05); 风扩散的种子比脊椎动物扩散和无助扩散的种子有更高的萌发率(F = 9.219, p<0.001) 和较早的萌发开始时间(F = 6.772, p<0.01)。说明植物生活史特征如种子萌发与扩散方式之间存在固有而紧密的联系, 扩散能力强的种子(如风扩散)由于能散布到较远而空旷的生境, 可能避免了各种有害因素或个体竞争, 因而种子以较强的萌发能力进行拓殖; 扩散能力弱的种子(如无助扩散)由于其散布距离较近, 种子以休眠或降低萌发的方式来避免个体或同胞竞争以及各种有害因素, 以获得最大的生态利益并确保物种延续。
|
| [28] |
Comparative evolutionary ecology of seed size .https://doi.org/10.1016/0169-5347(92)90006-W URL PMID: 21236070 [本文引用: 1] 摘要
A seedling's chances of establishing successfully are likely to be affected by the quantity of metabolic reserves in the seed. Seed size is thought to evolve as a compromise between producing numerous smaller seeds, each with few resources, and fewer larger seeds, each with more resources. Seed size varies 10 11 -fold across plant species, so the compromise has been struck at very different levels. These basic ideas have been accepted for 50 years, and many studies have interpreted seed size differences between species by reference to larger seed size being adaptive under a variety of hazards. However, experimental tests of the benefits of large seed size in relation to particular hazards have been rare. More experiments are now being reported, but a consistent picture has yet to emerge. There is typically at least a 10 5 -fold range of seed mass between species even within a single area, suggesting that much seed size variation is evolutionarily associated with other plant attributes.
|
| [29] |
Comparative ecology of seed size and dispersal .https://doi.org/10.1098/rstb.1996.0114 URL [本文引用: 1] 摘要
ABSTRACT Seed mass is correlated with a number of other plant traits, including dispersal mode, growth form and specific leaf area. Specific leaf area is the main determinant of potential relative growth rate and an indicator of the site quality to which a species is adapted. The relationships with dispersal mode and growth form have consistent form in five datasets from three continents, and each account for about 20-30% of variation in log seed mass. Thus, there is also very substantial variation within growth form and dispersal categories. Much, but not all, of the 20-30% is associated with shifted family composition between growth forms or dispersal modes. Experiments have shown that seedlings of larger-seeded species are better able to survive hazards including deep shade, drought, physical damage and the presence of competing vegetation. If there is a common mechanism under these different hazards, it seemingly must be a 'reserve effect', whereby during deployment and early growth larger-seeded species hold a bigger percentage of seed reserves uncommitted to seedling structure and available to support respiration or repair damage. A reserve effect has not yet been demonstrated directly. It remains possible that different mechanisms operate under different hazards. Under a reserve effect, advantages of larger seed size should be temporary, and temporary advantage has indeed been observed with regard to seedling survival under dense shade. Although larger seed mass confers benefits on seedlings, larger seeds must necessarily be produced in smaller numbers per unit of resource allocated. Seed mass is presumed to have evolved as a compromise between these counterposed pressures. Yet there has proved to be surprisingly little difference in average seed mass between very different vegetation regions, at least in temperate climates. Rather, there is startlingly wide variation in seed mass among species growing interspersed with each other. Recent applications of game theory may be capable of accounting for this wide variation between coexisting species, but at present these models are driven by competition among seedling species (as opposed to between seedlings and adults). It remains unclear whether competition among seedlings is a decisive influence on species composition in most of the world's vegetation types.
|
| [30] |
Seed size and plant- growth form as factors in dispersal spectra .https://doi.org/10.2307/1938268 URL [本文引用: 1] 摘要
All vascular plant species were listed on eight sites near Sydney, Australia. Four sites were on infertile and four on fertile soil. Each of the 355 species was classified according to seed mass (measured as fresh mass of embryo plus endosperm), morphological adaptations for dispersal by different vectors, and growth form. The infertile-soil sites had more species and more cover adapted for dispersal by ants, and the fertile-soil sites had more species and more cover with fleshy fruits adapted for dispersal by vertebrates. We tested the hypothesis that this difference could be interpreted as a secondary correlate of seed size or of growth form. Species with smaller seed mass of growing to <2 m tall were significantly more likely to be adapted for dispersal by ants relative to vertebrates. An indirect association via growth form was capable of accounting for up to 84% of the relationship between soil type and dispersal mode, and indirect association via seed mass for up to 23% of the relationship; the two together could account for a maximum of 85%. The indirect association association via seed mass relatively weaker because seed masses proved not to be very different between these soil types. There remained a significant residual tendency for species in any given seed size class and growth form to be more likely to be dispersed by vertebrates relative to ants on fertile soils than on infertile soils.
|
| [31] |
Dispersal mode, seed shadows, and colonization patterns .https://doi.org/10.1007/BF00052229 URL [本文引用: 1] 摘要
This review assesses the state of our knowledge about comparative seed shadows. Using data presently available in the literature, I compare the slopes (on a log-linear scale) of seed shadows for plants with different morphologically characterized modes of dispersal. The seeds of many species have no evident morphological adaptation for dispersal and seem to achieve only short-distance dispersal. Seed shadows for herbaceous species with devices for wind have flatter slopes and more distant modes and maxima than those of ballists, which in turn exceed those with no special devices. Seed shadows for wind-dispersed trees and shrubs had similar or steeper slopes than those for vertebrate-dispersed species in this sample. Species with poor mechanisms for dispersal in space only sometimes had the capacity for better dispersal in time (dormancy). Although some species exhibited seed shadows sufficiently steep to be predicted to colonize new-habitat in a ‘front’ or phalanx pattern, actual colonization patterns must reflect many other factors.
|
| [32] |
Seed dispersal spectra: a comparison of temperate plant-communities .https://doi.org/10.2307/3235789 URL [本文引用: 1] 摘要
Abstract Abstract. We compare the dispersal spectra of diaspores from varied plant communities in Australia, New Zealand, and North America, assigning dispersal mode to each diaspore type on the basis of apparent morphological adaptations. Species with ballistic and external dispersal modes were uncommon in most communities we surveyed. Ant dispersal was also rather uncommon, except in some Australian sclerophyll vegetation types. The frequency of vertebrate dispersal ranged up to 60% of the flora, the highest frequencies occurring in New Zealand forests. Wind dispersal ranged as high as 70% of the flora, with the highest values in Alaska, but usually comprised 10-30% of the flora. Many species in most communities had diaspores with no special morphological device for dispersal. Physiognomically similar vegetation types indifferentbiogeographic regions usually had somewhat dissimilar dispersal spectra. The frequency of dispersal by vertebrates often increased and the frequency of species with no special dispersal device decreased along gradients of increasing vertical diversity of vegetation structure. Elevation and moisture gradients also exhibited shifts in dispersal spectra. Within Australia, vertebrate- and wind-dispersal increased in frequency along a soil-fertility gradient, and dispersal by ants and by no special device decreased. Habitat breadths (across plant communities) and microhabitat breadths (within communities) for species of each major dispersal type did not show consistent differences, in general. Ant-dispersed species often had lower cover-values than other species in several Australian vegetation types. We discuss the ecological bases of these differences in dispersal spectra in terms of the availability of dispersal agents, seed size, and other ecological constraints. Seed size is suggested to be one ecological factor that is probably of general relevance to the evolution of dispersal syndromes.
|
| [33] |
Effect of seed traits on spatial aggregation of trees in a subtropical evergreen .https://doi.org/10.17520/biods.2014007 URL [本文引用: 1] 摘要
种子的特征如种子的重量、大小和传播方式直接影响物种的扩散能力,从而影响物种空间分布的聚集强度.作者以古田山亚热带常绿阔叶林24 ha样地中89种木本植物为研究对象,分析了它们的种子大小、重量和扩散方式与物种聚集强度的关系.结果表明:在20 m范围内,样地内90%的物种为聚集分布,聚集强度随着尺度的增大而减弱,稀有种的聚集强度大于常见种.用系统发育独立比较 (phylogenetically independent contrasts)方法排除了系统发育影响后,物种的多度与聚集强度呈显著相关(在15m尺度,R2=0.32,P<0.001);种子大小与物种聚集 强度的相关性较弱(R2=0.05,P<0.05),而种子重量与物种聚集强度之间的相关性相对较强(R2=0.14,P<0.05).种子的传播方式显 著地影响物种的聚集强度:无助力扩散物种的聚集强度显著地高于有助力扩散物种的聚集强度(F1,87=4.439,P=0.038),即长距离扩散导致物 种的聚集强度减弱.可见,在亚热带森林中,种子性状通过影响种子扩散进而引起物种聚集强度的变化,是影响物种空间分布格局的重要生态因素.
亚热带常绿阔叶林种子性状对木本植物聚集格局的影响 .https://doi.org/10.17520/biods.2014007 URL [本文引用: 1] 摘要
种子的特征如种子的重量、大小和传播方式直接影响物种的扩散能力,从而影响物种空间分布的聚集强度.作者以古田山亚热带常绿阔叶林24 ha样地中89种木本植物为研究对象,分析了它们的种子大小、重量和扩散方式与物种聚集强度的关系.结果表明:在20 m范围内,样地内90%的物种为聚集分布,聚集强度随着尺度的增大而减弱,稀有种的聚集强度大于常见种.用系统发育独立比较 (phylogenetically independent contrasts)方法排除了系统发育影响后,物种的多度与聚集强度呈显著相关(在15m尺度,R2=0.32,P<0.001);种子大小与物种聚集 强度的相关性较弱(R2=0.05,P<0.05),而种子重量与物种聚集强度之间的相关性相对较强(R2=0.14,P<0.05).种子的传播方式显 著地影响物种的聚集强度:无助力扩散物种的聚集强度显著地高于有助力扩散物种的聚集强度(F1,87=4.439,P=0.038),即长距离扩散导致物 种的聚集强度减弱.可见,在亚热带森林中,种子性状通过影响种子扩散进而引起物种聚集强度的变化,是影响物种空间分布格局的重要生态因素.
|
| [34] |
Advances in ecology of mass .https://doi.org/10.17521/cjpe.2007.0126 URL [本文引用: 2] 摘要
作为植物生活史中的一个关键性特征,种子重量与其它许多植物性状和生态因子有关,种子重量的分异与其它一些植物性状及环境的变化关系在进化生物学上已经成为一个非常有意义的研究内容,且具有一定的实践意义.种子重量被发现与下列的一些植物学和群落学性状有关:植物的生活型、种子的散布能力、种子的散布方式、植物的高度、植物的冠幅、植物的比叶面积、植物的寿命、动物的捕食、植被中植物的数量或多度、土壤中种子的数量或多度、种子的休眠、种子在土壤中的持久性和植物的净初级生产力等,另外生态因子如降雨、温度、坡向、海拔、经度、纬度、光强和干扰等都影响种子的重量.种子的重量被认为是在大量小种子和少量大种子之间的进化折衷,在一定的能量限度内,较大重量的种子一般具有较少的数量,而较小重量的种子一般数量较多,这是种子重量和数量方面具有的一种反向关系.与其它性状相比,很多研究都表明种子重量和植物的生活型的关系密切.没有散布结构或风散布的种子比以动物和水作为散布媒介的种子重量要小.种子重量与捕食的关系现发现有3种格局.种子重量和形状与种子在土壤中的持久 性的关系有4种格局.在干旱和阴暗的环境条件下,种子有变大的趋势.大重量种子比小种子赋予幼苗较优势的竞争地位,其原理尚有争论,尚不清楚是否是幼苗阶段的竞争决定了世界上大部分植被类型的物种组成.未来的研究方向主要有以下几个方面:1)种子重量与植物系统学相结合,探索种子重量的变化规律;2)调查群落三向(纬度、经度和海拔)性的种子重量谱变化规律;3)群落演替与群落种子重量谱的变化;4)种子重量与群落中植物个体和种子的数量的关系及机理研究;5) 微生境、微地形如坡向、坡位和林间隙等对种子重量的影响;6) 全球气候变化和种子重量变化的关系.
种子重量的生态学研究进展 .https://doi.org/10.17521/cjpe.2007.0126 URL [本文引用: 2] 摘要
作为植物生活史中的一个关键性特征,种子重量与其它许多植物性状和生态因子有关,种子重量的分异与其它一些植物性状及环境的变化关系在进化生物学上已经成为一个非常有意义的研究内容,且具有一定的实践意义.种子重量被发现与下列的一些植物学和群落学性状有关:植物的生活型、种子的散布能力、种子的散布方式、植物的高度、植物的冠幅、植物的比叶面积、植物的寿命、动物的捕食、植被中植物的数量或多度、土壤中种子的数量或多度、种子的休眠、种子在土壤中的持久性和植物的净初级生产力等,另外生态因子如降雨、温度、坡向、海拔、经度、纬度、光强和干扰等都影响种子的重量.种子的重量被认为是在大量小种子和少量大种子之间的进化折衷,在一定的能量限度内,较大重量的种子一般具有较少的数量,而较小重量的种子一般数量较多,这是种子重量和数量方面具有的一种反向关系.与其它性状相比,很多研究都表明种子重量和植物的生活型的关系密切.没有散布结构或风散布的种子比以动物和水作为散布媒介的种子重量要小.种子重量与捕食的关系现发现有3种格局.种子重量和形状与种子在土壤中的持久 性的关系有4种格局.在干旱和阴暗的环境条件下,种子有变大的趋势.大重量种子比小种子赋予幼苗较优势的竞争地位,其原理尚有争论,尚不清楚是否是幼苗阶段的竞争决定了世界上大部分植被类型的物种组成.未来的研究方向主要有以下几个方面:1)种子重量与植物系统学相结合,探索种子重量的变化规律;2)调查群落三向(纬度、经度和海拔)性的种子重量谱变化规律;3)群落演替与群落种子重量谱的变化;4)种子重量与群落中植物个体和种子的数量的关系及机理研究;5) 微生境、微地形如坡向、坡位和林间隙等对种子重量的影响;6) 全球气候变化和种子重量变化的关系.
|
| [35] |
|
| [36] |
Advances in model construction of anemochoric seed long distance dispersal .
植物种子的长距离传播在物种迁移、生物入侵、保护生物学等领域有重要的生态和进化意义。种子传播有很多方式,开阔草原等地区的草本植物和许多热带和温带的树木都是通过风传播种子的。风传播的方式最适合进行种子长距离传播现象的模拟研究。种子的风传播模型是传播生态研究的一个重要领域,尤其是种子的长距离风传播模型,对于外来入侵植物的扩散和破碎化景观中植物种群的基因交流等生态过程研究举足轻重,然而国内鲜见这方面的研究成果。本文综述了种子长距离风传播现象研究的背景和意义,分析了风传播种子模型的基本形式和构成原理,并分别就现象模型和机理模型的相关研究进展进行了总结,同时指出了未来发展的几个重要方向。种子的风传播模型可以分为现象模型和机理模型两类,现象模型按种子传播核心的形式包括短尾模型、偏峰长尾模型和混合传播核心模型,后两者对于长距离传播数据的模拟可以取得很好的效果。机理模型按照模拟机制可分为欧拉对流扩散模型和拉格郎日随机模型两类。本文重点介绍了种子的长距离风传播现象的形成机理和两类机理模型的参数构成和处理方式。适合种子脱落的天气和适合传播的天气的同步性可能是形成种子长距离风传播的一个重要前提,林缘和地表存在的上升气流及大风和暴风中形成的速度梯度都可能对于种子的长距离传播有重要的作用。机理模型的操作因子主要包括生物方面的因子、气象方面的因子和地形方面的因子。同时对目前几个应用比较成功的机理模型进行了简要的介绍和评价,包括倾斜羽毛模型、对流-扩散-下降模型、无掩蔽模型、背景模型、WINDISPER及其改进模型和PAPPUS模型。最后指出,目前在风传播种子的长距离模型研究中,对草本植物种子的传播模拟的投入明显不如树木种子的长距离传播模拟,对于破碎化景观中种子长距离的风传播的研究还存在很大的差距,而对提高机理模型预测能力的高分辨率物理环境数据输入技术的需求则为多学科交叉提供了很好的机会。
种子的长距离风传播模型研究进展 .
植物种子的长距离传播在物种迁移、生物入侵、保护生物学等领域有重要的生态和进化意义。种子传播有很多方式,开阔草原等地区的草本植物和许多热带和温带的树木都是通过风传播种子的。风传播的方式最适合进行种子长距离传播现象的模拟研究。种子的风传播模型是传播生态研究的一个重要领域,尤其是种子的长距离风传播模型,对于外来入侵植物的扩散和破碎化景观中植物种群的基因交流等生态过程研究举足轻重,然而国内鲜见这方面的研究成果。本文综述了种子长距离风传播现象研究的背景和意义,分析了风传播种子模型的基本形式和构成原理,并分别就现象模型和机理模型的相关研究进展进行了总结,同时指出了未来发展的几个重要方向。种子的风传播模型可以分为现象模型和机理模型两类,现象模型按种子传播核心的形式包括短尾模型、偏峰长尾模型和混合传播核心模型,后两者对于长距离传播数据的模拟可以取得很好的效果。机理模型按照模拟机制可分为欧拉对流扩散模型和拉格郎日随机模型两类。本文重点介绍了种子的长距离风传播现象的形成机理和两类机理模型的参数构成和处理方式。适合种子脱落的天气和适合传播的天气的同步性可能是形成种子长距离风传播的一个重要前提,林缘和地表存在的上升气流及大风和暴风中形成的速度梯度都可能对于种子的长距离传播有重要的作用。机理模型的操作因子主要包括生物方面的因子、气象方面的因子和地形方面的因子。同时对目前几个应用比较成功的机理模型进行了简要的介绍和评价,包括倾斜羽毛模型、对流-扩散-下降模型、无掩蔽模型、背景模型、WINDISPER及其改进模型和PAPPUS模型。最后指出,目前在风传播种子的长距离模型研究中,对草本植物种子的传播模拟的投入明显不如树木种子的长距离传播模拟,对于破碎化景观中种子长距离的风传播的研究还存在很大的差距,而对提高机理模型预测能力的高分辨率物理环境数据输入技术的需求则为多学科交叉提供了很好的机会。
|
| [37] |
Seed predation and dispersal of Hevea brasiliensis in Xishuangbanna introduction area .
在纯橡胶林、胶.竹复合林、沟谷季节雨林、次生林及毗邻次生林的橡胶林共5种不同生境中释放标记的橡胶树种子,观测种子捕食者以及种子被捕食、埋藏和扩散情况。结果表明:纯橡胶林和沟谷季节雨林内的橡胶树种子大多数都原地存留(>80%和>98%),而胶一竹复合林内几乎100%的种子均被捕食;次生林内释放的橡胶树种子捕食率也几为100%,而毗邻次生林的橡胶林种子捕食率约50%;卡方检验表明,胶-竹复合林的种子捕食率显著高于沟谷季节雨林和纯橡胶林,次生林内显著高于毗邻的橡胶林(P<0.001)。绝大多数橡胶树种子被捕食者原地或搬运后取食,仅有少量种子被埋藏。大多数种子(>80%)被搬运距离在10m以内。红外感应相机拍摄和饲喂实验发现,黄胸鼠(Rattus anezumi)和红刺鼠(Rattus urifer)为主要的种子捕食者。可见,在西双版纳引种区橡胶树种子主要是被小型鼠类取食,但捕食强度较大,而被埋藏种子比例很低,散布距离较短(<10m)。
橡胶树在西双版纳引种区的种子捕食与散布 .
在纯橡胶林、胶.竹复合林、沟谷季节雨林、次生林及毗邻次生林的橡胶林共5种不同生境中释放标记的橡胶树种子,观测种子捕食者以及种子被捕食、埋藏和扩散情况。结果表明:纯橡胶林和沟谷季节雨林内的橡胶树种子大多数都原地存留(>80%和>98%),而胶一竹复合林内几乎100%的种子均被捕食;次生林内释放的橡胶树种子捕食率也几为100%,而毗邻次生林的橡胶林种子捕食率约50%;卡方检验表明,胶-竹复合林的种子捕食率显著高于沟谷季节雨林和纯橡胶林,次生林内显著高于毗邻的橡胶林(P<0.001)。绝大多数橡胶树种子被捕食者原地或搬运后取食,仅有少量种子被埋藏。大多数种子(>80%)被搬运距离在10m以内。红外感应相机拍摄和饲喂实验发现,黄胸鼠(Rattus anezumi)和红刺鼠(Rattus urifer)为主要的种子捕食者。可见,在西双版纳引种区橡胶树种子主要是被小型鼠类取食,但捕食强度较大,而被埋藏种子比例很低,散布距离较短(<10m)。
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}