岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 )。海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿。海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 )。
海南岛生物多样性的起源及其与周边地区的联系一直存在争议(Francisco-Ortega et al, 2010 ; Zhu, 2016 )。张宏达(1962, 2001)指出, 海南岛、台湾岛、广东、广西、福建南部、云南东南部及中南半岛北部等地区的植物均属华南亚区; 张超常和刘兰芳(1983)认为, 海南岛属于华夏植物区系; 而蒋有绪(1988)认为, 海南岛的植物区系应属印度-马来西亚区系。王荷生(2000)、吴征镒等(2011)以及Zhu (2016)基本赞同蒋有绪的结论, 认为海南岛及邻近地区植物区系应属马来西亚亚区。然而, 传统植物区系地理学研究主要以植物区系的分类群组成及其分布区类型的分析为主, 相对忽视了进化历史的分析(李嵘和孙航, 2017 )。
海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 )。苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 )。海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 )。苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口。
本文首先比较分析了海南岛及其周边地区的苦苣苔科物种多样性与地理分布特点, 然后利用核基因与叶绿体基因重建海南岛自然分布的苦苣苔科物种系统关系, 并结合海南岛物种地理分布格局、相关地质历史及与周边地区物种组成的比较等, 分析了海南岛苦苣苔科物种迁移历史与特有种形成时间, 拟为揭示海南岛植物多样性形成与演化历史提供依据。
1 材料与方法
1.1 海南岛及邻近地区的苦苣苔科物种多样性
为确定海南岛苦苣苔科植物的物种多样性水平及其与周边地区的联系, 本文收集了海南岛、云南、广西、贵州、广东及邻国越南苦苣苔科物种多样性数据(Pham, 1991; 李振宇和王印政, 2005; 韦毅刚, 2010; 表1 )。为降低这6个地区因为面积差异过大可能产生的“抽样效应” (sampling effects), 本文参照Tang等(2006) , 通过面积归一化(自然对数转换)之后计算各地区的物种密度(species density, SD)和特有指数(endemic index, EI):
SD = N /ln(A ) (1)
EI = [NE /(N -NE )] / ln(A ) (2)
其中, N 为该地区苦苣苔科物种总数, NE 为该地区苦苣苔科特有种数, A 为该地区地理面积(单位: km2 )。
根据陈圣宾等(2010), 我们使用目前广泛应用的Jaccard相似性指数(Jaccard, 1912 )来度量海南岛与其邻近地区的物种组成相似性程度: C j = c / (a + b - c )。其中, a 为海南岛苦苣苔科物种数, b 为邻近某地区的物种数, c 为两地共有物种数。
1.2 海南岛苦苣苔科植物的地理分布格局
水平分布格局的研究选择海南岛8个主要山区, 即五指山(主峰海拔1,867 m)、鹦哥岭(1,812 m)、霸王岭(1,654 m)、吊罗山(1,499 m)、尖峰岭(1,413 m)、黎母山(1,412 m)、俄贤岭(1,238 m)和仙安石林 (1,107 m)等。这些地点涵盖了海南岛整个中南部山区, 也覆盖了苦苣苔科在海南岛的整个分布区域, 能客观反映海南岛苦苣苔科的物种多样性及水平分布格局。
以200 m的海拔间隔, 对苦苣苔科所有物种和特有种在以上8个地点不同海拔区间的垂直分布格局进行分析, 确定物种多样性和特有种丰富度最高的海拔位置。
1.3 海南岛苦苣苔科的物种系统发育关系
在海南岛全岛范围采集了海南特有种烟叶唇柱苣苔(Chirita heterotricha )、海南蛛毛苣苔(Paraboea hainanensis )、盾叶苣苔(Metapetrocosmea peltata )、黄花马铃苣苔(Oreocharis flavida )、毛花马铃苣苔(O. dasyantha )以及广布种红花芒毛苣苔(Aeschynanthus moningerae )各不少于5个个体的幼叶, 经变色硅胶干燥, 然后用改良的CTAB (Cetyl Trimethyl Ammonium Bromide, 十六烷基三甲基溴化铵)法提取叶片总DNA (Doyle & Doyle, 1987 )。选择变异程度大的叶绿体基因片段trn L-F (Wang et al, 2010 )和核基因片段ITS1/2 (White et al, 1990 )进行扩增和序列分析(表2 )。
PCR反应体系25 µL, 包括17 µL去离子水, 2.5 µL 10 ×缓冲液, 0.5 µL 10 mM dNTPs, 5 µM引物各0.5 uL, 1 µL DNA和 0.5 µL 5 U/µL Taq 酶 (Aidlab公司)。ITS1/2的反应程序: 94℃预变性5min, 94℃变性1 min, 55℃退火1 min, 72℃延伸1 min, 共35个循环, 最后在72℃延伸10 min。Trn L-F的反应程序: 94℃预变性4 min, 94℃变性30 s, 58℃退火40 s, 72℃延伸1 min, 共35个循环, 最后72℃延伸10 min。PCR产物送华大基因测序。
本研究中, 除通过测序获得6个苦苣苔科物种DNA序列外, 海南岛分布的苦苣苔科有14个物种(包括海南特有种扁蒴苣苔(Cathayanthe biflora ))的ITS1/2和trn L-F序列都可从GenBank获得(附录1)。为标定物种分化时间, 补充与亚洲苦苣苔科亲缘关系最近的、物种分化时间(57 Ma)已知的Sanango racemosum (Perret et al, 2013 )一起建树。因此, 一共有21个海南岛苦苣苔科植物的DNA序列(附录1)用于建立系统发育树。各物种的核基因ITS1/2片段和叶绿体trn L-F片段用SequenceMatrix软件(Vaidya et al, 2011 )拼接; 用MEGA 6.0软件(Kumar et al, 2008 )整合拼接好的片段。
用BEAST v1.7.5 (Drummond & Rambaut, 2007 )建立海南岛苦苣苔科植物的系统发育树。首先, 碱基替代模型为根据MODELTEST v3.6 (Posada & Crandall, 1998 )软件计算出的GTR+G模型; 其次, 用BESAT v1.7软件包中BEAUti生成.xml文件, 对所有21个物种拼接好的片段计算物种分化时间, 以Sanango racemosum (57 Ma) (Perret et al, 2013 )来限制某一分支出现的时间。设置的主要参数为: 核苷酸替代模型为GTR, 选择Gamma+Invariant位点, Gamma型分布及4种变化速率类型, 分子钟模型采用不相关对数松弛分子钟(uncorrelated lognormal relaxed molecular clock model), 树的后验拓扑结构为Yule speciation。独立计算2次, 每次运算8,000万代, 每1,000代取样1次, 舍弃10%的树(8,000棵) (Perret et al, 2013 )。最后, 通过TRACER v1.5 (http: //beast.bio.ed.ac.uk/Tracer/)检验相同分布的收敛性, 确保样本足够大小, 用TreeAnnotator v1.7.4软件建树, 所得的MCC树用FigTree v.1.3.1 (htpp://tree.bio. Ed.ac.uk/software/figtree/)打开, 将具有平均分化时间和95%置信区间的树以.pdf文件形式导出。
1.4 物种分化与花部特征和生境类型的关系
为分析海南岛苦苣苔科植物的物种分化是否伴随着花部特征与生境类型的适应变化, 所有21个物种的主要花部特征如花对称性以及生境(石灰岩地貌、海拔高度)等数据也标注在系统发育树上进行比对。
2 结果
2.1 物种多样性
经过面积归一化处理, 海南岛苦苣苔科植物的物种密度为18.985, 略高于越南(17.175)。海南岛苦苣苔科物种特有指数为0.396 (表1 ), 仅次于最高的广西(0.417), 略高于物种多样性高达236 (其中特有种106)的云南(0.222)。
海南岛与广东省共有种有12种, 物种相似性Jaccard指数达0.167, 远高于贵州(0.052)、越南(0.050)、广西(0.044)、云南(0.040)等地区与海南岛的物种相似性(表1 )。
2.2 地理分布格局
2.2.1 水平方向
海南岛苦苣苔科物种多样性最高的地区是五指山(15种)、鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种); 特有种分布最多的为五指山(6种)和尖峰岭(5种) (图1 )。红花芒毛苣苔、吊石苣苔(Lysionotus pauciflorus )、椭圆线柱苣苔(Rhynchotechum ellipticum )和冠萼线柱苣苔(R. formosanum )在海南岛各大山区均有分布, 其余的大多数苦苣苔科物种仅在五指山、鹦哥岭和尖峰岭等少数山区有分布。
图1 海南岛地形及本文研究的山峰
Fig. 1 Topography of Hainan Island and the mountains studied
海南岛的8个特有种中, 烟叶唇柱苣苔、盾叶苣苔、毛花马铃苣苔、黄花马铃苣苔在整个海南岛的各山区均有分布。而其他特有种分布非常狭窄, 如扁蒴苣苔仅在五指山有发现, 昌江蛛毛苣苔(Paraboea changjiangensis )与海南蛛毛苣苔仅分布在俄贤岭岩溶地貌上, 毛花马铃苣苔变种锈毛马铃苣苔(O. dasyantha var. ferruginose )只在五指山、吊罗山等零星分布。
2.2.2 垂直方向
海南岛苦苣苔科植物在600-800 m的中海拔区域分布最为集中, 然后向高海拔和低海拔依次递减。特有种的多样性在中海拔以及高海拔(1,600- 1,800 m)出现了两个峰值, 如五指山、鹦哥岭和尖峰岭(图2 )。芒毛苣苔(Aeschynanthus bracteatus )、横蒴苣苔(Beccarinda tonkinensis )、椭圆线柱苣苔和十字苣苔(Stauranthera umbrosa )等物种的海拔分布广, 从沟谷到山顶都有分布。海拔分布最低的物种有网脉蛛毛苣苔(Paraboea dictyoneura )和异色线柱苣苔(Rhynchotechum discolor ), 可低至海拔200 m。
图2 海南岛苦苣苔科物种多样性的垂直分布。□所有物种; ■ 特有种。
Fig. 2 Altitudinal distribution of species diversity of Gesneriaceae on Hainan Island. □ All species, ■ Hainan-endemic species.
海南岛苦苣苔科特有物种的分布海拔集中在高海拔(> 1200 m)和中海拔(600-1200 m)区域(图2 ), 在石灰岩分布的物种并不多(图4 ), 但4个海南岛特有种有3个是石灰岩专性或兼性物种。
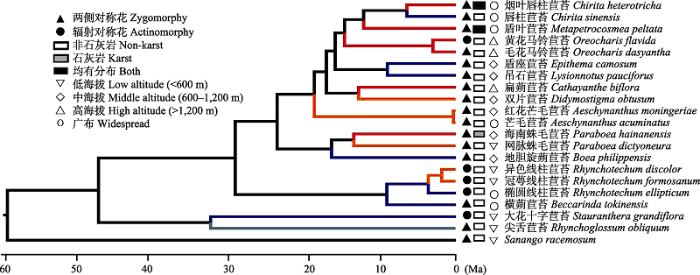
图3 根据ITS和trn L-F序列建立的海南岛苦苣苔科物种系统发育树。上方的地图显示了苦苣苔亚科在亚洲的地理分布, 示海南岛位于分布区中心。分支上的数值表示支持率。
Fig. 3 Phylogenetic tree of Hainan Gesneriaceae based on ITS and trn L-F. The insert map shows the geographical distribution of Asian Gesneriaceae. Note that Hainan Island locates at the center of the distribution range. The values above branches represent Bayesian posterior probabilities.
图4 海南岛苦苣苔科植物花对称性、分布生境与海拔的转变。分支颜色含义同图3 。
Fig. 4 Transitions of floral symmetry, habitat type and distribution altitude of Gesneriaceae on Hainan Isand. See Fig. 3 for the indication of branch color.
2.3 物种系统发育关系
尖舌苣苔属(Rhynchoglossum )在海南岛苦苣苔科植物系统发育树上位于基部, 大概在49 Ma分化出来, 在32 Ma前后分化出十字苣苔属(Stauranthera ) (图3 )。线柱苣苔属(Rhynchotechum )和横蒴苣苔属(Beccarinda )亲缘关系较近, 形成另一支系。蛛毛苣苔属(Paraboea )和旋蒴苣苔属(Boea )则形成另外一支, 并在大约14 Ma分化出海南岛特有种海南蛛毛苣苔。
海南岛特有的2个单种属, 盾叶苣苔属(Metapetrocosmea )形成时间约在12 Ma, 扁蒴苣苔属(Cathayanthe )则在大约13 Ma从双片苣苔属分化出来(图3 )。
海南岛苦苣苔科植物的基部类群都是与东南亚共享的, 而海南岛-中国大陆分布的类群均处于系统树20 Ma以后的较近分化出的分支。
2.4 花部特征与生境类型的转变
从花的对称性来看, 海南岛苦苣苔科植物与整个唇形目一致, 两侧对称花是较为原始的状态, 只在十字苣苔属和线柱苣苔属物种以及黄花马铃苣苔中出现了辐射对称花(图4 )。
海南岛苦苣苔科植物与石灰岩地貌的相关性很小, 大多数物种都分布在非石灰岩地貌, 仅有海南蛛毛苣苔以及另一个因缺少相关DNA序列而未列入系统发育研究的昌江蛛毛苣苔是石灰岩地貌专性物种; 而烟叶唇柱苣苔和盾叶苣苔则广布于石灰岩与非石灰岩地貌(图4 )。
从分布海拔来看, 海南岛苦苣苔科的原始类群多分布在低海拔地区, 后期分化出来的支系出现向中、高海拔迁移的趋势, 而海南岛特有种则多集中在高海拔区域(图4 )。
3 讨论
3.1 物种多样性及其分布格局
海南岛苦苣苔科植物有24种, 其中8个特有种, 与邻近地区分布的苦苣苔科植物相比其丰富度较低。这可能是因为海南岛面积仅有3.54万km2 , 且是海岛, 是本文这些区域中面积最小的孤立区域。
虽然海南岛苦苣苔科物种丰富度较低, 但物种特有比例高达33% (8/24), 且经过面积归一化处理后得到的特有指数较高(0.396)。这些特有植物主要分布于海南岛中南部山区, 且物种数最多的山峰往往是海拔最高的, 如五指山(15种), 其次是鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种) (图1 )。海南岛特有种基本都分布在高海拔区域(图2 ), 如扁蒴苣苔、黄花马铃苣苔仅分布在海拔1,400 m以上。因此, 高山深谷形成的生境隔离可能促进了苦苣苔科的物种形成与维持(韦毅刚等, 2004 )。与近缘种相比, 海南岛特有种的海拔分布表现出向上迁移的趋势(图2 )。这表明高耸入云的山顶形成的“天岛” (sky islands; Knowles, 2001 ; Robin et al, 2015 )隔离作用较强, 可能是扩散能力较弱的苦苣苔科特有种得以形成与维持的重要原因。
海南岛苦苣苔科特有种多数分布广泛, 如烟叶唇柱苣苔、盾叶苣苔、毛花马铃苣苔、锈毛马铃苣苔和黄花马铃苣苔在海南岛主要山峰都有较多的自然分布。这主要是因为海南岛中南部山区连续性较高、生境破碎化程度较小, 而且这些物种适应性较强(Francisco-Ortega et al, 2010 ; 邢福武, 2012 )。如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 )。
3.2 物种迁移历史
在海南岛自然分布的苦苣苔科植物中, 分化时间最早的类群是尖舌苣苔属(图3 )。尖舌苣苔属间断分布于亚洲和中南美洲(李振宇, 1996 ), 约在49 Ma左右起源于南美, 之后扩散至亚洲的热带地区即现今的东南亚群岛一带(Perret et al, 2013 )。海南岛有着大量该科的基部类群如尖舌苣苔属、十字苣苔属、横蒴苣苔属以及线柱苣苔属(图3 ), 而且都与东南亚共享, 表明苦苣苔科植物在很早的时候就已从东南亚迁入了海南岛。海南岛的地质历史也证实了这种可能性, 海南岛所处的陆地在早期的时候(印度板块撞击欧亚板块之前)与中南半岛相连(Zhu, 2016 )。
海南岛-中国大陆共同分布的类群处于海南岛苦苣苔科系统发育树末端的较新分支, 表明海南岛苦苣苔科植物可能是在后期才与中国大陆有着较强的交流。海南岛两个特有属的形成时间约在12 Ma, 特有种也多集中在15-5 Ma才分化出来(图3 )。在15 Ma左右, 海南岛在印度板块撞击欧亚板块、中南半岛向南挤压的作用下从北部湾脱离出来并已远离周边大陆约100 km (Zhu, 2016 )。这种海岛隔离作用可能导致了海南岛苦苣苔科植物特有种的形成(Ling et al, 2017 )。
海南岛苦苣苔科植物与邻近的广东省有着最高的Jaccard相似性系数(表1 ), 这可能是因为上新世末的海退和晚更新世末海平面的下降, 导致海南岛与广东省的雷州半岛曾两度相连(Liang, 2013 ; Zhu, 2016 )。这个过程中, 广东地区的苦苣苔科植物如芒毛苣苔属、蛛毛苣苔属和线柱苣苔属等植物可能多次进入海南岛(图3 ), 造成两地苦苣苔科物种组成的相似性很高。
3.3 物种适应进化
苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 )。两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 )。两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 )。一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 )。本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 )。
虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 )。这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 )。
从海拔分布区域来看, 苦苣苔科原始类型大多分布在低海拔地区, 之后逐渐向中高海拔迁移, 并在这个过程中由于山顶“天岛”效应及河谷隔离作用而分化出了一些海南岛特有种(图4 ), 这说明在向中高海拔迁移的过程中, 山顶形成的孤岛效应以及山脉河谷的隔离作用也促进了海南岛特有种的形成与维持(邢福武, 1995 ; Ling et al, 2017 )。
亚洲的苦苣苔科植物是从南美扩散而来, 海南岛的苦苣苔科植物在历史上很可能最先来自亚洲典型热带地区如东南亚, 后期才与中国大陆发生多次、频繁的物种交流。在这期间, 由于海岛隔离与山顶“天岛”效应等的影响在12 Ma前后分化出了一些特有属和特有种。海南岛自然分布的24个苦苣苔科物种中, 既有与东南亚共享的, 也有与中国大陆共同分布的类群。因此, 海南岛可能在亚洲热带植物长距离迁移过程中起着一定的作用。由于亚洲热带植物多分布在隔离程度大的群岛、半岛及大陆边缘, 将来有必要结合岛屿生物地理学和分子生物学研究方法综合研究海南岛、东南亚群岛、中南半岛等地典型热带植物的迁移历史, 深入认识亚洲热带植物多样性形成与演化。
附录 Supplementary Material
附录1 海南岛苦苣苔科物种系统发育树所用的物种及其序列在GenBank的编号
Appendix 1 The species used to construct phylogenetic tree of Hainan Gesneriaceae with their accession numbers in GenBank
http://www.biodiversity-science.net/fileup/PDF/2016360-1.pdf
致谢: 感谢温放博士提供《华南苦苣苔科植物》和越南苦苣苔科植物资料, 史佑海副教授对初稿修改提出宝贵意见。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[9]
Jaccard P 1912 ) The distribution of the flora in the alpine zone
. New Phytologist , 11 (2 ), 37 -50 .
[本文引用: 1]
[10]
Jiang YX 1988 ) A background analysis of the flora and the characteristics of tropical vegetation in Hainan Island
. Natural Sciences of Journal of Hainan University , 6 (3 ), 1 -8 . (in Chinese with English abstract)
[蒋有绪 (1988 ) 海南岛植物区系与热带植被性质的背景分析
. 海南大学学报(自然科学版) , 6 (3 ), 1 -8 .]
[11]
Knowles LL 2001 ) Genealogical portraits of speciation in montane grasshoppers (genus Malanoplus) from the sky island of the Rocky Mountains
. Proceeding of the Royal Society B: Biological Sciences , 268 , 319 -324 .
[本文引用: 1]
[12]
Kumar S Nei M Dudley J Tamura K 2008 ) MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences
. Briefings in Bioinformatics , 9 , 299 -306 .
[本文引用: 1]
[13]
Li R Sun H 2017 ) Phylofloristics: a case study from Yunnan, China
. Biodiversity Science , 25 , 195 -203 . (in Chinese with English abstract)
[本文引用: 1]
[李嵘 , 孙航 (2017 ) 植物系统发育区系地理学研究: 以云南植物区系为例
. 生物多样性 , 25 , 195 -203 .]
[本文引用: 1]
[14]
Li ZY 1996 ) The geographical distribution of the subfamily Cyrtanroideae Endl. Emend. Burtt (Gesneriaceae)
. Acta Phytotaxonmica Sinica , 34 , 341 -360 . (in Chinese with English abstract)
[本文引用: 3]
[李振宇 (1996 ) 苦苣苔亚科的地理分布
. 植物分类学报 , 34 , 341 -360 .]
[本文引用: 3]
[15]
Li ZY Wang YZ 2005 ) Plants of Gesneriaceae in China . Henan Science and Technology Publishing House , Zhengzhou . (in Chinese)
[本文引用: 2]
[李振宇 , 王印政 (2005 ) 中国苦苣苔科植物 . 河南科学技术出版社 , 郑州 .]
[本文引用: 2]
[16]
Liang GH 2013 ) Eight evidences on Hainan Island separating from China’s Beibuwan Gulf with rotation
. Acta Geologica Sinica , 87 , 73 -76 .
[本文引用: 1]
[17]
Ling SJ Meng QW Tang L Ren MX 2017 ) Pollination syndromes of Chinese Gesneriaceae: a comparative study between Hainan Island and neighboring regions
. Botanical Review , 83 , 59 -74 .
[本文引用: 2]
[18]
Losos JB Ricklefs RE 2010 ) The Theory of Island Biogeography Revisited . Princeton University Press , New Jersey .
[本文引用: 1]
[19]
MacArthur RH Wilson EO 1967 ) The Theory of Island Biogeography . Princeton University Press , New Jersey .
[本文引用: 1]
[20]
Martén-Rodríguez SA Quesada M Castro AA Lopezaraiza-Mikel M Fenster CB 2015 ) A comparison of reproductive strategies between island and mainland Caribbean Gesneriaceae
. Journal of Ecology , 103 , 1190 -1204 .
[本文引用: 2]
[21]
Myers N Mittermeier RA Mittermeier CG Da FG Kent J 2000 ) Biodiversity hotspots for conservation priorities
. Nature , 403 , 853 -858 .
[本文引用: 1]
[22]
Möller M Forrest A Wei YG Weber A 2010 ) A molecular phylogenetic assessment of the advanced Asiatic and Malesian didymocarpoid Gesneriaceae with focus on non-monophyletic and monotypic genera
. Plant Systematics and Evolution , 292 , 223 -248 .
[本文引用: 1]
[23]
Möller M Wei YG Wen F Clark JL Weber A 2016 ) You win some you lose some: updated generic delineations and classification of Gesneriaceae—implications for the family in China
. Guihaia , 36 , 44 -60 .
[本文引用: 1]
[24]
Perret M Chautems A Araujo AOD Salamin N 2013 ) Temporal and spatial origin of Gesneriaceae in the New World inferred from plastid DNA sequences
. Botanical Journal of the Linnean Society , 171 , 61 -79 .
[本文引用: 4]
[25]
Pham HH 1991 ) An Illustrated Flora in Vietnam
. Mekong Printing, Can Tho City .
[26]
Posada D Crandall KA 1998 ) MODELTEST: testing the model of DNA substitution
. Bioinformatics , 14 , 817 -818 .
[本文引用: 1]
[27]
Robin VV Vishnudas CK Gupta P Ramakrishnan U 2015 ) Deep and wide valleys drive nested phylogeographic patterns across a montane bird community
. Proceedings of the Royal Society B: Biological Sciences , 282 , 20150861 .
[本文引用: 1]
[28]
Su ZY Zhang HD 1994 ) The relationship between Guangxi’s flora and each of the adjacent floras
. Journal of South China Agricultural University , 15 (2 ), 38 -43 . (in Chinese with English abstract)
[本文引用: 1]
[苏志尧 , 张宏达 (1994 ) 广西植物区系与邻近地区植物区系的关系
. 华南农业大学学报 , 15 (2 ), 38 -43 .]
[本文引用: 1]
[29]
Tang ZY Wang ZH Zheng CY Fang JY 2006 ) Biodiversity in China’s mountains
. Frontiers in Ecology and the Environment , 4 , 347 -352 .
[本文引用: 1]
[30]
Vaidya G Lohman DJ Meier R 2011 ) SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information
. Cladistics , 27 , 171 -180 .
[本文引用: 1]
[31]
Wang FG Qin XS Chen HF Zhang RJ Liu DM Xing FW 2006 ) Endemic plants in limestone region on Hainan Island
. Journal of Tropical and Subtropical Botany , 14 , 45 -54 . (in Chinese with English abstract)
[本文引用: 2]
[王发国 , 秦新生 , 陈红锋 , 张荣京 , 刘东明 , 邢福武 (2006 ) 海南岛石灰岩特有植物的初步研究
. 热带亚热带植物学报 , 14 , 45 -54 .]
[本文引用: 2]
[32]
Wang HS 2000 ) The nature of China’s flora and the relationships between its different elements
. Acta Botanica Yunnanica , 22 , 119 -126 . (in Chinese with English abstract)
[王荷生 (2000 ) 中国植物区系的性质和各成分间的关系
. 云南植物研究 , 22 , 119 -126 .]
[1]
Barrett SCH Emerson B Mallet J 1996 ) The reproductive biology and genetics of island plants
. Philosophical Transactions of the Royal Society B: Biological Sciences , 351 , 725 -733 .
[本文引用: 1]
[2]
Bramwell D Caujapé-Castells J 2011 ) The Biology of Island Floras . Cambridge University Press , Cambridge .
[本文引用: 1]
[33]
Wang YZ Liang RH Wang BH Li JM Qiu ZY Weber A 2010 ) Origin and phylogenetic relationships of the Old World Gesneriaceae with actinomorphic flowers inferred from ITS and trnL-trnF sequences
. Taxon , 59 , 1044 -1052 .
[本文引用: 3]
[34]
Wei YG Zhong SH Wen HQ 2004 ) Studies of the flora and ecology Gesneriaceae in Guangxi Province
. Acta Botanica Yunnanica , 26 , 173 -182 . (in Chinese with English abstract)
[本文引用: 3]
[3]
Chen SB Ouyang ZY Xu WH Xiao Y 2010 ) A review of beta diversity studies
. Biodiversity Science , 18 , 323 -335 . (in Chinese with English abstract)
[陈圣宾 , 欧阳志云 , 徐卫华 , 肖燚 (2010 ) Beta多样性研究进展
. 生物多样性 , 18 , 323 -335 .]
[34]
[韦毅刚 , 钟树华 , 文和群 (2004 ) 广西苦苣苔科植物区系和生态特点研究
. 云南植物研究 , 26 , 173 -182 .]
[本文引用: 3]
[35]
Wei YG 2010 ) Gesneriaceae of South China . Guangxi Science and Technology Publishing House , Nanning . (in Chinese)
[本文引用: 3]
[4]
Ding L Tang WX Luo WH Pan B Wei YG Huang SX 2010 ) Floristic characteristics of endemic plants to Guangxi
. Guihaia , 30 , 202 -208 . (in Chinese with English abstract)
[本文引用: 1]
[丁莉 , 唐文秀 , 骆文华 , 盘波 , 韦毅刚 , 黄仕训 (2010 ) 广西特有植物区系特征研究
. 广西植物 , 30 , 202 -208 .]
[本文引用: 1]
[35]
[韦毅刚 (2010 ) 华南苦苣苔科植物 . 广西科学技术出版社 , 南宁 .]
[本文引用: 3]
[36]
White TJ Bruns T Lee S Taylor J 1990 ) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics
. In: PCR Protocols: A Guide to Methods and Applications (eds Innis MA, Gelfand DH, Sninsky JJ, White TJ) , pp. 315 -322. Academic Press, New York.
[本文引用: 1]
[5]
Doley JJ Doley JL 1987 ) A rapid DNA isolation procedure for small quantities of fresh leaf tissue
. Phytochemical Bulletin , 19 , 11 -15 .
[本文引用: 1]
[6]
Drummond AJ Rambaut A 2007 ) BEAST: Bayesian evolutionary analysis by sampling trees
. BMC Evolutionary Biology , 7 , 214 .
[本文引用: 1]
[37]
Wu ZY Sun H Zhou ZK Li DZ Peng H 2011 ) Floristics of Seed Plants from China . Science Press , Beijing . (in Chinese)
[吴征镒 , 孙航 , 周浙昆 , 李德铢 , 彭华 (2011 ) 中国种子植物区系地理 . 科学出版社 , 北京.]
[38]
Xing FW 1995 ) Endemic plants of Hainan Island
. Journal of Tropical and Subtropical Botany , 3 , 1 -12 . (in Chinese with English abstract)
[本文引用: 1]
[邢福武 (1995 ) 海南岛特有植物的研究
. 热带亚热带植物学报 , 3 , 1 -12 .]
[本文引用: 1]
[39]
Xing FW 2012 ) Inventory of Plant Species Diversity of Hainan . Huazhong University of Science and Technology Press , Wuhan . (in Chinese)
[本文引用: 3]
[7]
Francisco-Ortega J Wang ZS Wang FG Xing FW Liu H Xiu H Xu WX Luo YB Song XQ Gale S Boufford DE Maunder M An SQ 2010 ) Seed plant endemism on Hainan Island: a framework for conservation action
. Botanical Review , 76 , 346 -376 .
[本文引用: 2]
[8]
Gong YB Huang SQ 2009 ) Floral symmetry: pollinator-mediated stabilizing selection on flower size in bilateral species
. Proceedings of the Royal Society B: Biological Sciences , 276 , 4013 -4020 .
[本文引用: 1]
[39]
[邢福武 (2012 ) 海南植物物种多样性编目 . 华中科技大学出版社 , 武汉 .]
[本文引用: 3]
[40]
Yang XB 2013 ) Flora of Hainan . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[杨小波 (2013 ) 海南植物名录 . 科学出版社 , 北京 .]
[本文引用: 1]
[41]
Zhang CC Liu LF 1983 ) The angiospermous flora of Hainan
. Journal of Sun Yat-Sen University , (3 ), 67 -73 .
(in Chinese with English abstract) [张超常 , 刘兰芳 (1983 ) 海南岛被子植物区系
. 中山大学学报 , (3 ), 67 -73 .]
[42]
Zhang HD 1962 ) Characteristics of Guangdong flora
. Acta Scientiarum Naturalium Universitatis Sunyatseni , (1 ), 3 -36 . (in Chinese with English abstract)
[张宏达 (1962 ) 广东植物区系的特点
. 中山大学学报(自然科学版) , (1 ) , 3 -36 .]
[43]
Zhang HD 2001 ) The diversity of the Hainan Flora
. Ecological Science , 20 , 1 -10 . (in Chinese with English abstract)
[张宏达 (2001 ) 海南植物区系的多样性
. 生态科学 , 20 , 1 -10 .]
[44]
Zhu H 2016 ) Biogeographical evidences help revealing the origin of Hainan Island
. PLoS ONE , 11 , e0151941 .
[本文引用: 4]
The distribution of the flora in the alpine zone
1
1912
... 根据陈圣宾等(2010), 我们使用目前广泛应用的Jaccard相似性指数(Jaccard, 1912 )来度量海南岛与其邻近地区的物种组成相似性程度: C j = c / (a + b - c ).其中, a 为海南岛苦苣苔科物种数, b 为邻近某地区的物种数, c 为两地共有物种数. ...
Genealogical portraits of speciation in montane grasshoppers (genus Malanoplus) from the sky island of the Rocky Mountains
1
2001
... 虽然海南岛苦苣苔科物种丰富度较低, 但物种特有比例高达33% (8/24), 且经过面积归一化处理后得到的特有指数较高(0.396).这些特有植物主要分布于海南岛中南部山区, 且物种数最多的山峰往往是海拔最高的, 如五指山(15种), 其次是鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种) (图1 ).海南岛特有种基本都分布在高海拔区域(图2 ), 如扁蒴苣苔、黄花马铃苣苔仅分布在海拔1,400 m以上.因此, 高山深谷形成的生境隔离可能促进了苦苣苔科的物种形成与维持(韦毅刚等, 2004 ).与近缘种相比, 海南岛特有种的海拔分布表现出向上迁移的趋势(图2 ).这表明高耸入云的山顶形成的“天岛” (sky islands; Knowles, 2001 ; Robin et al, 2015 )隔离作用较强, 可能是扩散能力较弱的苦苣苔科特有种得以形成与维持的重要原因. ...
MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences
1
2008
... 本研究中, 除通过测序获得6个苦苣苔科物种DNA序列外, 海南岛分布的苦苣苔科有14个物种(包括海南特有种扁蒴苣苔(Cathayanthe biflora ))的ITS1/2和trn L-F序列都可从GenBank获得(附录1).为标定物种分化时间, 补充与亚洲苦苣苔科亲缘关系最近的、物种分化时间(57 Ma)已知的Sanango racemosum (Perret et al, 2013 )一起建树.因此, 一共有21个海南岛苦苣苔科植物的DNA序列(附录1)用于建立系统发育树.各物种的核基因ITS1/2片段和叶绿体trn L-F片段用SequenceMatrix软件(Vaidya et al, 2011 )拼接; 用MEGA 6.0软件(Kumar et al, 2008 )整合拼接好的片段. ...
植物系统发育区系地理学研究: 以云南植物区系为例
1
2017
... 海南岛生物多样性的起源及其与周边地区的联系一直存在争议(Francisco-Ortega et al, 2010 ; Zhu, 2016 ).张宏达(1962, 2001)指出, 海南岛、台湾岛、广东、广西、福建南部、云南东南部及中南半岛北部等地区的植物均属华南亚区; 张超常和刘兰芳(1983)认为, 海南岛属于华夏植物区系; 而蒋有绪(1988)认为, 海南岛的植物区系应属印度-马来西亚区系.王荷生(2000)、吴征镒等(2011)以及Zhu (2016)基本赞同蒋有绪的结论, 认为海南岛及邻近地区植物区系应属马来西亚亚区.然而, 传统植物区系地理学研究主要以植物区系的分类群组成及其分布区类型的分析为主, 相对忽视了进化历史的分析(李嵘和孙航, 2017 ). ...
植物系统发育区系地理学研究: 以云南植物区系为例
1
2017
... 海南岛生物多样性的起源及其与周边地区的联系一直存在争议(Francisco-Ortega et al, 2010 ; Zhu, 2016 ).张宏达(1962, 2001)指出, 海南岛、台湾岛、广东、广西、福建南部、云南东南部及中南半岛北部等地区的植物均属华南亚区; 张超常和刘兰芳(1983)认为, 海南岛属于华夏植物区系; 而蒋有绪(1988)认为, 海南岛的植物区系应属印度-马来西亚区系.王荷生(2000)、吴征镒等(2011)以及Zhu (2016)基本赞同蒋有绪的结论, 认为海南岛及邻近地区植物区系应属马来西亚亚区.然而, 传统植物区系地理学研究主要以植物区系的分类群组成及其分布区类型的分析为主, 相对忽视了进化历史的分析(李嵘和孙航, 2017 ). ...
苦苣苔亚科的地理分布
3
1996
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 在海南岛自然分布的苦苣苔科植物中, 分化时间最早的类群是尖舌苣苔属(图3 ).尖舌苣苔属间断分布于亚洲和中南美洲(李振宇, 1996 ), 约在49 Ma左右起源于南美, 之后扩散至亚洲的热带地区即现今的东南亚群岛一带(Perret et al, 2013 ).海南岛有着大量该科的基部类群如尖舌苣苔属、十字苣苔属、横蒴苣苔属以及线柱苣苔属(图3 ), 而且都与东南亚共享, 表明苦苣苔科植物在很早的时候就已从东南亚迁入了海南岛.海南岛的地质历史也证实了这种可能性, 海南岛所处的陆地在早期的时候(印度板块撞击欧亚板块之前)与中南半岛相连(Zhu, 2016 ). ...
苦苣苔亚科的地理分布
3
1996
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 在海南岛自然分布的苦苣苔科植物中, 分化时间最早的类群是尖舌苣苔属(图3 ).尖舌苣苔属间断分布于亚洲和中南美洲(李振宇, 1996 ), 约在49 Ma左右起源于南美, 之后扩散至亚洲的热带地区即现今的东南亚群岛一带(Perret et al, 2013 ).海南岛有着大量该科的基部类群如尖舌苣苔属、十字苣苔属、横蒴苣苔属以及线柱苣苔属(图3 ), 而且都与东南亚共享, 表明苦苣苔科植物在很早的时候就已从东南亚迁入了海南岛.海南岛的地质历史也证实了这种可能性, 海南岛所处的陆地在早期的时候(印度板块撞击欧亚板块之前)与中南半岛相连(Zhu, 2016 ). ...
2
2005
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
2
2005
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
Eight evidences on Hainan Island separating from China’s Beibuwan Gulf with rotation
1
2013
... 海南岛苦苣苔科植物与邻近的广东省有着最高的Jaccard相似性系数(表1 ), 这可能是因为上新世末的海退和晚更新世末海平面的下降, 导致海南岛与广东省的雷州半岛曾两度相连(Liang, 2013 ; Zhu, 2016 ).这个过程中, 广东地区的苦苣苔科植物如芒毛苣苔属、蛛毛苣苔属和线柱苣苔属等植物可能多次进入海南岛(图3 ), 造成两地苦苣苔科物种组成的相似性很高. ...
Pollination syndromes of Chinese Gesneriaceae: a comparative study between Hainan Island and neighboring regions
2
2017
... 海南岛-中国大陆共同分布的类群处于海南岛苦苣苔科系统发育树末端的较新分支, 表明海南岛苦苣苔科植物可能是在后期才与中国大陆有着较强的交流.海南岛两个特有属的形成时间约在12 Ma, 特有种也多集中在15-5 Ma才分化出来(图3 ).在15 Ma左右, 海南岛在印度板块撞击欧亚板块、中南半岛向南挤压的作用下从北部湾脱离出来并已远离周边大陆约100 km (Zhu, 2016 ).这种海岛隔离作用可能导致了海南岛苦苣苔科植物特有种的形成(Ling et al, 2017 ). ...
... 从海拔分布区域来看, 苦苣苔科原始类型大多分布在低海拔地区, 之后逐渐向中高海拔迁移, 并在这个过程中由于山顶“天岛”效应及河谷隔离作用而分化出了一些海南岛特有种(图4 ), 这说明在向中高海拔迁移的过程中, 山顶形成的孤岛效应以及山脉河谷的隔离作用也促进了海南岛特有种的形成与维持(邢福武, 1995 ; Ling et al, 2017 ). ...
1
2010
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
1
1967
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
A comparison of reproductive strategies between island and mainland Caribbean Gesneriaceae
2
2015
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
... ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
Biodiversity hotspots for conservation priorities
1
2000
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
A molecular phylogenetic assessment of the advanced Asiatic and Malesian didymocarpoid Gesneriaceae with focus on non-monophyletic and monotypic genera
1
2010
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
You win some you lose some: updated generic delineations and classification of Gesneriaceae—implications for the family in China
1
2016
... 虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 ).这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
Temporal and spatial origin of Gesneriaceae in the New World inferred from plastid DNA sequences
4
2013
... 本研究中, 除通过测序获得6个苦苣苔科物种DNA序列外, 海南岛分布的苦苣苔科有14个物种(包括海南特有种扁蒴苣苔(Cathayanthe biflora ))的ITS1/2和trn L-F序列都可从GenBank获得(附录1).为标定物种分化时间, 补充与亚洲苦苣苔科亲缘关系最近的、物种分化时间(57 Ma)已知的Sanango racemosum (Perret et al, 2013 )一起建树.因此, 一共有21个海南岛苦苣苔科植物的DNA序列(附录1)用于建立系统发育树.各物种的核基因ITS1/2片段和叶绿体trn L-F片段用SequenceMatrix软件(Vaidya et al, 2011 )拼接; 用MEGA 6.0软件(Kumar et al, 2008 )整合拼接好的片段. ...
... 用BEAST v1.7.5 (Drummond & Rambaut, 2007 )建立海南岛苦苣苔科植物的系统发育树.首先, 碱基替代模型为根据MODELTEST v3.6 (Posada & Crandall, 1998 )软件计算出的GTR+G模型; 其次, 用BESAT v1.7软件包中BEAUti生成.xml文件, 对所有21个物种拼接好的片段计算物种分化时间, 以Sanango racemosum (57 Ma) (Perret et al, 2013 )来限制某一分支出现的时间.设置的主要参数为: 核苷酸替代模型为GTR, 选择Gamma+Invariant位点, Gamma型分布及4种变化速率类型, 分子钟模型采用不相关对数松弛分子钟(uncorrelated lognormal relaxed molecular clock model), 树的后验拓扑结构为Yule speciation.独立计算2次, 每次运算8,000万代, 每1,000代取样1次, 舍弃10%的树(8,000棵) (Perret et al, 2013 ).最后, 通过TRACER v1.5 (http: //beast.bio.ed.ac.uk/Tracer/)检验相同分布的收敛性, 确保样本足够大小, 用TreeAnnotator v1.7.4软件建树, 所得的MCC树用FigTree v.1.3.1 (htpp://tree.bio. Ed.ac.uk/software/figtree/)打开, 将具有平均分化时间和95%置信区间的树以.pdf文件形式导出. ...
... )来限制某一分支出现的时间.设置的主要参数为: 核苷酸替代模型为GTR, 选择Gamma+Invariant位点, Gamma型分布及4种变化速率类型, 分子钟模型采用不相关对数松弛分子钟(uncorrelated lognormal relaxed molecular clock model), 树的后验拓扑结构为Yule speciation.独立计算2次, 每次运算8,000万代, 每1,000代取样1次, 舍弃10%的树(8,000棵) (Perret et al, 2013 ).最后, 通过TRACER v1.5 (http: //beast.bio.ed.ac.uk/Tracer/)检验相同分布的收敛性, 确保样本足够大小, 用TreeAnnotator v1.7.4软件建树, 所得的MCC树用FigTree v.1.3.1 (htpp://tree.bio. Ed.ac.uk/software/figtree/)打开, 将具有平均分化时间和95%置信区间的树以.pdf文件形式导出. ...
... 在海南岛自然分布的苦苣苔科植物中, 分化时间最早的类群是尖舌苣苔属(图3 ).尖舌苣苔属间断分布于亚洲和中南美洲(李振宇, 1996 ), 约在49 Ma左右起源于南美, 之后扩散至亚洲的热带地区即现今的东南亚群岛一带(Perret et al, 2013 ).海南岛有着大量该科的基部类群如尖舌苣苔属、十字苣苔属、横蒴苣苔属以及线柱苣苔属(图3 ), 而且都与东南亚共享, 表明苦苣苔科植物在很早的时候就已从东南亚迁入了海南岛.海南岛的地质历史也证实了这种可能性, 海南岛所处的陆地在早期的时候(印度板块撞击欧亚板块之前)与中南半岛相连(Zhu, 2016 ). ...
An Illustrated Flora in Vietnam
1991
MODELTEST: testing the model of DNA substitution
1
1998
... 用BEAST v1.7.5 (Drummond & Rambaut, 2007 )建立海南岛苦苣苔科植物的系统发育树.首先, 碱基替代模型为根据MODELTEST v3.6 (Posada & Crandall, 1998 )软件计算出的GTR+G模型; 其次, 用BESAT v1.7软件包中BEAUti生成.xml文件, 对所有21个物种拼接好的片段计算物种分化时间, 以Sanango racemosum (57 Ma) (Perret et al, 2013 )来限制某一分支出现的时间.设置的主要参数为: 核苷酸替代模型为GTR, 选择Gamma+Invariant位点, Gamma型分布及4种变化速率类型, 分子钟模型采用不相关对数松弛分子钟(uncorrelated lognormal relaxed molecular clock model), 树的后验拓扑结构为Yule speciation.独立计算2次, 每次运算8,000万代, 每1,000代取样1次, 舍弃10%的树(8,000棵) (Perret et al, 2013 ).最后, 通过TRACER v1.5 (http: //beast.bio.ed.ac.uk/Tracer/)检验相同分布的收敛性, 确保样本足够大小, 用TreeAnnotator v1.7.4软件建树, 所得的MCC树用FigTree v.1.3.1 (htpp://tree.bio. Ed.ac.uk/software/figtree/)打开, 将具有平均分化时间和95%置信区间的树以.pdf文件形式导出. ...
Deep and wide valleys drive nested phylogeographic patterns across a montane bird community
1
2015
... 虽然海南岛苦苣苔科物种丰富度较低, 但物种特有比例高达33% (8/24), 且经过面积归一化处理后得到的特有指数较高(0.396).这些特有植物主要分布于海南岛中南部山区, 且物种数最多的山峰往往是海拔最高的, 如五指山(15种), 其次是鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种) (图1 ).海南岛特有种基本都分布在高海拔区域(图2 ), 如扁蒴苣苔、黄花马铃苣苔仅分布在海拔1,400 m以上.因此, 高山深谷形成的生境隔离可能促进了苦苣苔科的物种形成与维持(韦毅刚等, 2004 ).与近缘种相比, 海南岛特有种的海拔分布表现出向上迁移的趋势(图2 ).这表明高耸入云的山顶形成的“天岛” (sky islands; Knowles, 2001 ; Robin et al, 2015 )隔离作用较强, 可能是扩散能力较弱的苦苣苔科特有种得以形成与维持的重要原因. ...
广西植物区系与邻近地区植物区系的关系
1
1994
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
广西植物区系与邻近地区植物区系的关系
1
1994
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
Biodiversity in China’s mountains
1
2006
... 为确定海南岛苦苣苔科植物的物种多样性水平及其与周边地区的联系, 本文收集了海南岛、云南、广西、贵州、广东及邻国越南苦苣苔科物种多样性数据(Pham, 1991; 李振宇和王印政, 2005; 韦毅刚, 2010; 表1 ).为降低这6个地区因为面积差异过大可能产生的“抽样效应” (sampling effects), 本文参照Tang等(2006) , 通过面积归一化(自然对数转换)之后计算各地区的物种密度(species density, SD)和特有指数(endemic index, EI): ...
SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information
1
2011
... 本研究中, 除通过测序获得6个苦苣苔科物种DNA序列外, 海南岛分布的苦苣苔科有14个物种(包括海南特有种扁蒴苣苔(Cathayanthe biflora ))的ITS1/2和trn L-F序列都可从GenBank获得(附录1).为标定物种分化时间, 补充与亚洲苦苣苔科亲缘关系最近的、物种分化时间(57 Ma)已知的Sanango racemosum (Perret et al, 2013 )一起建树.因此, 一共有21个海南岛苦苣苔科植物的DNA序列(附录1)用于建立系统发育树.各物种的核基因ITS1/2片段和叶绿体trn L-F片段用SequenceMatrix软件(Vaidya et al, 2011 )拼接; 用MEGA 6.0软件(Kumar et al, 2008 )整合拼接好的片段. ...
海南岛石灰岩特有植物的初步研究
2
2006
... 虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 ).这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
... )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
海南岛石灰岩特有植物的初步研究
2
2006
... 虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 ).这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
... )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
The reproductive biology and genetics of island plants
1
1996
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
1
2011
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
Origin and phylogenetic relationships of the Old World Gesneriaceae with actinomorphic flowers inferred from ITS and trnL-trnF sequences
3
2010
... 在海南岛全岛范围采集了海南特有种烟叶唇柱苣苔(Chirita heterotricha )、海南蛛毛苣苔(Paraboea hainanensis )、盾叶苣苔(Metapetrocosmea peltata )、黄花马铃苣苔(Oreocharis flavida )、毛花马铃苣苔(O. dasyantha )以及广布种红花芒毛苣苔(Aeschynanthus moningerae )各不少于5个个体的幼叶, 经变色硅胶干燥, 然后用改良的CTAB (Cetyl Trimethyl Ammonium Bromide, 十六烷基三甲基溴化铵)法提取叶片总DNA (Doyle & Doyle, 1987 ).选择变异程度大的叶绿体基因片段trn L-F (Wang et al, 2010 )和核基因片段ITS1/2 (White et al, 1990 )进行扩增和序列分析(表2 ). ...
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
... ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
广西苦苣苔科植物区系和生态特点研究
3
2004
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 虽然海南岛苦苣苔科物种丰富度较低, 但物种特有比例高达33% (8/24), 且经过面积归一化处理后得到的特有指数较高(0.396).这些特有植物主要分布于海南岛中南部山区, 且物种数最多的山峰往往是海拔最高的, 如五指山(15种), 其次是鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种) (图1 ).海南岛特有种基本都分布在高海拔区域(图2 ), 如扁蒴苣苔、黄花马铃苣苔仅分布在海拔1,400 m以上.因此, 高山深谷形成的生境隔离可能促进了苦苣苔科的物种形成与维持(韦毅刚等, 2004 ).与近缘种相比, 海南岛特有种的海拔分布表现出向上迁移的趋势(图2 ).这表明高耸入云的山顶形成的“天岛” (sky islands; Knowles, 2001 ; Robin et al, 2015 )隔离作用较强, 可能是扩散能力较弱的苦苣苔科特有种得以形成与维持的重要原因. ...
... 虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 ).这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
广西苦苣苔科植物区系和生态特点研究
3
2004
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 虽然海南岛苦苣苔科物种丰富度较低, 但物种特有比例高达33% (8/24), 且经过面积归一化处理后得到的特有指数较高(0.396).这些特有植物主要分布于海南岛中南部山区, 且物种数最多的山峰往往是海拔最高的, 如五指山(15种), 其次是鹦哥岭(11种)、尖峰岭(11种)和霸王岭(10种) (图1 ).海南岛特有种基本都分布在高海拔区域(图2 ), 如扁蒴苣苔、黄花马铃苣苔仅分布在海拔1,400 m以上.因此, 高山深谷形成的生境隔离可能促进了苦苣苔科的物种形成与维持(韦毅刚等, 2004 ).与近缘种相比, 海南岛特有种的海拔分布表现出向上迁移的趋势(图2 ).这表明高耸入云的山顶形成的“天岛” (sky islands; Knowles, 2001 ; Robin et al, 2015 )隔离作用较强, 可能是扩散能力较弱的苦苣苔科特有种得以形成与维持的重要原因. ...
... 虽然石灰岩地貌造成的生境隔离作用被认为是苦苣苔科物种分化的一个极其重要的因素(韦毅刚等, 2004 ; Möller et al, 2016 ), 但海南岛的石灰岩地貌总面积仅约400 km2 , 只占海南岛面积的1%左右, 零星分布于海南岛西部、南部的部分山地(王发国等, 2006 ).这些分散的小面积石灰岩地貌造成的生境隔离作用可能很弱, 使得海南岛特有的石灰岩专性物种非常少, 仅有海南蛛毛苣苔(图4 )和昌江蛛毛苣苔出现在海南岛石灰岩地貌最集中分布的俄贤岭(王发国等, 2006 ). ...
3
2010
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
广西特有植物区系特征研究
1
2010
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
广西特有植物区系特征研究
1
2010
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
3
2010
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics
1
1990
... 在海南岛全岛范围采集了海南特有种烟叶唇柱苣苔(Chirita heterotricha )、海南蛛毛苣苔(Paraboea hainanensis )、盾叶苣苔(Metapetrocosmea peltata )、黄花马铃苣苔(Oreocharis flavida )、毛花马铃苣苔(O. dasyantha )以及广布种红花芒毛苣苔(Aeschynanthus moningerae )各不少于5个个体的幼叶, 经变色硅胶干燥, 然后用改良的CTAB (Cetyl Trimethyl Ammonium Bromide, 十六烷基三甲基溴化铵)法提取叶片总DNA (Doyle & Doyle, 1987 ).选择变异程度大的叶绿体基因片段trn L-F (Wang et al, 2010 )和核基因片段ITS1/2 (White et al, 1990 )进行扩增和序列分析(表2 ). ...
A rapid DNA isolation procedure for small quantities of fresh leaf tissue
1
1987
... 在海南岛全岛范围采集了海南特有种烟叶唇柱苣苔(Chirita heterotricha )、海南蛛毛苣苔(Paraboea hainanensis )、盾叶苣苔(Metapetrocosmea peltata )、黄花马铃苣苔(Oreocharis flavida )、毛花马铃苣苔(O. dasyantha )以及广布种红花芒毛苣苔(Aeschynanthus moningerae )各不少于5个个体的幼叶, 经变色硅胶干燥, 然后用改良的CTAB (Cetyl Trimethyl Ammonium Bromide, 十六烷基三甲基溴化铵)法提取叶片总DNA (Doyle & Doyle, 1987 ).选择变异程度大的叶绿体基因片段trn L-F (Wang et al, 2010 )和核基因片段ITS1/2 (White et al, 1990 )进行扩增和序列分析(表2 ). ...
BEAST: Bayesian evolutionary analysis by sampling trees
1
2007
... 用BEAST v1.7.5 (Drummond & Rambaut, 2007 )建立海南岛苦苣苔科植物的系统发育树.首先, 碱基替代模型为根据MODELTEST v3.6 (Posada & Crandall, 1998 )软件计算出的GTR+G模型; 其次, 用BESAT v1.7软件包中BEAUti生成.xml文件, 对所有21个物种拼接好的片段计算物种分化时间, 以Sanango racemosum (57 Ma) (Perret et al, 2013 )来限制某一分支出现的时间.设置的主要参数为: 核苷酸替代模型为GTR, 选择Gamma+Invariant位点, Gamma型分布及4种变化速率类型, 分子钟模型采用不相关对数松弛分子钟(uncorrelated lognormal relaxed molecular clock model), 树的后验拓扑结构为Yule speciation.独立计算2次, 每次运算8,000万代, 每1,000代取样1次, 舍弃10%的树(8,000棵) (Perret et al, 2013 ).最后, 通过TRACER v1.5 (http: //beast.bio.ed.ac.uk/Tracer/)检验相同分布的收敛性, 确保样本足够大小, 用TreeAnnotator v1.7.4软件建树, 所得的MCC树用FigTree v.1.3.1 (htpp://tree.bio. Ed.ac.uk/software/figtree/)打开, 将具有平均分化时间和95%置信区间的树以.pdf文件形式导出. ...
海南岛特有植物的研究
1
1995
... 从海拔分布区域来看, 苦苣苔科原始类型大多分布在低海拔地区, 之后逐渐向中高海拔迁移, 并在这个过程中由于山顶“天岛”效应及河谷隔离作用而分化出了一些海南岛特有种(图4 ), 这说明在向中高海拔迁移的过程中, 山顶形成的孤岛效应以及山脉河谷的隔离作用也促进了海南岛特有种的形成与维持(邢福武, 1995 ; Ling et al, 2017 ). ...
海南岛特有植物的研究
1
1995
... 从海拔分布区域来看, 苦苣苔科原始类型大多分布在低海拔地区, 之后逐渐向中高海拔迁移, 并在这个过程中由于山顶“天岛”效应及河谷隔离作用而分化出了一些海南岛特有种(图4 ), 这说明在向中高海拔迁移的过程中, 山顶形成的孤岛效应以及山脉河谷的隔离作用也促进了海南岛特有种的形成与维持(邢福武, 1995 ; Ling et al, 2017 ). ...
3
2012
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 海南岛苦苣苔科特有种多数分布广泛, 如烟叶唇柱苣苔、盾叶苣苔、毛花马铃苣苔、锈毛马铃苣苔和黄花马铃苣苔在海南岛主要山峰都有较多的自然分布.这主要是因为海南岛中南部山区连续性较高、生境破碎化程度较小, 而且这些物种适应性较强(Francisco-Ortega et al, 2010 ; 邢福武, 2012 ).如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 ). ...
... ).如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 ). ...
Seed plant endemism on Hainan Island: a framework for conservation action
2
2010
... 海南岛生物多样性的起源及其与周边地区的联系一直存在争议(Francisco-Ortega et al, 2010 ; Zhu, 2016 ).张宏达(1962, 2001)指出, 海南岛、台湾岛、广东、广西、福建南部、云南东南部及中南半岛北部等地区的植物均属华南亚区; 张超常和刘兰芳(1983)认为, 海南岛属于华夏植物区系; 而蒋有绪(1988)认为, 海南岛的植物区系应属印度-马来西亚区系.王荷生(2000)、吴征镒等(2011)以及Zhu (2016)基本赞同蒋有绪的结论, 认为海南岛及邻近地区植物区系应属马来西亚亚区.然而, 传统植物区系地理学研究主要以植物区系的分类群组成及其分布区类型的分析为主, 相对忽视了进化历史的分析(李嵘和孙航, 2017 ). ...
... 海南岛苦苣苔科特有种多数分布广泛, 如烟叶唇柱苣苔、盾叶苣苔、毛花马铃苣苔、锈毛马铃苣苔和黄花马铃苣苔在海南岛主要山峰都有较多的自然分布.这主要是因为海南岛中南部山区连续性较高、生境破碎化程度较小, 而且这些物种适应性较强(Francisco-Ortega et al, 2010 ; 邢福武, 2012 ).如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 ). ...
Floral symmetry: pollinator-mediated stabilizing selection on flower size in bilateral species
1
2009
... 苦苣苔科植物的花部特征非常特化, 是对昆虫传粉适应程度很高的一个类群(Wang et al, 2010 ; 韦毅刚, 2010 ; Martén-Rodríguez et al, 2015 ).两侧对称的唇形花冠可能是苦苣苔科作为唇形目一员的原始状态(Wang et al, 2010 ).两侧对称花的访花昆虫更专一, 传粉精确性更高; 而辐射对称花的访花昆虫较多, 花粉浪费较大, 但也不容易受到专一昆虫丧失带来的灭绝风险(Gong & Huang, 2009 ).一般而言, 海岛的昆虫种类与数量少, 植物传粉方式多依靠多种昆虫进行泛化传粉或自交, 花冠多为辐射对称(Barrett et al, 1996 ; Martén-Rodríguez et al, 2015 ).本研究发现, 海南岛苦苣苔科植物大多仍为两侧对称花, 只有冠萼线柱苣苔、大花十字苣苔(Stauranthera grandiflora )和海南岛特有种黄花马铃苣苔出现了辐射对称花(图4 ). ...
3
2012
... 海南岛及其邻近地区是苦苣苔亚科(Cyrtandroideae)的集中分布区(李振宇和王印政, 2005 ; 韦毅刚, 2010 ).苦苣苔亚科约有1,700种, 物种分化与特有中心有两个: 一个是位于中国西南部的广西、云南、贵州交界地区(苏志尧和张宏达, 1994 ; 李振宇, 1996 ; 韦毅刚等, 2004 ; 丁莉等, 2010 ); 另一个位于东南亚的马来群岛(李振宇, 1996 ; 李振宇和王印政, 2005 ; Möller et al, 2010 ).海南岛位于这两个中心之间, 分布着24种苦苣苔科植物, 其中8个特有种(含1变种)、2个特有属(韦毅刚, 2010 ; 邢福武, 2012 ).苦苣苔亚科在海南岛及其一南一北两个物种多样性中心之间的地理分布格局及物种迁移历史, 可能是认识海南岛植物多样性起源与演化、揭示海南岛在亚洲热带植物物种迁移过程中作用的一个窗口. ...
... 海南岛苦苣苔科特有种多数分布广泛, 如烟叶唇柱苣苔、盾叶苣苔、毛花马铃苣苔、锈毛马铃苣苔和黄花马铃苣苔在海南岛主要山峰都有较多的自然分布.这主要是因为海南岛中南部山区连续性较高、生境破碎化程度较小, 而且这些物种适应性较强(Francisco-Ortega et al, 2010 ; 邢福武, 2012 ).如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 ). ...
... ).如烟叶唇柱苣苔既喜阴也耐旱, 在石灰岩与花岗岩石壁上皆可大量生长, 海拔分布范围也较广; 盾叶苣苔虽然仅生长于阴湿土壤或流水石上, 但这种小生境遍布海南岛各大山区和不同海拔; 毛花马铃苣苔耐旱、耐寒, 分布在高海拔的砂砾岩石壁上或者林下岩石上, 在海拔超过1,000 m的五指山、鹦哥岭、霸王岭等地都有分布(邢福武, 2012 ). ...
1
2013
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
1
2013
... 岛屿是开展生物多样性保护、物种迁移与适应进化和生物地理学研究的天然实验室(MacArthur & Wilson, 1967 ; Losos & Ricklefs, 2010 ; Bramwell & Caujapé-Castells, 2011 ).海南岛位于中国大陆最南端、亚洲热带北缘, 面积达3.4万km2 , 是中国第一大全热带岛屿.海南岛也是全球生物多样性热点地区之一(Myers et al, 2000 ), 维管植物多达4,000多种, 其中特有植物有480多种(杨小波, 2013 ). ...
Biogeographical evidences help revealing the origin of Hainan Island
4
2016
... 海南岛生物多样性的起源及其与周边地区的联系一直存在争议(Francisco-Ortega et al, 2010 ; Zhu, 2016 ).张宏达(1962, 2001)指出, 海南岛、台湾岛、广东、广西、福建南部、云南东南部及中南半岛北部等地区的植物均属华南亚区; 张超常和刘兰芳(1983)认为, 海南岛属于华夏植物区系; 而蒋有绪(1988)认为, 海南岛的植物区系应属印度-马来西亚区系.王荷生(2000)、吴征镒等(2011)以及Zhu (2016)基本赞同蒋有绪的结论, 认为海南岛及邻近地区植物区系应属马来西亚亚区.然而, 传统植物区系地理学研究主要以植物区系的分类群组成及其分布区类型的分析为主, 相对忽视了进化历史的分析(李嵘和孙航, 2017 ). ...
... 在海南岛自然分布的苦苣苔科植物中, 分化时间最早的类群是尖舌苣苔属(图3 ).尖舌苣苔属间断分布于亚洲和中南美洲(李振宇, 1996 ), 约在49 Ma左右起源于南美, 之后扩散至亚洲的热带地区即现今的东南亚群岛一带(Perret et al, 2013 ).海南岛有着大量该科的基部类群如尖舌苣苔属、十字苣苔属、横蒴苣苔属以及线柱苣苔属(图3 ), 而且都与东南亚共享, 表明苦苣苔科植物在很早的时候就已从东南亚迁入了海南岛.海南岛的地质历史也证实了这种可能性, 海南岛所处的陆地在早期的时候(印度板块撞击欧亚板块之前)与中南半岛相连(Zhu, 2016 ). ...
... 海南岛-中国大陆共同分布的类群处于海南岛苦苣苔科系统发育树末端的较新分支, 表明海南岛苦苣苔科植物可能是在后期才与中国大陆有着较强的交流.海南岛两个特有属的形成时间约在12 Ma, 特有种也多集中在15-5 Ma才分化出来(图3 ).在15 Ma左右, 海南岛在印度板块撞击欧亚板块、中南半岛向南挤压的作用下从北部湾脱离出来并已远离周边大陆约100 km (Zhu, 2016 ).这种海岛隔离作用可能导致了海南岛苦苣苔科植物特有种的形成(Ling et al, 2017 ). ...
... 海南岛苦苣苔科植物与邻近的广东省有着最高的Jaccard相似性系数(表1 ), 这可能是因为上新世末的海退和晚更新世末海平面的下降, 导致海南岛与广东省的雷州半岛曾两度相连(Liang, 2013 ; Zhu, 2016 ).这个过程中, 广东地区的苦苣苔科植物如芒毛苣苔属、蛛毛苣苔属和线柱苣苔属等植物可能多次进入海南岛(图3 ), 造成两地苦苣苔科物种组成的相似性很高. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}