Beta diversity describes the variation in species composition among communities within a region and it is determined by two antithetic processes: species turnover (or species replacement), and nestedness (or difference in richness). Beta-diversity partitioning aims to separate these two processes when examining species composition among communities, and to reveal their underlying mechanisms. Since 2010, the partitioning methods were proposed following two dominant frameworks: the BAS method proposed by Andrés Baselga in 2010 (partitioning overall beta diversity into turnover and nestedness components) and the POD method proposed by János Podani and Dénes Schmera in 2011 and José C. Carvalho et al. in 2012 (partitioning overall beta diversity into species replacement and richness difference components). With the continuous debate on the nature of the BAS and POD methods, studies on beta-diversity partitioning have developed rapidly worldwide. We reviewed journal articles in the field of beta-diversity partitioning since 2010. Results showed that the number of publications and citations using the BAS method were greater than those using the POD method (75% vs. 20%). In those publications, most of study sites were located in Europe (45%) and research taxa were dominated by animals (64%). Here, we introduce the history and development of beta-diversity partitioning, potential applications in studying biodiversity distributions across spatial-temporal scales (latitudinal/altitudinal gradients, habitat fragmentation, seasonal and annual dynamics), multiple-faceted diversity (taxonomic, functional and phylogenetic diversity), and comparisons among various biological taxa. We point out the following directions in the field of beta-diversity partitioning in the future: (1) the synthesis and comparative analysis of the methods of beta-diversity partitioning; (2) examining patterns of overall beta diversity and its components by incorporating species abundance; and (3) testing the generality of results yielded from beta-diversity partitioning across large scales.
Fig. 1 Publications for studies in the field of beta-diversity partitioning from 2010 to 2016. (A) Number of annual publications cited BAS and POD methods, and using BAS or POD method to analyze the data; (B) Distribution of research areas
Table 1 Pairwise dissimilarities and their components of BAS method (Baselga, 2010) and POD method (Podani & Schmera, 2011). a is the number of shared species between two communities, b and c are the numbers of exclusive species of either.
Fig. 3 Demonstration of species compositions between two sites. a is the number of shared species, b and c are the species exclusive to each site. The number of compositional differences between two sites is b + c. Richness difference is |b - c|, and species replacement is given by 2 min (b, c).
1.1.3 BAS法和POD法的分解组分对照
表1罗列了BAS法和POD法中使用的成对相异性指数。其中, βjne的前半部分|b - c| / (a + b + c)等于βrich。BAS法和POD法的Jaccard成对相异性指数(βjac和βcc)具有相同的表达式(表1)。自2010年之后, 基于BAS法和POD法的beta多样性分解研究迅速发展。目前, 两种分解方法可由R语言中的betapart包(Baselga & Orme, 2012)和BAT包(Cardoso et al, 2015)分别实现。本文的附录3中, 我们简要演示了基于BAS法和POD法的成对和多点相异性指数的计算过程。
1.1.4 BAS法和POD法的争论
2010年, 在西班牙生态学家Andrés Baselga提出BAS法之后, 学者们对该方法的争论也随即出现。例如, Podani和Schmera (2011)、Carvalho等(2012)认为BAS法中提到的嵌套只是物种丰富度差异的一种特殊情况, 因此他们基于Jaccard相异性指数把beta多样性分解为物种替换和物种丰富度差异两部分(即POD法)。此后, Almeida-Neto等(2012)指出, BAS法不能计算物种丧失或增加的程度, 因为BAS法的嵌套组分并不能准确计算严格意义上的嵌套格局, 因此建议应该用诸如NODF (the Nestedness metric based on Overlap and Decreasing Fill)等指数(Atmar & Patterson, 1993; Almeida-Neto et al, 2008)计算真正的嵌套格局。同时, 由于BAS法分解组分的分母不同, 因而尺度也并不相同, 从而会导致βsim高估物种替换的作用(Carvalho et al, 2013)。除此之外, Almeida-Neto等(2012)认为, 在某些特殊情况下, 即使共有物种数为零, BAS法中的多点相异性指数的嵌套组分仍然能够得到非零结果。
Appendix 2 Number of total publications for each journal published studies of beta-diversity partitioning (list journals with ≥ 4 publications only) from 2010 to 2016
Revealing a conservation challenge through partitioned long-term beta diversity: increasing turnover and decreasing nestedness of boreal lake metacommunities.
Distance decay of similarity, effects of environmental noise and ecological heterogeneity among species in the spatio-temporal dynamics of a dispersal-limited community.
Vertebrate dissimilarity due to turnover and richness differences in a highly beta-diverse region: the role of spatial grain size, dispersal ability and distance.
CodyML (1975) Towards a theory of continental species diversities: bird distributions over Mediterranean habitat gradients. In: Ecology and Evolution of Communities (eds Cody ML, Diamond JM), pp. 214-257. Harvard University Press, Cambridge.
[35]
ColwellRK, CoddingtonJA (1994)
Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London,
Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world.
Water-energy balance, past ecological perturbations and evolutionary constraints shape the latitudinal diversity gradient of soil testate amoebae in south- western South America.
Revealing beta-diversity patterns of breeding bird and lizard communities on inundated land-bridge islands by separating the turnover and nestedness components.
Low plant diversity and floristic homogenization in fast-urbanizing towns in Shandong Peninsular, China: effects of urban greening at regional scale for ecological engineering.
Multiscale partitioning of small mammal β-diversity provides novel insights into the Quaternary faunal history of Qinghai-Tibetan Plateau and Hengduan Mountains.
a) Complimentary analysis of metacommunity nestedness and diversity partitioning highlights the need for a holistic conservation strategy for highland lake fish assemblages.
Rethinking the relationship between nestedness and beta diversity: a comment on Baselga (2010).
2012
A consistent metric for nestedness analysis in ecological systems: reconciling concept and measurement.
1
2008
... 2010年, 在西班牙生态学家Andrés Baselga提出BAS法之后, 学者们对该方法的争论也随即出现.例如, Podani和Schmera (2011)、Carvalho等(2012)认为BAS法中提到的嵌套只是物种丰富度差异的一种特殊情况, 因此他们基于Jaccard相异性指数把beta多样性分解为物种替换和物种丰富度差异两部分(即POD法).此后, Almeida-Neto等(2012)指出, BAS法不能计算物种丧失或增加的程度, 因为BAS法的嵌套组分并不能准确计算严格意义上的嵌套格局, 因此建议应该用诸如NODF (the Nestedness metric based on Overlap and Decreasing Fill)等指数(Atmar & Patterson, 1993; Almeida-Neto et al, 2008)计算真正的嵌套格局.同时, 由于BAS法分解组分的分母不同, 因而尺度也并不相同, 从而会导致βsim高估物种替换的作用(Carvalho et al, 2013).除此之外, Almeida-Neto等(2012)认为, 在某些特殊情况下, 即使共有物种数为零, BAS法中的多点相异性指数的嵌套组分仍然能够得到非零结果. ...
Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist.
2
2011
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Beta-diversity gradients of butterflies along productivity axes.
1
2012
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Revealing a conservation challenge through partitioned long-term beta diversity: increasing turnover and decreasing nestedness of boreal lake metacommunities.
... 运用beta多样性分解的方法研究群落在时间维度上的变化, 将有助于了解生态系统对局部环境乃至全球气候变化的响应规律, 有利于生态学家更为深入地理解群落的构建机制并制定更加完善的生物多样性保护策略(Sheldon et al, 2011; Boieiro et al, 2013; Si et al, 2014; Urban, 2015).在时间维度上, 如果周转组分的增加程度大于嵌套组分的降低程度, 则表明所有群落对区域多样性具有更加相似的贡献, 因此所有群落均可能是潜在的保护目标; 如果嵌套组分的增加程度大于周转组分的减小程度, 则表明部分群落对区域多样性的贡献更大, 从而需要优先考虑这些特定的群落.同理, 如果周转和嵌套组分随时间稳定变化(temporal stability), 那么对于维持区域多样性而言, 无需额外实施保护对策; 但是, 如果周转和嵌套组分的大小随时间交错变化, 则难以基于目前的结果提出生物多样性保护建议(Angeler, 2013; 斯幸峰, 2014; Lamy et al, 2015; Si et al, 2015).Angeler (2013)对瑞典26个湖泊的水生无脊椎动物长达17年的研究发现, 总体beta多样性并未发生显著性变化, 但物种空间周转组分却一直在增加, 而嵌套组分一直在减少.因此, 从生物多样性保护角度来讲, 在时间维度上所有生态系统对局域多样性具有更加相似的贡献而均需要保护.Lewis等(2016)对苏格兰滨海草地34年的研究发现, 相较于周转组分, 气候变化对嵌套组分影响更大.在群落构建机制的研究中, Baselga等(2015)发现法国西南地区农田生境中的鸟类群落在25年间没有明显的时间分布格局, 推测可能是由于较强的随机性过程所导致.Habel等(2016)对德国东南部蝶类和蛾类的近2个世纪的研究发现嵌套组分占据主导地位, 并且随着时间推移群落组成更为相似, 这跟生境特化种的减少和泛化种的增加有关.在对农田生境的研究中, Uchida和Ushimaru (2015)发现抛荒地(abandoned terrace)里的植物和直翅目昆虫在时空维度上均具有较高的周转组分, 而在耕地(intensive terrace)里的植物在时间维度上具有较高的嵌套组分, 表明对耕地减少刈割频度并对抛荒地进行刈割将有助于生物多样性恢复. ...
The measure of order and disorder in the distribution of species in fragmented habitat.
1
1993
... 2010年, 在西班牙生态学家Andrés Baselga提出BAS法之后, 学者们对该方法的争论也随即出现.例如, Podani和Schmera (2011)、Carvalho等(2012)认为BAS法中提到的嵌套只是物种丰富度差异的一种特殊情况, 因此他们基于Jaccard相异性指数把beta多样性分解为物种替换和物种丰富度差异两部分(即POD法).此后, Almeida-Neto等(2012)指出, BAS法不能计算物种丧失或增加的程度, 因为BAS法的嵌套组分并不能准确计算严格意义上的嵌套格局, 因此建议应该用诸如NODF (the Nestedness metric based on Overlap and Decreasing Fill)等指数(Atmar & Patterson, 1993; Almeida-Neto et al, 2008)计算真正的嵌套格局.同时, 由于BAS法分解组分的分母不同, 因而尺度也并不相同, 从而会导致βsim高估物种替换的作用(Carvalho et al, 2013).除此之外, Almeida-Neto等(2012)认为, 在某些特殊情况下, 即使共有物种数为零, BAS法中的多点相异性指数的嵌套组分仍然能够得到非零结果. ...
Partitioning the turnover and nestedness components of beta diversity.
13
2010
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... Pairwise dissimilarities and their components of BAS method (Baselga, 2010) and POD method (Podani & Schmera, 2011). a is the number of shared species between two communities, b and c are the numbers of exclusive species of either. ...
... 由于Simpson成对相异性指数(Simpson pairwise dissimilarity index) (βsim)可以排除物种丰富度的影响(Lennon et al, 2001), 因此当两个群落完全不嵌套时, βsor将等于βsim (Baselga, 2010; Leprieur et al, 2012).对于两个不同的群落, βsor和βsim两者之差(βsne)则可以表示beta多样性的嵌套组分(Baselga, 2010).如表1所示, βsne指数的前半部分[|b - c|] / (2a + b + c)], 与βgl指数[2|b - c| / (2a + max(b, c) + min(b, c)]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
... )则可以表示beta多样性的嵌套组分(Baselga, 2010).如表1所示, βsne指数的前半部分[|b - c|] / (2a + b + c)], 与βgl指数[2|b - c| / (2a + max(b, c) + min(b, c)]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
... )两个组分(Baselga, 2010). ...
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
The relationship between species replacement, dissimilarity derived from nestedness, and nestedness.
9
2012
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
betapart: an R package for the study of beta diversity.
1
2012
... 表1罗列了BAS法和POD法中使用的成对相异性指数.其中, βjne的前半部分|b - c| / (a + b + c)等于βrich.BAS法和POD法的Jaccard成对相异性指数(βjac和βcc)具有相同的表达式(表1).自2010年之后, 基于BAS法和POD法的beta多样性分解研究迅速发展.目前, 两种分解方法可由R语言中的betapart包(Baselga & Orme, 2012)和BAT包(Cardoso et al, 2015)分别实现.本文的附录3中, 我们简要演示了基于BAS法和POD法的成对和多点相异性指数的计算过程. ...
Distance decay of similarity, effects of environmental noise and ecological heterogeneity among species in the spatio-temporal dynamics of a dispersal-limited community.
1
2014
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
Impacts of rain forest fragmentation on butterflies in northern Borneo: species richness, turnover and the value of small fragments.
1
2006
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
The utility of DNA metabarcoding for studying the response of arthropod diversity and composition to land-use change in the tropics.
1
2016
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
Contrasting species and functional beta diversity in montane ant assemblages.
1
2015
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Spatial factors play a major role as determinants of endemic ground beetle beta diversity of Madeira Island Laurisilva.
1
2013
... 运用beta多样性分解的方法研究群落在时间维度上的变化, 将有助于了解生态系统对局部环境乃至全球气候变化的响应规律, 有利于生态学家更为深入地理解群落的构建机制并制定更加完善的生物多样性保护策略(Sheldon et al, 2011; Boieiro et al, 2013; Si et al, 2014; Urban, 2015).在时间维度上, 如果周转组分的增加程度大于嵌套组分的降低程度, 则表明所有群落对区域多样性具有更加相似的贡献, 因此所有群落均可能是潜在的保护目标; 如果嵌套组分的增加程度大于周转组分的减小程度, 则表明部分群落对区域多样性的贡献更大, 从而需要优先考虑这些特定的群落.同理, 如果周转和嵌套组分随时间稳定变化(temporal stability), 那么对于维持区域多样性而言, 无需额外实施保护对策; 但是, 如果周转和嵌套组分的大小随时间交错变化, 则难以基于目前的结果提出生物多样性保护建议(Angeler, 2013; 斯幸峰, 2014; Lamy et al, 2015; Si et al, 2015).Angeler (2013)对瑞典26个湖泊的水生无脊椎动物长达17年的研究发现, 总体beta多样性并未发生显著性变化, 但物种空间周转组分却一直在增加, 而嵌套组分一直在减少.因此, 从生物多样性保护角度来讲, 在时间维度上所有生态系统对局域多样性具有更加相似的贡献而均需要保护.Lewis等(2016)对苏格兰滨海草地34年的研究发现, 相较于周转组分, 气候变化对嵌套组分影响更大.在群落构建机制的研究中, Baselga等(2015)发现法国西南地区农田生境中的鸟类群落在25年间没有明显的时间分布格局, 推测可能是由于较强的随机性过程所导致.Habel等(2016)对德国东南部蝶类和蛾类的近2个世纪的研究发现嵌套组分占据主导地位, 并且随着时间推移群落组成更为相似, 这跟生境特化种的减少和泛化种的增加有关.在对农田生境的研究中, Uchida和Ushimaru (2015)发现抛荒地(abandoned terrace)里的植物和直翅目昆虫在时空维度上均具有较高的周转组分, 而在耕地(intensive terrace)里的植物在时间维度上具有较高的嵌套组分, 表明对耕地减少刈割频度并对抛荒地进行刈割将有助于生物多样性恢复. ...
Mammals on mountainsides: elevational patterns of diversity.
1
2001
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Microbes on mountainsides: contrasting elevational patterns of bacterial and plant diversity. Proceedings of the National Academy of Sciences,
2
2008
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Linking global turnover of species and environments. Proceedings of the National Academy of Sciences,
1
2008
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Vertebrate dissimilarity due to turnover and richness differences in a highly beta-diverse region: the role of spatial grain size, dispersal ability and distance.
1
2013
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Testing the performance of beta diversity measures based on incidence data: the robustness to undersampling.
1
2009
... 在成对相异性指数中, 物种替换表示一个物种被另外一个物种替换, 物种丰富度的差异则表示各个群落中特有物种数差值的绝对值, 这与物种组成是否嵌套无关(Carvalho et al, 2012).因此, 总体beta多样性(βcc)可以分解为物种替换(β-3)和物种丰富度的差异(βrich) (Podani & Schmera, 2011; Carvalho et al, 2012) (表1) (简称POD法).两个群落间物种的组成差异(b + c)与两群落总物种数(a + b + c)的比为总体beta多样性(Jaccard相异性指数, βcc) (图3) (Colwell & Coddington, 1994) (该表达式与BAS法的Jaccard成对相异性指数相同).由于群落间会发生物种替换, 两两群落间替换的物种数[2min(b,c)]在总物种数中所占的比例则为β-3 (Williams, 1996; Cardoso et al, 2009).物种丰富度差异等于一个群落的物种数(b + a)和另外一个群落物种数(c + a)差值的绝对值|(b + a) - (c + a)|, 该值在总物种数中的比例为βrich (Schmera & Podani, 2011; Carvalho et al,2012).因此, POD法的表达式为: βcc = β-3 + βrich, 其中 β-3 = 2min(b, c) / (a + b + c), 以及βrich = |(b + a) - (c + a)| / (a + b + c) = |b - c| / (a + b + c) (表1). ...
BAT—biodiversity assessment tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity.
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Measuring fractions of beta diversity and their relationships to nestedness: a theoretical and empirical comparison of novel approaches.
2
2013
... 2010年, 在西班牙生态学家Andrés Baselga提出BAS法之后, 学者们对该方法的争论也随即出现.例如, Podani和Schmera (2011)、Carvalho等(2012)认为BAS法中提到的嵌套只是物种丰富度差异的一种特殊情况, 因此他们基于Jaccard相异性指数把beta多样性分解为物种替换和物种丰富度差异两部分(即POD法).此后, Almeida-Neto等(2012)指出, BAS法不能计算物种丧失或增加的程度, 因为BAS法的嵌套组分并不能准确计算严格意义上的嵌套格局, 因此建议应该用诸如NODF (the Nestedness metric based on Overlap and Decreasing Fill)等指数(Atmar & Patterson, 1993; Almeida-Neto et al, 2008)计算真正的嵌套格局.同时, 由于BAS法分解组分的分母不同, 因而尺度也并不相同, 从而会导致βsim高估物种替换的作用(Carvalho et al, 2013).除此之外, Almeida-Neto等(2012)认为, 在某些特殊情况下, 即使共有物种数为零, BAS法中的多点相异性指数的嵌套组分仍然能够得到非零结果. ...
Determinants of beta diversity of spiders in coastal dunes along a gradient of mediterraneity.
1
2011
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns.
3
2012
... 在成对相异性指数中, 物种替换表示一个物种被另外一个物种替换, 物种丰富度的差异则表示各个群落中特有物种数差值的绝对值, 这与物种组成是否嵌套无关(Carvalho et al, 2012).因此, 总体beta多样性(βcc)可以分解为物种替换(β-3)和物种丰富度的差异(βrich) (Podani & Schmera, 2011; Carvalho et al, 2012) (表1) (简称POD法).两个群落间物种的组成差异(b + c)与两群落总物种数(a + b + c)的比为总体beta多样性(Jaccard相异性指数, βcc) (图3) (Colwell & Coddington, 1994) (该表达式与BAS法的Jaccard成对相异性指数相同).由于群落间会发生物种替换, 两两群落间替换的物种数[2min(b,c)]在总物种数中所占的比例则为β-3 (Williams, 1996; Cardoso et al, 2009).物种丰富度差异等于一个群落的物种数(b + a)和另外一个群落物种数(c + a)差值的绝对值|(b + a) - (c + a)|, 该值在总物种数中的比例为βrich (Schmera & Podani, 2011; Carvalho et al,2012).因此, POD法的表达式为: βcc = β-3 + βrich, 其中 β-3 = 2min(b, c) / (a + b + c), 以及βrich = |(b + a) - (c + a)| / (a + b + c) = |b - c| / (a + b + c) (表1). ...
... ; Carvalho et al, 2012) (表1) (简称POD法).两个群落间物种的组成差异(b + c)与两群落总物种数(a + b + c)的比为总体beta多样性(Jaccard相异性指数, βcc) (图3) (Colwell & Coddington, 1994) (该表达式与BAS法的Jaccard成对相异性指数相同).由于群落间会发生物种替换, 两两群落间替换的物种数[2min(b,c)]在总物种数中所占的比例则为β-3 (Williams, 1996; Cardoso et al, 2009).物种丰富度差异等于一个群落的物种数(b + a)和另外一个群落物种数(c + a)差值的绝对值|(b + a) - (c + a)|, 该值在总物种数中的比例为βrich (Schmera & Podani, 2011; Carvalho et al,2012).因此, POD法的表达式为: βcc = β-3 + βrich, 其中 β-3 = 2min(b, c) / (a + b + c), 以及βrich = |(b + a) - (c + a)| / (a + b + c) = |b - c| / (a + b + c) (表1). ...
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
The merging of community ecology and phylogenetic biology.
1
2009
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
1975
Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London,
1
1994
... 在成对相异性指数中, 物种替换表示一个物种被另外一个物种替换, 物种丰富度的差异则表示各个群落中特有物种数差值的绝对值, 这与物种组成是否嵌套无关(Carvalho et al, 2012).因此, 总体beta多样性(βcc)可以分解为物种替换(β-3)和物种丰富度的差异(βrich) (Podani & Schmera, 2011; Carvalho et al, 2012) (表1) (简称POD法).两个群落间物种的组成差异(b + c)与两群落总物种数(a + b + c)的比为总体beta多样性(Jaccard相异性指数, βcc) (图3) (Colwell & Coddington, 1994) (该表达式与BAS法的Jaccard成对相异性指数相同).由于群落间会发生物种替换, 两两群落间替换的物种数[2min(b,c)]在总物种数中所占的比例则为β-3 (Williams, 1996; Cardoso et al, 2009).物种丰富度差异等于一个群落的物种数(b + a)和另外一个群落物种数(c + a)差值的绝对值|(b + a) - (c + a)|, 该值在总物种数中的比例为βrich (Schmera & Podani, 2011; Carvalho et al,2012).因此, POD法的表达式为: βcc = β-3 + βrich, 其中 β-3 = 2min(b, c) / (a + b + c), 以及βrich = |(b + a) - (c + a)| / (a + b + c) = |b - c| / (a + b + c) (表1). ...
Midpoint attractors and species richness: modelling the interaction between environmental drivers and geometric constraints.
1
2016
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
The mid-domain effect: geometric constraints on the geography of species richness.
1
2000
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world.
1
2010
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Ecological responses to habitat fragmentation per se. Annual Review of Ecology, Evolution,
1
2017
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Conservation evaluation and phylogenetic diversity.
1
1992
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Water-energy balance, past ecological perturbations and evolutionary constraints shape the latitudinal diversity gradient of soil testate amoebae in south- western South America.
1
2016
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
The effect of elevation and time on mountain spider diversity: a view of two aspects in the Cederberg mountains of South Africa.
Species divergence and trait convergence in experimental plant community assembly.
1
2005
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Near-complete extinction of native small mammal fauna 25 years after forest fragmentation.
1
2013
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Phylogenetic beta diversity: linking ecological and evolutionary processes across space in time.
1
2008
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Spatial and temporal patterns of waterbird assemblages in fragmented wetlands of southern Brazil.
1
2005
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Contrasting effects of natural and anthropogenic stressors on beta diversity in river organisms.
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Butterfly community shifts over 2 centuries.
2016
Neutral theory as a predictor of avifaunal extinctions after habitat loss. Proceedings of the National Academy of Sciences,
1
2011
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Beta diversity on geographic gradients in Britain.
1
1992
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
The elevational gradient in Andean bird species richness at the local scale: a foothill peak and a high-elevation plateau.
1
2005
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
On the generality of the latitudinal diversity gradient.
1
2004
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Ice age climate, evolutionary constraints and diversity patterns of European dung beetles.
1
2011
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Disentangling nestedness from models of ecological complexity.
1
2012
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
Beta diversity along environmental gradients: implications of habitat specialization in tropical montane landscapes.
1
2009
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
The global diversity of birds in space and time.
1
2012
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Spatial components of bird diversity in the Andes of Colombia: implications for designing a regional reserve system.
1
2006
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
a) Measuring beta diversity for presence-absence data.
1
2003
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
b) Are there latitudinal gradients in species turnover?
2
2003
... 由于Simpson成对相异性指数(Simpson pairwise dissimilarity index) (βsim)可以排除物种丰富度的影响(Lennon et al, 2001), 因此当两个群落完全不嵌套时, βsor将等于βsim (Baselga, 2010; Leprieur et al, 2012).对于两个不同的群落, βsor和βsim两者之差(βsne)则可以表示beta多样性的嵌套组分(Baselga, 2010).如表1所示, βsne指数的前半部分[|b - c|] / (2a + b + c)], 与βgl指数[2|b - c| / (2a + max(b, c) + min(b, c)]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Disentangling the drivers of β diversity along latitudinal and elevational gradients.
1
2011
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels.
1
2010
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Understanding the spatio-temporal response of coral reef fish communities to natural disturbances: insights from beta-diversity decomposition.
1
2015
... 运用beta多样性分解的方法研究群落在时间维度上的变化, 将有助于了解生态系统对局部环境乃至全球气候变化的响应规律, 有利于生态学家更为深入地理解群落的构建机制并制定更加完善的生物多样性保护策略(Sheldon et al, 2011; Boieiro et al, 2013; Si et al, 2014; Urban, 2015).在时间维度上, 如果周转组分的增加程度大于嵌套组分的降低程度, 则表明所有群落对区域多样性具有更加相似的贡献, 因此所有群落均可能是潜在的保护目标; 如果嵌套组分的增加程度大于周转组分的减小程度, 则表明部分群落对区域多样性的贡献更大, 从而需要优先考虑这些特定的群落.同理, 如果周转和嵌套组分随时间稳定变化(temporal stability), 那么对于维持区域多样性而言, 无需额外实施保护对策; 但是, 如果周转和嵌套组分的大小随时间交错变化, 则难以基于目前的结果提出生物多样性保护建议(Angeler, 2013; 斯幸峰, 2014; Lamy et al, 2015; Si et al, 2015).Angeler (2013)对瑞典26个湖泊的水生无脊椎动物长达17年的研究发现, 总体beta多样性并未发生显著性变化, 但物种空间周转组分却一直在增加, 而嵌套组分一直在减少.因此, 从生物多样性保护角度来讲, 在时间维度上所有生态系统对局域多样性具有更加相似的贡献而均需要保护.Lewis等(2016)对苏格兰滨海草地34年的研究发现, 相较于周转组分, 气候变化对嵌套组分影响更大.在群落构建机制的研究中, Baselga等(2015)发现法国西南地区农田生境中的鸟类群落在25年间没有明显的时间分布格局, 推测可能是由于较强的随机性过程所导致.Habel等(2016)对德国东南部蝶类和蛾类的近2个世纪的研究发现嵌套组分占据主导地位, 并且随着时间推移群落组成更为相似, 这跟生境特化种的减少和泛化种的增加有关.在对农田生境的研究中, Uchida和Ushimaru (2015)发现抛荒地(abandoned terrace)里的植物和直翅目昆虫在时空维度上均具有较高的周转组分, 而在耕地(intensive terrace)里的植物在时间维度上具有较高的嵌套组分, 表明对耕地减少刈割频度并对抛荒地进行刈割将有助于生物多样性恢复. ...
Interpreting the replacement and richness difference components of beta diversity.
8
2014
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... Legendre (2014)基于POD法, 提出了将Sørensen成对相异性指数分解为物种替换(Repls)和物种丰富度差异(RichDiffs)的分解方法, 其中Repls = 2min(b, c) / (2a + b + c); RichDiffs = |b - c| / (2a + b + c).通过比较可以发现, Repls和RichDiffs与POD法分解得出的Jaccard组分分母发生了变化, 而分子等于β-3和βrich的分子部分. ...
The geographical structure of British bird distributions: diversity, spatial turnover and scale.
3
2001
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... 由于Simpson成对相异性指数(Simpson pairwise dissimilarity index) (βsim)可以排除物种丰富度的影响(Lennon et al, 2001), 因此当两个群落完全不嵌套时, βsor将等于βsim (Baselga, 2010; Leprieur et al, 2012).对于两个不同的群落, βsor和βsim两者之差(βsne)则可以表示beta多样性的嵌套组分(Baselga, 2010).如表1所示, βsne指数的前半部分[|b - c|] / (2a + b + c)], 与βgl指数[2|b - c| / (2a + max(b, c) + min(b, c)]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
... )]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
Quantifying phylogenetic beta diversity: distinguishing between ‘true’ turnover of lineages and phylogenetic diversity gradients.
1
2012
... 由于Simpson成对相异性指数(Simpson pairwise dissimilarity index) (βsim)可以排除物种丰富度的影响(Lennon et al, 2001), 因此当两个群落完全不嵌套时, βsor将等于βsim (Baselga, 2010; Leprieur et al, 2012).对于两个不同的群落, βsor和βsim两者之差(βsne)则可以表示beta多样性的嵌套组分(Baselga, 2010).如表1所示, βsne指数的前半部分[|b - c|] / (2a + b + c)], 与βgl指数[2|b - c| / (2a + max(b, c) + min(b, c)]相近(Lennon et al, 2001; Koleff et al, 2003a), 均是计算物种丰富度的差异(difference in richness); βsne指数的后半部分[a / (a + min(b, c))]是Simpson相似性指数(1 - βsim), 因此βsne能够计算物种丰富度差异中因嵌套而产生的部分.因此, 所有形式的总体beta多样性(βsor)均可以分解成物种周转(βsim)和嵌套(βsne)两个组分(Baselga, 2010). ...
The need for richness-independent measures of turnover when delineating biogeographical regions.
2014
Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes.
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
... 目前在有关beta多样性分解的研究中, 与区域尺度相比, 在更大时空维度下对beta多样性分解的结果进行普适性验证的研究相对较少(Leprieur et al, 2011; Dobrovolski et al, 2012).此外, 应加强在同一区域内对多个生物类群进行综合分析, 并探讨不同生物类群间的总体beta多样性及其组分的贡献程度是否一致.对于岛屿系统中物种beta多样性的周转和嵌套组分, Wu等(2017)整合分析了全球岛屿上陆生动物的群落结构(共18个研究案例), 结果表明94%的研究中周转组分大于嵌套组分, 这说明至少在岛屿系统中总体beta多样性主要取决于物种的周转过程.此外, 从不同时空维度或者多样性的不同方面整合分析周转和嵌套组分在总体beta多样性中所占比例的研究还鲜有报道.在同一地点对多个生物类群进行综合分析, 虽然国际上有不少研究平台具有这样的研究条件(Si et al, 2015; Thorn et al, 2016), 但至今未有类似的研究报道. ...
Climate drives temporal replacement and nested-resultant richness patterns of Scottish coastal vegetation.
2016
Convergence and divergence in a long-term old-field succession: the importance of spatial scale and species abundance.
1
2016
... 相较于物种频度分布数据(presence/absence distributions), 基于物种多度数据(abundance)的研究同样具有重要的生态学意义, 并有利于探讨beta多样性及其组分的时空格局(Legendre, 2014; Li et al, 2016).由于优势种在群落中占据着核心的生态位, 因此优势种可能更容易受到生境过滤等确定性过程的影响(Umaña et al, 2015), 而稀有种由于其种群较小, 更易受到随机过程的影响(Orrock & Watling, 2010).目前, 基于多度数据的beta多样性分解方法已经提出(Baselga, 2013b; Podani et al, 2013; Baselga, 2017), 在今后的研究中可以通过整合物种分布和多度的信息来探讨beta多样性及其组分的时空分布格局和形成机制. ...
Mountain metacommunities: climate and spatial connectivity shape ant diversity in a complex landscape.
1
2017
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Contribution of environmental filtering and dispersal limitation to species turnover of temperate deciduous broad-leaved forests in China.
1
2015
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Performance of partitioning functional beta-diversity indices: influence of functional representation and partitioning methods.
1
2017
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Elevation gradients of species-density: historical and prospective views.
1
2001
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Phylogenetic perspectives on community ecology.
1
1996
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Beta-diversity patterns elucidate mechanisms of alien plant invasion in mountains.
1
2013
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Microbial biogeography: putting microorganisms on the map.
1
2006
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Global analysis of bird elevational diversity.
1
2009
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Species turnover in vertebrate communities along elevational gradients is idiosyncratic and unrelated to species richness.
1
2016
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Rebuilding community ecology from functional traits.
1
2006
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Putting beta-diversity on the map: broad-scale congruence and coincidence in the extremes.
1
2007
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
Environmental drivers of beta-diversity patterns in New-World birds and mammals.
1
2009
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Functional redundancy of multiple forest taxa along an elevational gradient: predicting the consequences of non-random species loss.
1
2015
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Community assembly processes shape an altitudinal gradient of forest biodiversity.
1
2013
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
The challenge of delineating biogeographical regions: nestedness matters for Indo-Pacific coral reef fishes.
1
2013
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
The distance decay of similarity in biogeography and ecology.
2
1999
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Local community size mediates ecological drift and competition in metacommunities.
1
2010
... 相较于物种频度分布数据(presence/absence distributions), 基于物种多度数据(abundance)的研究同样具有重要的生态学意义, 并有利于探讨beta多样性及其组分的时空格局(Legendre, 2014; Li et al, 2016).由于优势种在群落中占据着核心的生态位, 因此优势种可能更容易受到生境过滤等确定性过程的影响(Umaña et al, 2015), 而稀有种由于其种群较小, 更易受到随机过程的影响(Orrock & Watling, 2010).目前, 基于多度数据的beta多样性分解方法已经提出(Baselga, 2013b; Podani et al, 2013; Baselga, 2017), 在今后的研究中可以通过整合物种分布和多度的信息来探讨beta多样性及其组分的时空分布格局和形成机制. ...
Nested subsets and the structure of insular mammalian faunas and archipelagos.
2
1986
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
Functional diversity (FD), species richness and community composition.
1
2002
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Functional diversity: back to basics and looking forward.
1
2006
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Latitudinal gradients in species diversity: a review of concepts.
1
1966
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Times to extinction for small populations of large birds. Proceedings of the National Academy of Sciences,
1
1993
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
The biodiversity of species and their rates of extinction, distribution, and protection.
1
2014
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
A general framework for analyzing beta diversity, nestedness and related community-level phenomena based on abundance data.
1
2013
... 相较于物种频度分布数据(presence/absence distributions), 基于物种多度数据(abundance)的研究同样具有重要的生态学意义, 并有利于探讨beta多样性及其组分的时空格局(Legendre, 2014; Li et al, 2016).由于优势种在群落中占据着核心的生态位, 因此优势种可能更容易受到生境过滤等确定性过程的影响(Umaña et al, 2015), 而稀有种由于其种群较小, 更易受到随机过程的影响(Orrock & Watling, 2010).目前, 基于多度数据的beta多样性分解方法已经提出(Baselga, 2013b; Podani et al, 2013; Baselga, 2017), 在今后的研究中可以通过整合物种分布和多度的信息来探讨beta多样性及其组分的时空分布格局和形成机制. ...
A new conceptual and methodological framework for exploring and explaining pattern in presence-absence data.
7
2011
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... Pairwise dissimilarities and their components of BAS method (Baselga, 2010) and POD method (Podani & Schmera, 2011). a is the number of shared species between two communities, b and c are the numbers of exclusive species of either. ...
... 在成对相异性指数中, 物种替换表示一个物种被另外一个物种替换, 物种丰富度的差异则表示各个群落中特有物种数差值的绝对值, 这与物种组成是否嵌套无关(Carvalho et al, 2012).因此, 总体beta多样性(βcc)可以分解为物种替换(β-3)和物种丰富度的差异(βrich) (Podani & Schmera, 2011; Carvalho et al, 2012) (表1) (简称POD法).两个群落间物种的组成差异(b + c)与两群落总物种数(a + b + c)的比为总体beta多样性(Jaccard相异性指数, βcc) (图3) (Colwell & Coddington, 1994) (该表达式与BAS法的Jaccard成对相异性指数相同).由于群落间会发生物种替换, 两两群落间替换的物种数[2min(b,c)]在总物种数中所占的比例则为β-3 (Williams, 1996; Cardoso et al, 2009).物种丰富度差异等于一个群落的物种数(b + a)和另外一个群落物种数(c + a)差值的绝对值|(b + a) - (c + a)|, 该值在总物种数中的比例为βrich (Schmera & Podani, 2011; Carvalho et al,2012).因此, POD法的表达式为: βcc = β-3 + βrich, 其中 β-3 = 2min(b, c) / (a + b + c), 以及βrich = |(b + a) - (c + a)| / (a + b + c) = |b - c| / (a + b + c) (表1). ...
Once again on the components of pairwise beta diversity.
2016
On-farm habitat restoration counters biotic homogenization in intensively managed agriculture.
1
2016
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
Beta diversity in relation to dispersal ability for vascular plants in North America.
1
2009
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Drivers of β-diversity along latitudinal gradients revisited.
3
2013
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... ; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
A latitudinal gradient in large-scale beta diversity for vascular plants in North America.
3
2007
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
The elevational gradient of species richness: a uniform pattern?
1
1995
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
The role of spatial scale and the perception of large-scale species-richness patterns.
1
2005
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Community diversity: relative roles of local and regional processes.
1
1987
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
Computing β-diversity from species-area curves.
1
2002
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
A multiple-site dissimilarity measure for species presence/absence data and its relationship with nestedness and turnover.
Revealing beta-diversity patterns of breeding bird and lizard communities on inundated land-bridge islands by separating the turnover and nestedness components.
4
2015
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
... 运用beta多样性分解的方法研究群落在时间维度上的变化, 将有助于了解生态系统对局部环境乃至全球气候变化的响应规律, 有利于生态学家更为深入地理解群落的构建机制并制定更加完善的生物多样性保护策略(Sheldon et al, 2011; Boieiro et al, 2013; Si et al, 2014; Urban, 2015).在时间维度上, 如果周转组分的增加程度大于嵌套组分的降低程度, 则表明所有群落对区域多样性具有更加相似的贡献, 因此所有群落均可能是潜在的保护目标; 如果嵌套组分的增加程度大于周转组分的减小程度, 则表明部分群落对区域多样性的贡献更大, 从而需要优先考虑这些特定的群落.同理, 如果周转和嵌套组分随时间稳定变化(temporal stability), 那么对于维持区域多样性而言, 无需额外实施保护对策; 但是, 如果周转和嵌套组分的大小随时间交错变化, 则难以基于目前的结果提出生物多样性保护建议(Angeler, 2013; 斯幸峰, 2014; Lamy et al, 2015; Si et al, 2015).Angeler (2013)对瑞典26个湖泊的水生无脊椎动物长达17年的研究发现, 总体beta多样性并未发生显著性变化, 但物种空间周转组分却一直在增加, 而嵌套组分一直在减少.因此, 从生物多样性保护角度来讲, 在时间维度上所有生态系统对局域多样性具有更加相似的贡献而均需要保护.Lewis等(2016)对苏格兰滨海草地34年的研究发现, 相较于周转组分, 气候变化对嵌套组分影响更大.在群落构建机制的研究中, Baselga等(2015)发现法国西南地区农田生境中的鸟类群落在25年间没有明显的时间分布格局, 推测可能是由于较强的随机性过程所导致.Habel等(2016)对德国东南部蝶类和蛾类的近2个世纪的研究发现嵌套组分占据主导地位, 并且随着时间推移群落组成更为相似, 这跟生境特化种的减少和泛化种的增加有关.在对农田生境的研究中, Uchida和Ushimaru (2015)发现抛荒地(abandoned terrace)里的植物和直翅目昆虫在时空维度上均具有较高的周转组分, 而在耕地(intensive terrace)里的植物在时间维度上具有较高的嵌套组分, 表明对耕地减少刈割频度并对抛荒地进行刈割将有助于生物多样性恢复. ...
... 目前在有关beta多样性分解的研究中, 与区域尺度相比, 在更大时空维度下对beta多样性分解的结果进行普适性验证的研究相对较少(Leprieur et al, 2011; Dobrovolski et al, 2012).此外, 应加强在同一区域内对多个生物类群进行综合分析, 并探讨不同生物类群间的总体beta多样性及其组分的贡献程度是否一致.对于岛屿系统中物种beta多样性的周转和嵌套组分, Wu等(2017)整合分析了全球岛屿上陆生动物的群落结构(共18个研究案例), 结果表明94%的研究中周转组分大于嵌套组分, 这说明至少在岛屿系统中总体beta多样性主要取决于物种的周转过程.此外, 从不同时空维度或者多样性的不同方面整合分析周转和嵌套组分在总体beta多样性中所占比例的研究还鲜有报道.在同一地点对多个生物类群进行综合分析, 虽然国际上有不少研究平台具有这样的研究条件(Si et al, 2015; Thorn et al, 2016), 但至今未有类似的研究报道. ...
Selective extinction drives taxonomic and functional alpha and beta diversities in island bird assemblages.
4
2016
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Functional and phylogenetic structure of island bird communities.
1
2017
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
欧美陆地鸟类监测的历史、现状与我国的对策. 生物多样性,
1
2011
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
欧美陆地鸟类监测的历史、现状与我国的对策. 生物多样性,
1
2011
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Turnover of breeding bird communities on islands in an inundated lake.
2014
Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees.
1
2013
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
How should beta-diversity inform biodiversity conservation?
3
2016
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
Deforestation and avian extinction on tropical landbridge islands.
1
2010
... 由于人类活动的加剧, 生境丧失与片断化对生物多样性的影响成为保护生物学的重要研究内容(Wilcove et al, 1998; Sodhi et al, 2010; Pimm et al, 2014; Wilson et al, 2016).生境的丧失将导致较小斑块中的动物群落具有更高的灭绝风险, 因此普遍认为较小的斑块由于具有较低的保护优先等级而受到较少的关注(Pimm et al, 1993; Krauss et al, 2010; Halley & Iwasa, 2011; Gibson et al, 2013).但是, 因生境片断化形成的单个小斑块虽然难以承载一个种群, 而多个邻近的小斑块却因为分散了灭绝风险或通过“景观补偿效应” (landscape complementation)对生物多样性的维持有所贡献(Guadagnin et al, 2005; Fahrig, 2017), 因此小斑块可能依旧拥有潜在的保护价值而不能忽略(Benedick et al, 2006; Kattan et al, 2006; Si et al, 2015).在空间维度上, beta多样性的物种周转和嵌套组分均能反映不同的保护学策略, 即如果嵌套组分占优势, 那么该研究区域内具有较多物种的地点拥有较高的保护等级; 而如果空间周转组分占主导则意味着所有研究地点对beta多样性的贡献相似, 那么所有地点都需要保护, 没有优先级之分(Baselga, 2010; Gutiérrez-Cánovas et al, 2013).Si等(2015)对千岛湖片断化生境岛屿上的繁殖鸟类和蜥蜴群落的研究发现, 不管是鸟类还是蜥蜴, 总体beta多样性均是由物种周转过程主导.Wu等(2017)对千岛湖岛屿上的蜘蛛群落研究同样发现周转过程在总体beta多样性中占主导地位.这表明在开展千岛湖生物多样性保护时, 除了考虑较大岛屿或者生境较为丰富的岛屿之外, 还应该包括其他小型岛屿或生境较为单一的岛屿.因此在片断化生境中开展物种保护以及保护区设计时, 除了考虑不同斑块内的物种丰富度, 还需要分析不同群落间的beta多样性分布格局及其背后的驱动过程, 这将有利于制定科学有效且具有针对性的生物多样性保护策略(Wiersma & Urban, 2005; 斯幸峰, 2014; Socolar et al, 2016). ...
The distance decay of similarity in ecological communities.
1
2007
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
The ghost of nestedness in ecological networks.
1
2013
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
Inferring ecological processes from taxonomic, phylogenetic and functional trait beta-diversity.
1
2011
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Environment, dispersal and patterns of species similarity.
1
2006
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Human impacts on functional and taxonomic homogenization of plateau fish assemblages in Yunnan, China.
1
2015
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
Climate, history and neutrality as drivers of mammal beta diversity in Europe: insights from multiscale deconstruction.
1
2011
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
a) Deterministic tropical tree community turnover: evidence from patterns of functional beta diversity along an elevational gradient.
1
2011
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
b) Temporal turnover in the composition of tropical tree communities: functional determinism and phylogenetic stochasticity.
2
2011
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Patterns of plant beta-diversity along elevational and latitudinal gradients in mountain forests of China.
1
2012
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Toward a trophic theory of species diversity. Proceedings of the National Academy of Sciences,
1
2015
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Changes in the dominant assembly mechanism drive species loss caused by declining resources.
1
2016
... 目前在有关beta多样性分解的研究中, 与区域尺度相比, 在更大时空维度下对beta多样性分解的结果进行普适性验证的研究相对较少(Leprieur et al, 2011; Dobrovolski et al, 2012).此外, 应加强在同一区域内对多个生物类群进行综合分析, 并探讨不同生物类群间的总体beta多样性及其组分的贡献程度是否一致.对于岛屿系统中物种beta多样性的周转和嵌套组分, Wu等(2017)整合分析了全球岛屿上陆生动物的群落结构(共18个研究案例), 结果表明94%的研究中周转组分大于嵌套组分, 这说明至少在岛屿系统中总体beta多样性主要取决于物种的周转过程.此外, 从不同时空维度或者多样性的不同方面整合分析周转和嵌套组分在总体beta多样性中所占比例的研究还鲜有报道.在同一地点对多个生物类群进行综合分析, 虽然国际上有不少研究平台具有这样的研究条件(Si et al, 2015; Thorn et al, 2016), 但至今未有类似的研究报道. ...
1
2001
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
a) A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity.
2
2010
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
b) A diversity of beta diversities: straightening up a concept gone awry. Part 2. Quantifying beta diversity and related phenomena.
2
2010
... 随着beta多样性研究的逐渐深入, 加性分配法(β = γ - α)和相似性(或相异性)指数等多种beta多样性测定方法被提出(Tuomisto, 2010a, b; Anderson et al, 2011).在这些方法中, 相似性或相异性指数最为常用, 并被广泛用来测定群落物种组成在时间或空间维度上的变化.这些指数中Jaccard和Sørensen指数应用最广, 其次为Simpson和Bray-Curtis指数(陈圣宾等, 2010).此外, beta多样性亦被定义为 种-面积曲线的斜率(z) (Rosenzweig, 1995; Ricotta et al, 2002; Qian et al, 2013)、距离衰减效应(distance- decay effect) (Nekola & White, 1999; Soininen et al, 2007; Bellier et al, 2014)、群落总方差(total variance of community data) (Legendre & De Cáceres, 2013)和区域与局域物种丰富度间的差异(Ricklefs, 1987; Qian et al, 2013)等各种形式.Beta多样性测量方法的发展进一步扩展了其在生态学和保护生物学中的应用, 诸如被用来进行生物地理区域划分(Williams, 1996; Baselga et al, 2012; Mouillot et al, 2013), 以及保护区选址和保护区网络设置等(Wiersma & Urban, 2005; McKnight et al, 2007; Socolar et al, 2016). ...
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Land abandonment and intensification diminish spatial and temporal β-diversity of grassland plants and herbivorous insects within paddy terraces.
2015
A consumer’s guide to nestedness analysis.
1
2009
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
Commonness, rarity, and intraspecific variation in traits and performance in tropical tree seedlings.
1
2015
... 相较于物种频度分布数据(presence/absence distributions), 基于物种多度数据(abundance)的研究同样具有重要的生态学意义, 并有利于探讨beta多样性及其组分的时空格局(Legendre, 2014; Li et al, 2016).由于优势种在群落中占据着核心的生态位, 因此优势种可能更容易受到生境过滤等确定性过程的影响(Umaña et al, 2015), 而稀有种由于其种群较小, 更易受到随机过程的影响(Orrock & Watling, 2010).目前, 基于多度数据的beta多样性分解方法已经提出(Baselga, 2013b; Podani et al, 2013; Baselga, 2017), 在今后的研究中可以通过整合物种分布和多度的信息来探讨beta多样性及其组分的时空分布格局和形成机制. ...
Accelerating extinction risk from climate change.
1
2015
... 运用beta多样性分解的方法研究群落在时间维度上的变化, 将有助于了解生态系统对局部环境乃至全球气候变化的响应规律, 有利于生态学家更为深入地理解群落的构建机制并制定更加完善的生物多样性保护策略(Sheldon et al, 2011; Boieiro et al, 2013; Si et al, 2014; Urban, 2015).在时间维度上, 如果周转组分的增加程度大于嵌套组分的降低程度, 则表明所有群落对区域多样性具有更加相似的贡献, 因此所有群落均可能是潜在的保护目标; 如果嵌套组分的增加程度大于周转组分的减小程度, 则表明部分群落对区域多样性的贡献更大, 从而需要优先考虑这些特定的群落.同理, 如果周转和嵌套组分随时间稳定变化(temporal stability), 那么对于维持区域多样性而言, 无需额外实施保护对策; 但是, 如果周转和嵌套组分的大小随时间交错变化, 则难以基于目前的结果提出生物多样性保护建议(Angeler, 2013; 斯幸峰, 2014; Lamy et al, 2015; Si et al, 2015).Angeler (2013)对瑞典26个湖泊的水生无脊椎动物长达17年的研究发现, 总体beta多样性并未发生显著性变化, 但物种空间周转组分却一直在增加, 而嵌套组分一直在减少.因此, 从生物多样性保护角度来讲, 在时间维度上所有生态系统对局域多样性具有更加相似的贡献而均需要保护.Lewis等(2016)对苏格兰滨海草地34年的研究发现, 相较于周转组分, 气候变化对嵌套组分影响更大.在群落构建机制的研究中, Baselga等(2015)发现法国西南地区农田生境中的鸟类群落在25年间没有明显的时间分布格局, 推测可能是由于较强的随机性过程所导致.Habel等(2016)对德国东南部蝶类和蛾类的近2个世纪的研究发现嵌套组分占据主导地位, 并且随着时间推移群落组成更为相似, 这跟生境特化种的减少和泛化种的增加有关.在对农田生境的研究中, Uchida和Ushimaru (2015)发现抛荒地(abandoned terrace)里的植物和直翅目昆虫在时空维度上均具有较高的周转组分, 而在耕地(intensive terrace)里的植物在时间维度上具有较高的嵌套组分, 表明对耕地减少刈割频度并对抛荒地进行刈割将有助于生物多样性恢复. ...
Decomposing functional β-diversity reveals that low functional β-diversity is driven by low functional turnover in European fish assemblages.
1
2013
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
New multidimensional functional diversity indices for a multifaceted framework in functional ecology.
1
2008
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
Low functional β-diversity despite high taxonomic β-diversity among tropical estuarine fish communities.
2
2012
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Low plant diversity and floristic homogenization in fast-urbanizing towns in Shandong Peninsular, China: effects of urban greening at regional scale for ecological engineering.
1
2014
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
Regional and global elevational patterns of microbial species richness and evenness.
1
2017
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
Nutrient enrichment modifies temperature-biodiversity relationships in large-scale field experiments.
1
2016
... 不同的生物类群具有不同的进化历史和生物学特征, 因此形成不同的beta多样性分布格局.在相同的环境梯度或者地理区域中对比研究不同生物类群, 可以将不同类群群落间的组成变化与类群特征联系起来(Bryant et al, 2008; Dobrovolski et al, 2012; Calderón-Patrón et al, 2013).其中, 物种扩散能力是影响物种分布的重要因素(Nekola & White, 1999), 因此具有较强扩散能力的陆栖脊椎动物能够较为容易地越过地理屏障, 从而形成较低的beta多样性(Steinitz et al, 2006; Qian, 2009)和较高的周转组分(Baselga et al, 2012).例如, 相比于扩散能力相对较弱的兽类、两栖类和爬行类等陆生脊椎动物类群, 鸟类具有较强的扩散能力(飞行能力)而呈现出相对较低的beta多样性(Harrison et al, 1992).Si等(2015)在千岛湖的研究中同样发现, 相较于鸟类, 蜥蜴由于其较弱的扩散能力, 更容易受到不同岛屿间水域隔离作用的影响而具有较高的总体beta多样性以及相对较高的嵌套组分.在微生物的beta多样性研究中发现, 由于微生物具有较强的扩散作用, 容易形成较低的beta多样性, 但同时由于微生物具有较高的环境选择性, 也可能因此形成较高的beta多样性(Martiny et al, 2006; Wang et al, 2016).此外, 长期的进化历史过程也会影响物种在大尺度上的分布格局(Qian & Ricklefs, 2007; Jetz et al, 2012).例如, 冰川效应影响了欧洲生物的地理分布(Svenning et al, 2011).由于变温动物对低温更为敏感(Buckley & Jetz, 2008), 因此相较于恒温的鸟类和兽类, 两栖类动物更有可能由于冰川效应的影响而快速消失.同时, 由于其较低的扩散能力, 在冰期之后, 两栖类的物种迁入率(拓殖率)也较低(Baselga, 2010).Dobrovolski等(2012)研究新大陆(New World)的冰川效应后也发现了类似的格局: 鸟类具有最大的空间周转组分, 而两栖类的空间周转组分最低.除了生物类群间扩散能力的差异之外, 不同动物类群间的食性差异、活动节律, 植物类群中的物候差异, 以及它们对气候变化的响应都可能影响物种的分布格局, 从而对组成总体beta多样性的周转和嵌套组分产生影响.随着beta多样性分解研究的迅速发展, 以及全球范围内物种分布数据的积累, 例如通过公民科学(citizen science)的方式收集众多鸟类分布的数据(斯幸峰和丁平, 2011), 将有助于探讨不同生物类群间群落组成的时空变化. ...
Patterns of elevational beta diversity in micro- and macroorganisms.
1
2012
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
黄土高原植物群落演替过程中的β多样性变化. 生态学杂志,
1
2013
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
黄土高原植物群落演替过程中的β多样性变化. 生态学杂志,
1
2013
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
黄土高原油松林林下群落异质性分析. 生态学杂志,
1
2016
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
黄土高原油松林林下群落异质性分析. 生态学杂志,
1
2016
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
Nestedness for different reasons: the distributions of birds, lizards and small mammals on islands of an inundated lake.
2
2010
... 广义的beta多样性是指不同地点间物种组成的差异, 然而不同地点间的物种丰富度差异会导致地点间物种组成的变化, 因而在分析beta多样性的空间格局时应避免物种丰富度差异的干扰(Kraft et al, 2011; Qian et al, 2013).早在20世纪90年代和本世纪初, Harrison等(1992)、Williams (1996)和Lennon等(2001)就开创性地提出群落间物种组成差异起源于两种不同的过程: 物种周转或替换(species turnover, or replacement)和物种丧失或增加(species loss or gain).其中, 物种周转表示不同群落间的物种替换, 而物种丧失会导致群落间物种丰富度产生差异(Lennon et al, 2001).当物种丰富度差异沿着某一梯度呈现出有序排列时, 拥有较少物种的群落将是拥有较多物种的群落的子集, 此时就会呈现出嵌套格局(nestedness pattern)(Patterson & Atmar, 1986; Wang et al, 2010; James et al, 2012; Staniczenko et al, 2013).Beta多样性分解方法就是区分这两种过程对总体beta多样性的作用, 以及探讨这两种过程如何共同影响不同时空维度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011).目前, 有关beta多样性分解的研究快速发展, 这引起了众多生态学家和生物地理学家的关注(Legendre, 2014). ...
... 较之于物种周转组分, 物种丰富度差异组分表示在特定环境梯度下因物种的丧失或增加而引起的群落间物种组成的差异, 可能的驱动机制包括生态位的多样性(diversity of niches)以及导致嵌套格局的生态过程等(Legendre, 2014).其中, 嵌套格局典型的解释机理为选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness)等(Patterson & Atmar, 1986; Ulrich et al, 2009; Wang et al, 2010; Ponisio et al, 2016).例如, 由于物种对环境变化的敏感性不同, 受胁物种容易因为选择性灭绝而在环境压力较高的生境中消失, 但对环境变化具有较高容忍度的物种则能在不同的生境中存活(Gutiérrez-Cánovas et al, 2013; Si et al, 2016).另外, 历史因素同样可能导致不同区域间具有较大的嵌套组分(Dobrovolski et al, 2012).比如在冰川消退过程中, 由于物种扩散能力的差异而导致高纬度地区的物种是低纬度地区物种的子集(Baselga et al, 2012).因此, 通过对beta多样性分解, 我们可以区分不同组分的空间分布格局, 进而解释各自组分的形成机制, 理解可能的生态学和进化历史过程.但是, 在beta多样性分解的研究中, 目前提出的机制并不能解释所有的研究案例, 因此在分析不同生境中不同类群的分布格局时, 需要结合具体的生态和进化背景进行探讨(Baselga, 2010; Legendre, 2014). ...
Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
1
2000
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Phylogenies and community ecology.
2
2002
... 虽然早在20世纪60年代, beta多样性概念已由美国生态学家Robert H. Whittaker提出(Whittaker, 1960, 1972), 用以表示不同研究地点之间生物群落的组成差异(Tuomisto, 2010a, b; Anderson et al, 2011), 但是传统的物种beta多样性分析难以确定不同群落间物种在功能上的组成差异(Petchey & Gaston, 2002; Stegen & Hurlbert, 2011; Villéger et al, 2012).因此, 基于功能alpha多样性(functional alpha diversity) (Tilman, 2001; Petchey & Gaston, 2006; Villéger et al, 2008), 最近有学者提出了功能beta多样性的概念(functional beta diversity), 以期描述不同群落之间功能多样性的组成变化(Swenson et al, 2011a; Si et al, 2016).除了物种在功能上的差异, 物种间还具有不同的进化历史(Faith, 1992; Webb et al, 2002), 因此在度量不同群落间的物种组成变化时, 需要基于谱系beta多样性来探讨群落的构建机制(Cavender-Bares et al, 2009; Swenson et al, 2011b; Cardoso et al, 2014). ...
... 由于物种beta多样性既不考虑物种的进化历史, 也不考虑物种的功能特征(Fukami et al, 2005; Siefert et al, 2013), 因此物种组成相异的两个群落, 可能会具有非常相似的功能特征, 例如具有相同的营养级、相似的形态特征等等(Villéger et al, 2012; Terborgh, 2015; Loiseau et al, 2017; Si et al, 2017).另一方面, 当不同群落的物种beta多样性较高时, 并不一定意味着具有较高的功能beta多样性或者谱系beta多样性, 因为不同的物种可能具有相似的功能特征或者进化历史(Webb et al, 2002; McGill et al, 2006; Si et al, 2016; Liu et al, 2017).例如, Villéger等(2013)对欧洲25个主要河流流域的鱼类分析后发现, 总体beta多样性的差异是由于周转组分导致的, 即物种beta多样性远大于功能beta多样性, 却具有相似的嵌套组分.这表明这些流域的不同鱼类具有相似的功能性状, 从而出现较高的物种周转和较低的功能周转组分.对于物种beta多样性与谱系beta多样性而言, 两者亦反映了各自不同的维持机理(Losos, 1996; Webb, 2000; Graham & Fine, 2008; Swenson et al, 2011b).例如, Cardoso等(2014)对欧洲的兽类和葡萄牙亚速尔群岛的节肢动物研究发现, 谱系beta多样性的丰富度差异组分变化反映了扩散限制和生境过滤的作用; 而物种替换组分的变化可能与环境的干扰有关.因此, 对于不同的群落, 分别基于物种、功能和谱系beta多样性进行综合分析, 将能更为深入地理解驱动不同群落间组成变化的生态学机理(Devictor et al, 2010; Villéger et al, 2013; Cardoso et al, 2014; Bishop et al, 2015). ...
Multiscale partitioning of small mammal β-diversity provides novel insights into the Quaternary faunal history of Qinghai-Tibetan Plateau and Hengduan Mountains.
1
2016
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
Vegetation of the Siskiyou Mountains, Oregon and California.
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
物种丰富度垂直分布格局及影响机制. 动物学杂志,
1
2013
... 物种丰富度沿海拔梯度的分布格局包括随海拔升高而呈现单调递减模式、单峰模式和单调递增模式等(Rahbek, 2005; 吴永杰和雷富民, 2013; Colwell et al, 2016; Wang et al, 2017), 这同样强烈地吸引了众多生态学家和生物地理学家的关注(Rahbek, 1995; Colwell & Lees, 2000; Lomolino, 2001; McCain, 2009).不过相较于alpha或gamma多样性, 探讨beta多样性随海拔梯度变化的研究至今还相对比较薄弱(Tang et al, 2012; Mori et al, 2013).对于较为常见的单峰模式, 诸多研究表明这可能仅仅是由于低山物种和高山物种在中度海拔地区交汇而发生了物种周转(Brown, 2001; Herzog et al, 2005; McCain & Beck, 2016).由于扩散限制、环境过滤等过程均可能决定beta多样性的海拔梯度分布(Qian & Ricklefs, 2007; Jankowski et al, 2009; Andrew et al, 2012; Liu et al, 2015), 所以需要对组成beta多样性的物种周转和嵌套这两种过程分别进行探讨(Baselga, 2010; Carvalho et al, 2011).例如, Bishop等(2015)对南非高山蚂蚁群落的研究发现, 物种组成差异随着海拔升高而增大, 这主要是由物种周转引起的.标准化后的物种周转和嵌套组分随海拔升高两者呈现相反的趋势, 表明群落的构建是由确定性过程(deterministic process)导致的.Foord和Dippenaar-Schoeman (2016)对南非山地蜘蛛群落的研究发现, 虽然总体beta多样性主要都是由物种周转组成, 但物种周转和嵌套组分对东西坡向的群落组成差异的贡献程度不同.标准化后的群落格局与随机群落没有显著性差别, 表明随机过程(stochastic process)对蜘蛛群落的构建具有重要作用.随着beta多样性分解方法的发展, 单独分析组成beta多样性的周转和嵌套过程将有助于更好地理解海拔梯度上的群落构建机制(Bryant et al, 2008; Wang et al, 2012; Marini et al, 2013; Mori et al, 2015). ...
a) Complimentary analysis of metacommunity nestedness and diversity partitioning highlights the need for a holistic conservation strategy for highland lake fish assemblages.
1
2015
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...
b) Latitudinal differences in species abundance distributions, rather than spatial aggregation, explain beta-diversity along latitudinal gradients.
1
2015
... 物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一(Pianka, 1966; Willig et al, 2003; Hillebrand, 2004).同时, beta多样性的纬度梯度分布格局也逐渐引起众多生态学家的关注(Koleff et al, 2003b; Qian & Ricklefs, 2007; Melo et al, 2009; Xu et al, 2015b).Koleff等(2003b)通过综合研究全球多个类群后发现, 仅部分研究证实了低纬度地区具有较高的beta多样性.这可能是由于衡量beta多样性的指数众多且意义并不一致所致(Koleff et al, 2003a).另外, 物种周转和嵌套(物种丧失或增加)两个过程会同时影响beta多样性分布格局(Baselga, 2010; Carvalho et al, 2012).例如, Baselga等(2012)发现全球两栖类的总体beta多样性(Sørensen相异性指数)并不随纬度变化而变化, 但是把总体beta多样性分解之后, 则可以清晰地发现低纬度地区具有更高的物种周转, 而高纬度地区具有更高的嵌套组分.该研究凸显了大尺度研究中历史气候因子对物种分布的重要性, 即随着冰川的消退, 两栖类逐渐往两极扩散而导致高纬度地区的物种组成是低纬度地区的子集, 而低纬度地区由于受到冰川的影响较小而具有较高的成种效应, 从而导致较高的物种周转(Baselga et al, 2012).最近不少基于纬度梯度的研究都发现历史气候因素是决定beta多样性分布格局的主要因子, 并建议在分析纬度梯度时把形成beta多样性的两个组成过程进行独立分析(Hortal et al, 2011; Leprieur et al, 2011; Dobrovolski et al, 2012; Fernandez et al, 2016). ...
Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia grassland.
1
2014
... 相较于国际前沿, 我国的beta多样性分解研究仍较为落后.在本文所统计的349篇论文中, 我国学者的研究仅占2%, 研究区域主要在黄土高原(王世雄等, 2013, 2016)、山东半岛(Wang et al, 2014)、内蒙古草原(Zhang et al, 2014)、浙江千岛湖(Si et al, 2015, 2016)、青藏高原-横断山脉(Wen et al, 2016)和云南西双版纳(Beng et al, 2016)等地.此外, 另有两篇论文报道了云南高原湖泊中淡水鱼类的研究(Su et al, 2015; Xu et al, 2015a).我国从南到北覆盖了热带至寒温带的一系列生物地理区域, 具有丰富的生境多样性.这些特殊的地理条件造就了我国极高的物种多样性, 开展有关beta多样性分解的研究将有利于更为深入地了解我国生物多样性的分布格局及其维持机制. ...



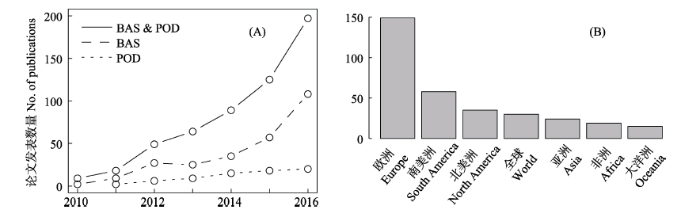
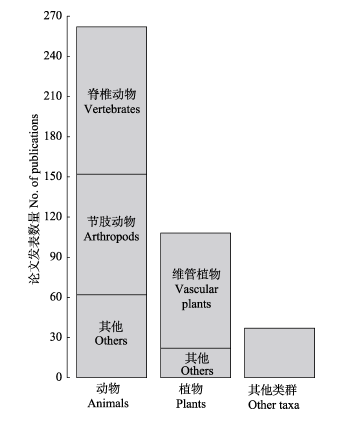
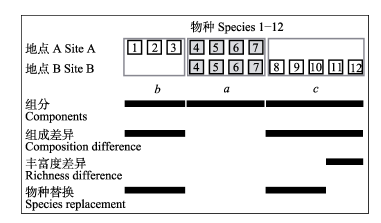
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}