解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容。自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 )。周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响。这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用。然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题。
局域尺度上的物种共存问题是群落构建和生物多样性维持机制研究的重要组成部分。人们常常将物种共存、群落构建和生物多样性维持相提并论、混淆使用, 但实际上物种共存问题与后两者的研究存在着研究尺度上的差异。物种共存通常以较小尺度上的群落为研究对象, 根据Vellend (2010) 的群落综合概念框架, 其侧重于非生物环境和生物选择下群落内相互作用的组分种之间的关系, 而较少考虑局域生态漂变的影响、物种扩散的作用以及物种形成的贡献。简而言之, 物种共存理论主要研究生物和非生物选择下群落组分种的生态位分化和适合度差异。
本文首先简要回顾了经典的物种共存理论及其局限性; 在此基础上, 介绍了Chesson (2000) 提出的当代物种共存理论, 包括理论框架的定义和部分实验验证工作; 进而探讨当代物种共存理论在群落生态学和全球变化生态学中的应用, 包括如何更好地从格局来推断内在的群落构建过程和生物多样性维持机制, 以及如何应用当代物种共存理论来预测气候变化的影响等; 最后通过介绍当代物种共存理论与当代生态位理论、群落构建和生物多样性维持机制理论之间的联系, 试图阐明以物种共存为核心的群落生态学未来可能的研究方向。
1 经典的物种共存理论
经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 )。Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche)。尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的。Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 )。俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 )。
在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求。在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化。MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等。极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 )。换言之, 共存物种之间的生态位不能无限重叠。从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础。Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 )。
另外一条主线以机理模型为基础, 主要是MacArthur提出的消费者-资源模型(MacArthur, 1972 )。机理模型相对现象模型的优势在于同时考虑了物种对环境的“需求”和“影响”, 这也是生态位概念的一次完善, 是Grinnell生态位概念和Elton生态位概念的综合。以机理模型为基础解释物种共存的工作始于MacArthur (1972), 而让该模型人所共知的是Tilman (1980, 1982), 比如其提出的R* 理论和资源比例假说便是生态学发展史上的一个重要节点。在机理模型中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响。换言之, 物种受不同资源的限制, 且该物种消耗最多的资源恰好是限制其增长的那个资源。理论上, MacArthur的消费者-资源模型可以解释无数物种在群落内的共存问题(Tilman, 1982 )。然而, 这种理论上的完美性伴随的是其在实际群落中较低的可操作性, 较难进行实验验证。
无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架。然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 )。研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要。因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型。回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的。对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 )。此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键。同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用。这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )?
因此, 在20世纪末群落生态学中是否存在普适性的理论成为了争论的焦点。回溯这次争论, 我们发现群落生态学迎来了新一轮的快速发展。
2 当代物种共存理论
当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律。伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架。一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响。尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖。因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述。下面, 我们单就Chesson的当代物种共存理论作出详尽说明。
2.1 生态位差异和平均适合度差异
相对于中性理论假定物种功能等同, 当代物种共存理论将物种之间的差异分为两类: 生态位差异(niche difference, ND)和平均适合度差异(average fitness difference, AFD) (Chesson, 2000 ; 2013 ) (图1 )。生态位差异和平均适合度差异是两个非常抽象的概念, 均不涉及具体的物种差异而是对真实的物种差异的高度概括和抽象。对于生态位差异, 我们可以作如下理解: 深根系物种与浅根系物种通过根系的长短实现对资源的分化利用; 对于平均适合度差异, 类似地我们可以将其设想为物种对相同资源在利用效率上的差异(HilleRisLambers et al, 2012 )。 由此可见, 当代物种共存理论中的生态位差异包括了经典的物种共存理论的精髓, 即生态位分化 (Letten et al, 2017 )。对于稳定的物种共存而言, 生态位差异越大越好, 而平均适合度差异则越小越好。换言之, 生态位差异越大越有利于稳定共存, 而平均适合度差异越大则越有利于竞争排除。对于任何一个现实的群落, 生态位差异和平均适合度差异的相对大小决定了物种是稳定共存还是会发生竞争排除(Adler et al, 2007 ), 这是当代物种共存理论最为核心的内容。当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局。从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 )。
图1 当代物种共存理论框架。物种间差异分为生态位差异和平均适合度差异, 前者促进物种共存, 后者利于竞争排除。物种能否稳定共存取决于生态位差异和平均适合度差异的相对大小: 灰色区域, 生态位差异大于平均适合度差 异——稳定共存; 白色区域, 平均适合度差异大于生态位差异——竞争排除。生态位差异对应稳定化机制, 平均适合度差异对应均等化机制。
Fig. 1 The conceptual diagram of contemporary coexistence theory. Species differences are categorized into two groups: niche differences and average fitness differences. Niche differences maintain species stable coexistence, and average fitness differences drive competitive exclusion. For a given community, the balance between niche differences and average fitness differences determines the outcome of competition. In the gray region, niche differences are larger than average fitness differences, which results in stable coexistence. In the white region, average fitness differences are stronger than niche differences, which results in competitive exclusion. Niche differences correspond to stabilizing mechanisms, and average fitness differences correspond to equalizing mechanisms.
2.2 理论定义
如上所述, 生态位差异(ND)和平均适合度差异(AFD)是两类抽象的物种间差异, 如何量化这两类差异是当代物种共存理论框架的关键。Chesson (2000, 2013)在提出当代物种共存理论框架的同时, 给出了基于现象模型即Lotka-Volterra竞争模型的计算公式:
(1)
(2)
其中, αii 和αjj 为种内竞争系数, αij 和αji 为种间竞争系数; ρ 表示生态位重叠, 1 - ρ 即为生态位差异; κj /κi
表示适合度差异(κ 读音为Kappa, 不同于Lotka- Volterra竞争模型中的环境容纳量K )。物种稳定共存的标准为种内竞争系数同时大于种间竞争系数: αii > αji 并且αjj > αij 。
Carroll等(2011) 从机理模型即MacArthur的消费者-资源模型出发, 基于种群的入侵增长率, 提出了另外一种量化生态位差异(ND)和平均适合度差异(AFD)的方法:
(3)
(4)
其中, n 为群落内的物种数, S 表示某物种对竞争的敏感性, 通过下面的公式计算:
(5)
其中, pgrinvasion 为种群的入侵增长率, pgralone 为种群的内禀增长率。实际上, 此处的生态位差异ND是1减去物种竞争敏感性S 的几何平均数, 而平均适合度差异AFD则是物种竞争敏感性S 的几何标准差。物种稳定共存需要满足AFD ≤ 1/(1 - ND)。
生态位差异的取值范围为0-1, 0表示物种生态位完全重叠, 1表示物种生态位完全分离; 平均适合度差异的取值范围为≥ 1, 1表示物种间的适合度相等即没有适合度的差异。因此, 当生态位差异ND = 0且平均适合适度差异AFD = 1的时候, 群落即为中性。
自2000年Chesson提出当代物种共存的理论框架之后将近10年左右的时间内, 该框架一直停留在理论层面而鲜有实验验证, 这可能有以下3个方面的原因: 第一, 在实际工作中无论是测量竞争系数还是种群的内禀增长率和入侵增长率都具有很大
的挑战性, 一定程度上限制了对生态位差异和平均适合度差异的量化和理解; 第二, Hubbell于2001年提出的群落中性理论因其简洁性和解释力改变了群落生态学的发展轨迹, 使得很多优秀的群落生态学家的注意力被吸引到中性理论与生态位理论的争论之上; 第三, Chesson于2000年发表的原始论文相对比较晦涩, 虽然这一点并不是主要原因, 但从整个学界的反馈来看确实也是一个不容忽视的影响因素。
2.3 稳定化机制和均等化机制
物种的生态位差异导致了稳定化机制, 平均适合度差异导致了均等化机制。如上所述, 在当代物种共存理论当中, 稳定化机制与均等化机制共同决定了物种能否实现稳定共存(Chesson, 2000 , 2013 ): 稳定化机制通过物种之间的生态位分化减弱或者消除竞争的影响, 而均等化机制通过降低物种之间的适合度差异来促进共存。
种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 )。在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 )。稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等。
均等化机制是通过降低物种之间的适合度差异或者物种在竞争能力方面的差异来促进共存的。以对资源的竞争为例, 物种对资源的竞争能力可以通过R *值来表示, 即某物种在群落中能够维持下去所需的最低的资源水平(Tilman, 1982 )。当多个物种同时利用相同的限制性资源时, 具有最低R *值的物种将竞争排除掉所有其他的物种。因此, 任何能减少物种之间R *值差异的因子都可被归为均等化的机制, 比如对竞争优势种的部分去除等。不同于稳定化机制, 均等化机制不会导致负密度制约。经典物种共存理论的工作几乎都是以稳定化机制为研究对象的, 强调物种之间的生态位分化, 却很少关注均等化机制。实际上, 均等化机制的研究是在群落中性理论提出后才得到了较为快速的发展的(Chave, 2004 ; Turnbull et al, 2008 ; Ostling, 2012 )。
2.4 实验验证
对当代物种共存理论的实验验证是该理论框架日趋成熟和为人所接受的重要一步。下面我们将简要回顾部分验证工作, 一方面阐明该领域所取得的进展, 另一方面是希望通过研究案例的方式介绍如何开展实验来验证当代物种共存理论。
从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率。目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据。Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数。如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异。结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性。Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异。实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局。
相比于一年生草地植物群落, 当代物种共存理论在多年生植物群落中的验证得益于长期观测数据的积累(Adler et al, 2010 ; Chu et al, 2014 )。Chu和Adler (2015)收集了北美5套长期定位观测的草地野外数据, 构建了种群统计学参数模型(包括存活率、生长率和更新率), 然后通过积分投影模型(integral projection model)的手段计算种群入侵增长率, 进而量化生态位差异和平均适合度差异。分析结果同样支持了当代物种共存理论的预测, 同时还发现在这些自然群落中物种间的生态位差异非常大而平均适合度差异相对比较小, 这也就意味着这些多年生草地植物群落受物种间生态位差异的强烈影响, 相对来说非常稳定(Chu & Adler, 2015 )。
此外, 还有研究者采用其他更易操作的实验系统, 比如微生物系统(Zhao et al, 2016 )和藻类系统(Narwani et al, 2013 )来验证当代物种共存理论。所有这些研究都从实验的角度支持了当代物种共存理论的预期, 也为当代物种共存理论的传播及其被认可提供了基础。
3 当代物种共存理论的应用
当代物种共存理论在提出后先是度过了将近10年的沉寂期, 此后便引起了越来越多的生态学家的兴趣和关注。这不仅仅是因为它为物种共存提供了一个综合性、普适性的理论框架, 更为重要的是具有非常重要的应用价值, 为解决生态学中很多关键的科学问题都提供了新的思路和视角(Chesson, 2013 )。在此, 我们列举两个方面的应用: 群落构建机制的推断和预测气候变化的间接影响。
3.1 基于格局推断过程
在竞争能力上可能差异较大), 则会使群落谱系呈现聚集的格局。换言之, 竞争和环境过滤都有可能产生聚集式的格局, 上述“一一对应”的关系并非是普遍存在的(Mayfield & Levine, 2010 )。
当代物种共存理论为解决这个问题提供了最直接和最有效的方法。在当代物种共存理论中, 物种间的差异分为生态位差异和平均适合度差异, 而这两类差异真实地代表了物种共存的两类驱动力。我们可以将这两类差异视为抽象的性状。如果能够将这两类抽象性状与具体性状如根系深浅结合在一起(谱系距离可依此类推), 找到二者之间的联系, 就能有效地揭示内在的群落构建机制(Adler et al, 2013 ; Kraft et al, 2015 )。比如, 如果平均适合度差异与谱系距离差异呈现负相关关系, 那么就意味着亲缘关系越近的物种其适合度差异越大, 而平均适合度差异越大物种间竞争越强, 越不容易稳定共存。目前已有一些工作报道了相关的研究成果。以一年生草本植物为研究对象, Kraft等(2015)发现在其选择的11个功能性状中, 大部分功能性状与平均适合度差异之间的相关性很强, 而与生态位差异的相关性不明显, 这意味着这些功能性状主要体现的是物种之间的适合度差异。以绿藻群落为研究对象, Narwani等(2013)发现物种间的谱系距离与生态位差异和平均适合度差异均无明显相关关系, 与物种是否共存也不相关。这表明基于谱系距离来推断竞争的重要性并不合适, 即上述的“亲缘关系越近竞争越强, 亲缘关系越远竞争越弱”的假定可能是不成立的。Germain等(2016)也发现谱系距离与生态位差异和平均适合度差异并无一一对应的关系。
3.2 预测气候变化的间接影响
温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 )。这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 )。那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 )。由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大。换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度。基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小。这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响。
4 研究展望
从前面的介绍中我们可以看到, 当代物种共存理论为局域群落物种共存提供了一个普适性的理论框架: 从纷繁复杂的具体的共存机制中抽象出物种的两类差异, 生态位差异促进物种共存, 平均适合度差异促进竞争排除。所有有关物种共存的研究均可以放到这个框架中进行理解和解释。然而, 这样一个“万能”的理论本身实际上也并非是“万能”的, 而是存在着一定的局限。下面我们将从3个方面入手探讨如何更好地应用当代物种共存理论来解决群落生态学家所关注的问题。
4.1 对理论框架本身的检验
经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退。然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落。
在当代物种共存理论中, 物种间竞争的结局有两类: 稳定共存或者竞争排除。而在自然群落中, 要观察到竞争排除或者稳定共存通常都需要相对较长的时间, 这实际上是很难实现的。但任何一个物种种群的增长或消亡最终都要体现在种群的统计学参数上, 包括个体的存活率、生长率和繁殖率, 所以相对于观察稳定共存或竞争排除, 转而关注竞争对种群统计学参数的影响是目前检验当代物种共存理论的替代方法(Harms et al, 2000 ; Funk & Wolf, 2016 )。这是其一。
同时, 从理论上来说, 量化生态位差异和平均适合度差异需要测量竞争系数或者种群的入侵增长率。而在复杂的自然系统中, 这一点同样很难实现。为此我们同样需要寻找一个替代的方案。对于功能性状而言, 假设有两个物种A和B, 其性状值分别为TA 和TB 。人们提出了用物种间绝对性状距离(性状间差异的绝对值, |TA -TB |)来代表生态位差异, 而用物种间等级性状距离(性状间差异, TA -TB )来代表平均适合度差异。换言之, 绝对性状距离是没有方向的, 只要存在差异就有利于物种的共存, 体现的是物种间生态位的分化; 而等级性状距离具有方向, 体现的是物种间适合度的差异(Kunstler et al, 2012 )。这是其二。
通过上述两个方面的替代, 就可以对当代物种共存理论开展行之有效的验证。利用法国全国林业调查的数据, Kunstler等(2012) 发现物种间的竞争强度主要受性状等级(等级性状距离)所驱动, 而与性状或谱系的相似性(性状相似性即绝对性状距离)没有明显的相关关系。Gross等(2015)在新西兰开展的草地实验表明, 性状间的差异主要体现的是竞争的等级, 即在该实验中所选取的性状如植株高度和特定叶面积指数代表的主要是物种间的平均适合度差异。
需要指出的是, 这种替代方法只是对当代物种共存理论的一个近似验证。随着长期观测数据的积累, 在复杂自然群落中直接检验当代物种共存理论将会逐渐成为现实, 也为更准确和全面地揭示群落内在的构建机制提供了可能。
4.2 与具体物种共存机制研究的结合
当代物种共存理论是对过去所有具体共存机制的综合、抽象和升华, 生态位差异和平均适合度差异是对物种功能性状的简化处理。伴随当代物种共存理论“万能”解释能力的是对具体物种共存机制的忽略。例如, 假定有一个自然群落, 我们知道了其物种间生态位差异是0.9, 平均适合度差异是2.6, 那么我们能够从中得到多少有关该群落的有价值的信息?很少!再者, 这个群落主要受什么因素 的影响?物种间的性状和谱系差异如何?群落对于环境条件的变化会做出什么样的响应?都不得而知。
将当代物种共存理论与具体共存机制相结合是当代物种共存理论发展的必然。而具体物种共存机制正是生态位理论的核心。生态位理论按照生态位的定义大体上可以划分为经典生态位理论和当代生态位理论, 这在本文的第一部分均有介绍。经典生态位理论主要是以现象模型如Lotka-Volterra模型为基础, 侧重物种对环境的需求。当代生态位理论以机理模型如消费者-资源模型为基础, 既包含了物种对环境的需求, 又包含了物种对环境的影响。这里我们侧重当代物种共存理论与当代生态位理论的结合, 而这两个理论正是通过生态位的概念密切联系在一起(Kleinhesselink & Adler, 2015 ; Letten et al, 2017 )。在当代生态位理论中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响。那么这3个因素与当代物种共存理论中的生态位差异(稳定化机制)和平均适合度差异(均等化机制)有什么关系?Letten等(2017)发现资源的供给率体现的是均等化的机制, 物种对环境的影响体现的是稳定化的机制, 而物种对环境的需求可能是均等化机制也可能是稳定化机制。这也就是说当代物种共存理论与当代生态位理论确实是可以有机地结合在一起的; 也只有将二者紧密结合才能最大限度地发挥二者的作用, 从而为更全面地理解物种共存提供基础。
4.3 与基于过程的群落构建机制研究的整合
从格局推断过程因其不具有唯一性而显得非常复杂和不可靠。比如前面所介绍的从功能性状或谱系结构推断群落内在的构建机制。群落生态学在一定程度上过于注重对格局的发现和描述, 比如物种多度分布, 而忽视了对内在过程的深入探讨。这解释了中性理论在其提出后的十多年里在群落生态学的研究中占据的主导地位。如果一直按照“从格局推断过程”这一思路发展下去, 那么群落生态学可能就只会原地踏步。当代物种共存理论在这个方面前进了一步, 其与功能性状和谱系关系的结合将能更可靠地揭示出竞争的相对重要性。
Vellend (2010, 2016)提出的基于过程的群落构建框架被认为是关于群落构建机制普适性理论的有益尝试。朱璧如和张大勇(2011)对该理论构建框架进行了详细的介绍。简言之, 该理论框架以过程为出发点, 认为群落的构建由4个基本过程所共同决定: 选择、漂变、扩散和成种。在不同的群落中各个过程的相对重要性可能不同。其中, 平衡选择(负密度制约或负频度制约)所对应的正是我们所熟知的生态位理论和当代物种共存理论。如何将当代物种共存理论和生态位理论与基于过程的群落构建框架有机地结合在一起, 从而揭示群落构建和生物多样性维持的内在机制, 是未来研究工作的一个重要发展方向。随着当代物种共存理论和基于过程的群落构建框架日趋成熟和完善, 群落生态学正在迎来一个新的发展时期。
(责任编委: 马克平 责任编辑: 黄祥忠)
群落生态学的一项重要任务是从群落格局推断内在的群落构建机制。一是基于功能性状的分布格局来探讨环境过滤和竞争在群落构建过程中的相对作用: 如果共存物种的性状分布相对于零模型来说表现为聚集的格局, 那么推断环境过滤是群落构建的主要驱动力; 反之, 如果性状分布表现为发散的格局, 那么物种之间的竞争则起着主导作用(Ackerly & Cornwell, 2007 )。二是基于亲缘关系的谱系群落生态学, 与基于功能性状的方法类似: 如果生境过滤作用占主导地位, 那么相似生境将筛选出适应能力相似、亲缘关系较近的物种, 从而表现为谱系的聚集; 相反, 竞争排斥会使得生态位相似的物种无法共存于同一环境, 群落内物种的亲缘关系则较远, 表现为谱系的发散(Webb et al, 2002 )。这种从格局推断过程的方法在2002年Webb等发表其综述性论文后被普遍采纳。同时, 在谱系群落生态学中有一个非常常见的重要前提假定, 即亲缘关系越近竞争越强, 亲缘关系越远竞争越弱。
然而, 这种简洁的“一一对应”的关系在自然群落中是否成立?Mayfield和Levine (2010)对此提出了质疑: 竞争如果排除的是亲缘关系较近的物种(如亲缘关系较近的物种具有相似的生态位需求), 那么会使群落谱系呈发散格局; 反之, 竞争若排除的是亲缘关系较远的物种(如亲缘关系较远的物种
致谢: 本文是在2016年“第十届海峡两岸森林动态样区研讨会”报告的基础上整理而成, 感谢中国科学院植物研究所马克平研究员的支持和鼓励。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ackerly DD Cornwell WK 2007 ) A trait-based approach to community assembly: partitioning of species trait values into within- and among-community components.
Ecology Letters , 10 , 135 -145 .
[本文引用: 1]
[2]
Adler PB Dalgleish HJ Ellner SP 2012 ) Forecasting plant community impacts of climate variability and change: when do competitive interactions matter?
Journal of Ecology , 100 , 478 -487 .
[本文引用: 2]
[3]
Adler PB Ellner SP Levine JM 2010 ) Coexistence of perennial plants: an embarrassment of niches.
Ecology Letters , 13 , 1019 -1029 .
[本文引用: 1]
[4]
Adler PB Fajardo A Kleinhesselink AR Kraft NJB 2013 ) Trait-based tests of coexistence mechanisms.
Ecology Letters , 16 , 1294 -1306 .
[本文引用: 1]
[5]
Adler PB HilleRisLambers J Levine JM 2007 ) A niche for neutrality.
Ecology Letters , 10 , 95 -104 .
[本文引用: 2]
[6]
Angert AL LaDeau SL Ostfeld RS 2013 ) Climate change and species interactions: ways forward.
Annals of the New York Academy of Sciences , 1297 , 1 -7 .
[本文引用: 1]
[7]
Bell G 2001 ) Neutral macroecology.
Science , 293 , 2413 -2418 .
[本文引用: 1]
[8]
Carroll IT Cardinale BJ Nisbet RM 2011 ) Niche and fitness differences relate the maintenance of diversity to ecosystem function.
Ecology , 92 , 1157 -1165 .
[本文引用: 2]
[9]
Chase JM Leibold MA 2003 ) Ecological Niches : Linking Classical and Contemporary Approaches. University of Chicago Press, Chicago .
[本文引用: 2]
[10]
Chave J 2004 ) Neutral theory and community ecology.
Ecology Letters , 7 , 241 -253 .
[本文引用: 1]
[11]
Chen L Mi XC Ma KP 2014 ) Niche differentiation and its consequence on biodiversity maintenance in forest communities.
Chinese Bulletin of Life Sciences , 26 , 112 -117 . (in Chinese)
[本文引用: 1]
[陈磊 , 米湘成 , 马克平 (2014 ) 生态位分化与森林群落物种多样性维持研究展望
. 生命科学 , 26 , 112 -117 .]
[本文引用: 1]
[12]
Chesson P 1985 ) Coexistence of competitors in spatially and temporally varying environments: a look at the combined effects of different sorts of variability.
Theoretical Population Biology , 28 , 263 -287 .
[本文引用: 1]
[13]
Chesson P 1994 ) Multispecies competition in variable environments.
Theoretical Population Biology , 45 , 227 -276 .
[本文引用: 1]
[14]
Chesson P 2000 ) Mechanisms of maintenance of species diversity.
Annual Review of Ecology and Systematics , 31 , 343 -366 .
[本文引用: 3]
[15]
Chesson P 2013 ) Species competition and predation. In: Encyclopedia of Sustainability Science and Technology (ed. Meyers RA) , pp. 223 -256. Springer-Verlag, New York.
[本文引用: 4]
[16]
Chu CJ Bartlett M Wang YS He FL Weiner J Chave J Sack L 2016 ) Does climate directly influence NPP globally?
Global Change Biology , 22 , 12 -24 .
[本文引用: 1]
[17]
Chu CJ Adler PB 2015 ) Large niche differences emerge at the recruitment stage to stabilize grassland coexistence.
Ecological Monographs , 85 , 373 -392 .
[本文引用: 2]
[18]
Chu CJ Havstad KM Kaplan N Lauenroth WK McClaran MP Peters DP Vermeire LT Adler PB 2014 ) Life form influences survivorship patterns for 109 herbaceous perennials from six semi-arid ecosystems.
Journal of Vegetation Science , 25 , 947 -954 .
[本文引用: 1]
[19]
Connell JH 1971 ) On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: Dynamics of Populations (eds den Boer PJ, Gradwell G) , pp. 298 -312. Pudoc, Oosterbeek.
[本文引用: 1]
[20]
Elton C 1927 ) Animal Ecology. Sidgwick and Jackson
, London.
[本文引用: 1]
[21]
Funk JL Wolf AA 2016 ) Testing the trait-based community framework: do functional traits predict competitive outcomes?
Ecology , 97 , 2206 -2211 .
[本文引用: 1]
[22]
Gause GF 1934 ) The Struggle of Existence. Williams & Wilkins, Baltimore.
[本文引用: 1]
[23]
Germain RM Weir JT Gilbert B 2016 ) Species coexistence: macroevolutionary relationships and the contingency of historical interactions.
Proceedings of the Royal Society B: Biological Sciences , 283 , 20160047 .
[本文引用: 1]
[24]
Gilman SE Urban MC Tewksbury J Gilchrist GW Holt RD 2010 ) A framework for community interactions under climate change.
Trends in Ecology & Evolution , 25 , 325 -331 .
[本文引用: 1]
[25]
Godoy O Kraft NJB Levine JM 2014 ) Phylogenetic relatedness and the determinants of competitive outcomes.
Ecology Letters , 17 , 836 -844 .
[本文引用: 2]
[26]
Godoy O Levine JM 2014 ) Phenology effects on invasion success: insights from coupling field experiments to coexistence theory.
Ecology , 95 , 726 -736 .
[本文引用: 1]
[27]
Grinnell J 1917 ) The niche-relationships of the California thrasher.
Auk , 34 , 427 -433 .
[本文引用: 2]
[28]
Gross N Liancourt P Butters R Duncan RP Hulme PE 2015 ) Functional equivalence, competitive hierarchy and facilitation determine species coexistence in highly invaded grasslands.
New Phytologist , 206 , 175 -186 .
[29]
Hardin G 1960 ) The competitive exclusion principle.
Science , 131 , 1292 -1297 .
[本文引用: 1]
[30]
Harms K Wright S Calderon O Hernandez A Herre E 2000 ) Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest.
Nature , 404 , 493 -495 .
[本文引用: 1]
[31]
HilleRisLambers J Adler PB Harpole WS Levine JM Mayfield MM 2012 ) Rethinking community assembly through the lens of coexistence theory.
Annual Review of Ecology and Systematics , 43 , 227 -248 .
[本文引用: 1]
[32]
Hubbell SP 2001 ) The Unified Neutral Theory of Biodiversity and Biogeography . Princeton University Press, Princeton .
[本文引用: 1]
[33]
Hutchinson GE 1957 ) Concluding remarks.
Cold Spring Harbor Symposia on Quantitative Biology , 22 , 415 -427 .
[本文引用: 1]
[34]
Janzen DH 1970 ) Herbivores and the number of tree species in tropical forests.
The American Naturalist , 104 , 501 -528 .
[本文引用: 1]
[35]
Kleinhesselink AR Adler PB 2015 ) Indirect effects of environmental change in resource competition models.
The American Naturalist , 186 , 766 -776 .
[本文引用: 2]
[36]
Kraft NJB Godoy O Levine JM 2015 ) Plant functional traits and the multidimensional nature of species coexistence. Proceedings of the National Academy of Sciences,
USA , 112 , 797 -802 .
[本文引用: 2]
[37]
Kunstler G Lavergne S Courbaud B Thuiller W Vieilledent G Zimmermann NE Kattge J Coomes DA 2012 ) Competitive interactions between forest trees are driven by species’ trait hierarchy, not phylogenetic or functional similarity: implications for forest community assembly.
Ecology Letters , 15 , 831 -840 .
[本文引用: 2]
[38]
Lawton JH 1999 ) Are there general laws in ecology?
Oikos , 84 , 177 -192 .
[本文引用: 1]
[39]
Leibold MA 1995 ) The niche concept revisited: mechanistic models and community context.
Ecology , 76 , 1371 -1382 .
[本文引用: 1]
[40]
Letten AD Ke PJ Fukami T 2017 ) Linking modern coexistence theory and contemporary niche theory.
Ecological Monographs , 87 , 161 -177 .
[本文引用: 2]
[41]
Levine JM HilleRisLambers J 2009 ) The importance of niches for the maintenance of species diversity.
Nature , 461 , 254 -257 .
[本文引用: 1]
[42]
Lewin R 1983 ) Santa Rosalia was a goat.
Science , 221 , 636 -639 .
[本文引用: 1]
[43]
Lotka AJ 1925 ) Elements of Physical Biology. Williams & Wilkins Company, Baltimore.
[本文引用: 2]
[44]
MacArthur RH 1958 ) Population ecology of some warblers of northeastern coniferous forests.
Ecology , 39 , 599 -619 .
[本文引用: 2]
[45]
MacArthur RH 1969 ) The theory of the niche. In: Population Biology and Evolution (ed. Lewontin RC) , pp. 159 -176. Syracuse University Press, Syracuse.
[本文引用: 1]
[46]
MacArthur RH 1972 ) Geographical Ecology : Patterns in the Distribution of Species. Princeton University Press, Princeton .
[本文引用: 2]
[47]
MacArthur RH Levins R 1967 ) The limiting similarity, convergence, and divergence of coexisting species.
The American Naturalist , 101 , 377 -385 .
[本文引用: 2]
[48]
May RM MacArthur RH 1972 ) Niche overlap as a function of environmental variability. Proceedings of the National Academy of Science,
USA , 69 , 1109 -1113 .
[本文引用: 1]
[49]
Mayfield MM Levine JM 2010 ) Opposing effects of competitive exclusion on the phylogenetic structure of communities.
Ecology Letters , 13 , 1085 -1093 .
[本文引用: 1]
[50]
Narwani A Alexandrou MA Oakley TH Carroll IT Cardinale BJ 2013 ) Experimental evidence that evolutionary relatedness does not affect the ecological mechanisms of coexistence in freshwater green algae.
Ecology Letters , 16 , 1373 -1381 .
[本文引用: 2]
[51]
Newman EI 1973 ) Competition and diversity in herbaceous vegetation.
Nature , 244 , 310 .
[本文引用: 1]
[52]
Niu HY Wang ZF Lian JY Ye WH Shen H 2011 ) New progress in community assembly: community phylogenetic structure combining evolution and ecology.
Biodiversity Science , 19 , 275 -283 . (in Chinese with English abstract)
[本文引用: 1]
[牛红玉 , 王峥峰 , 练琚愉 , 叶万辉 , 沈浩 (2011 ) 群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究
. 生物多样性 , 19 , 275 -283 .]
[本文引用: 1]
[53]
Niu KC Liu YN Shen ZH He FL Fang JY 2009 ) Community assembly: the relative importance of neutral theory and niche theory.
Biodiversity Science , 17 , 579 -593 . (in Chinese with English abstract)
[本文引用: 2]
[牛克昌 , 刘怿宁 , 沈泽昊 , 何芳良 , 方精云 (2009 ) 群落构建的中性理论和生态位理论
. 生物多样性 , 17 , 579 -593 .]
[本文引用: 2]
[54]
Ostling A 2012 ) Do fitness-equalizing tradeoffs lead to neutral communities?
Theoretical Ecology , 5 , 181 -194 .
[本文引用: 1]
[55]
Siepielski AM McPeek MA 2010 ) On the evidence for species coexistence: a critique of the coexistence program.
Ecology , 91 , 3153 -3164 .
[本文引用: 1]
[56]
Simberloff D Boecklen W 1981 ) Santa Rosalia reconsidered: size ratios and competition.
Evolution , 35 , 1206 -1228 .
[本文引用: 1]
[57]
Soberón J 2007 ) Grinnellian and Eltonian niches and geographic distributions of species.
Ecology Letters , 10 , 1115 -1123 .
[本文引用: 1]
[58]
Strong DR Szyska LA Simberloff D 1979 ) Tests of community-wide character displacement against null hypotheses.
Evolution , 33 , 897 -913 .
[本文引用: 1]
[59]
Tilman D 1980 ) Resource: a graphical-mechanistic approach to competition and predation.
The American Naturalist , 116 , 362 -393 .
[60]
Tilman D 1982 ) Resource Competition and Community Structure . Princeton University Press, Princeton .
[本文引用: 4]
[61]
Turnbull LA Rees M Purves DW 2008 ) Why equalising trade-offs aren’t always neutral?
Ecology Letters , 11 , 1037 -1046 .
[本文引用: 1]
[62]
Vellend BM 2010 ) Conceptual synthesis in community ecology.
The Quarterly Review of Biology , 85 , 183 -206 .
[63]
Vellend M 2016 ) The Theory of Ecological Communities . Princeton University Press, Princeton and Oxford .
[64]
Volterra V 1926 ) Variations and fluctuations of the number of individuals in animal species living together. In: Animal Ecology (ed. Chapman RN) (Reprinted in 1931). McGraw Hill, New York.
[本文引用: 2]
[65]
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002 ) Phylogenies and community ecology.
Annual Review of Ecology and Systematics , 33 , 475 -505 .
[本文引用: 1]
[66]
Zhao L Zhang QG Zhang DY 2016 ) Evolution alters ecological mechanisms of coexistence in experimental microcosms.
Functional Ecology , 30 , 1440 -1446 .
[本文引用: 1]
[67]
Zhou SR Zhang DY 2006 ) Neutral theory in community ecology.
Journal of Plant Ecology (Chinese Version) , 30 , 868 -877 . (in Chinese with English abstract)
[本文引用: 2]
[周淑荣 , 张大勇 (2006 ) 群落生态学的中性理论
. 植物生态学报 , 30 , 868 -877 .]
[本文引用: 2]
[68]
Zhu BR Zhang DY 2011 ) A process-based theoretical framework for community ecology.
Biodiversity Science , 19 , 389 -399 . (in Chinese with English abstract)
[本文引用: 1]
[朱璧如 , 张大勇 (2011 ) 基于过程的群落生态学理论框架
. 生物多样性 , 19 , 389 -339 .]
[本文引用: 1]
A trait-based approach to community assembly: partitioning of species trait values into within- and among-community components.
1
2007
... 群落生态学的一项重要任务是从群落格局推断内在的群落构建机制.一是基于功能性状的分布格局来探讨环境过滤和竞争在群落构建过程中的相对作用: 如果共存物种的性状分布相对于零模型来说表现为聚集的格局, 那么推断环境过滤是群落构建的主要驱动力; 反之, 如果性状分布表现为发散的格局, 那么物种之间的竞争则起着主导作用(Ackerly & Cornwell, 2007 ).二是基于亲缘关系的谱系群落生态学, 与基于功能性状的方法类似: 如果生境过滤作用占主导地位, 那么相似生境将筛选出适应能力相似、亲缘关系较近的物种, 从而表现为谱系的聚集; 相反, 竞争排斥会使得生态位相似的物种无法共存于同一环境, 群落内物种的亲缘关系则较远, 表现为谱系的发散(Webb et al, 2002 ).这种从格局推断过程的方法在2002年Webb等发表其综述性论文后被普遍采纳.同时, 在谱系群落生态学中有一个非常常见的重要前提假定, 即亲缘关系越近竞争越强, 亲缘关系越远竞争越弱. ...
Forecasting plant community impacts of climate variability and change: when do competitive interactions matter?
2
2012
... 温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 ).这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
... ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
Coexistence of perennial plants: an embarrassment of niches.
1
2010
... 相比于一年生草地植物群落, 当代物种共存理论在多年生植物群落中的验证得益于长期观测数据的积累(Adler et al, 2010 ; Chu et al, 2014 ).Chu和Adler (2015)收集了北美5套长期定位观测的草地野外数据, 构建了种群统计学参数模型(包括存活率、生长率和更新率), 然后通过积分投影模型(integral projection model)的手段计算种群入侵增长率, 进而量化生态位差异和平均适合度差异.分析结果同样支持了当代物种共存理论的预测, 同时还发现在这些自然群落中物种间的生态位差异非常大而平均适合度差异相对比较小, 这也就意味着这些多年生草地植物群落受物种间生态位差异的强烈影响, 相对来说非常稳定(Chu & Adler, 2015 ). ...
Trait-based tests of coexistence mechanisms.
1
2013
... 当代物种共存理论为解决这个问题提供了最直接和最有效的方法.在当代物种共存理论中, 物种间的差异分为生态位差异和平均适合度差异, 而这两类差异真实地代表了物种共存的两类驱动力.我们可以将这两类差异视为抽象的性状.如果能够将这两类抽象性状与具体性状如根系深浅结合在一起(谱系距离可依此类推), 找到二者之间的联系, 就能有效地揭示内在的群落构建机制(Adler et al, 2013 ; Kraft et al, 2015 ).比如, 如果平均适合度差异与谱系距离差异呈现负相关关系, 那么就意味着亲缘关系越近的物种其适合度差异越大, 而平均适合度差异越大物种间竞争越强, 越不容易稳定共存.目前已有一些工作报道了相关的研究成果.以一年生草本植物为研究对象, Kraft等(2015)发现在其选择的11个功能性状中, 大部分功能性状与平均适合度差异之间的相关性很强, 而与生态位差异的相关性不明显, 这意味着这些功能性状主要体现的是物种之间的适合度差异.以绿藻群落为研究对象, Narwani等(2013)发现物种间的谱系距离与生态位差异和平均适合度差异均无明显相关关系, 与物种是否共存也不相关.这表明基于谱系距离来推断竞争的重要性并不合适, 即上述的“亲缘关系越近竞争越强, 亲缘关系越远竞争越弱”的假定可能是不成立的.Germain等(2016)也发现谱系距离与生态位差异和平均适合度差异并无一一对应的关系. ...
A niche for neutrality.
2
2007
... 相对于中性理论假定物种功能等同, 当代物种共存理论将物种之间的差异分为两类: 生态位差异(niche difference, ND)和平均适合度差异(average fitness difference, AFD) (Chesson, 2000 ; 2013 ) (图1 ).生态位差异和平均适合度差异是两个非常抽象的概念, 均不涉及具体的物种差异而是对真实的物种差异的高度概括和抽象.对于生态位差异, 我们可以作如下理解: 深根系物种与浅根系物种通过根系的长短实现对资源的分化利用; 对于平均适合度差异, 类似地我们可以将其设想为物种对相同资源在利用效率上的差异(HilleRisLambers et al, 2012 ). 由此可见, 当代物种共存理论中的生态位差异包括了经典的物种共存理论的精髓, 即生态位分化 (Letten et al, 2017 ).对于稳定的物种共存而言, 生态位差异越大越好, 而平均适合度差异则越小越好.换言之, 生态位差异越大越有利于稳定共存, 而平均适合度差异越大则越有利于竞争排除.对于任何一个现实的群落, 生态位差异和平均适合度差异的相对大小决定了物种是稳定共存还是会发生竞争排除(Adler et al, 2007 ), 这是当代物种共存理论最为核心的内容.当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局.从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 ). ...
... ), 这是当代物种共存理论最为核心的内容.当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局.从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 ). ...
Climate change and species interactions: ways forward.
1
2013
... 温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 ).这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
Neutral macroecology.
1
2001
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
Niche and fitness differences relate the maintenance of diversity to ecosystem function.
2
2011
... Carroll等(2011) 从机理模型即MacArthur的消费者-资源模型出发, 基于种群的入侵增长率, 提出了另外一种量化生态位差异(ND)和平均适合度差异(AFD)的方法: ...
... 经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退.然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落. ...
2
2003
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
... 在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求.在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化.MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
Neutral theory and community ecology.
1
2004
... 均等化机制是通过降低物种之间的适合度差异或者物种在竞争能力方面的差异来促进共存的.以对资源的竞争为例, 物种对资源的竞争能力可以通过R *值来表示, 即某物种在群落中能够维持下去所需的最低的资源水平(Tilman, 1982 ).当多个物种同时利用相同的限制性资源时, 具有最低R *值的物种将竞争排除掉所有其他的物种.因此, 任何能减少物种之间R *值差异的因子都可被归为均等化的机制, 比如对竞争优势种的部分去除等.不同于稳定化机制, 均等化机制不会导致负密度制约.经典物种共存理论的工作几乎都是以稳定化机制为研究对象的, 强调物种之间的生态位分化, 却很少关注均等化机制.实际上, 均等化机制的研究是在群落中性理论提出后才得到了较为快速的发展的(Chave, 2004 ; Turnbull et al, 2008 ; Ostling, 2012 ). ...
生态位分化与森林群落物种多样性维持研究展望
1
2014
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
生态位分化与森林群落物种多样性维持研究展望
1
2014
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
Coexistence of competitors in spatially and temporally varying environments: a look at the combined effects of different sorts of variability.
1
1985
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
Multispecies competition in variable environments.
1
1994
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Mechanisms of maintenance of species diversity.
3
2000
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
... 相对于中性理论假定物种功能等同, 当代物种共存理论将物种之间的差异分为两类: 生态位差异(niche difference, ND)和平均适合度差异(average fitness difference, AFD) (Chesson, 2000 ; 2013 ) (图1 ).生态位差异和平均适合度差异是两个非常抽象的概念, 均不涉及具体的物种差异而是对真实的物种差异的高度概括和抽象.对于生态位差异, 我们可以作如下理解: 深根系物种与浅根系物种通过根系的长短实现对资源的分化利用; 对于平均适合度差异, 类似地我们可以将其设想为物种对相同资源在利用效率上的差异(HilleRisLambers et al, 2012 ). 由此可见, 当代物种共存理论中的生态位差异包括了经典的物种共存理论的精髓, 即生态位分化 (Letten et al, 2017 ).对于稳定的物种共存而言, 生态位差异越大越好, 而平均适合度差异则越小越好.换言之, 生态位差异越大越有利于稳定共存, 而平均适合度差异越大则越有利于竞争排除.对于任何一个现实的群落, 生态位差异和平均适合度差异的相对大小决定了物种是稳定共存还是会发生竞争排除(Adler et al, 2007 ), 这是当代物种共存理论最为核心的内容.当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局.从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 ). ...
... 物种的生态位差异导致了稳定化机制, 平均适合度差异导致了均等化机制.如上所述, 在当代物种共存理论当中, 稳定化机制与均等化机制共同决定了物种能否实现稳定共存(Chesson, 2000 , 2013 ): 稳定化机制通过物种之间的生态位分化减弱或者消除竞争的影响, 而均等化机制通过降低物种之间的适合度差异来促进共存. ...
4
2013
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
... 物种的生态位差异导致了稳定化机制, 平均适合度差异导致了均等化机制.如上所述, 在当代物种共存理论当中, 稳定化机制与均等化机制共同决定了物种能否实现稳定共存(Chesson, 2000 , 2013 ): 稳定化机制通过物种之间的生态位分化减弱或者消除竞争的影响, 而均等化机制通过降低物种之间的适合度差异来促进共存. ...
... 当代物种共存理论在提出后先是度过了将近10年的沉寂期, 此后便引起了越来越多的生态学家的兴趣和关注.这不仅仅是因为它为物种共存提供了一个综合性、普适性的理论框架, 更为重要的是具有非常重要的应用价值, 为解决生态学中很多关键的科学问题都提供了新的思路和视角(Chesson, 2013 ).在此, 我们列举两个方面的应用: 群落构建机制的推断和预测气候变化的间接影响. ...
... 经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退.然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落. ...
Does climate directly influence NPP globally?
1
2016
... 温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 ).这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
Large niche differences emerge at the recruitment stage to stabilize grassland coexistence.
2
2015
... 相比于一年生草地植物群落, 当代物种共存理论在多年生植物群落中的验证得益于长期观测数据的积累(Adler et al, 2010 ; Chu et al, 2014 ).Chu和Adler (2015)收集了北美5套长期定位观测的草地野外数据, 构建了种群统计学参数模型(包括存活率、生长率和更新率), 然后通过积分投影模型(integral projection model)的手段计算种群入侵增长率, 进而量化生态位差异和平均适合度差异.分析结果同样支持了当代物种共存理论的预测, 同时还发现在这些自然群落中物种间的生态位差异非常大而平均适合度差异相对比较小, 这也就意味着这些多年生草地植物群落受物种间生态位差异的强烈影响, 相对来说非常稳定(Chu & Adler, 2015 ). ...
... 经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退.然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落. ...
Life form influences survivorship patterns for 109 herbaceous perennials from six semi-arid ecosystems.
1
2014
... 相比于一年生草地植物群落, 当代物种共存理论在多年生植物群落中的验证得益于长期观测数据的积累(Adler et al, 2010 ; Chu et al, 2014 ).Chu和Adler (2015)收集了北美5套长期定位观测的草地野外数据, 构建了种群统计学参数模型(包括存活率、生长率和更新率), 然后通过积分投影模型(integral projection model)的手段计算种群入侵增长率, 进而量化生态位差异和平均适合度差异.分析结果同样支持了当代物种共存理论的预测, 同时还发现在这些自然群落中物种间的生态位差异非常大而平均适合度差异相对比较小, 这也就意味着这些多年生草地植物群落受物种间生态位差异的强烈影响, 相对来说非常稳定(Chu & Adler, 2015 ). ...
1
1971
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Animal Ecology. Sidgwick and Jackson
1
1927
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Testing the trait-based community framework: do functional traits predict competitive outcomes?
1
2016
... 在当代物种共存理论中, 物种间竞争的结局有两类: 稳定共存或者竞争排除.而在自然群落中, 要观察到竞争排除或者稳定共存通常都需要相对较长的时间, 这实际上是很难实现的.但任何一个物种种群的增长或消亡最终都要体现在种群的统计学参数上, 包括个体的存活率、生长率和繁殖率, 所以相对于观察稳定共存或竞争排除, 转而关注竞争对种群统计学参数的影响是目前检验当代物种共存理论的替代方法(Harms et al, 2000 ; Funk & Wolf, 2016 ).这是其一. ...
The Struggle of Existence. Williams & Wilkins, Baltimore.
1
1934
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Species coexistence: macroevolutionary relationships and the contingency of historical interactions.
1
2016
... 从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率.目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据.Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数.如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异.结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性.Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异.实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局. ...
A framework for community interactions under climate change.
1
2010
... 温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 ).这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
Phylogenetic relatedness and the determinants of competitive outcomes.
2
2014
... 从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率.目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据.Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数.如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异.结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性.Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异.实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局. ...
... 经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退.然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落. ...
Phenology effects on invasion success: insights from coupling field experiments to coexistence theory.
1
2014
... 从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率.目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据.Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数.如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异.结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性.Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异.实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局. ...
The niche-relationships of the California thrasher.
2
1917
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
... ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Functional equivalence, competitive hierarchy and facilitation determine species coexistence in highly invaded grasslands.
2015
The competitive exclusion principle.
1
1960
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest.
1
2000
... 在当代物种共存理论中, 物种间竞争的结局有两类: 稳定共存或者竞争排除.而在自然群落中, 要观察到竞争排除或者稳定共存通常都需要相对较长的时间, 这实际上是很难实现的.但任何一个物种种群的增长或消亡最终都要体现在种群的统计学参数上, 包括个体的存活率、生长率和繁殖率, 所以相对于观察稳定共存或竞争排除, 转而关注竞争对种群统计学参数的影响是目前检验当代物种共存理论的替代方法(Harms et al, 2000 ; Funk & Wolf, 2016 ).这是其一. ...
Rethinking community assembly through the lens of coexistence theory.
1
2012
... 相对于中性理论假定物种功能等同, 当代物种共存理论将物种之间的差异分为两类: 生态位差异(niche difference, ND)和平均适合度差异(average fitness difference, AFD) (Chesson, 2000 ; 2013 ) (图1 ).生态位差异和平均适合度差异是两个非常抽象的概念, 均不涉及具体的物种差异而是对真实的物种差异的高度概括和抽象.对于生态位差异, 我们可以作如下理解: 深根系物种与浅根系物种通过根系的长短实现对资源的分化利用; 对于平均适合度差异, 类似地我们可以将其设想为物种对相同资源在利用效率上的差异(HilleRisLambers et al, 2012 ). 由此可见, 当代物种共存理论中的生态位差异包括了经典的物种共存理论的精髓, 即生态位分化 (Letten et al, 2017 ).对于稳定的物种共存而言, 生态位差异越大越好, 而平均适合度差异则越小越好.换言之, 生态位差异越大越有利于稳定共存, 而平均适合度差异越大则越有利于竞争排除.对于任何一个现实的群落, 生态位差异和平均适合度差异的相对大小决定了物种是稳定共存还是会发生竞争排除(Adler et al, 2007 ), 这是当代物种共存理论最为核心的内容.当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局.从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 ). ...
1
2001
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
Concluding remarks.
1
1957
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Herbivores and the number of tree species in tropical forests.
1
1970
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Indirect effects of environmental change in resource competition models.
2
2015
... 温度和降水等环境条件既可以通过影响光合作用和呼吸作用等对植物产生直接的影响, 也可以通过影响生物间的相互作用如竞争产生间接的影响(Chu et al, 2016 ).这种因环境变化所导致的间接效应已成为预测气候变化影响的重要的不确定性来源(Gilman et al, 2010 ; Adler et al, 2012 ; Angert et al, 2013 ).那么如何有效地预测气候因子间接影响的强度呢?对该问题的回答能够为判断是否需要考虑气候的间接影响提供依据(Adler et al, 2012 ; Kleinhesselink & Adler, 2015 ).由于这种间接影响是通过种间相互作用来调节的, 因此基于当代物种共存理论, 我们可以做出如下预测: 物种间的生态位差异越大, 物种间的竞争可能越弱, 气候的间接影响就越小; 而物种间的生态位差异越小, 物种间的竞争可能越强, 则气候的间接影响越大.换言之, 可以通过物种间的生态位差异来预测气候间接影响的强度.基于长期的野外观测数据, 结合积分投影模型等手段, Chu等(2016)发现物种间的生态位差异确实与气候间接影响的强度呈现一定的相关关系: 当生态位差异很大的时候, 气候间接影响的强度将变得很小.这也就意味着在物种间生态位差异较大的情况下, 气候对植物群落的直接影响将强于通过物种间相互作用所产生的间接影响, 此时就可以忽略气候的间接影响. ...
... 将当代物种共存理论与具体共存机制相结合是当代物种共存理论发展的必然.而具体物种共存机制正是生态位理论的核心.生态位理论按照生态位的定义大体上可以划分为经典生态位理论和当代生态位理论, 这在本文的第一部分均有介绍.经典生态位理论主要是以现象模型如Lotka-Volterra模型为基础, 侧重物种对环境的需求.当代生态位理论以机理模型如消费者-资源模型为基础, 既包含了物种对环境的需求, 又包含了物种对环境的影响.这里我们侧重当代物种共存理论与当代生态位理论的结合, 而这两个理论正是通过生态位的概念密切联系在一起(Kleinhesselink & Adler, 2015 ; Letten et al, 2017 ).在当代生态位理论中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响.那么这3个因素与当代物种共存理论中的生态位差异(稳定化机制)和平均适合度差异(均等化机制)有什么关系?Letten等(2017)发现资源的供给率体现的是均等化的机制, 物种对环境的影响体现的是稳定化的机制, 而物种对环境的需求可能是均等化机制也可能是稳定化机制.这也就是说当代物种共存理论与当代生态位理论确实是可以有机地结合在一起的; 也只有将二者紧密结合才能最大限度地发挥二者的作用, 从而为更全面地理解物种共存提供基础. ...
Plant functional traits and the multidimensional nature of species coexistence. Proceedings of the National Academy of Sciences,
2
2015
... 从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率.目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据.Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数.如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异.结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性.Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异.实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局. ...
... 当代物种共存理论为解决这个问题提供了最直接和最有效的方法.在当代物种共存理论中, 物种间的差异分为生态位差异和平均适合度差异, 而这两类差异真实地代表了物种共存的两类驱动力.我们可以将这两类差异视为抽象的性状.如果能够将这两类抽象性状与具体性状如根系深浅结合在一起(谱系距离可依此类推), 找到二者之间的联系, 就能有效地揭示内在的群落构建机制(Adler et al, 2013 ; Kraft et al, 2015 ).比如, 如果平均适合度差异与谱系距离差异呈现负相关关系, 那么就意味着亲缘关系越近的物种其适合度差异越大, 而平均适合度差异越大物种间竞争越强, 越不容易稳定共存.目前已有一些工作报道了相关的研究成果.以一年生草本植物为研究对象, Kraft等(2015)发现在其选择的11个功能性状中, 大部分功能性状与平均适合度差异之间的相关性很强, 而与生态位差异的相关性不明显, 这意味着这些功能性状主要体现的是物种之间的适合度差异.以绿藻群落为研究对象, Narwani等(2013)发现物种间的谱系距离与生态位差异和平均适合度差异均无明显相关关系, 与物种是否共存也不相关.这表明基于谱系距离来推断竞争的重要性并不合适, 即上述的“亲缘关系越近竞争越强, 亲缘关系越远竞争越弱”的假定可能是不成立的.Germain等(2016)也发现谱系距离与生态位差异和平均适合度差异并无一一对应的关系. ...
Competitive interactions between forest trees are driven by species’ trait hierarchy, not phylogenetic or functional similarity: implications for forest community assembly.
2
2012
... 同时, 从理论上来说, 量化生态位差异和平均适合度差异需要测量竞争系数或者种群的入侵增长率.而在复杂的自然系统中, 这一点同样很难实现.为此我们同样需要寻找一个替代的方案.对于功能性状而言, 假设有两个物种A和B, 其性状值分别为TA 和TB .人们提出了用物种间绝对性状距离(性状间差异的绝对值, |TA -TB |)来代表生态位差异, 而用物种间等级性状距离(性状间差异, TA -TB )来代表平均适合度差异.换言之, 绝对性状距离是没有方向的, 只要存在差异就有利于物种的共存, 体现的是物种间生态位的分化; 而等级性状距离具有方向, 体现的是物种间适合度的差异(Kunstler et al, 2012 ).这是其二. ...
... 通过上述两个方面的替代, 就可以对当代物种共存理论开展行之有效的验证.利用法国全国林业调查的数据, Kunstler等(2012) 发现物种间的竞争强度主要受性状等级(等级性状距离)所驱动, 而与性状或谱系的相似性(性状相似性即绝对性状距离)没有明显的相关关系.Gross等(2015)在新西兰开展的草地实验表明, 性状间的差异主要体现的是竞争的等级, 即在该实验中所选取的性状如植株高度和特定叶面积指数代表的主要是物种间的平均适合度差异. ...
Are there general laws in ecology?
1
1999
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
The niche concept revisited: mechanistic models and community context.
1
1995
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Linking modern coexistence theory and contemporary niche theory.
2
2017
... 相对于中性理论假定物种功能等同, 当代物种共存理论将物种之间的差异分为两类: 生态位差异(niche difference, ND)和平均适合度差异(average fitness difference, AFD) (Chesson, 2000 ; 2013 ) (图1 ).生态位差异和平均适合度差异是两个非常抽象的概念, 均不涉及具体的物种差异而是对真实的物种差异的高度概括和抽象.对于生态位差异, 我们可以作如下理解: 深根系物种与浅根系物种通过根系的长短实现对资源的分化利用; 对于平均适合度差异, 类似地我们可以将其设想为物种对相同资源在利用效率上的差异(HilleRisLambers et al, 2012 ). 由此可见, 当代物种共存理论中的生态位差异包括了经典的物种共存理论的精髓, 即生态位分化 (Letten et al, 2017 ).对于稳定的物种共存而言, 生态位差异越大越好, 而平均适合度差异则越小越好.换言之, 生态位差异越大越有利于稳定共存, 而平均适合度差异越大则越有利于竞争排除.对于任何一个现实的群落, 生态位差异和平均适合度差异的相对大小决定了物种是稳定共存还是会发生竞争排除(Adler et al, 2007 ), 这是当代物种共存理论最为核心的内容.当物种既无生态位差异也无适合度差异的时候, 群落即为中性格局.从这个意义上来讲, 中性理论和当代物种共存理论本质上是不冲突的, 前者只是后者的一个特例(Adler et al, 2007 ). ...
... 将当代物种共存理论与具体共存机制相结合是当代物种共存理论发展的必然.而具体物种共存机制正是生态位理论的核心.生态位理论按照生态位的定义大体上可以划分为经典生态位理论和当代生态位理论, 这在本文的第一部分均有介绍.经典生态位理论主要是以现象模型如Lotka-Volterra模型为基础, 侧重物种对环境的需求.当代生态位理论以机理模型如消费者-资源模型为基础, 既包含了物种对环境的需求, 又包含了物种对环境的影响.这里我们侧重当代物种共存理论与当代生态位理论的结合, 而这两个理论正是通过生态位的概念密切联系在一起(Kleinhesselink & Adler, 2015 ; Letten et al, 2017 ).在当代生态位理论中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响.那么这3个因素与当代物种共存理论中的生态位差异(稳定化机制)和平均适合度差异(均等化机制)有什么关系?Letten等(2017)发现资源的供给率体现的是均等化的机制, 物种对环境的影响体现的是稳定化的机制, 而物种对环境的需求可能是均等化机制也可能是稳定化机制.这也就是说当代物种共存理论与当代生态位理论确实是可以有机地结合在一起的; 也只有将二者紧密结合才能最大限度地发挥二者的作用, 从而为更全面地理解物种共存提供基础. ...
The importance of niches for the maintenance of species diversity.
1
2009
... 从2.2节介绍的两类量化生态位差异和平均适合度差异的方法可以看出, 验证的关键是获取种内和种间竞争系数或者种群的内禀和入侵增长率.目前针对当代物种共存理论的实验验证主要来自一年生草地植物群落(Levine & HilleRisLambers, 2009 ; Godoy et al, 2014 ; Godoy & Levine, 2014 ; Kraft et al, 2015 ; Germain et al, 2016 ), 这主要是因为一年生植物群落的实验周期短, 易于获取种群增长和竞争系数的相关数据.Levine和HilleRisLambers (2009)通过在美国加利福尼亚州的野外控制实验将理论模型参数化, 然后在理论模型中设置种内竞争系数等于种间竞争系数.如上所述, 稳定共存需要满足种内竞争系数大于种间竞争系数, 所以这种处理相当于去除了物种间生态位的差异.结果表明, 去除生态位差异后, 能共存的物种数目明显下降, 这进一步说明了生态位差异对于物种共存的重要性.Kraft等(2015)利用类似的方法, 通过物种单播和两两混种的方式获得了种内竞争系数和种间竞争系数, 进而量化了物种间的生态位差异和平均适合度差异.实验结果支持了当代物种共存理论的预测: 生态位差异和平均适合度差异共同决定了物种竞争的结局. ...
Santa Rosalia was a goat.
1
1983
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
Elements of Physical Biology. Williams & Wilkins Company, Baltimore.
2
1925
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Population ecology of some warblers of northeastern coniferous forests.
2
1958
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
1
1969
... 在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求.在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化.MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
2
1972
... 在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求.在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化.MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
... 另外一条主线以机理模型为基础, 主要是MacArthur提出的消费者-资源模型(MacArthur, 1972 ).机理模型相对现象模型的优势在于同时考虑了物种对环境的“需求”和“影响”, 这也是生态位概念的一次完善, 是Grinnell生态位概念和Elton生态位概念的综合.以机理模型为基础解释物种共存的工作始于MacArthur (1972), 而让该模型人所共知的是Tilman (1980, 1982), 比如其提出的R* 理论和资源比例假说便是生态学发展史上的一个重要节点.在机理模型中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响.换言之, 物种受不同资源的限制, 且该物种消耗最多的资源恰好是限制其增长的那个资源.理论上, MacArthur的消费者-资源模型可以解释无数物种在群落内的共存问题(Tilman, 1982 ).然而, 这种理论上的完美性伴随的是其在实际群落中较低的可操作性, 较难进行实验验证. ...
The limiting similarity, convergence, and divergence of coexisting species.
2
1967
... 在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求.在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化.MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
... ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
Niche overlap as a function of environmental variability. Proceedings of the National Academy of Science,
1
1972
... 在Grinnell (1917)生态位概念的影响下, Hutchinson (1957)提出了超体积生态位的概念, 即物种的适合度是由多个因素所共同决定的, 相对较为定量地描述了物种对外界环境的需求.在这个框架下, Hutchinson试图寻找影响共存物种间相似性的因素, 其中具有重要影响的工作是关于动物体型大小的Hutchinson比例: 当两个相似的物种共存时, 较大的物种体型是较小物种的1.3倍: 竞争使得物种个体大小发生了变化.MacArthur等在Hutchinson生态位概念的基础上开展了一系列时至今日仍对生态学有重要影响的研究工作(MacArthur & Levins, 1967 ; MacArthur, 1969 , 1972 ; May & MacArthur, 1972 ), 也促使当时的生态学家开始测量和计算物种的生态位宽度、生态位重叠程度等.极限相似性假说即为一例, 其假定能够稳定共存的两个物种之间的相似性程度存在一个上限, 超过此上限, 物种间即发生竞争排除从而不能稳定共存(MacArthur & Levins, 1967 ).换言之, 共存物种之间的生态位不能无限重叠.从Grinnell (1917)到Hutchinson (1957)再到MacArthur (1969), 都侧重于物种对环境的需求, 这是经典物种共存理论发展的其中一条主线, 其本质主要是以Lotka-Volterra竞争模型为基础.Lotka- Volterra竞争模型是现象模型, 以竞争系数这个抽象的指数来笼统代表具体机制对物种共存的影响(Chase & Leibold, 2003 ). ...
Opposing effects of competitive exclusion on the phylogenetic structure of communities.
1
2010
... 在竞争能力上可能差异较大), 则会使群落谱系呈现聚集的格局.换言之, 竞争和环境过滤都有可能产生聚集式的格局, 上述“一一对应”的关系并非是普遍存在的(Mayfield & Levine, 2010 ). ...
Experimental evidence that evolutionary relatedness does not affect the ecological mechanisms of coexistence in freshwater green algae.
2
2013
... 此外, 还有研究者采用其他更易操作的实验系统, 比如微生物系统(Zhao et al, 2016 )和藻类系统(Narwani et al, 2013 )来验证当代物种共存理论.所有这些研究都从实验的角度支持了当代物种共存理论的预期, 也为当代物种共存理论的传播及其被认可提供了基础. ...
... 经过近20年的发展, 当代物种共存理论本身已相对成熟, 这主要体现在两个方面: 一是无论是基于现象模型(如Lotka-Volterra模型)还是机理模型(如消费者-资源模型), 量化物种间的生态位差异和平均适合度差异在理论上和实践中都是可行的(Carroll et al, 2011 ; Chesson, 2013 ; Narwani et al, 2013 ; Godoy et al, 2014 ; Chu & Adler, 2015 ); 二是越来越多的群落生态学家对当代物种共存理论的接受和认可部分源于中性理论的影响逐渐消退.然而, 正如我们前面所提到的, 无论是测量竞争系数还是种群入侵增长率, 在实际操作中都存在着很大的困难, 这也是为什么到目前为止针对该理论的实验验证都主要集中在相对简单的系统, 如一年生草地植物群落、微生物群落和藻类群落. ...
Competition and diversity in herbaceous vegetation.
1
1973
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究
1
2011
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究
1
2011
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
群落构建的中性理论和生态位理论
2
2009
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
群落构建的中性理论和生态位理论
2
2009
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
Do fitness-equalizing tradeoffs lead to neutral communities?
1
2012
... 均等化机制是通过降低物种之间的适合度差异或者物种在竞争能力方面的差异来促进共存的.以对资源的竞争为例, 物种对资源的竞争能力可以通过R *值来表示, 即某物种在群落中能够维持下去所需的最低的资源水平(Tilman, 1982 ).当多个物种同时利用相同的限制性资源时, 具有最低R *值的物种将竞争排除掉所有其他的物种.因此, 任何能减少物种之间R *值差异的因子都可被归为均等化的机制, 比如对竞争优势种的部分去除等.不同于稳定化机制, 均等化机制不会导致负密度制约.经典物种共存理论的工作几乎都是以稳定化机制为研究对象的, 强调物种之间的生态位分化, 却很少关注均等化机制.实际上, 均等化机制的研究是在群落中性理论提出后才得到了较为快速的发展的(Chave, 2004 ; Turnbull et al, 2008 ; Ostling, 2012 ). ...
On the evidence for species coexistence: a critique of the coexistence program.
1
2010
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Santa Rosalia reconsidered: size ratios and competition.
1
1981
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
Grinnellian and Eltonian niches and geographic distributions of species.
1
2007
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
Tests of community-wide character displacement against null hypotheses.
1
1979
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
Resource: a graphical-mechanistic approach to competition and predation.
1980
4
1982
... 另外一条主线以机理模型为基础, 主要是MacArthur提出的消费者-资源模型(MacArthur, 1972 ).机理模型相对现象模型的优势在于同时考虑了物种对环境的“需求”和“影响”, 这也是生态位概念的一次完善, 是Grinnell生态位概念和Elton生态位概念的综合.以机理模型为基础解释物种共存的工作始于MacArthur (1972), 而让该模型人所共知的是Tilman (1980, 1982), 比如其提出的R* 理论和资源比例假说便是生态学发展史上的一个重要节点.在机理模型中, 物种能否共存取决于3个方面: 资源的供给率、物种对环境的需求以及物种对环境的影响.换言之, 物种受不同资源的限制, 且该物种消耗最多的资源恰好是限制其增长的那个资源.理论上, MacArthur的消费者-资源模型可以解释无数物种在群落内的共存问题(Tilman, 1982 ).然而, 这种理论上的完美性伴随的是其在实际群落中较低的可操作性, 较难进行实验验证. ...
... 无论是基于现象模型还是基于机理模型的物种共存理论都为生态位分化和竞争在物种共存中的作用提供了一个相对综合性的理论框架.然而, 在20世纪80-90年代, 这两类模型都遭到了强烈的质疑(Strong et al, 1979 ; Simberloff & Boecklen, 1981 ; Lewin, 1983 ).研究发现, 很多格局, 如Hutchinson比例, 实际上并不具有统计上的显著性, 这意味着生态位分化或者竞争对物种共存的作用并非像之前假定的那么重要.因此, 人们转而倾向于认为物种共存机制依赖于具体的群落类型.回顾群落生态学的发展, 人们的确发现不同群落类型物种共存的具体机制是不同的.对一些群落而言, 土壤的营养元素含量和比例很重要(Tilman, 1982 ), 而在另外一些群落中物种对光照的竞争则起着主要作用(Newman, 1973 ).此外, 物种的空间生态位的分离(MacArthur, 1958 )和时间生态位的分化(Chesson, 1985 )是维持某些群落物种共存的关键.同时, 对于某个具体的群落而言, 物种共存与否是多个因素共同作用的结果, 包括因素的独立影响、不同因素之间的交互影响以及这些因素时空变异的作用.这一度让一些生态学家陷入了绝望的境地: 关于物种共存机制还存在普适性的理论框架吗(Lawton, 1999 )? ...
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
... 均等化机制是通过降低物种之间的适合度差异或者物种在竞争能力方面的差异来促进共存的.以对资源的竞争为例, 物种对资源的竞争能力可以通过R *值来表示, 即某物种在群落中能够维持下去所需的最低的资源水平(Tilman, 1982 ).当多个物种同时利用相同的限制性资源时, 具有最低R *值的物种将竞争排除掉所有其他的物种.因此, 任何能减少物种之间R *值差异的因子都可被归为均等化的机制, 比如对竞争优势种的部分去除等.不同于稳定化机制, 均等化机制不会导致负密度制约.经典物种共存理论的工作几乎都是以稳定化机制为研究对象的, 强调物种之间的生态位分化, 却很少关注均等化机制.实际上, 均等化机制的研究是在群落中性理论提出后才得到了较为快速的发展的(Chave, 2004 ; Turnbull et al, 2008 ; Ostling, 2012 ). ...
Why equalising trade-offs aren’t always neutral?
1
2008
... 均等化机制是通过降低物种之间的适合度差异或者物种在竞争能力方面的差异来促进共存的.以对资源的竞争为例, 物种对资源的竞争能力可以通过R *值来表示, 即某物种在群落中能够维持下去所需的最低的资源水平(Tilman, 1982 ).当多个物种同时利用相同的限制性资源时, 具有最低R *值的物种将竞争排除掉所有其他的物种.因此, 任何能减少物种之间R *值差异的因子都可被归为均等化的机制, 比如对竞争优势种的部分去除等.不同于稳定化机制, 均等化机制不会导致负密度制约.经典物种共存理论的工作几乎都是以稳定化机制为研究对象的, 强调物种之间的生态位分化, 却很少关注均等化机制.实际上, 均等化机制的研究是在群落中性理论提出后才得到了较为快速的发展的(Chave, 2004 ; Turnbull et al, 2008 ; Ostling, 2012 ). ...
Conceptual synthesis in community ecology.
2010
2
1926
... 经典物种共存理论强调具体的物种共存机制, 比如植物物种对土壤资源的分化利用、物种的时间生态位分化和空间生态位分化等, 其历史可追溯到生态位概念的提出(Grinnell, 1917 ).Grinnell的生态位概念侧重物种对外界环境条件的“需求” (需求生态位, requirement niche), 而Elton (1927)的生态位概念则强调物种在群落中的作用和对环境的“影响” (影响生态位, impact niche).尽管人们对生态位概念本身的认识存在诸多分歧(Grinnell, 1917 ; Elton, 1927 ; Gause, 1934 ; Hutchinson, 1957 ; Leibold, 1995 ; Chase & Leibold, 2003 ; Soberón, 2007 ), 但是生态位分化对于物种共存的重要性是普遍认可的.Lotka- Volterra竞争模型在理论上表明了种内竞争与种间竞争的相对强度决定了物种是稳定共存还是发生竞争排除(Lotka, 1925 ; Volterra, 1926 ).俄国生物学家Gause (1934)通过草履虫实验验证了Lotka-Volterra竞争模型的预测结果, 这意味着物种对资源的分化利用是维持物种共存的必要条件, 即“竞争排除法则” (Hardin, 1960 ). ...
... 种群增长的负密度制约或者稀有种优势是稳定化机制的体现: 在一个稳定的群落中, 如果任何一个物种变得特别稀少, 那么该物种的种群数量将会表现出增加的趋势(Siepielski & McPeek, 2010 ).在Lotka-Volterra竞争模型中, 种内竞争强于种间竞争是生态位分化和稳定化机制的具体体现; 当且仅当种内竞争大于种间竞争时, 物种才会实现稳定的共存(Lotka, 1925 ; Volterra, 1926 ).稳定化机制涵盖了大部分经典的有关物种共存的假设和理论, 包括物种间的时间生态位分化(Chesson, 1994 )、空间生态位分化(MacArthur, 1958 )、资源分化(Tilman, 1982 )和物种特异性天敌(Janzen, 1970 ; Connell, 1971 )等. ...
Phylogenies and community ecology.
1
2002
... 群落生态学的一项重要任务是从群落格局推断内在的群落构建机制.一是基于功能性状的分布格局来探讨环境过滤和竞争在群落构建过程中的相对作用: 如果共存物种的性状分布相对于零模型来说表现为聚集的格局, 那么推断环境过滤是群落构建的主要驱动力; 反之, 如果性状分布表现为发散的格局, 那么物种之间的竞争则起着主导作用(Ackerly & Cornwell, 2007 ).二是基于亲缘关系的谱系群落生态学, 与基于功能性状的方法类似: 如果生境过滤作用占主导地位, 那么相似生境将筛选出适应能力相似、亲缘关系较近的物种, 从而表现为谱系的聚集; 相反, 竞争排斥会使得生态位相似的物种无法共存于同一环境, 群落内物种的亲缘关系则较远, 表现为谱系的发散(Webb et al, 2002 ).这种从格局推断过程的方法在2002年Webb等发表其综述性论文后被普遍采纳.同时, 在谱系群落生态学中有一个非常常见的重要前提假定, 即亲缘关系越近竞争越强, 亲缘关系越远竞争越弱. ...
Evolution alters ecological mechanisms of coexistence in experimental microcosms.
1
2016
... 此外, 还有研究者采用其他更易操作的实验系统, 比如微生物系统(Zhao et al, 2016 )和藻类系统(Narwani et al, 2013 )来验证当代物种共存理论.所有这些研究都从实验的角度支持了当代物种共存理论的预期, 也为当代物种共存理论的传播及其被认可提供了基础. ...
群落生态学的中性理论
2
2006
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
群落生态学的中性理论
2
2006
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
... 当代物种共存理论忽略了具体的共存机制, 转而关注影响物种共存的一般性规律.伴随着上述争论, 关于物种共存发展出两类相对独立的理论框架.一是Chesson提出的关于物种共存的综合性理论框架, 也就是大家现在俗称的“当代物种共存理论” (Chesson, 2000 , 2013 ); 二是群落中性理论(Bell, 2001 ; Hubbell, 2001 ), 它忽略物种间的差异或者假定物种间差异对于物种共存没有重要影响.尽管中性理论有很好的拟合观测数据的能力, 但是其关键的前提假定与人们的常识相悖.因已有多篇综述性论文对中性理论进行过详细的介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ), 所以本文不再赘述.下面, 我们单就Chesson的当代物种共存理论作出详尽说明. ...
基于过程的群落生态学理论框架
1
2011
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
基于过程的群落生态学理论框架
1
2011
... 解释多物种如何在现实群落中的共存是群落生态学和生物多样性研究的核心内容.自2006年以来, 国内一些有关群落构建和生物多样性维持机制的综述对此有所介绍(周淑荣和张大勇, 2006 ; 牛克昌等, 2009 ; 牛红玉等, 2011 ; 朱璧如和张大勇, 2011 ; 陈磊等, 2014 ).周淑荣和张大勇(2006)介绍了群落中性理论的前提假设、预测能力和物种形成等方面的研究进展, 分析了扩散限制条件下群落中性构建中物种共存的可能机制; 牛克昌等(2009)在回顾生态位理论和群落中性理论发展的基础上, 分析了二者之间的主要分歧和互补性, 梳理了二者整合的可能途径, 提出群落的多物种共存可能是随机过程和确定性作用的共同结果, 而针对具体群落需要深入研究二者的相对贡献; 牛红玉等(2011)综述了群落谱系生态学的相关理论和研究进展, 讨论了如何理解长期进化过程对群落中多物种共存的影响; 朱璧如和张大勇(2011)对Vellend (2010)提出的基于过程的群落构建概念框架进行了介绍, 为国内工作者理解格局与过程之间的关系提供了新视角; 陈磊等(2014)介绍了生态位分化在物种多样性维持过程中的作用, 提出在将来的研究中需要考虑个体分化对局域群落构建的影响.这些综述性论文为国内从事群落生态学和生物多样性研究的工作者理解群落构建和生物多样性的维持机制起到了积极的推动作用.然而, 以上综述均没有专门介绍当代物种共存理论这一近十年来群落生态学发展最为活跃的主题. ...
 , 王酉石
, 王酉石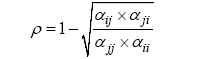
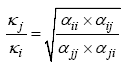
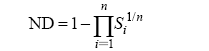
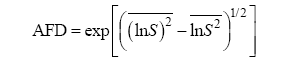
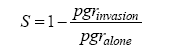


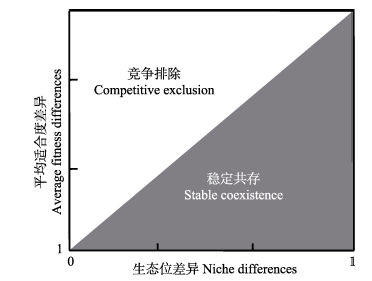
{kind=link}
{kind=link}