植物区系(flora)是某一特定地区生长着的全部植物种类, 是植物科属种的自然综合体(吴征镒等, 2006 )。植物区系研究在植物种类清查、生物资源开发、环境保护、国民经济建设等方面起着至关重要的作用(吴征镒等, 2006 )。我国植物区系地理学研究主要集中于对一个自然地理区域内所有植物的地理成分从科、属、种三个层次进行分析, 并通过与邻近地区的植物区系比较, 确定该地区现代植物区系的性质、来源和地位, 如吴征镒等(2010)关于我国种子植物区系的研究, 表明中国种子植物区系的亚热带性质及热带亲缘等特点。
然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 )。除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 )。因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 )。早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用。而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度。系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 )。
Swenson和Umaña (2014)在研究加勒比海小安第列斯群岛的植物区系时, 首次提出了植物系统发育区系学(phylofloristics)的概念, 即应用系统发育的方法对现代植物区系进行研究。但在此之前, 植物系统发育区系学已被用于全球不同地区的研究, 如: Forest等(2007)对南非好望角植物区系的研究, Kooyman等(2011)对澳洲不同森林类型的植物区系研究和Qian等(2013a , b ; 2015 )对北美被子植物区系的研究。这些研究均表明结合进化历史的植物区系研究对理解植物区系的形成起着至关重要的作用。
我国有很好的植物区系研究基础, 但从进化历史的角度研究植物区系才刚刚起步。当前研究主要以大型森林动态样地为平台或以木本植物为主, 探讨不同生态过程对不同空间尺度树种系统发育组成的影响(如: Wang et al, 2012 ; Qiao et al, 2015 ; Qian et al, 2016 ), 但这些研究忽视了反映林下小生境对植物区系形成起重要作用的草本植物的影响, 而目前基于特定地区所有植物种类的植物系统发育区系地理学研究仅有零星报道(Li et al, 2014 , 2015a, 2015b )。我国幅员辽阔, 不同地区的地质历史与现代自然环境千差万别, 植物区系的形成原因大相径庭, 因此, 在我国开展植物系统发育区系地理学研究, 对深入理解中国植物区系的成因具有重要意义。
云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 )。在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 )。近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论。
鉴于此, 本文在云南植物区系分区研究的基础上(Li et al, 2015a ), 从进化历史的角度分析不同地理单元的分类群组成、系统发育组成及其相似性, 探讨各个地理单元的系统发育结构及地理单元间的系统发育相似程度, 以期更全面地了解云南现代植物区系的性质和来源。
1 方法
1.1 研究地区
云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份。自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 )。因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 )。正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 )。
1.2 数据来源
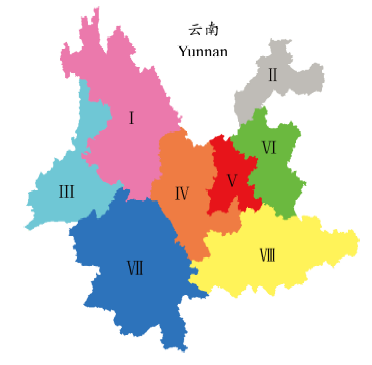
基于云南植物区系分区研究(Li et al, 2015a )划分的8个地理单元(图1 ), 以《云南植物志》为基础(吴征镒, 1977-2006), 构建不同地理单元种子植物的属级“有/无”数据矩阵。区系组成中的外来成分或栽培种类不列入统计。科属范畴的界定和划分依据被子植物系统发育分类系统(Angiosperm Phylogeny Group Classification, APG)的观点(APG III, 2009 )。本研究共计包括8,072条数据记录。
图1 基于云南植物区系分区研究划分的8个地理单元(引自Li et al, 2015a )
Fig. 1 The delineation of eight floristic regions in Yunnan, China (adopted from Li et al, 2015a )
1.3 系统发育树构建
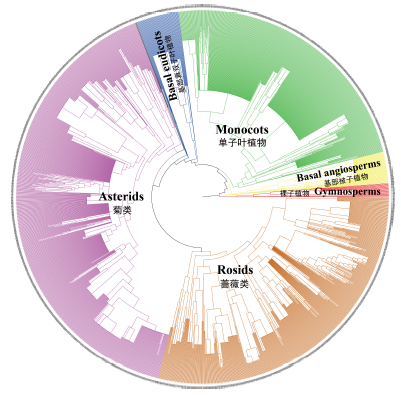
将云南植物区系的所有种子植物属作为物种库, 以种子植物系统发育关系(Zanne et al, 2014 )为骨架, 利用Phylomatic软件构建基于全部属的系统发育树(Webb & Donoghue, 2005 )。对于科内无法解决的属间关系, 则以多歧分支(polytomy)的方式进行处理(Hardy et al, 2012 ) (图2 )。
图2 云南植物区系的宏系统发育树
Fig. 2 The mega-phylogeny of seed plants in Yunnan
1.4 分类群组成与系统发育组成的多样性测量
采用科或属的丰富度(richness)与Faith (1992)的系统发育多样性(phylogenetic diversity, PD)来量化不同地理单元的分类群组成与系统发育组成多样性。丰富度以各个地理单元内科或属的绝对数量来衡量。系统发育多样性为地理单元内所有分类群在有根系统发育树上的支长总和。使用线性回归模型检测丰富度与系统发育多样性之间的相关性。
采用净相关指数(net relatedness index, NRI)度量不同地理单元的系统发育结构(Webb, 2000 ), 其计算公式如下:
(1)
式中, MPDobserved 是观察地理单元分类群间的平均谱系距离(mean phylogenetic distance, MPD), MPDrandomized 表示随机过程产生的零地理单元分类群间的平均谱系距离, sdMPDrandomized 表示零地理单元分类群间平均谱系距离的方差。如果NRI值为正, 则观察地理单元的MPD值小于通过零地理单元计算的MPD值, 表示植物区系的系统发育结构为聚集型(phylogenetic clustering); 反之, 如果NRI值为负, 则MPD值大于通过零地理单元计算的期望值, 表示系统发育结构为离散型(phylogenetic overdispersion)。
1.5 分类群组成与系统发育组成的相似性测度
采用Sørenson分类群相似性指数(Sørenson, 1948 )及PhyloSor系统发育相似性指数(Bryant et al, 2008 )测度云南植物区系不同地理单元的分类群组成与系统发育组成的相似程度, 其计算公式如下:
(2)
(3)
式中, Sij 是i 、j 两个地理单元共有的分类群数目, Si 与Sj 是i 、j 两个地理单元各自的分类群数目; 类似地, BLij 是i 、j 两个地理单元共有的支长之和, BLi 与BLj 是i 、j 两个地理单元各自的支长总和。Sørenson与PhyloSor的值在0与1之间。如为0, 则表示植物区系完全不相同; 如为1, 则表示植物区系完全相同。运用Mantel检验分析分类群组成相似性与系统发育组成相似性之间的相关性(Legendre & Fortin, 2010 )。
为了直观显示不同地理单元之间的区系组成相似程度, 采用Ward的方法对分类群组成及系统发育组成相似性系数进行等级聚类分析(Kreft & Jetz, 2010 )。
上述所有分析均采用R语言(R Core Team, 2014 )的vegan (Oksanen et al, 2015 )、picante (Kembel et al, 2010 )、cluster (Maechler et al, 2015 )等程序包计算。
2 结果
2.1 分类群多样性与系统发育多样性
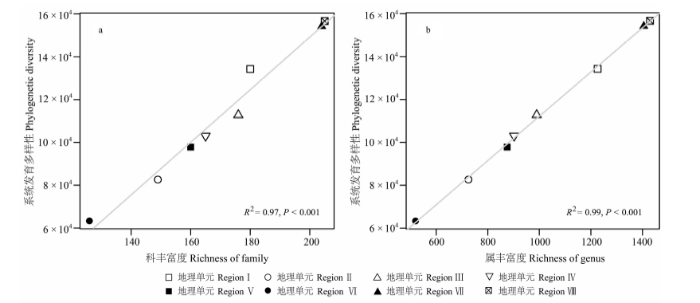
基于被子植物系统发育分类系统(APG III, 2009 )的统计, 云南种子植物区系计有225个科1,983个属。8个地理单元的科、属组成及系统发育多样性见表1 。地理单元VIII具有最高的科、属丰富度及系统发育组成多样性, 地理单元VI具有最低的科、属丰富度及系统发育多样性。不同地理单元植物区系的系统发育多样性与科(R2 = 0.97, P < 0.001)或属(R2 = 0.99, P < 0.001)的丰富度呈显著正相关(图3 )。
图3 云南植物区系不同地理单元系统发育多样性与科丰富度(a)和属丰富度(b)的关系(I-VIII代表的地理单元同图1 )
Fig. 3 Relationships between phylogenetic diversity and family richness (a) or genus richness (b) among the floristic regions in Yunnan. I-VIII indicate different floristic regions as in Fig. 1 .
2.2 系统发育结构
云南种子植物区系在不同地理单元的系统发育结构不一致(表2 ), 地理单元III的系统发育结构为离散型, 其余地理单元均为聚集型。
2.3 分类群组成与系统发育组成相似性
云南种子植物区系不同地理单元间的系统发育组成相似性与分类群组成相似性呈显著正相关(R2 = 0.99, P = 0.001)。不同地理单元两两间(n = 28)的平均分类群组成相似性(Sørenson指数)为0.665, 地理单元VII与VIII的分类群组成最为相似(0.822), 地理单元VII与VI的分类群组成差异最大(0.476)。不同地理单元两两间(n = 28)的平均系统发育组成相似性(PhyloSor指数)为0.698, 地理单元VII与VIII的系统发育组成最为相似(0.836), 地理单元VII与VI的系统发育组成差异最大(0.526), 系统发育组成平均相似性比分类群组成平均相似性高3.3%。
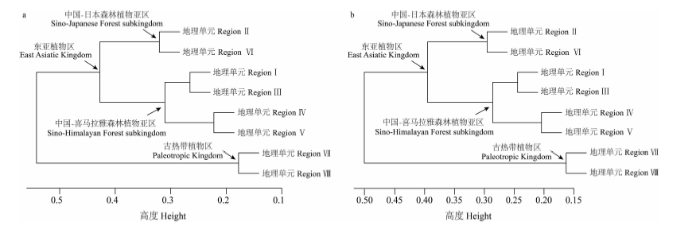
分类群组成与系统发育组成的相似性聚类分析(图4 )也显示了云南种子植物区系不同地理单元间分类群组成与系统发育组成的相关性。不论是分类群组成聚类分析还是系统发育组成聚类分析, 地理单元VII与VIII均聚为一类(南部区), 其他6个地理单元聚为另一类(北部区); 在北部区内, 地理单元II与VI聚为一支, 地理单元I、III、IV及V聚为另一支。
图4 云南植物区系不同地理单元间分类学组成(a)与系统发育组成(b)的相似性聚类
Fig. 4 A cluster dendrogram representing the taxonomic floristic similarity (a) and the phylofloristic similarity (b) between the geographic regions of Yunnan. I-VIII indicate different floristic regions as in Fig. 1 .
3 讨论
3.1 现代植物区系的进化历史
云南植物种类丰富, 早期根据科属分布区类型的分析表明, 云南植物区系地理成分复杂、联系广泛(李锡文, 1985 ; 吴征镒, 1987 ), 该结论得到本研究的支持。从云南种子植物所含1,983个属所展示的进化关系来看(图2 ), 这些属在系统发育树上的各个主要分支均有分布(如: 祼子植物、基部被子植物、单子叶植物、基部真双子叶植物、蔷薇类、菊类等), 表明云南植物区系在地质历史的不同时期与其他地区的植物区系有过广泛的联系和交流, 并在适应当地的自然环境下产生了大量的后裔, 逐渐演变为当今植物区系的外貌。
整体来说, 云南植物区系的分类群多样性(科或属的丰富度)与系统发育多样性显著相关(图3 ), 然而, 使用零模型(null model)保持植物区系分类群组成不变的情况下, 随机产生的零地理单元系统发育组成与观察地理单元系统发育组成的比较发现, 分类群多样性或系统发育多样性丰富的区域并不一定具有多样的系统发育组成, 即云南植物区系显示了非随机的系统发育结构, 不同地理单元植物区系的进化组成不一致(表2 )。8个地理单元中, 有7个地理单元的系统发育结构为聚集型, 表明组成这7个地理单元的分类群(科或属)在系统发育树上的位置相对较为聚集或者这些分类群主要来自相同的进化谱系(lineages), 如构成地理单元I的高山灌丛和高山草甸植被主要是由物种多样性高的杜鹃花科、报春花科及马先蒿属(Pedicularis )、龙胆属(Gentiana )、风毛菊属(Saussurea )等类群组成, 这些科属均来源于进化树上的菊类谱系; 而地理单元III是离散型的系统发育结构, 即这个地理单元植物区系的进化谱系组成较为多样, 组成这个地理单元的分类群来自系统发育树上的更多谱系, 如构成地理单元III的常绿阔叶林建群种主要是樟科、壳斗科及山茶科等类群的物种, 这些科则分别来自于进化树上的基部被子植物、蔷薇类及菊类等谱系。
云南植物区系非随机的系统发育结构与其进化历史有关。自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 )。地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成。与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低。
3.2 现代植物区系不同地理单元间的相似性
整体来看, 云南植物区系不同地理单元间的系统发育组成相似性与分类群组成相似性显著相关, 且前者比后者高, 表明云南植物区系不同地理单元间的分类群组成差异较系统发育组成差异大, 即不同地理单元的分类群组成主要由进化树上相同谱系内的不同类群构成, 如组成云南植物区系主体的4个大科禾本科(158属)、兰科(134属)、菊科(117属)及豆科(105属), 从科一级来看, 在云南广泛分布, 但从科内属一级来看, 则不同的属在云南有不同的地理分布, 这就导致不同地理单元间系统发育组成相似性比分类群组成相似性高, 这从另一侧面也说明云南多样且独特的自然环境是生物分化和演化的舞台。
云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 )。在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持。南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 )。北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 )。北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分。新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 )。
3.3 问题与不足
植物系统发育区系地理学研究在我国刚刚起步, 本文以云南植物区系为例, 从进化历史的角度探讨不同地理单元的系统发育结构及它们之间的相似度, 与仅基于分类群的区系分析相比, 系统发育的区系分析能更深入地理解植物区系的性质和来源。然而, 本研究中属间进化关系未解决及属的地理分布以县为单位限制了对植物区系的起源时间、演化、特有现象及区系分区等问题的深入了解, 克服这些瓶颈则成为将来植物系统发育区系地理学的重要方向。第一, 利用DNA条形码或基因组数据, 重建高精度区域性宏系统发育树(mega- phylogeny), 将加深对植物区系的起源及进化历史的理解; 第二, 基于标本馆馆藏标本数据, 构建精准的地区性植物地理分布网络, 结合地理信息系统(GIS)和系统发育的分析, 有助于深入认识植物区系特有现象及区系分区的本质; 第三, 综合进化、生态及地理分布等信息的全面分析, 有助于阐明生态过程与进化历史在植物区系形成中的作用。同时, 相关研究对生物多样性保护也具有重要的参考价值。
(责任编委: 葛学军 责任编辑: 时意专)
致谢: 感谢中国科学院昆明植物研究所信息中心的王雨华博士、李拓径博士提供本研究所需的基础数据。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[8]
Ge XJ 2015 ) Application of DNA barcoding in phylofloristics study.
Biodiversity Science , 23 , 295 -296 .
[本文引用: 2]
(in Chinese) [葛学军 (2015 ) DNA条形码在植物系统发育区系学研究中的应用. 生物多样性,
23 , 295 -296 .]
[本文引用: 2]
[9]
Hardy OJ Couteron P Munoz F Ramesh BR Pélissier R 2012 ) Phylogenetic turnover in tropical tree communities: impact of environmental filtering, biogeography and mesoclimatic niche conservatism.
Global Ecology and Biogeography , 21 , 1007 -1016 .
[本文引用: 1]
[10]
Harrison TM Copeland P Kidd WSF Yin A 1992 ) Raising Tibet.
Science , 255 , 1663 -1670 .
[本文引用: 1]
[11]
Kembel SW Cowan PD Helmus MR Cornwell WK Morlon H Ackerly DD Blomberg SP Webb CO 2010 ) Picante: R tools for integrating phylogenies and ecology.
Bioinformatics , 26 , 1463 -1464 .
[本文引用: 1]
[12]
Kooyman R Rossetto M Cornwell W Westoby M 2011 ) Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests.
Global Ecology and Biogeography , 20 , 707 -716 .
[13]
Kreft H Jetz W 2010 ) A framework for delineating biogeographical regions based on species distributions.
Journal of Biogeography , 37 , 2029 -2053 .
[本文引用: 1]
[14]
Legendre P Fortin MJ 2010 ) Comparison of the Mantel test and alternative approaches for detecting complex multivariate relationships in the spatial analysis of genetic data.
Molecular Ecology Resources , 10 , 831 -844 .
[本文引用: 1]
[15]
Li R Dao ZL Ji YH Li H 2007 ) A floristic study on the seed plants of the northern Gaoligong Mountains in western Yunnan, China.
Acta Botanica Yunnanica , 29 , 601 -615 .
[本文引用: 1]
(in Chinese with English abstract) [李嵘 , 刀志灵 , 纪运恒 , 李恒 (2007 ) 高黎贡山北段种子植物区系研究. 云南植物研究,
29 , 601 -615 .]
[本文引用: 1]
[16]
Li R Kraft NJB Yang J Wang YH 2015 a) A phylogenetically informed delineation of floristic regions within a biodiversity hotspot in Yunnan, China.
Scientific Reports , 5 , 9396 .
[17]
Li R Kraft NJB Yu HY Li H 2015 b) Seed plant phylogenetic diversity and species richness in conservation planning within a global biodiversity hotspot in eastern Asia.
Conservation Biology , 29 , 1552 -1562 .
[18]
Li XH Zhu XX Niu Y Sun H 2014 ) Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains region, Southwest China.
Journal of Systematics and Evolution , 52 , 280 -288 .
[本文引用: 1]
[19]
Li XW 1985 ) Floristic study of Yunnan Province.
Acta Botanica Yunnanica , 7 , 361 -382 .
[本文引用: 2]
(in Chinese with English abstract) [李锡文 (1985 ) 云南植物区系. 云南植物研究,
7 , 361 -382 .]
[本文引用: 2]
[20]
Li XW 1994 ) Two big biodiversity centers of Chinese endemic genera of seed plants and their characteristics in Yunnan Province.
Acta Botanica Yunnanica , 16 , 221 -227 .
[本文引用: 3]
(in Chinese with English abstract) [李锡文 (1994 ) 中国特有属在云南的两大生物多样性中心及其特征. 云南植物研究,
16 , 221 -227 .]
[本文引用: 3]
[21]
Li XW 1995 a) A floristic study on the seed plants from tropical Yunnan.
Acta Botanica Yunnanica , 17 , 115 -128 .
(in Chinese with English abstract) [李锡文 (1995 a) 云南热带种子植物区系. 云南植物研究,
17 , 115 -128 .]
[22]
Li XW 1995 b) A floristic study on the seed plants from the region of Yunnan Plateau.
Acta Botanica Yunnanica , 17 , 1 -14 .
(in Chinese with English abstract) [李锡文 (1995 b) 云南高原地区种子植物区系. 云南植物研究,
17 , 1 -14 .]
[23]
Li XW Li J 1992 ) On the validity of Tanaka line and its significance viewed from the distribution of eastern Asiatic genera in Yunnan.
Acta Botanica Yunnanica , 14 , 1 -12 .
[本文引用: 1]
(in Chinese with English abstract) [李锡文 , 李捷 (1992 ) 从滇产东亚属的分布论述“田中线”的真实性和意义. 云南植物研究,
14 , 1 -12 .]
[本文引用: 1]
[24]
Li XW Li J 1993 ) A preliminary floristic study of the seed plants from the region of the Hengduan Mountains.
Acta Botanica Yunnanica , 15 , 217 -231 .
[本文引用: 1]
(in Chinese with English abstract) [李锡文 , 李捷 (1993 ) 横断山脉地区种子植物区系的初步研究. 云南植物研究,
15 , 217 -231 .]
[本文引用: 1]
[25]
Li XW Li J 1997 ) The Tanaka-Kaiyong line—an important floristic line for the study of the flora of East Asia.
Annals of the Missouri Botanical Garden , 84 , 888 -892 .
[本文引用: 1]
[26]
Li XW Walker D 1986 ) The plant geography of Yunnan Province, Southwest China.
Journal of Biogeography , 13 , 367 -397 .
[本文引用: 1]
[27]
Liu JQ Duan YW Hao G Ge XJ Sun H 2014 ) Evolutionary history and underlying adaptation of alpine plants on the Qinghai-Tibet Plateau.
Journal of Systematics and Evolution , 52 , 241 -249 .
[本文引用: 1]
[28]
López-Pujol J Zhang FM Sun HQ Ying TS Ge S 2011 ) Centres of plant endemism in China: places for survival or for speciation?
Journal of Biogeography , 38 , 1267 -1280 .
[本文引用: 1]
[29]
Luo D Yue JP Sun WG Xu B Li ZM Comes HP Sun H 2016 ) Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains: first insights from comparative phylogeography of four perennial herbs.
Journal of Biogeography , 43 , 31 -43 .
[本文引用: 1]
[1]
An ZS Kutzbach JE Prell WL Porter SC 2001 ) Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since late Miocene times.
Nature , 411 , 62 -66 .
[本文引用: 1]
[2]
APG III 2009 ) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
Botanical Journal of the Linnean Society , 161 , 105 -121 .
[本文引用: 2]
[30]
Maechler M Rousseeuw P Struyf A Hubert M Hornik K 2015 ) Cluster: Cluster Analysis Basics and Extensions. R package version 2.
0 .3 .
[本文引用: 1]
[31]
Mittermeier RA Robles PG Hoffmann M Pilgrim J Brooks T Mittermeier CG da Fonseca GAB 2004 ) Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. CEMEX/Agrupaciaon Sierra Madre, Mexico City.
[本文引用: 1]
[3]
Bryant JA Lamanna C Morlon H Kerkhoff AJ Enquist BJ Green JL 2008 ) Microbes on mountainsides: contrasting elevational patterns of bacterial and plant diversity. Proceedings of the National Academy of Sciences,
USA , 105 , 11505 -11511 .
[本文引用: 1]
[4]
Elton C 1946 ) Competition and the structure of ecological communities.
Journal of Animal Ecology , 15 , 54 -68 .
[本文引用: 1]
[32]
Oksanen J Blanchet FG Kindt R Legendre P Minchin PR O’Hara RB Simpson GL Solymos P Stevens MHH Wagner H 2015 ) Vegan: Community Ecology Package. R package version 2.
0 .3 .
[本文引用: 1]
[33]
Peng H 1996 ) The floristic equilibrium point of seed plants in Mt. Wuliangshan.
Acta Botanica Yunnanica , 18 , 385 -397 .
[本文引用: 1]
(in Chinese with English abstract) [彭华 (1996 ) 无量山种子植物的区系平衡点. 云南植物研究,
18 , 385 -397 .]
[本文引用: 1]
[34]
Qian H Richard F Zhang JL Zhang J Chen SB 2016 ) Phylogenetic structure and ecological and evolutionary determinants of species richness for angiosperm trees in forest communities in China.
Journal of Biogeography , 43 , 603 -615 .
[本文引用: 1]
[5]
Emerson BC Gillespie RG 2008 ) Phylogenetic analysis of community assembly and structure over space and time.
Trends in Ecology and Evolution , 23 , 619 -630 .
[本文引用: 1]
[6]
Faith DP 1992 ) Conservation evaluation and phylogenetic diversity.
Biological Conservation , 61 , 1 -10 .
[7]
Forest F Grenyer R Rouget M Davies TJ Cowling RM Faith DP Balmford A Manning JC Proches S van der Bank M Reeves G Hedderson TAJ Savolainen V 2007 ) Preserving the evolutionary potential of floras in biodiversity hotspots.
Nature , 445 , 757 -760 .
[本文引用: 1]
[35]
Qian H Swenson NG Zhang JL 2013 a) Phylogenetic beta diversity of angiosperms in North America.
Global Ecology and Biogeography , 22 , 1152 -1161 .
[36]
Qian H Wiens JJ Zhang J Zhang YJ 2015 ) Evolutionary and ecological causes of species richness patterns in North American angiosperm trees.
Ecography , 38 , 241 -250 .
[37]
Qian H Zhang YJ Zhang J Wang XL 2013 b) Latitudinal gradients in phylogenetic relatedness of angiosperm trees in North America.
Global Ecology and Biogeography , 22 , 1183 -1191 .
[38]
Qiao XJ Jabot F Tang ZY Jiang MX Fang JY 2015 ) A latitudinal gradient in tree community assembly processes evidence in Chinese forests.
Global Ecology and Biogeography , 24 , 314 -323 .
[本文引用: 1]
[39]
R Core Team 2014 ) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
[本文引用: 1]
[40]
Ricklefs RE 1987 ) Community diversity: relative roles of local and regional processes.
Science , 235 , 161 -171 .
[本文引用: 1]
[41]
Sørensen T 1948 ) A method of establishing groups of equal amplitude in plant sociology based on similarity of species, and its application to analyses of the vegetation on Danish commons.
Biologiske Skrifter , 5 , 1 -34 .
[42]
Spicer RA Harris NBW Widdowson M Herman AB Guo S Valdes PJ Wolfe JA Kelley SP 2003 ) Constant elevation of southern Tibet over the past 15 million years.
Nature , 421 , 622 -624 .
[本文引用: 1]
[43]
Sun H 2002 ) Evolution of Arctic-Tertiary flora in Himalayan-Hengduan Mountains.
Acta Botanica Yunnanica , 24 , 671 -688 .
[本文引用: 1]
(in Chinese with English abstract) [孙航 (2002 ) 北极-第三纪成分在喜马拉雅-横断山的发展及演化. 云南植物研究,
24 , 671 -688 .]
[本文引用: 1]
[44]
Swenson NG Umaña MN 2014 ) Phylofloristics: an example from the Lesser Antilles.
Journal of Plant Ecology , 7 , 166 -175 .
[45]
Tucker CM Cadotte MW Davies TJ Rebelo AG 2012 ) Incorporating geographical and evolutionary rarity into conservation prioritization.
Conservation Biology , 26 , 593 -601 .
[本文引用: 1]
[46]
Wang Y 2006 ) Yunnan Mountain Climate . Yunnan Science and Technology Press, Kunming .
[本文引用: 1]
(in Chinese) [王宇 (2006 ) 云南山地气候. 云南科技出版社 , 昆明.]
[本文引用: 1]
[47]
Wang ZH Fang JY Tang ZY Lin X 2012 ) Relative role of contemporary environment versus history in shaping diversity patterns of China’s woody plants.
Ecography , 35 , 1124 -1133 .
[48]
Webb CO 2000 ) Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
The American Naturalist , 156 , 145 -155 .
[本文引用: 1]
[49]
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002 ) Phylogenies and community ecology.
Annual Review of Ecology and Systematics , 33 , 475 -505 .
[本文引用: 1]
[50]
Webb CO Donoghue MJ 2005 ) Phylomatic: tree assembly for applied phylogenetics.
Molecular Ecology Notes , 5 , 181 -183 .
[本文引用: 1]
[51]
Wu ZY 1977 -2006) Flora Yunnanica . Science Press, Beijing .
(in Chinese) [吴征镒 (1977 -2006) 云南植物志. 科学出版社 , 北京.]
[52]
Wu ZY 1987 ) Vegetation of Yunnan . Science Press, Beijing .
[本文引用: 4]
(in Chinese) [吴征镒 (1987 ) 云南植被. 科学出版社 , 北京.]
[本文引用: 4]
[53]
Wu ZY Sun H Zhou ZK Li DZ Peng H 2010 ) Floristics of Seed Plants from China . Science Press, Beijing .
[本文引用: 3]
(in Chinese) [吴征镒 , 孙航 , 周浙昆 , 李德铢 , 彭华 (2010 ) 中国种子植物区系地理. 科学出版社 , 北京.]
[本文引用: 3]
[54]
Wu ZY Wu SG 1996 ) A proposal for a new floristic kingdom (realm)—the E. Asiatic Kingdom, its delineation and characteristics. In: Floristic Characteristics and Diversity of East Asian Plants (eds Zhang AL, Wu SG) , pp. 3 -42. China Higher Education Press, Beijing; Springer-Verlag, Berlin, Heidelberg.
[55]
Wu ZY Zhou ZK Sun H Li DZ Peng H 2006 ) The Areal-Types of Seed Plants and Their Origin and Differentiation . Yunnan Science and Technology Press, Kunming .
[本文引用: 4]
(in Chinese) [吴征镒 , 周浙昆 , 孙航 , 李德铢 , 彭华 (2006 ) 种子植物分布区类型及其起源和分化. 云南科技出版社 , 昆明.]
[本文引用: 4]
[56]
Yang YG 1990 ) Comprehensive Physical Regionalization in Yunnan . China Higher Education Press, Beijing .
[本文引用: 1]
(in Chinese) [杨一光 (1990 ) 云南省综合自然区划. 中国高等教育出版社 , 北京.]
[本文引用: 1]
[57]
Zanne AE Tank DC Cornwell WK Eastman JM Smith SA FitzJohn RG McGlinn DJ O’Meara BC Moles AT Reich PB Royer DL Soltis DE Stevens PF Westoby M Wright IJ Aarssen L Bertin RL Calaminus A Govaerts R Hemmings F Leishman MR Oleksyn J Soltis PS Swenson NG Warman L Beaulieu JM 2014 ) Three keys to the radiation of angiosperms into freezing environments.
Nature , 506 , 89 -92 .
[本文引用: 1]
[58]
Zhu H 2008 ) The tropical flora of southern Yunnan, China, and its biogeographical affinities.
Annals of the Missouri Botanical Garden , 95 , 661 -680 .
[本文引用: 1]
[59]
Zhu H 2012 ) Biogeographical divergence of the flora of Yunnan, southwestern China initiated by the uplift of Himalaya and extrusion of Indochina block.
PLoS ONE , 7 , e45601 .
[本文引用: 1]
[60]
Zhu H 2013 ) The floras of southern and tropical southeastern Yunnan have been shaped by divergent geological histories.
PLoS ONE , 8 , e64213 .
[本文引用: 1]
[61]
Zhu H Cao M Hu H 2006 ) Geological history, flora, and vegetation of Xishuangbanna, southern Yunnan, China.
Biotropica , 38 , 310 -317 .
[本文引用: 1]
DNA条形码在植物系统发育区系学研究中的应用. 生物多样性,
2
2015
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
... ; 葛学军, 2015 ). ...
DNA条形码在植物系统发育区系学研究中的应用. 生物多样性,
2
2015
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
... ; 葛学军, 2015 ). ...
Phylogenetic turnover in tropical tree communities: impact of environmental filtering, biogeography and mesoclimatic niche conservatism.
1
2012
... 将云南植物区系的所有种子植物属作为物种库, 以种子植物系统发育关系(Zanne et al, 2014 )为骨架, 利用Phylomatic软件构建基于全部属的系统发育树(Webb & Donoghue, 2005 ).对于科内无法解决的属间关系, 则以多歧分支(polytomy)的方式进行处理(Hardy et al, 2012 ) (图2 ). ...
Raising Tibet.
1
1992
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
Picante: R tools for integrating phylogenies and ecology.
1
2010
... 上述所有分析均采用R语言(R Core Team, 2014 )的vegan (Oksanen et al, 2015 )、picante (Kembel et al, 2010 )、cluster (Maechler et al, 2015 )等程序包计算. ...
Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests.
2011
A framework for delineating biogeographical regions based on species distributions.
1
2010
... 为了直观显示不同地理单元之间的区系组成相似程度, 采用Ward的方法对分类群组成及系统发育组成相似性系数进行等级聚类分析(Kreft & Jetz, 2010 ). ...
Comparison of the Mantel test and alternative approaches for detecting complex multivariate relationships in the spatial analysis of genetic data.
1
2010
... 式中, Sij 是i 、j 两个地理单元共有的分类群数目, Si 与Sj 是i 、j 两个地理单元各自的分类群数目; 类似地, BLij 是i 、j 两个地理单元共有的支长之和, BLi 与BLj 是i 、j 两个地理单元各自的支长总和.Sørenson与PhyloSor的值在0与1之间.如为0, 则表示植物区系完全不相同; 如为1, 则表示植物区系完全相同.运用Mantel检验分析分类群组成相似性与系统发育组成相似性之间的相关性(Legendre & Fortin, 2010 ). ...
高黎贡山北段种子植物区系研究. 云南植物研究,
1
2007
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
高黎贡山北段种子植物区系研究. 云南植物研究,
1
2007
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
a) A phylogenetically informed delineation of floristic regions within a biodiversity hotspot in Yunnan, China.
2015
b) Seed plant phylogenetic diversity and species richness in conservation planning within a global biodiversity hotspot in eastern Asia.
2015
Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains region, Southwest China.
1
2014
... 我国有很好的植物区系研究基础, 但从进化历史的角度研究植物区系才刚刚起步.当前研究主要以大型森林动态样地为平台或以木本植物为主, 探讨不同生态过程对不同空间尺度树种系统发育组成的影响(如: Wang et al, 2012 ; Qiao et al, 2015 ; Qian et al, 2016 ), 但这些研究忽视了反映林下小生境对植物区系形成起重要作用的草本植物的影响, 而目前基于特定地区所有植物种类的植物系统发育区系地理学研究仅有零星报道(Li et al, 2014 , 2015a, 2015b ).我国幅员辽阔, 不同地区的地质历史与现代自然环境千差万别, 植物区系的形成原因大相径庭, 因此, 在我国开展植物系统发育区系地理学研究, 对深入理解中国植物区系的成因具有重要意义. ...
云南植物区系. 云南植物研究,
2
1985
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物种类丰富, 早期根据科属分布区类型的分析表明, 云南植物区系地理成分复杂、联系广泛(李锡文, 1985 ; 吴征镒, 1987 ), 该结论得到本研究的支持.从云南种子植物所含1,983个属所展示的进化关系来看(图2 ), 这些属在系统发育树上的各个主要分支均有分布(如: 祼子植物、基部被子植物、单子叶植物、基部真双子叶植物、蔷薇类、菊类等), 表明云南植物区系在地质历史的不同时期与其他地区的植物区系有过广泛的联系和交流, 并在适应当地的自然环境下产生了大量的后裔, 逐渐演变为当今植物区系的外貌. ...
云南植物区系. 云南植物研究,
2
1985
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物种类丰富, 早期根据科属分布区类型的分析表明, 云南植物区系地理成分复杂、联系广泛(李锡文, 1985 ; 吴征镒, 1987 ), 该结论得到本研究的支持.从云南种子植物所含1,983个属所展示的进化关系来看(图2 ), 这些属在系统发育树上的各个主要分支均有分布(如: 祼子植物、基部被子植物、单子叶植物、基部真双子叶植物、蔷薇类、菊类等), 表明云南植物区系在地质历史的不同时期与其他地区的植物区系有过广泛的联系和交流, 并在适应当地的自然环境下产生了大量的后裔, 逐渐演变为当今植物区系的外貌. ...
中国特有属在云南的两大生物多样性中心及其特征. 云南植物研究,
3
1994
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
... )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
中国特有属在云南的两大生物多样性中心及其特征. 云南植物研究,
3
1994
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
... )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
a) 云南热带种子植物区系. 云南植物研究,
1995
a) 云南热带种子植物区系. 云南植物研究,
1995
b) 云南高原地区种子植物区系. 云南植物研究,
1995
b) 云南高原地区种子植物区系. 云南植物研究,
1995
从滇产东亚属的分布论述“田中线”的真实性和意义. 云南植物研究,
1
1992
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
从滇产东亚属的分布论述“田中线”的真实性和意义. 云南植物研究,
1
1992
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
横断山脉地区种子植物区系的初步研究. 云南植物研究,
1
1993
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
横断山脉地区种子植物区系的初步研究. 云南植物研究,
1
1993
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
The Tanaka-Kaiyong line—an important floristic line for the study of the flora of East Asia.
1
1997
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
The plant geography of Yunnan Province, Southwest China.
1
1986
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
Evolutionary history and underlying adaptation of alpine plants on the Qinghai-Tibet Plateau.
1
2014
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
Centres of plant endemism in China: places for survival or for speciation?
1
2011
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains: first insights from comparative phylogeography of four perennial herbs.
1
2016
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since late Miocene times.
1
2001
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
2
2009
... 基于云南植物区系分区研究(Li et al, 2015a )划分的8个地理单元(图1 ), 以《云南植物志》为基础(吴征镒, 1977-2006), 构建不同地理单元种子植物的属级“有/无”数据矩阵.区系组成中的外来成分或栽培种类不列入统计.科属范畴的界定和划分依据被子植物系统发育分类系统(Angiosperm Phylogeny Group Classification, APG)的观点(APG III, 2009 ).本研究共计包括8,072条数据记录. ...
... 基于被子植物系统发育分类系统(APG III, 2009 )的统计, 云南种子植物区系计有225个科1,983个属.8个地理单元的科、属组成及系统发育多样性见表1 .地理单元VIII具有最高的科、属丰富度及系统发育组成多样性, 地理单元VI具有最低的科、属丰富度及系统发育多样性.不同地理单元植物区系的系统发育多样性与科(R2 = 0.97, P < 0.001)或属(R2 = 0.99, P < 0.001)的丰富度呈显著正相关(图3 ). ...
Cluster: Cluster Analysis Basics and Extensions. R package version 2.
1
2015
... 上述所有分析均采用R语言(R Core Team, 2014 )的vegan (Oksanen et al, 2015 )、picante (Kembel et al, 2010 )、cluster (Maechler et al, 2015 )等程序包计算. ...
Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. CEMEX/Agrupaciaon Sierra Madre, Mexico City.
1
2004
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
Microbes on mountainsides: contrasting elevational patterns of bacterial and plant diversity. Proceedings of the National Academy of Sciences,
1
2008
... 采用Sørenson分类群相似性指数(Sørenson, 1948 )及PhyloSor系统发育相似性指数(Bryant et al, 2008 )测度云南植物区系不同地理单元的分类群组成与系统发育组成的相似程度, 其计算公式如下: ...
Competition and the structure of ecological communities.
1
1946
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
Vegan: Community Ecology Package. R package version 2.
1
2015
... 上述所有分析均采用R语言(R Core Team, 2014 )的vegan (Oksanen et al, 2015 )、picante (Kembel et al, 2010 )、cluster (Maechler et al, 2015 )等程序包计算. ...
无量山种子植物的区系平衡点. 云南植物研究,
1
1996
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
无量山种子植物的区系平衡点. 云南植物研究,
1
1996
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
Phylogenetic structure and ecological and evolutionary determinants of species richness for angiosperm trees in forest communities in China.
1
2016
... 我国有很好的植物区系研究基础, 但从进化历史的角度研究植物区系才刚刚起步.当前研究主要以大型森林动态样地为平台或以木本植物为主, 探讨不同生态过程对不同空间尺度树种系统发育组成的影响(如: Wang et al, 2012 ; Qiao et al, 2015 ; Qian et al, 2016 ), 但这些研究忽视了反映林下小生境对植物区系形成起重要作用的草本植物的影响, 而目前基于特定地区所有植物种类的植物系统发育区系地理学研究仅有零星报道(Li et al, 2014 , 2015a, 2015b ).我国幅员辽阔, 不同地区的地质历史与现代自然环境千差万别, 植物区系的形成原因大相径庭, 因此, 在我国开展植物系统发育区系地理学研究, 对深入理解中国植物区系的成因具有重要意义. ...
Phylogenetic analysis of community assembly and structure over space and time.
1
2008
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
Conservation evaluation and phylogenetic diversity.
1992
Preserving the evolutionary potential of floras in biodiversity hotspots.
1
2007
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
a) Phylogenetic beta diversity of angiosperms in North America.
2013
Evolutionary and ecological causes of species richness patterns in North American angiosperm trees.
2015
b) Latitudinal gradients in phylogenetic relatedness of angiosperm trees in North America.
2013
A latitudinal gradient in tree community assembly processes evidence in Chinese forests.
1
2015
... 我国有很好的植物区系研究基础, 但从进化历史的角度研究植物区系才刚刚起步.当前研究主要以大型森林动态样地为平台或以木本植物为主, 探讨不同生态过程对不同空间尺度树种系统发育组成的影响(如: Wang et al, 2012 ; Qiao et al, 2015 ; Qian et al, 2016 ), 但这些研究忽视了反映林下小生境对植物区系形成起重要作用的草本植物的影响, 而目前基于特定地区所有植物种类的植物系统发育区系地理学研究仅有零星报道(Li et al, 2014 , 2015a, 2015b ).我国幅员辽阔, 不同地区的地质历史与现代自然环境千差万别, 植物区系的形成原因大相径庭, 因此, 在我国开展植物系统发育区系地理学研究, 对深入理解中国植物区系的成因具有重要意义. ...
R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
1
2014
... 上述所有分析均采用R语言(R Core Team, 2014 )的vegan (Oksanen et al, 2015 )、picante (Kembel et al, 2010 )、cluster (Maechler et al, 2015 )等程序包计算. ...
Community diversity: relative roles of local and regional processes.
1
1987
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
A method of establishing groups of equal amplitude in plant sociology based on similarity of species, and its application to analyses of the vegetation on Danish commons.
1948
Constant elevation of southern Tibet over the past 15 million years.
1
2003
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
北极-第三纪成分在喜马拉雅-横断山的发展及演化. 云南植物研究,
1
2002
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
北极-第三纪成分在喜马拉雅-横断山的发展及演化. 云南植物研究,
1
2002
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
Phylofloristics: an example from the Lesser Antilles.
2014
Incorporating geographical and evolutionary rarity into conservation prioritization.
1
2012
... 云南植物区系非随机的系统发育结构与其进化历史有关.自第三纪以来, 伴随着印度板块与欧亚板块的碰撞, 大规模的造山运动导致云南及其周边地区产生了复杂多样的地理环境(Harrison et al, 1992 ; An et al, 2001 ; Spicer et al, 2003 ), 第四纪的气候变化更促进了云南植物区系的发展(Li & Walker, 1986 ).地理单元III异质的生境及多样的气候条件(如: 热带、亚热带与温带气候), 加之处于热带与温带植物区系的过渡带, 有助于不同生态耐受性的物种同域生存, 从而造就了该区域多样的系统发育组成; 相反, 对其他地理单元来说, 尽管也具有多样的生境与气候条件(如: 地理单元I与VII), 但因不是植物区系的交汇区, 不利于不同生态耐受性的物种同域共存, 从而导致相对较低的系统发育组成.与此相似的系统发育结构模式在南非好望角植物区系研究中已有报道(Forest et al, 2007 ; Tucker et al, 2012 ), 因为地质历史及气候因素的不同, 尽管好望角西部的物种多样性比东部高, 但西部的系统发育组成却较东部低. ...
1
2006
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
1
2006
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
Relative role of contemporary environment versus history in shaping diversity patterns of China’s woody plants.
2012
Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
1
2000
... 采用净相关指数(net relatedness index, NRI)度量不同地理单元的系统发育结构(Webb, 2000 ), 其计算公式如下: ...
Phylogenies and community ecology.
1
2002
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
Phylomatic: tree assembly for applied phylogenetics.
1
2005
... 将云南植物区系的所有种子植物属作为物种库, 以种子植物系统发育关系(Zanne et al, 2014 )为骨架, 利用Phylomatic软件构建基于全部属的系统发育树(Webb & Donoghue, 2005 ).对于科内无法解决的属间关系, 则以多歧分支(polytomy)的方式进行处理(Hardy et al, 2012 ) (图2 ). ...
4
1987
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
... 云南植物种类丰富, 早期根据科属分布区类型的分析表明, 云南植物区系地理成分复杂、联系广泛(李锡文, 1985 ; 吴征镒, 1987 ), 该结论得到本研究的支持.从云南种子植物所含1,983个属所展示的进化关系来看(图2 ), 这些属在系统发育树上的各个主要分支均有分布(如: 祼子植物、基部被子植物、单子叶植物、基部真双子叶植物、蔷薇类、菊类等), 表明云南植物区系在地质历史的不同时期与其他地区的植物区系有过广泛的联系和交流, 并在适应当地的自然环境下产生了大量的后裔, 逐渐演变为当今植物区系的外貌. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
4
1987
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
... 云南植物种类丰富, 早期根据科属分布区类型的分析表明, 云南植物区系地理成分复杂、联系广泛(李锡文, 1985 ; 吴征镒, 1987 ), 该结论得到本研究的支持.从云南种子植物所含1,983个属所展示的进化关系来看(图2 ), 这些属在系统发育树上的各个主要分支均有分布(如: 祼子植物、基部被子植物、单子叶植物、基部真双子叶植物、蔷薇类、菊类等), 表明云南植物区系在地质历史的不同时期与其他地区的植物区系有过广泛的联系和交流, 并在适应当地的自然环境下产生了大量的后裔, 逐渐演变为当今植物区系的外貌. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
3
2010
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
... ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
3
2010
... 云南是全球34个生物多样性热点地区中3个热点地区(喜马拉雅、印度-缅甸及中国西南山地)的交汇区(Mittermeier et al, 2004 ), 植物种类位居全国之首(吴征镒, 1987 ), 区系起源古老、地理成分复杂、特有现象丰富, 是很多第三纪孑遗植物的避难所(李锡文, 1985 , 1994 ; López-Pujol et al, 2011 ).在中国植物区系区划中, 本区域地跨古热带植物区、中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区(吴征镒等, 2010 ).近来基于系统发育的区系研究将云南分为8个地理单元(Li et al, 2015a ), 然而, 各个地理单元的系统发育组成、系统发育结构及不同地理单元间的进化历史相似性仍缺乏深入的 讨论. ...
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
... ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
4
2006
... 植物区系(flora)是某一特定地区生长着的全部植物种类, 是植物科属种的自然综合体(吴征镒等, 2006 ).植物区系研究在植物种类清查、生物资源开发、环境保护、国民经济建设等方面起着至关重要的作用(吴征镒等, 2006 ).我国植物区系地理学研究主要集中于对一个自然地理区域内所有植物的地理成分从科、属、种三个层次进行分析, 并通过与邻近地区的植物区系比较, 确定该地区现代植物区系的性质、来源和地位, 如吴征镒等(2010)关于我国种子植物区系的研究, 表明中国种子植物区系的亚热带性质及热带亲缘等特点. ...
... ).植物区系研究在植物种类清查、生物资源开发、环境保护、国民经济建设等方面起着至关重要的作用(吴征镒等, 2006 ).我国植物区系地理学研究主要集中于对一个自然地理区域内所有植物的地理成分从科、属、种三个层次进行分析, 并通过与邻近地区的植物区系比较, 确定该地区现代植物区系的性质、来源和地位, 如吴征镒等(2010)关于我国种子植物区系的研究, 表明中国种子植物区系的亚热带性质及热带亲缘等特点. ...
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
... ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
4
2006
... 植物区系(flora)是某一特定地区生长着的全部植物种类, 是植物科属种的自然综合体(吴征镒等, 2006 ).植物区系研究在植物种类清查、生物资源开发、环境保护、国民经济建设等方面起着至关重要的作用(吴征镒等, 2006 ).我国植物区系地理学研究主要集中于对一个自然地理区域内所有植物的地理成分从科、属、种三个层次进行分析, 并通过与邻近地区的植物区系比较, 确定该地区现代植物区系的性质、来源和地位, 如吴征镒等(2010)关于我国种子植物区系的研究, 表明中国种子植物区系的亚热带性质及热带亲缘等特点. ...
... ).植物区系研究在植物种类清查、生物资源开发、环境保护、国民经济建设等方面起着至关重要的作用(吴征镒等, 2006 ).我国植物区系地理学研究主要集中于对一个自然地理区域内所有植物的地理成分从科、属、种三个层次进行分析, 并通过与邻近地区的植物区系比较, 确定该地区现代植物区系的性质、来源和地位, 如吴征镒等(2010)关于我国种子植物区系的研究, 表明中国种子植物区系的亚热带性质及热带亲缘等特点. ...
... 然而, 植物区系是一个地区植物发生发展过程的反映, 它是在地质历史长河中与环境共同作用的结果(吴征镒等, 2006 ).除现代自然环境的影响外, 进化历史过程中可能发生的隔离、分化、扩散或绝灭等事件对植物区系的形成也有重要影响(Ricklefs, 1987 ).因此, 植物区系的研究需包含进化历史的信息, 方能更全面地理解植物区系的性质和来源(葛学军, 2015 ).早期研究中, 利用科属或属种的比例来反映一个地区植物区系的进化历史(Elton, 1946 ; 彭华, 1996 ; 李嵘等, 2007 ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
... ), 或者通过科或属的分类群统计和分析来比较不同植物区系的相似程度(吴征镒等, 2006 ), 但这些方法难以量化进化过程在区系形成中的作用.而且, 这些分析忽略了物种或类群间的进化关系, 从而难以揭示不同区系间进化历史的相似程度.系统发育分析方法在区域尺度上的应用为理解进化历史对现代植物区系的影响及比较不同区系间系统发育的相似性提供了可量化的指标和途径(Webb et al, 2002 ; Emerson & Gillespie, 2008 ; 葛学军, 2015 ). ...
1
1990
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
1
1990
... 云南地处中国西南, 位于21º8′32″ - 29º15′8″ N, 97º31′39″ - 106º11′47″ E, 是一个高原内陆省份.自晚第三纪末至第四纪初以来的大规模山体隆升和深刻的高原解体, 导致境内河谷深嵌、地势高耸, 整体呈西北向东南倾斜, 最高峰海拔6,740 m, 最低点海拔76.4 m (杨一光, 1990 ).因受印度洋西南季风及太平洋东南季风的影响, 气候具有干湿季分明、降水丰沛、年温差小、日温差大等特点(王宇, 2006 ).正是由于这种复杂的地形地貌加之丰厚的水热条件, 造就了云南丰富多样的植物区系种类(吴征镒, 1987 ). ...
Three keys to the radiation of angiosperms into freezing environments.
1
2014
... 将云南植物区系的所有种子植物属作为物种库, 以种子植物系统发育关系(Zanne et al, 2014 )为骨架, 利用Phylomatic软件构建基于全部属的系统发育树(Webb & Donoghue, 2005 ).对于科内无法解决的属间关系, 则以多歧分支(polytomy)的方式进行处理(Hardy et al, 2012 ) (图2 ). ...
The tropical flora of southern Yunnan, China, and its biogeographical affinities.
1
2008
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
Biogeographical divergence of the flora of Yunnan, southwestern China initiated by the uplift of Himalaya and extrusion of Indochina block.
1
2012
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
The floras of southern and tropical southeastern Yunnan have been shaped by divergent geological histories.
1
2013
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
Geological history, flora, and vegetation of Xishuangbanna, southern Yunnan, China.
1
2006
... 云南植物区系的分类群组成与系统发育组成相似性的聚类分析均表明南部地理单元VII与VIII的区系相似, 而北部6个地理单元的区系相近(图4 ).在中国植物区系区划中, 地理单元VII与VIII基本位于古热带植物区内, 其他6个地理单元处于东亚植物区内(吴征镒等, 2010 ), 这从局域尺度和系统发育的角度为Wu和Wu (1996)将东亚植物区提升为“Kingdom”提供了支持.南部地理单元VII与VIII为低海拔山地, 主要受热带气候的影响, 自第三纪以来, 随着印度板块与欧亚板块的碰撞, 热带亚洲成分由南向北渗入, 从而演变成以热带亚洲成分为主的热带植物区系(Li, 1995a ; Zhu et al, 2006 ; Zhu, 2008 , 2013 ), 该区域由于相对稳定的地质构造和气候环境, 成为伯乐树属(Bretschneidera )、蒜头果属(Malania )等大量第三纪孑遗植物的避难所(Li, 1994 ).北部6个地理单元为中海拔山地、云贵高原或高山峡谷区, 主要为亚热带和温带气候, 随着第三纪古地中海的退却及喜马拉雅的隆升, 北温带植物区系成分由北向南渗透, 从而造就了现今以北温带成分为主的温带植物区系(Li & Li, 1993 ; Li, 1995b ; Sun, 2002 ; Zhu, 2012 ), 该地区(特别西北部)地质运动活跃、山体高大、生境类型多样, 成为绵参属(Eriophyton )、复芒菊属(Formania )等诸多新生类群的发源地(Li, 1994 ; Liu et al, 2014 ; Luo et al, 2016 ).北部的6个地理单元内, 地理单元II与VI的区系相似, 地理单元I、III、IV及V的区系相近, 这也印证了东亚植物区系中“中国-日本森林植物亚区及中国-喜马拉雅森林植物亚区”(吴征镒等, 2010 )的区系划分.新生代以来, 自然地理和气候条件在云南东西部的分化是形成不同地理单元间区系分化的重要因素之一(吴征镒, 1987 ), 东部(地理单元II与VI)以石灰岩地貌为主, 深受太平洋季风的影响, 西部(地理单元I、III、IV及V)属高原地貌, 基本为印度洋季风控制(Li & Li, 1992 , 1997 ). ...
 , 孙航
, 孙航

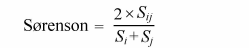
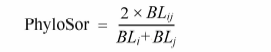


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}