1 .Plant Evolutionary Ecology, University of Tübingen, 72076 Tübingen, Germany;2.中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 1001933 .Department of Biology/Ecology & Evolution, University of Fribourg, 1700 Fribourg, Switzerland;
Biological control opportunities of ragweed are predicted to decrease with climate change in East Asia
Yan Sun1, *, , Zhongshi Zhou2, Rui Wang2, Heinz Müller-Schärer3
1 Plant Evolutionary Ecology, University of Tübingen, 72076 Tübingen, Germany ;2 State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China3 Department of Biology/ Ecology & Evolution, University of Fribourg, 1700 Fribourg, Switzerland;
基金资助: The Advance Postdoc. Mobility fellowship from the Swiss National Science Foundation (SNSF: P300PA-161014 to YS). The Swiss State Secretariat for Education, Research, and Innovation (SERI: 13.0146 to HMS). The Swiss National Science Foundation (SNSF: 31003A_166448/1 to HMS) and the EU COST Action FA1203 ‘Sustainable management of Ambrosia artemisiifolia in Europe (SMA-RTER to HMS)’
展开
摘要
外来入侵植物对入侵地的生态系统与社会经济可造成严重的威胁。如何控制入侵植物对自然资源的危害, 向人类提出了极大的挑战。利用原产地的专食性天敌来控制入侵植物已被广泛证明是一种经济、可持续的生物防治手段。在全球气候变暖的背景下, 生物防治研究的关键问题是全面了解物种的潜在分布区和传播范围, 进而优化入侵植物的生物防治方案。本研究利用生物地理模型, 通过预测两种生物防治昆虫和它们的组合在东亚地区的适宜分布区, 预测豚草(Ambrosia artemisiifolia)的生物防治区域。豚草原产北美, 目前已经入侵全球多个国家和地区。20世纪末, 豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)作为豚草的生物防治昆虫从原产地引入到东亚地区。本研究旨在探讨如下问题: (1)在豚草的适宜生长分布区内, 有多少区域也同样适宜其两种天敌的生存?(2)在目前和未来的气候背景下, 有多少区域适宜豚草生长但是不适宜它的两种天敌生长?(3)在这些适宜豚草生长却不适宜两种昆虫天敌生长的区域内, 需要选择哪些特定的生物型进行投放?为此, 我们基于入侵植物和两种生物防治昆虫的全球分布记录及其分布点的重要生物气候因子, 同时模拟了入侵植物及其两种生物防治天敌在东亚地区的分布范围。排序技术被用来探索气候因子对每个物种的限制作用, 同时也用来检验豚草在北美和东亚地区的生态位重叠和相似性。结果表明, 在当前和未来的气候背景下, 相较于豚草卷蛾, 豚草条纹叶甲与豚草的地理分布范围更加吻合(当前气候: 40.3% vs. 21.6%, 未来气候: 29.8% vs. 20.3%)。气候变化可能会导致两种生物防治天敌(尤其是豚草条纹叶甲)的地理分布与豚草的地理分布的重叠区域减少(42.9% vs. 29.9%)。本研究同时提出了温度和降水等气候因子可用于为特殊区域(生物防治天敌未覆盖的豚草分布区)筛选生物防治天敌的相应株系。
关键词:生物防治
;
生物入侵
;
豚草卷蛾
;
生态位重叠
;
豚草条纹叶甲
;
物种分布
Abstract
The control of invasive alien plants (IAP) that jeopardize our ecosystems and economy constitutes a significant challenge for natural resource management. Classical biological control referring to the introduction of specialist antagonists from the native range has proven to be a highly cost-effective management tool against IAP. A critical issue in biological control research is to guide informed decision-making on the potential spread and distribution and thus impact of biological control candidates, especially under climate change. Here we propose a biogeographic modeling approach to predict the cover of the suitable area of a plant invader in East Asia (EA) by two biological control agents and their combinations. Our study system is Ambrosia artemisiifolia, native to North America and invasive worldwide, and two North American biological control agents, Ophraella communa and Epiblema strenuana that were accidentally and deliberately introduced into East Asia (EA) in the late 20th century, respectively. Specifically, we ask: (1) what percentage of the suitable A. artemisiifolia area is also suitable for the two agents in EA, and (2) which part of the suitable A. artemisiifolia area in EA is likely to remain uncovered by these two agents, both under current and future climatic scenarios; and (3) which particular biotypes would be needed to fill in the yet uncovered part of the suitable A. artemisiifolia range in East Asia? For this, we simultaneously modelled the species distributions based on worldwide occurrences and important bioclimatic variables for the target invasive plant and its two biological control agents. Ordination techniques were used to explore climatic constraints of each species and to perform niche overlap and similarity tests with A. artemisiifolia between its native North American and introduced EA range. Our results show that O. communa has a larger overlap with the geographic range of A. artemisiifolia than E. strenuana, both under current (40.3% vs. 21.6% for O. communa and E. strenuana, respectively) and future climatic scenarios (29.8% vs. 20.3% for O. communa and E. strenuana, respectively). Importantly, climate change is expected to reduce the total geographic overlap of A. artemisiifolia by the two agents combined (42.9% vs. 29.8% for current and future climate conditions, respectively), with a higher reduction by O. communa than by E. strenuana. Our analyses also identified for which abiotic conditions to select in order to develop climatically adapted strains for particular regions, where A. artemisiifolia is presently unlikely to be covered.
Keywords:biological control
;
biological invasions
;
Epiblema strenuana
;
niche overlap
;
Ophraella communa
;
species distribution
YanSun, ZhongshiZhou, RuiWang, HeinzMüller-Schärer. Biological control opportunities of ragweed are predicted to decrease with climate change in East Asia[J]. Biodiversity Science, 2017, 25(12): 1285-1294 https://doi.org/10.17520/biods.2017096
我们通过文献资料收集全球范围内所有可用的豚草及其两种专食性天敌(豚草条纹叶甲和豚草卷蛾)的分布记录。3个物种分布点的地理坐标信息主要从以下4个网络资源中下载获得: (1) the Global Biodiversity Information Facility Online Database (GBIF; http://www.gbif.org); (2) the Southwest Environmental Information Network (SEINet; http://swbiodiversity.org); (3) the Barcode of Life Data Systems (BOLD; http://www.boldsystems.org/); (4) the Berkeley Ecoinformatics Engine (Ecoengine; https://ecoengine.berkeley.edu/)。更多分布数据可见于Sun等(2017)文章的附录信息。
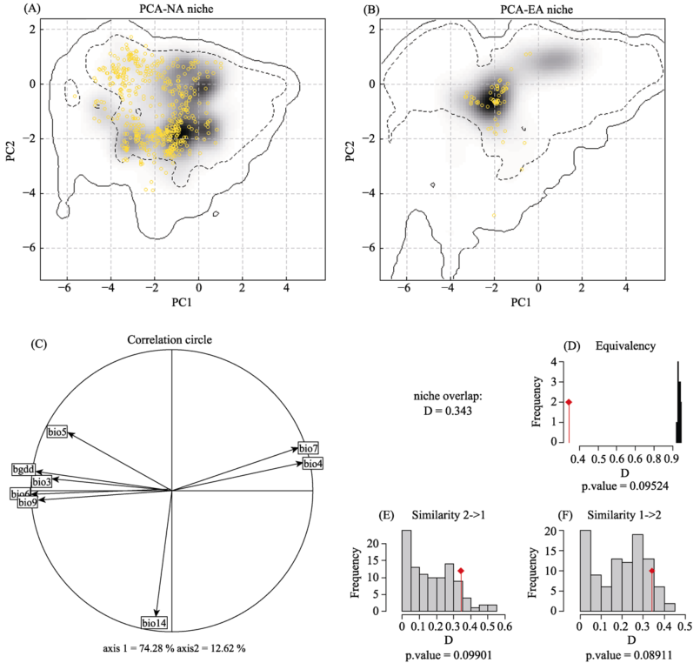
使用R软件中ecospat包的排序技术比较了北美原产地和东亚入侵地的豚草生态位(Broennimann et al, 2014)。基于豚草分布模型筛选出的9个生物气候变量矩阵, 利用PCA-env的前两个主成分坐标轴对生态位进行量化。在生态位重叠变量的排序中, ecospat已经被证实明显优于其他排序方法(Broennimann et al, 2012; Cola et al, 2016)。因此, 我们用Ecospat中的PCA-env来检测生态位的等价性和相似性。对两个区域的分布记录进行随机分配后, 可以通过等价性检验(niche equivalency)来计算其生态位重叠区域是否恒定。生态位相似性(niche similarity)检验了两个地区的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域。该检验同时从北美到东亚和从东亚到北美的两个方向上进行(Broennimann et al, 2012)。生态位等价性和相似性检验均使用100次重复。
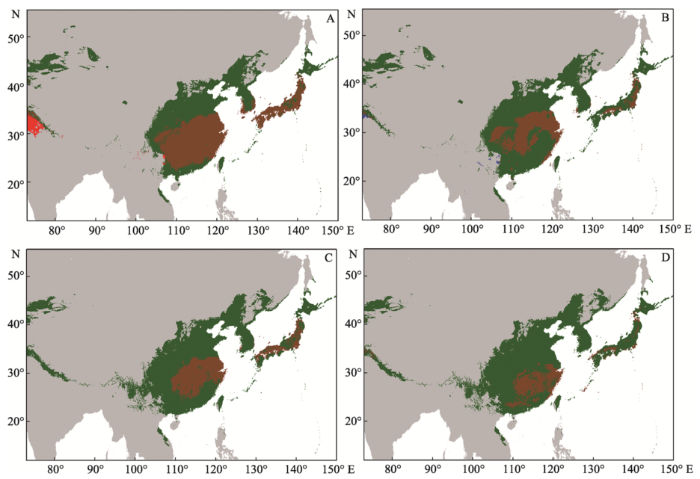
Fig. 1 Geographical predictions of Ambrosia artemisiifolia and two biological control insects for East Asia, under present and future climatic scenarios. The climatic suitability indicates the optimal threshold of the percentage of models predicting each species. Dark green in all figures, A. artemisiifolia; under current climatic conditions: (A) Red, Ophraella communa; sienna, overlap 40.3%; (B) Blue, Epiblema strenuana; sienna, overlap 21.6%; under future climatic scenarios: (C) Red, Ophraella communa; sienna, overlap 29.8%; (D) Blue, Epiblema strenuana; sienna, overlap 20.3%. Models calibrated in East Asia only.
Fig. 2 Niche of Ambrosia artemisiifolia in climatic space using principal component analysis (PCA-env). Panels (A) and (B) represent the niche of the species along the two first axes of the PCA for the native North American (NA) and introduced East Asian (EA) range, respectively. Gray shading shows the density of the occurrences of the species by the cell. The solid contour lines illustrate 100% of the available environment, and dashed lines indicate the 50% of the most common background environment. Yellow circles in (A) and (B) give the occurrences of two insect species in NA and in EA. The contribution of the climatic variables of the two axes of the PCA and the percentage of inertia explained by the two axes is given in (C). Histograms (D-F) show the observed niche overlap between the two ranges (bars and a diamond) and simulated niche overlaps (gray bars) on which tests of niche equivalency (D), niche similarity of EA and NA (E), and niche similarity of NA and EA (F) are calculated from 100 iterations, with the significance level of the tests.
Appendix 3 Geographic continence predictions of Ambrosia artemisiifolia and its two biological control agents, Ophraella communa and Epibleama strenuana under present climatic scenarios in East Asia with occurrences presented.
Appendix 4 Geographic predictions of overlap of Ambrosia artemisiifolia and its two biological control agents Ophraella communa and Epiblema strenuana under present and future climatic scenarios in East Asia.
Appendix 5 Mean and standard deviation of geographic predictions of Ambrosia artemisiifolia and its two biological control agents Ophraella communa and Epiblema strenuana under four future climatic scenarios in East Asia.
Post-release evaluation of biological control of Bemisia tabaci biotype “B” in the USA and the development of predictive tools to guide introductions for other countries.
Selecting arthropod biological control agents against arthropod pests: Can the science be improved to decrease the risk of releasing ineffective agents?
MaJ, GuoJY, WanFH, HuX, WanFH, LiB, GuoJ (2008) Biological control of Ambrosia artemisiifolia and A. trifida. In: Biological Invasions: Biological Control Theory and Practice, pp. 157-185. Science Press, Beijing. (in Chinese with English abstract)
MessengerP, van den BoschR(1971) The adaptability of introduced biological control agents. In: Biological Control (ed. Huffaker CB), pp. 68-92. Springer, Boston.
Climatic suitability ranking of candidate biological control agents for weeds: a biogeographic approach for ragweed management in Europe under present and future climatic conditions.
Rapid evolution of an introduced insect Ophraella communa LeSage in new environments: temporal changes and geographical differences in photoperiodic response.
WanFH, LiuWX, MaJ, GuoJ (2005) Ambrosia artemisiifolia and A. trifida. In: Biology and Management of Invasive Alien Species in Agriculture and Forestry (eds Wan FH, Zheng XB, Guo JY), pp. 662-688. Science Press, Beijing. (in Chinese)
Effects of sample size on the performance of species distribution models.
Diversity and Distributions, 14, 763-773.
[51]
YamazakiK, ImaiC, NatuharaY (2000)
Rapid population growth and food-plant exploitation pattern in an exotic leaf beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), in western Japan.
Effects of temperature on survival, development, longevity, and fecundity of Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent against Ambrosia artemisiifolia (Asterales: Asteraceae).
a) Variation in cold hardiness among geographic populations of the ragweed beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), a biological control agent of Ambrosia artemisiifolia L. (Asterales: Asteraceae), in China.
The hot summer of 2010: redrawing the temperature record map of Europe.
1
2011
... 尽管各物种对4种假设气候变化响应不一, 4种未来生物气候情景的平均值还是很好地预测了豚草及其生物防治天敌在东亚分布的重叠区域.气候变化将会使重叠部分进一步减少(当前气候对未来气候情景的重叠比例为42.9% vs. 29.8%).尤其是豚草条纹叶甲与豚草适宜生存区的空间重叠区域占豚草适宜生存区的比例从40.3%下降至29.8%.该结果与对欧洲两种物种分布的预测结果是一致的(Sun et al, 2017).在未来气候变化的情况下, 东亚地区适宜生物防治昆虫地理分布区减少的一个可能的原因是: 未来夏天变得更加炎热, 晚秋的霜冻和温度的季节性增强, 这些将导致豚草向北方和西方进一步扩散, 但是它的两种专食性天敌却并非如此.这进一步证明了入侵植物和它的生物防治天敌对气候变化并不一定有着相似的反应(Schweiger et al, 2008; Barriopedro et al, 2011; Björkman & Niemela, 2015). ...
Climate Change and Insect Pests. CABI, Oxfordshire, UK.
1
2015
... 尽管各物种对4种假设气候变化响应不一, 4种未来生物气候情景的平均值还是很好地预测了豚草及其生物防治天敌在东亚分布的重叠区域.气候变化将会使重叠部分进一步减少(当前气候对未来气候情景的重叠比例为42.9% vs. 29.8%).尤其是豚草条纹叶甲与豚草适宜生存区的空间重叠区域占豚草适宜生存区的比例从40.3%下降至29.8%.该结果与对欧洲两种物种分布的预测结果是一致的(Sun et al, 2017).在未来气候变化的情况下, 东亚地区适宜生物防治昆虫地理分布区减少的一个可能的原因是: 未来夏天变得更加炎热, 晚秋的霜冻和温度的季节性增强, 这些将导致豚草向北方和西方进一步扩散, 但是它的两种专食性天敌却并非如此.这进一步证明了入侵植物和它的生物防治天敌对气候变化并不一定有着相似的反应(Schweiger et al, 2008; Barriopedro et al, 2011; Björkman & Niemela, 2015). ...
Measuring ecological niche overlap from occurrence and spatial environmental data.
3
2012
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
... 使用R软件中ecospat包的排序技术比较了北美原产地和东亚入侵地的豚草生态位(Broennimann et al, 2014).基于豚草分布模型筛选出的9个生物气候变量矩阵, 利用PCA-env的前两个主成分坐标轴对生态位进行量化.在生态位重叠变量的排序中, ecospat已经被证实明显优于其他排序方法(Broennimann et al, 2012; Cola et al, 2016).因此, 我们用Ecospat中的PCA-env来检测生态位的等价性和相似性.对两个区域的分布记录进行随机分配后, 可以通过等价性检验(niche equivalency)来计算其生态位重叠区域是否恒定.生态位相似性(niche similarity)检验了两个地区的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域.该检验同时从北美到东亚和从东亚到北美的两个方向上进行(Broennimann et al, 2012).生态位等价性和相似性检验均使用100次重复. ...
... ).因此, 我们用Ecospat中的PCA-env来检测生态位的等价性和相似性.对两个区域的分布记录进行随机分配后, 可以通过等价性检验(niche equivalency)来计算其生态位重叠区域是否恒定.生态位相似性(niche similarity)检验了两个地区的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域.该检验同时从北美到东亚和从东亚到北美的两个方向上进行(Broennimann et al, 2012).生态位等价性和相似性检验均使用100次重复. ...
Package ‘ecospat’
1
2014
... 使用R软件中ecospat包的排序技术比较了北美原产地和东亚入侵地的豚草生态位(Broennimann et al, 2014).基于豚草分布模型筛选出的9个生物气候变量矩阵, 利用PCA-env的前两个主成分坐标轴对生态位进行量化.在生态位重叠变量的排序中, ecospat已经被证实明显优于其他排序方法(Broennimann et al, 2012; Cola et al, 2016).因此, 我们用Ecospat中的PCA-env来检测生态位的等价性和相似性.对两个区域的分布记录进行随机分配后, 可以通过等价性检验(niche equivalency)来计算其生态位重叠区域是否恒定.生态位相似性(niche similarity)检验了两个地区的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域.该检验同时从北美到东亚和从东亚到北美的两个方向上进行(Broennimann et al, 2012).生态位等价性和相似性检验均使用100次重复. ...
ecospat: an R package to support spatial analyses and modeling of species niches and distributions.
1
2016
... 使用R软件中ecospat包的排序技术比较了北美原产地和东亚入侵地的豚草生态位(Broennimann et al, 2014).基于豚草分布模型筛选出的9个生物气候变量矩阵, 利用PCA-env的前两个主成分坐标轴对生态位进行量化.在生态位重叠变量的排序中, ecospat已经被证实明显优于其他排序方法(Broennimann et al, 2012; Cola et al, 2016).因此, 我们用Ecospat中的PCA-env来检测生态位的等价性和相似性.对两个区域的分布记录进行随机分配后, 可以通过等价性检验(niche equivalency)来计算其生态位重叠区域是否恒定.生态位相似性(niche similarity)检验了两个地区的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域.该检验同时从北美到东亚和从东亚到北美的两个方向上进行(Broennimann et al, 2012).生态位等价性和相似性检验均使用100次重复. ...
Biological flora of the British Isles: Ambrosia artemisiifolia.
2
2015
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
... ; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
Population abundance and host use pattern of Ophraella communa (Coleoptera: Chrysomelidae) in its native and introduced range
1
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
Changes in defense of an alien plant Ambrosia artemisiifolia before and after the invasion of a native specialist enemy Ophraella communa.
1
2012
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
Phylogeny and the evolution of host plant associations in the leaf beetle genus Ophraella (Coleoptera, Chrysomelidae).
1
1990
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
Prospects for biological control of Ambrosia artemisiifolia in Europe: learning from the past.
2
2011
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
... 在中国的西部和北部、韩国和日本北部的大部分豚草分布区域将不再适合它的两种天敌的生存.韩国不适合豚草条纹叶甲的生存; 同时, 豚草卷蛾的适宜生存区也未覆盖到韩国和日本大部分地区.我们可以通过以下两种方法来弥补这一空间覆盖区域上的缺口: (1)探索、发现并引进其他的生物防治天敌(见Gerber et al, 2011); (2)筛选出适应这一空缺区域的豚草条纹叶甲和豚草卷蛾的株系.对此, 生态位分析清楚地表明气候因子可以为我们遴选出适应这些区域的特殊的昆虫天敌株系(比如筛选适应于夏季温度更低和温度季节性增强的品种).所以, 与这些特殊气候适应性状相关的遗传变异显得尤为重要.但是到目前为止, 针对生物防治昆虫的这些功能性状遗传变异的研究仍然十分匮乏(Roderick et al, 2012).近几年, 研究报道了豚草条纹叶甲的一些相关性状的遗传变异, 如飞行行为(Tanaka, 2009)和光周期对环境条件的应答(Tanaka et al, 2015).Zhou等(2013)已经通过低温驯化筛选出了一些适应低温的豚草条纹叶甲株系, 并可能将它们应用于华北地区的豚草生物防治当中.因此, 这一特殊的生物防治天敌株系能够追踪它们的寄主植物到更冷的区域, 并且通过增加世代的方式在这些区域形成更高的种群密度.在大量的商业生产和投放中利用特殊的选择机制可以加快特定天敌株系的培育, 并更好地控制东亚生物防治天敌未覆盖的豚草分布区. ...
Accommodating climate change contingencies in conservation strategy.
1
2013
... 全球气候变化会导致物种分布区迁移(IPCC, 2013), 从而对生态系统的资源动态产生显著的影响.气候变化对入侵植物和生物防治天敌的地理分布影响尤为显著(Theoharides & Dukes, 2007; Walther et al, 2009).气候因子对种间相互作用也有巨大的影响, 进而决定物种分布, 因此对生物防治方案实施的有效性起关键作用(Messenger & Van den Bosch, 1971).植物和节肢动物对气候变化的响应可能会是截然不同的(Hannah et al, 2002; Gillson et al, 2013), 因此, 为了制定可持续的生物防治管理措施, 了解未来气候变化对外来入侵植物与生物防治天敌的影响是至关重要的.生物气候模型能够对当前和未来的气候背景进行深入的考量, 这就为我们提供了一个经济的途径来解决这一未知的难题. ...
Climate and carbon cycle changes from 1850 to 2100 in MPI-ESM simulations for the Coupled Model Intercomparison Project phase 5.
Post-release evaluation of biological control of Bemisia tabaci biotype “B” in the USA and the development of predictive tools to guide introductions for other countries.
1
2005
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
Predictive habitat distribution models in ecology.
1
2000
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
Selecting arthropod biological control agents against arthropod pests: Can the science be improved to decrease the risk of releasing ineffective agents?
1
2005
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
Climate change 2013: The Physical Science Basis.
2
2013
... 全球气候变化会导致物种分布区迁移(IPCC, 2013), 从而对生态系统的资源动态产生显著的影响.气候变化对入侵植物和生物防治天敌的地理分布影响尤为显著(Theoharides & Dukes, 2007; Walther et al, 2009).气候因子对种间相互作用也有巨大的影响, 进而决定物种分布, 因此对生物防治方案实施的有效性起关键作用(Messenger & Van den Bosch, 1971).植物和节肢动物对气候变化的响应可能会是截然不同的(Hannah et al, 2002; Gillson et al, 2013), 因此, 为了制定可持续的生物防治管理措施, 了解未来气候变化对外来入侵植物与生物防治天敌的影响是至关重要的.生物气候模型能够对当前和未来的气候背景进行深入的考量, 这就为我们提供了一个经济的途径来解决这一未知的难题. ...
Climatic niche shifts are rare among terrestrial plant invaders.
1
2012
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
1
2016
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
Evolution and biological control.
2012
Climate change can cause spatial mismatch of trophically interacting species.
1
2008
... 所有模型使用R-3.3.2统计软件进行分析(R Development Core Team, 2016). ...
Biological control of invasive plant species: a reassessment for the Anthropocene.
1
2015
... 在中国的西部和北部、韩国和日本北部的大部分豚草分布区域将不再适合它的两种天敌的生存.韩国不适合豚草条纹叶甲的生存; 同时, 豚草卷蛾的适宜生存区也未覆盖到韩国和日本大部分地区.我们可以通过以下两种方法来弥补这一空间覆盖区域上的缺口: (1)探索、发现并引进其他的生物防治天敌(见Gerber et al, 2011); (2)筛选出适应这一空缺区域的豚草条纹叶甲和豚草卷蛾的株系.对此, 生态位分析清楚地表明气候因子可以为我们遴选出适应这些区域的特殊的昆虫天敌株系(比如筛选适应于夏季温度更低和温度季节性增强的品种).所以, 与这些特殊气候适应性状相关的遗传变异显得尤为重要.但是到目前为止, 针对生物防治昆虫的这些功能性状遗传变异的研究仍然十分匮乏(Roderick et al, 2012).近几年, 研究报道了豚草条纹叶甲的一些相关性状的遗传变异, 如飞行行为(Tanaka, 2009)和光周期对环境条件的应答(Tanaka et al, 2015).Zhou等(2013)已经通过低温驯化筛选出了一些适应低温的豚草条纹叶甲株系, 并可能将它们应用于华北地区的豚草生物防治当中.因此, 这一特殊的生物防治天敌株系能够追踪它们的寄主植物到更冷的区域, 并且通过增加世代的方式在这些区域形成更高的种群密度.在大量的商业生产和投放中利用特殊的选择机制可以加快特定天敌株系的培育, 并更好地控制东亚生物防治天敌未覆盖的豚草分布区. ...
Climatic suitability ranking of candidate biological control agents for weeds: a biogeographic approach for ragweed management in Europe under present and future climatic conditions.
1
2017
... 尽管各物种对4种假设气候变化响应不一, 4种未来生物气候情景的平均值还是很好地预测了豚草及其生物防治天敌在东亚分布的重叠区域.气候变化将会使重叠部分进一步减少(当前气候对未来气候情景的重叠比例为42.9% vs. 29.8%).尤其是豚草条纹叶甲与豚草适宜生存区的空间重叠区域占豚草适宜生存区的比例从40.3%下降至29.8%.该结果与对欧洲两种物种分布的预测结果是一致的(Sun et al, 2017).在未来气候变化的情况下, 东亚地区适宜生物防治昆虫地理分布区减少的一个可能的原因是: 未来夏天变得更加炎热, 晚秋的霜冻和温度的季节性增强, 这些将导致豚草向北方和西方进一步扩散, 但是它的两种专食性天敌却并非如此.这进一步证明了入侵植物和它的生物防治天敌对气候变化并不一定有着相似的反应(Schweiger et al, 2008; Barriopedro et al, 2011; Björkman & Niemela, 2015). ...
Genetic variation in flight activity of Ophraella communa (Coleoptera: Chrysomelidae): heritability estimated by artificial selection.
Rapid evolution of an introduced insect Ophraella communa LeSage in new environments: temporal changes and geographical differences in photoperiodic response.
5
2015
... 物种分布模型(species distribution models, SDMs)是利用已知的物种分布位点数据与环境因子研究物种与环境间的关系.环境因子是决定物种生存和繁殖的重要因素之一, 通过物种分布模型可以完成对物种潜在地理分布区的预测(Guisan & Zimmermann, 2000).此外, 物种分布模型也是预测物种分布对气候变化响应的主要方法(Peterson, 2011).近几年, 物种分布模型越来越多地用于预测在当前和未来气候背景下入侵植物的地理分布并识别高危生境(Peterson, 2003).此外, 物种分布模型也可以有效地预测入侵植物分布区内生物防治昆虫的最佳生存区域(Sun et al, 2017).由于生物物理学因素, 昆虫种群受气候因子的影响非常显著(Hoelmer & Kirk, 2005), 所以, 物种分布模型还能够帮助我们识别出生物防治昆虫在原产地的最佳采集地和在引入区域的最佳释放点(Mukherjee et al, 2011).已有研究发现, 生物防治昆虫的引入区域与原产地的气候越匹配, 它们就越容易在引入区域建立种群并更加有效地控制入侵植物(Goolsby et al, 2005).因排序技术能够直接在环境空间内比较物种与环境因子之间的关系, 利用该技术可以从物种分布位点数据推断相应的生物防治效果(Broennimann et al, 2012). ...
... 全球气候变化会导致物种分布区迁移(IPCC, 2013), 从而对生态系统的资源动态产生显著的影响.气候变化对入侵植物和生物防治天敌的地理分布影响尤为显著(Theoharides & Dukes, 2007; Walther et al, 2009).气候因子对种间相互作用也有巨大的影响, 进而决定物种分布, 因此对生物防治方案实施的有效性起关键作用(Messenger & Van den Bosch, 1971).植物和节肢动物对气候变化的响应可能会是截然不同的(Hannah et al, 2002; Gillson et al, 2013), 因此, 为了制定可持续的生物防治管理措施, 了解未来气候变化对外来入侵植物与生物防治天敌的影响是至关重要的.生物气候模型能够对当前和未来的气候背景进行深入的考量, 这就为我们提供了一个经济的途径来解决这一未知的难题. ...
1
1993
... 全球气候变化会导致物种分布区迁移(IPCC, 2013), 从而对生态系统的资源动态产生显著的影响.气候变化对入侵植物和生物防治天敌的地理分布影响尤为显著(Theoharides & Dukes, 2007; Walther et al, 2009).气候因子对种间相互作用也有巨大的影响, 进而决定物种分布, 因此对生物防治方案实施的有效性起关键作用(Messenger & Van den Bosch, 1971).植物和节肢动物对气候变化的响应可能会是截然不同的(Hannah et al, 2002; Gillson et al, 2013), 因此, 为了制定可持续的生物防治管理措施, 了解未来气候变化对外来入侵植物与生物防治天敌的影响是至关重要的.生物气候模型能够对当前和未来的气候背景进行深入的考量, 这就为我们提供了一个经济的途径来解决这一未知的难题. ...
Biological Control of Weeds: A World Catalogue of Agents and Their Target Weeds, 5th edn
3
2014
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
... ), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
... 豚草卷蛾是一种采食植物茎部、由虫瘿孵化而来的蛾类.它将纺锤形虫瘿产于成熟豚草属植物的侧枝上, 适宜条件下每年能够繁殖3-4个世代(Ma et al, 2008).在中国的华北地区一年可繁衍2-3个世代, 华中地区4-5个世代(Zhou et al, 2014).1982年, 豚草卷蛾被从墨西哥引入到澳大利亚来控制银胶菊(Parthenium hysterophorus), 但是随后它广泛分布在豚草植株上(McFadyen, 1992).1990年, 我国从澳大利亚引入豚草卷蛾来控制豚草, 经过宿主专一性检测后, 于1993年释放到野外, 随后豚草卷蛾广泛地分布于华南地区(Wan et al, 2005). ...
Effects of sample size on the performance of species distribution models.
2008
Rapid population growth and food-plant exploitation pattern in an exotic leaf beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), in western Japan.
2
2000
... 豚草(Ambrosia artemisiifolia)是一年生风媒传粉、雌雄同株的草本植物, 原产北美和墨西哥.它被无意地引入全球的多个地区, 包括亚洲、南美洲、大洋洲和欧洲.它的花粉具有致敏性, 其植株作为杂草在农田中极难控制; 所以不管是在原产地还是入侵地, 豚草都造成了极大的危害(Wan et al, 2005; Essl et al, 2015).中国的豚草分布区域内, 2-3%的人对其花粉有过敏症状, 这也直接导致了每年14亿元人民币的损失(Zhou et al, 2015).豚草入侵日本已经超过100年(Fukano & Yahara, 2012).20世纪30年代, 在中国首次发现了豚草的分布记录, 直至1997年才被认为是有害于多种农作物的杂草(Wan et al, 1993).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
... ).在中国(Wan et al, 1993)和日本(Hisauchi, 1950), 豚草除了分布在农田以外, 还广泛地分布在交通运输线上(比如铁路、公路和河流沿线).豚草经过收割和除草剂局部控制以后, 其侧枝的生长能力非常强, 植株的生长速度也非常快, 这对管理和控制是一个极大的挑战(Wan et al, 2005; Essl et al, 2015).在农田中, 除草剂对豚草生物量的控制是有效的, 但是没法有效地遏制其开花、传粉和结果(Gerber et al, 2011). ...
Control of the invasive weed Ambrosia artemisiifolia with Ophraella communa and Epiblema str¬enuana.
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
Effects of temperature on survival, development, longevity, and fecundity of Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent against Ambrosia artemisiifolia (Asterales: Asteraceae).
a) Variation in cold hardiness among geographic populations of the ragweed beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), a biological control agent of Ambrosia artemisiifolia L. (Asterales: Asteraceae), in China.
3
2011
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
... ).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
... ).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
Review on management of Ambrosia artemisiifolia using natural enemy insects.
4
2015
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
... ; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
... 豚草卷蛾是一种采食植物茎部、由虫瘿孵化而来的蛾类.它将纺锤形虫瘿产于成熟豚草属植物的侧枝上, 适宜条件下每年能够繁殖3-4个世代(Ma et al, 2008).在中国的华北地区一年可繁衍2-3个世代, 华中地区4-5个世代(Zhou et al, 2014).1982年, 豚草卷蛾被从墨西哥引入到澳大利亚来控制银胶菊(Parthenium hysterophorus), 但是随后它广泛分布在豚草植株上(McFadyen, 1992).1990年, 我国从澳大利亚引入豚草卷蛾来控制豚草, 经过宿主专一性检测后, 于1993年释放到野外, 随后豚草卷蛾广泛地分布于华南地区(Wan et al, 2005). ...
... 1993年, 两种专食性生物防治昆虫在湖南岳阳第一次大规模释放.1999年, 在湖南已控制了超过2,000 km2的豚草分布区.随后, 这两种专食性天敌扩散到湖北省和江西省(Li et al, 1999).在华南地区, 豚草条纹叶甲和豚草卷蛾共存于许多豚草分布区, 并被建议同时使用以更有效地控制豚草入侵(Zhou et al, 2014). ...
b) Reevaluation of biosecurity of Ophraella communa against sunflower (Helianthus annuus).
2
2011
... 经典的生物防治手段是可持续控制豚草的最佳策略.在中国, 利用豚草原产地的生物天敌豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)对豚草进行生物防治已经被证明是最有效的方式(Zhou et al, 2011b).两种天敌昆虫都原产北美.豚草条纹叶甲是一种偏爱豚草作为寄主植物和食物来源的寡食性叶甲(LeSage, 1986; Futuyma & McCafferty, 1990; Palmer & Goeden, 1991).在东亚地区, 于1996年在日本东部的千叶县首次被发现(偶然引入), 此后迅速扩散到日本全境(Yamazaki et al, 2000; Moriya & Shiyake, 2001).1999年, 豚草条纹叶甲第一次在中国发现(Zhou et al, 2014).良好的环境条件下, 豚草条纹叶甲每年可以繁殖4-7代, 这使它能够在豚草生长季的后半段形成局部高密度种群.不论是在实验条件还是野外条件下, 这种叶甲都能够导致豚草的叶片完全脱落和死亡(Palmer & Goeden, 1991; Zhou et al, 2014).在中国和日本, 豚草条纹叶甲均被认为是目前控制豚草最有效、最成功的生物防治手段(Yamazaki et al, 2000; Fukano & Doi, 2013).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
... ).豚草条纹叶甲主要分布在亚热带地区(Zhou et al, 2010).近几年, 中国学者尝试选出一些优良的豚草条纹叶甲品种, 使它们能够在高纬度地区生存并繁殖后代(Zhou et al, 2011a, 2013).但截至目前, 日本和韩国还没有进行人为释放豚草条纹叶甲来防治豚草的实验(Winston et al, 2014). ...
Cold temperatures increase cold hardiness in the next generation Ophraella communa beetles.
 , 周忠实
, 周忠实

{kind=link}
{kind=link}
{kind=link}
{kind=link}