洪德元
中国科学院植物研究所系统与进化植物学国家重点实验室, 北京 100093
De-Yuan Hong
通讯作者:
版权声明: 2016 《生物多样性》编辑部 版权所有.《生物多样性》编辑部
基金资助:
展开
摘要
物种概念(species concept)是生物学家们持续关注的中心问题。物种概念决定物种划分, 而物种划分的合理性关系到生物多样性的研究、保护和可持续利用。本文把现有较流行的物种概念分为6类, 并对它们予以述评后指出: 虽然生物学物种概念、遗传学物种概念、进化物种概念、系统发生物种概念等从不同方面认识了物种的客观真实性和物种的本质, 但在实践中都难以操作。绝大多数物种是由分类学家划分的, 但目前所有的分类学物种概念都包含有不同程度的主观因素, 从而造成物种划分的人为性, 对生物多样性研究造成负面影响。因此, 生物多样性事业需要科学、可操作的物种概念。本文在吸收了生物学物种概念、遗传学物种概念、进化物种概念以及系统发生物种概念等的长处, 也分析了它们的不足和问题的基础上提出一个新的物种概念, 即形态-生物学物种概念。最后, 以芍药属(Paeonia)几个物种的处理为例, 说明这一新的物种概念是可操作的, 划分的物种在形态上区别分明, 易于鉴别。更重要的是, 其结果得到基于25或26个单拷贝或寡拷贝核基因DNA序列所作的系统发生分析的强有力支持。各个物种在系统发生树上形成单系和独立的谱系, 表明其间各自形成独立的基因库, 没有基因交换, 它们独立进化, 有各自的生态位和独立的分布区。因此, 利用这一新的物种概念能够达到预期目标。
关键词:
Abstract
The future of human being may rely on biodiversity, and thus depends on how to investigate, conserve, and rationally use biodiversity. Species is the basic unit of biodiversity, and therefore rational delimitation of species is one of the crucial issues for biodiversity pursuits. However, no species concept published until now is both scientific and operative. A tentative species concept is proposed here just for discussion.
Keywords:
生物多样性与人类息息相关。过去知道, 人类的衣食住行离不开生物, 现在更知道, 人类未来的居住环境也离不开生物多样性。人类未来生活的质量要看人类如何对待生物多样性, 那就是深入研究它、保护它、可持续地利用它。
生物多样性由3个层次组成, 即生态系统多样性、物种多样性和遗传多样性。其中物种起着承上启下的作用, 向上组成生态系统, 向下是遗传多样性的载体。如果对物种多样性认识不到位就不可能真正认识生物多样性, 深入研究、有效保护和可持续利用就很可能是一句空话。
我数十年的研究工作都涉及物种多样性: 编写植物志, 作分类修订, 写类群专著, 主持有关生物多样性项目。因此我深深认识到, 物种的合理划分对于正确认识生物多样性和编撰植物志的水平和质量是何等重要, 以及物种划分的偏差对生物多样性认识造成了多大的误导。物种划分问题在国际上有以《苏联植物志》(Flora USSR)为代表的苏联学派和以《欧洲植物志》(Flora Europaea)为代表的欧美学派, 他们划分物种的结果有很大差别。中国这方面的问题更加突出。《中国植物志》和Flora of China 的完成, 在得到国际上赞扬的同时, 却也很有一些外国学者认为中国的某些植物分类学家是“分裂派”(splitters), 意指有乱造“新种”之嫌。在国内, 也有不少人在肯定“两志”重大贡献的同时, 抱怨许多类群的检索表没法使用(检索表是否好用是鉴别物种划分是否自然的试金石)。一些植物类群的物种数目在不同的分类处理中差异巨大。这种状况不仅损害生物多样性研究、保护和可持续利用的效果, 也影响生物多样性评估和地区之间比较的科学性。因此, 合理划分物种, 作出可检验和实用的分类处理是我追求的目标。本文集我一辈子努力实践和读书结果, 提出一个新的物种概念, 供大家讨论。
本文选择用中文发表, 是想先让国内的同事们对新提的物种概念加以讨论。文中凡我点评的物种概念都引出了原文并作了翻译, 是考虑到过去国内很少讨论物种概念问题, 许多有关译文不够准确, 甚至很不准确。希望今后有更多的学者参与物种概念的讨论。
物种问题持续成为生物学家和哲学家关注的中心(Mishler & Brandon, 1987)。著名植物学家Davis和Heywood (1963)在他们的名著Principles of Angiosperm Taxonomy中说: “物种不管从哪方面看都是分类学的基本单元”。
两部划时代的生物学著作都以“物种”(species)作标题。瑞典人林奈(1753)的《植物种志》(Species Plantarum)标志着现代生物学的诞生。英国人达尔文(1859)的《物种起源》(The Origin of Species)更是为人类开创了进化论时代。由此可见, 物种问题一直是生物学中的一个重点话题。
Simpson (1951)明确提出物种是进化的基本单位(a basic unit in evolution)。
物种是生物多样性的基本单元, 保护国际(Conservation International)确定的世界上34个生物多样性热点地区就是以物种数目和特有物种数目为依据的; 一个国家或地区生物多样性的丰富度和规模亦是以物种数目衡量的。如果没有一个客观的、可操作的物种概念, 物种划分各行其是, 那么生物多样性评估、地区之间的比较、热点地区的确定就难有客观性、合理性。
生物多样性保护是人类的伟大事业。完成这件大事首先要通过深入研究来确定保护对象, 也即保护单元。下面我们以牡丹为例, 说明只有科学地确定保护单元, 生物多样性保护才能达到预期效果。
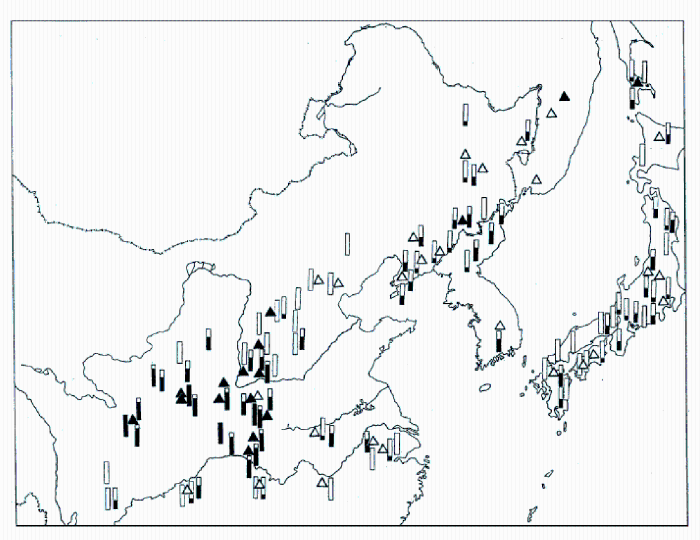
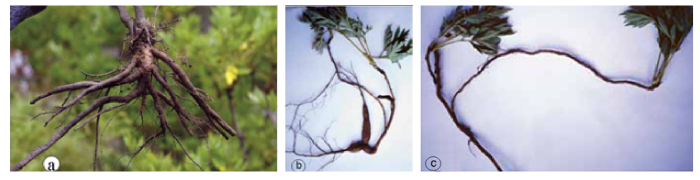
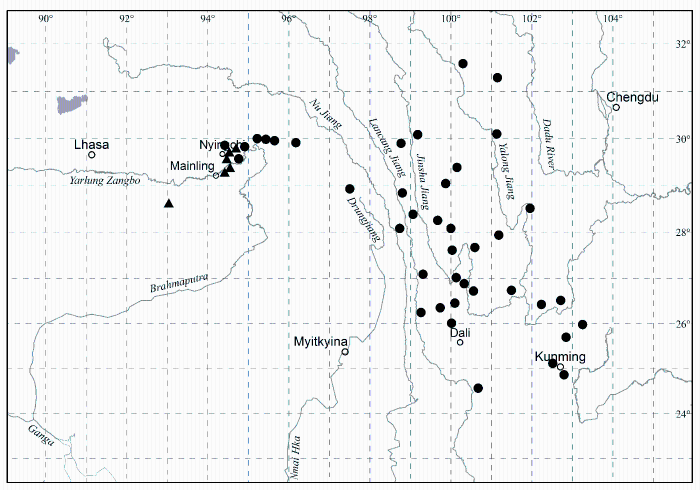
黄牡丹(Paeonia delavayi Franch. var. lutea (Franch.) Finet et Gagnep.)是《中国植物红皮书》(傅立国和金鉴明, 1992)收录的354个保护单元中的一个。书中描述黄牡丹花黄色, 分布于云南中部、西北部和西南部, 四川西南部的木里等县, 以及西藏东南部的波密、林芝、工布江达和隆子(图1), 相当广布。但书中所谓的“黄牡丹”其实是滇牡丹复合群(Paeonia delavayi Franch. complex)中的黄花类型。这一复合群的花色极其多变, 从深紫色、红色、橙色、黄色、黄中带紫或橙色斑块, 直至白色, 甚至在同一居群(population一词的中文翻译除人口外还有4种: 群体(主要用于遗传学和进化研究)、居群(主要用于分类学和进化研究)、种群(主要用于生态学)和族群(台湾学者)。)中都可以看到这种花色变异, 只不过黄色是其中最常见的一种花色。因此, 以花色划分物种和变种都是不科学的(Hong et al, 1998)。事实上, 西藏东南部的“黄牡丹”(当地称大花黄牡丹)与其他广大地区的“黄牡丹”区别分明。它虽然也是花黄色, 但它的根像灌木, 茎丛生(图2), 心皮单一, 蒴果大而长, 而其他地区的“黄牡丹”根白薯状, 茎散生(图2), 心皮2-5, 果小。因此, 它们是两个独立的物种, 即大花黄牡丹(Paeonia ludlowii)和滇牡丹(P. delavayi)(详见下文案例5)。滇牡丹由于具有块根、地下走茎、散生的习性以及相当广泛的分布, 并不是濒危物种; 而大花黄牡丹丛生习性, 全靠种子繁殖, 更由于它局部分布, 仅有5个居群, 才是真正的濒危物种, 但被红皮书掩埋在并不濒危的“黄牡丹”中。由于乱采滥挖(见图3)和建设林芝机场(在米林), 其有限的居群遭受到近乎毁灭性的破坏。而这种牡丹已于1936年被英国采集家引至英国, 在辉煌的伦敦自然博物馆大门两侧花坛里健壮生长(图4)。如再不采取有效的保护措施, 很有可能重演麋鹿(El- aphurus davidianus)重引入那使中国人尴尬的一幕。
此案例的教训很深刻: 保护单元的误定会错把普通物种作为濒危物种来保护, 劳而无功; 而真正的濒危物种却被淹没, 惨遭破坏!可见, 科学、合理的物种划分, 保证保护单元确定的科学性是何等重要!
图3 西藏东南部的“黄牡丹”[其实是大花黄牡丹Paeonia ludlowii (Stern & G. Taylor) D. Y. Hong]的遭遇(洪德元摄自西藏米林)
Mayden (1997)列举了22个物种概念, 5年后他增加为24个(Mayden, 2002); Wilkins (2006, 2009)列举了26个(表1); 我国学者周长发和杨光(2011)列出了68个。
表1 Wilkins (2006, 2009)列举的26个物种概念
| 物种概念 Species concept | |
|---|---|
| 1 | 无性繁殖物种 Agamospecies |
| 2 | 自衍征物种 Autapomorphic species |
| 3 | 生物学物种 Biospecies |
| 4 | 支序物种 Cladospecies |
| 5 | 内聚物种 Cohesion species |
| 6 | 掠夺物种 Compilospecies |
| 7 | 复合物种 Composite species |
| 8 | 生态学物种 Ecospecies |
| 9 | 进化物种 Evolutionary species |
| 10 | 进化显著单元 Evolutionary significant unit |
| 11 | 血缘和谐物种 Genealogical concordance species |
| 12 | 基因物种 Genic species |
| 13 | 遗传学物种 Genetic species |
| 14 | 基因型簇 Genotypic cluster |
| 15 | 亨氏物种 Hennigian species |
| 16 | 节点间物种 Internodal species |
| 17 | 涵盖范围最小分类单元 Least inclusive taxonomic unit (LITUS) |
| 18 | 形态学物种 Morphospecies |
| 19 | 无维物种 Non-dimensional species |
| 20 | 假物种 Nothospecies |
| 21 | 系统发生分类群物种 Phylogenetic taxon species |
| 22 | 表征物种 Phenospecies |
| 23 | 识别物种 Recognition species |
| 24 | 生殖竞争物种 Reproductive competition species |
| 25 | 接替物种 Successional (successive) species |
| 26 | 分类学物种 Taxonomic species |
Wilkins (2006)把所有物种概念归为7类: (1)无性繁殖物种概念; (2)生物学物种概念; (3)生态学物种概念; (4)进化物种概念; (5)遗传学物种概念; (6)形态学物种概念; (7)分类学物种概念。
但是我认为归成以下6类更合适, 即: (1)强调生殖隔离的物种概念, 包括生物学物种概念、内聚物种概念、识别物种概念、生殖竞争物种概念、无维物种概念等; (2)强调基因差异的物种概念, 包括遗传学物种概念、基因物种概念和基因型簇物种概念等; (3)强调进化的物种概念, 包括进化物种概念、进化显著单元、接替物种概念等; (4)强调系统发生或支序分类理论的物种概念, 包括系统发生物种概念、谱系物种概念、支序物种概念、亨氏物种概念、节点间物种概念、自衍征物种概念、涵盖范围最小分类单元等; (5)强调生态的物种概念, 其代表就是生态学物种概念; (6)强调外部形态的物种概念, 包括形态学物种概念、分类学物种概念、形态-地理学物种概念、表征物种概念等。
2.3.1 强调生殖隔离的物种概念
这类物种概念最具代表性的是生物学物种概念(biological species concept)。它由Dobzhansky于1935年提出, 经多次修改, 1977年在他离世两年后由他和他的同事共同发表了最后版本(Dobzhansky et al, 1977): “在有性生殖的生物中, 物种可定义为孟德尔居群或孟德尔居群的集合体, 它们之间的基因交换因有生殖隔离机制而受限或受阻”(In sexually reproducing organisms species can be defined as Mendelian populations, or arrays of Mendelian populations, between which the gene exchange is limited or prevented by reproductive isolating mechanisms.)。
生物学物种概念以Mayr (1942)的版本最为流行: “物种是一群实际上或潜在地异交繁育的自然居群, 与其他这样的居群存在生殖隔离”(Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups.)。有两个例子最能解释生物学物种概念的内涵。其一, 马和驴能杂交, 但产生的杂种骡是没有生育能力的, 因此马和驴之间存在生殖隔离, 故它们是两个物种。其二, 鹅掌楸(Liriodendron chinense (Hemsl.) Sarg.)分布于中国东部和越南北部, 北美鹅掌楸(L. tulipifera L.)分布于美国东南部, 两者不仅跨太平洋间断分布, 而且形态上也有明显区别, 一直被认为是两个物种, 但杂交试验表明, 它们的后代不仅能育, 而且生长强壮, 说明两者之间不存在生殖隔离。按照生物学物种概念, 它们应是一个物种。
Bock (1986, 2004)提出了最新的生物学物种概念: “物种是一群实际或潜在异交繁育的居群, 它们在自然界中与其他这样的群存在遗传隔离”(A species is a group of actually or potentially interbreeding populations which are genetically isolated in nature from other such groups.)。对于这一物种概念, 我有两点看法: (1)它把生物学物种概念和遗传学物种概念融合在一起, 只是把Mayr (1942)的“reproductively”换成了“genetically”。我赞成这一改换, “遗传隔离”比“生殖隔离”涵义更宽, 更有生物学意义; (2)既然强调“在自然界”, 那么如何知道居群之间是具有潜在的异交繁育能力, 还是生殖隔离的?
识别物种概念(recognition species concept)的代表人物是Paterson (1985, 1992), 他认为: “因此, 我们可以把物种看成是涵盖所有共享一个受精系统的双亲个体的居群”(We can, therefore, regard as a species that the most inclusive population of individual biparent organisms which share a common fertilization system.)。
总之, 这一类物种概念强调的是物种内所有成员共享一个基因库, 可进行基因交换, 但这个基因库对每个物种来说是独有的, 与别的物种没有基因交换。这是物种形成、生物多样性分化的关键步骤。这类物种概念的提出在理解物种本质、物种形成和生物多样性形成机制上是一大进步, 因此是至今最广泛接受但同时也是受到最多批评的物种概念。周长发和杨光(2011)列举了多达20条批评意见。就连Mayr本人也认识到应用生物学物种概念的难点多达6个(Mayr, 1992)。van Valen (1992)更是直截了当地说, Mayr的概念美其名曰“生物学物种概念”, 其实应该称之为“生殖物种概念”(reproductive species concept)。
对这类物种概念我仅简述5点: (1)只适用于两性生殖的生物。(2)往往难于操作。例如, 对于植物界的松柏类和动物界的龟鳖类等, 实验者能等着看到其F1的后代活得怎样、是否能育吗?对海洋中的鲸类能做杂交实验吗?(3)造成生殖隔离的原因复杂, 有的物种之间仅由1个基因决定是否存在生殖隔离。(4)生殖隔离也有程度问题, 绝不是“是”与“否”、“0”与“100”这么简单(Ge & Hong, 1994)。Mishler和Donoghue (1982)也指出了这一问题。(5)生殖隔离不是物种本质的全部, 有些其他方面明显不同、且广为承认的物种却无生殖隔离, 在禾本科小麦族和兰科中均存在这种情况。
2.3.2 强调基因差异的物种概念
包括遗传学物种概念、基因物种概念、基因差异物种概念和基因型簇物种概念, 这里介绍前两个。
(1)遗传学物种概念(genetic species concept)。首先由Simpson (1943)提出: “遗传学物种是这样一群生物个体, 它们在自然界中的组成和所处状况决定它们中任何一个个体的遗传性状都有可能(但不一定)传递给其他任何一个个体的后代”(A genetic species is a group of organisms so constituted and so situated in nature that a hereditary character of any one of these organisms may be (possibly, but not necessarily) transmitted to a descendant of any other.)。有些学者提出的遗传学物种概念则几乎是生物学物种概念的翻版, 如Baker和Bradley (2006): “遗传学物种是一群遗传上相融、互交繁育的自然居群, 它们与其他这样的群在遗传上隔离”(A genetic species is a group of genetically compatible interbreeding natural populations that is genetically isolated from other such groups.)。
(2)基因物种概念(genic species concept)。由中国学者吴仲义首次提出(Wu, 2001): “物种是适应上已经分化的类群, 即使接触也不能通过直接交换或经由中间的杂种居群共享控制这些适应性状的基因。这些类群在基因组别的位置上可能已经分化, 也可能还没有分化”(Species are groups that are differentially adapted and, upon contact, are not able to share genes controlling these adaptive characters, by direct exchange or through intermediate hybrid populations. These groups may or may not be differentiated elsewhere in the genome.)。
遗传学物种概念本质上与生物学物种概念没有显著区别, 而吴仲义的基因物种概念强调适应性基因群。它们从基因和基因组角度丰富了生物学物种概念, 但有同样的问题: 适应上分化到何种程度才是不同物种?如何快速鉴定“控制这些适应性状的基因”(genes controlling these adaptive characters)?目前恐怕还只能在个别生物中才有可能实现。总的看来, 基因物种概念也不比生物学物种概念更容易操作。
2.3.3 强调进化的物种概念
有3个物种概念可归入这一类: 进化物种概念(evolutionary species concept)、接替物种概念(successive species concept)和时间物种概念(chronospecies or chronological species concept)。这里介绍前两个。
(1)进化物种概念。这一概念首次由Simpson (1951)不明确地提出, 而于1961年(Simpson, 1961)正式提出: “进化物种是一个谱系(居群的祖-裔序列), 与其他谱系分别进化, 而且有独自的进化作用和趋势”(An evolutionary species is a lineage (an ancestral-descendent sequence of populations) evolving separately from others and with its own unitary evolutionary role and tendencies.)。Wiley (1992)作了修正: “物种是由祖-裔生物居群组成的单个谱系, 它与其他这样的谱系保持独立, 并有自身的进化趋势和历史命运”(A species is a single lineage of ancestral descendent populations of organisms which maintains its identity from other such lineages and which has its own evolutionary tendencies and historical fate.)。Grant (1971)也认为进化物种是一个居群系统, 具4个特点: (a)是一个谱系, 即居群在时空上的祖-裔顺序; (b)与其他谱系分别进化; (c)有独有的进化作用, 即占据适合自己的生态位; (d)有自身的进化趋势, 在历史进程中会发生进化作用的变化。
(2)接替物种概念(successive/successional species)。这是由Simpson (1961)首先提出的, 即一个谱系中时间短暂的一段。古生物学物种概念(paleo- species concept)也是由Simpson同年提出: “古生物物种即单个谱系中短暂接替的物种”(palaeospecies: temporally successive species in a single lineage)。Wilkins (2006)给出了他自己的版本: “人为划分的形态类型向前进化的阶段, 主要体现在古生物记录中”(Arbitrary anagenetic stages in morphological forms, mainly in the paleontological record.)。可见, 接替物种概念和古生物学物种概念实质上是统 一的。
这些物种概念强调物种是一个谱系, 在进化上具独立性, 这在理论上有助于更全面地认识物种。但如何确定谱系的组成和边界, 他们没有具体说明, 如何划分物种也没有给分类学家提供指导或参考意见。
2.3.4 强调系统发生或支序分类理论的物种概念
这一类物种概念很庞杂, 至少包括8个, 这里仅评述较流行的3个。
(1)系统发生物种概念(phylogenetic species concept)。常被引用的版本是Cracraft (1983)提出的, “物种是可鉴别的最小生物个体群, 群内存在祖-裔传承关系”(A species is the smallest diagnosable cluster of individual organisms within which there is a parental pattern of ancestry and descent.)。Mishler和Brandon (1987) 提出: “物种是分类中识别出的涵盖范围最小的分类群, 归入其中的是有单系证据(即通常但不限于存在共有衍征)的生物个体……”(A species is the least inclusive taxon recognized in a classification, into which organisms are grouped because of evidence of monophyly (usually, but not restricted to, the presence of synapomorphies), …。
(2)支序物种概念(cladistic species concept) = 亨氏物种概念(Hennigian species concept)。Ridley (1989)解释Hennig 1966年的支序物种概念是: “物种是两次物种形成事件中间, 或一次物种形成事件和一次物种灭绝事件之间, 或由一次物种形成事件传下来的那一群生物”(A species is then that set of organisms between two speciation events, or between one speciation event and one extinction event, or that are descended from a speciation event.)。Ridley本人则把这一物种概念缩简为: “支序物种是两次物种形成事件之间的生物个体群……”(a cladistic species is the set of organisms between two speciation events… (Ridley, 1989)。
(3)节点间物种概念(internodal species concept)。由Kornet (1993)提出: “亲缘网络中两次永久分裂事件之间或一次永久分裂事件和一次灭绝事件之间的那部分个体因为是共同成员而同属一个物种”(Individual organisms are conspecific in virture of their common membership of a part of genealogical network between two permanent splitting events or between a permanent splitting event and an extinction event.)。
系统发生和支序理论物种概念的流行程度仅次于生物学物种概念。它们的最大特点是强调单系、共有衍征、祖-裔谱系关系和物种形成的节点事件, 对于认识系统发生、进化和物种形成以及物种的本质都很有积极作用。系统发生物种概念是除分类学物种概念以外最接近实际运用的。Mishler和Brandon (1987)说他们讨论物种问题的目的是为系统学的基本单元寻找一个有实用价值的定义, 提出最理想的、普适的分类, 并反映当前关于进化过程的知识。他们用共有衍征来确定单系无疑是一大进步。但遗憾的是, 他们未提确定物种一级的单系应该用多少个共有衍征, 1个, 2个, 还是更多?我认为仅用1个共有衍征确定的单系很可能是生态型(ecotype)或亚种, 甚至是1个基因型, 但不太可能是物种。他们还提到, 确定单系“通常但不限于共有衍征”。那么, 如果不用共有衍征, 又如何来确定单系呢?
2.3.5 强调生态的物种概念
其代表就是生态学物种概念(ecological species concept), 首先由van Valen (1976)提出: “物种是一个或一组亲缘紧密的谱系, 与其分布范围内的任何其他谱系都在各自占据的适应带上有微小差异, 而与分布范围外的所有谱系都是分别进化的”(A species is a lineage (or a closely related set of lineages) which occupies an adaptive zone minimally different from that of any other lineage in its range and which evolves separately from all lineages outside its range.)。这一概念涉及谱系、适应带(adaptive zone)、分布范围、分别进化等, 强调分布范围内不同谱系之间在适应带方面有微小差异, 但内容抽象。谱系如何限定?如何确定“微小差异”? 如何知道“分别进化”? 这一物种概念除了让人认识到同一分布范围内的物种有生态位(niche)分化外, 对物种的认识和划分并无实质贡献。而且“谱系”和“亲缘紧密的谱系”是不能等同的, 这一提法与进化物种概念和系统发生物种概念并不吻合。谱系之间即使亲缘关系紧密也是没有基因交流的, 应属于不同的物种。
2.3.6 强调形态的物种概念
包括形态学物种概念(morphological species concept)、分类学物种概念(taxonomic species concept)、形态-地理学物种概念(morpho-geographical species concept)、表征物种概念(phenetic species concept)等。Simpson (1943)把形态学物种概念和分类学物种概念分开(见后文), 但Davis和Heywood (1963)认为应把涉及形态的物种概念统归于分类学物种概念之下, 因为它们都为分类学家所用。已有很多学者提出了分类学物种概念, 这里仅介绍几个比较流行的。
林奈(1758)认为: “物种是一群有着相同或相似形态性状的个体或居群”(A species is a group of individuals or populations with the same or similar morphological characters.) (Mayr, 1942)。尽管林奈划分的物种大多被后来人承认为自然的, 称为“林奈种”, 但不是所有后来人都能分清什么算“相同”, 什么算“相似”, 所以林奈的概念模糊了物种、亚种和属之间的界限(Mayr, 1942)。
达尔文为了证明物种是进化的产物, 不强调物种和变种之间的界限, 反而强调两者之间的过渡。所以他说: “物种是由有能力的系统学家(更确切地说是那个类群的专家)认定的系统学的一个单元”(A species is a systematic unit which is considered a species by a competent systematist (preferably a specialist of the group).)。又说: “在决定一个生物类型究竟应定为物种还是变种时, 似应依从判断力强、经验丰富的博物学家的意见”(In determining whether a form should be ranked as a species or a variety, the opinion of naturalists having sound judgment and wide experience seems the only guide to follow.)。很遗憾, 伟大的达尔文为分类学物种概念附上了人为成分, 这对后人有很大影响。
Du Rietz (1930)把物种定义为“所谓物种就是在一系列生物型上间断分明而彼此永久分开的最小自然居群”(The smallest natural populations permanently separated from each other by a distinct discontinuity in the series of biotypes, are called species.)。他接着补充说: “因此物种是一个居群, 它或者由一个严格的无性而有生命力的生物型组成, 或由一群实际上不可分辨的、严格的无性而有生命力的生物型组成, 或由许多有性生殖的、形成异交群①的生物型组成, 而异交群之间因完全或接近完全的性隔离或因相对小的过渡居群而分开”(A species thus is a population consisting either of one strictly asexual and vital biotype, or of a group of practically undistinguishable, strictly asexual and vital biotypes, or of many sexually propagating biotypes forming a syngameon(syngameon (异交群) = intercrossing population) separated from all other by more or less complete sexual isolation or by comparatively small transitional populations.)。就我所知, 在物种概念中出现“明显不连续”(distinct discontinuity)这一字眼还是第一次, 这是向科学的物种概念走近了一步, 但如何精准掌握“在一系列生物型上间断分明”(a distinct discontinuity in the series of biotypes)?多少个“生物型”(biotypes)? 恐怕生物学家不会有统一意见。他自己补充说“一个”、“一群”, 或“许多”。但即便这样, 分类学家仍然不能精确地掌握, 而且他的补充使他的定义更加抽象, 如何准确理解他的“完全或接近完全的有性隔离”(more or less complete sexual isolation)和“比较小的过渡居群” (comparatively small transitional populations)?
瑞典人Hedberg (1958)赞同Stebbins (1950)的见解, 即在有性生殖的生物中划分物种的标准是“不同物种的成员之间没有或罕有基因交换”(absence or rarity of gene interchange between members of different species)。Hedberg意识到直接得到这样的信息常常是不可能的, 用实验的方法则又费时、费力、费钱, 因而不是可以普遍进行的。于是他以间接的办法, 即用形态性状变异的不连续性来代之。他在处理东非高山植物的替代现象时采用的办法就是(Hedberg, 1958): “当两个这样的居群仅在一个性状上呈现完全不连续变异时(常伴有其他性状上的部分不连续), 就把它们区分为亚种, 而当它们之间的不连续变异表现在两个或多个独立性状上时, 就一律把它们分为不同的物种”(When two such populations display completely discontinuous variation in one character only (frequently supported by partial discontinuities in other characters) I have treated them as subspecifically distinct, whereas they are as a rule classified as different species if discontinuous variation occurs in two or more independent features.)。这是我认为迄今为止最客观、最可操作的划分物种的标准。但非常遗憾的是, 仅仅5年后就被两位权威学者Davis和Heywood (1963)批评“过分简单”: “作为例外, 人们可以用外部形态的单个差异, 如果它总是与互交不育相伴——常因染色体数目的不同所致……”(As an exception one might allow a single difference in external morphology if it were constantly associated with a high degree of intersterility—often associated with a difference in chromosome number…)( Davis和Heywood所说的是生物学物种概念的极端观点, 其代表人物是Löve夫妇, 他们(Löve & Löve, 1961)主张把染色体数目不同的生物类型都分成不同的物种, 因为它们之间存在生殖隔离。这也就是van Valen (1992)把生物学物种概念贬为生殖物种概念的原因。)。
Davis和Heywood (1963)的Principles of Angiosperm Taxonomy是一本很有影响的书, 对被子植物分类学原理作了详尽的阐述, 也花了大量笔墨讨论物种概念。他们也提出了自己的物种概念: “可把物种看成是形态上可界定的单元, 由被假定通常异交繁育的个体群(居群)组成, 是一个或多个基因库的容器和表现”(In summary we may regard species as morphologically definable units, made up of groups of individuals (populations), which it is assumed are usually interbreeding, the containers and expression of one or more gene pools.)。这一物种概念认为物种是形态上“可界定的”, 但未说明如何界定, 被界定的物种是否异交繁育还需“假定”。一个物种能有多个基因库吗?这与生物学物种概念和遗传学物种概念相悖。
Davis和Heywood (1963)赞赏Du Rietz (1930)的概念中提出“间断性”(discontinuity)一词, 也认为“分类学家在划分物种时依据居群在形态上是否显示出变异的间断作出决定”(They (指taxonomists) base their decision about specific delimitation on the presence or absence of morphological breaks in the variation shown by these populations.)。但是他们又接着说: “居群的范围是根据样本的分布判断的(也由其他深入研究过的案例外推); 所要求的形态间断的明显程度也必须依据每个场合的具体情况作出判断”(The limit of the populations is judged on the basis of the distribution of the samples (and by extrapolation from other well-studied situations); the degree of distinctness of the morphological discontinuity required has to be judged on its merits for each case.)。在他们看来, 物种划分离不开判断。
Cronquist (1978)的物种概念是: “物种是一致地和持续地区别分明、可用常规手段鉴别的最小类群”(Species are the smallest group that are consistently and persistently distinct, and distinguishable by ordinary means.)。但他本人也说过, 植物中物种划分的争论多数来自于这样的疑问: 什么样的“一致” (consistent)才算“一致”?什么样的“持续”(per- sistent)才叫“持续”?什么是“常规手段”?因此他的物种概念仍然没有避开主观因素。
2.3.7 其他领域的物种概念
动物学家和古生物学家们又如何看待物种概念呢?动物分类学家Blackwelder (1967)对物种的定义是: “物种是被特定的分类学家认为是或会认为是同一类成员的所有标本, 这或者依据显示的证据, 或者设想它们与其后代或数代之内的遗传上的亲族一样”(…a species consists of all the specimens which are, or would be, considered by a particular taxonomist to be member of a single kind as shown by the evidence or the assumption that they are alike as their offspring or their hereditary relatives within a few generations.)。这一定义含有“特定的分类学家”、 “认为”、“设想”等主观词语, 于是被周长发和杨光(2011)正确地归入人为物种概念之列。
我国著名昆虫学家陈世骧(1978)的物种概念是: “物种是繁殖单元, 由连续又间断的居群组成; 物种是进化单元, 是生物系统线上的基本环节, 是分类的基本单元”。陈先生的物种概念中提到3个单元, 理论上是对的, 但是过于抽象。分类学家能拿这一概念划分出实在而自然的物种吗?
古动物学家Simpson (1943)把形态学物种概念和分类学物种概念分得很清楚。他认为前者很接近遗传学物种概念, 是客观的, 但两者都是不适用的。他说(Simpson, 1943): “分类学物种是一个主观概念”(The taxonomic species is a subjective concept.)。他还说: “分类学物种是根据抽出来的一批标本就形态学物种的最有可能的性状和范围所作的一种判断”(a taxonomic species is an inference as to the most probable characters and limits of the morphological species from which a given series of specimens has been drawn.)。带有“判断”的概念至少是不够科学的。
2.3.8 小结
总结以上的介绍和评述, 除Hedberg (1958)采用的方法外, 所有其他的分类学物种概念, 不管是植物学家提出的还是动物学家提出的, 都带有人为因素。以至于Davis和Heywood (1963)说: “分类学的双重性质: 科学和艺术交汇的场所”(ambivalent nature of taxonomy: a meeting ground of Science and Art.)。对此我很不认同。理由将在讨论部分回答。
现有较流行的物种概念给我这样的印象: 前3类的关系密不可分, 它们清楚地阐述了物种的基本性质: 物种是客观存在的; 物种的所有成员共享一个基因库, 互换基因; 物种之间由于生殖隔离或遗传隔离的存在彼此不能进行基因互换, 因而独立进化。这在理论上基本回答了物种问题, 但是若按这些物种概念确定一个物种则必须进行实验, 而且在许多情况下必须进行长时间的实验, 甚至有不可能进行的实验。就是说, 这3类物种概念从理论上看很好, 但实际上难以操作。系统发生物种概念强调物种是单系, 并用共有衍征来认定单系。我很赞同这一点, 它体现了理论的发展, 又有可操作的方法, 但是过于简单, 既没有指出划分物种所需共有衍征的数目, 也没有提及数量性状。因此要使这一概念付诸实施也还是困难的。生态学物种概念过于抽象, 不过有一点可取, 那就是认识到物种在进化过程中占据独有的生态位。最后, 我很遗憾地说, 绝大多数物种是由分类学家划分的, 划分物种也是分类学家最大的责任, 但是除Hedberg (1958)的方法以外, 分类学物种概念全都含有人为因素。Hedberg (1958)划分物种的原则是分类学的一大进步, 但他并未作为物种概念发表, 而且其陈述被Davis和Heywood (1963)认为“过于简单”。因此, 至今还没有一个既科学又可操作的物种概念。
物种问题是生物学家共同关注的焦点问题之一, 而物种概念和物种划分关乎生物多样性的研究、有效保护和可持续利用。因此, 生物学的发展和生物多样性事业需要科学、可操作的物种概念。新的物种概念应该既反映物种的本质, 又可操作, 使分类学家划分的物种不仅在形态上区别分明, 易于鉴别, 而且能够反映物种的下述属性: (1)物种内的所有成员共享一个基因库, 个体间和居群间存在基因流, 而物种之间由于隔离(也包括地理隔离和生态隔离)不发生基因交换; (2)一个独立的谱系, 与别的谱系分别进化; (3)形成有两个或多个共有衍征的最小的单系群; (4)与别的物种在生态位上有分化; (5)有独立的地理分布。只有达到了这些标准, 才能经得起分子和/或基因组系统发生分析的考验。这里我提出一个新的物种概念迎接需求的挑战, 供讨论。
形态-生物学物种概念: 物种是由一个或多个自然居群组成的生物类群, 种内呈现形态性状的多态性和变异的连续性, 而种间则有两个或多个独立的形态性状显现变异的间断或统计上的间断。
下面我们用5个案例来展示这一物种概念的科学性和可操作性。
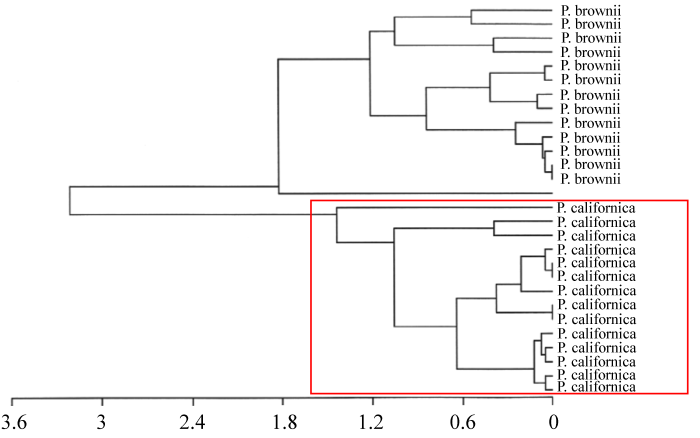
案例1: 北美西部Paeonia californica和P. brownii分与合的故事。
对Paeonia californica曾有4种处理: (1)作独立的物种, 即Paeonia californica Nutt. ex Torr. & A. Gray (1838); (2)作P. brownii的异名(Brewer & Watson, 1876; Jepson, 1909; Munz, 1935); (3)作P. brownii的变种, 即P. brownii var. californica (Nutt. ex Torr. & A. Gray) Lynch (1890); (4)作P. brownii的亚种, 即P. brownii subsp. californica (Nutt. ex Torr. & A. Gray) J. J. Halda (1997); Halda (2004)。这4种处理中究竟哪一种是自然的?
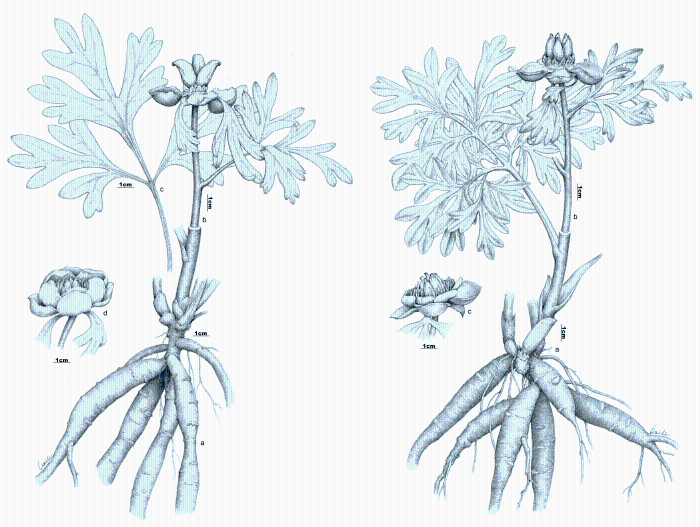
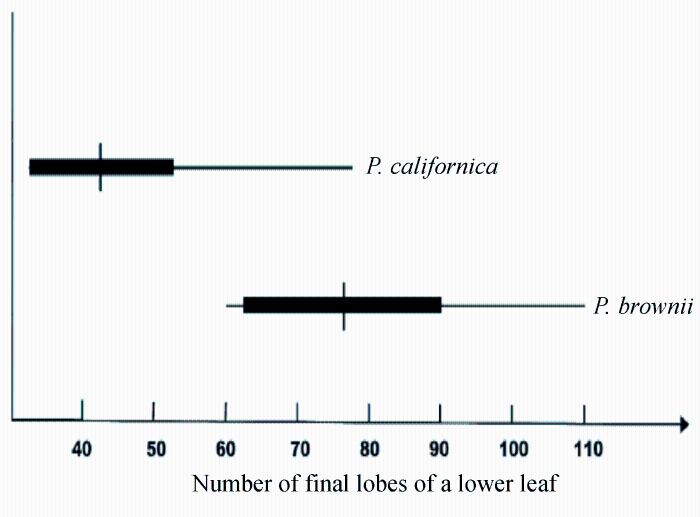
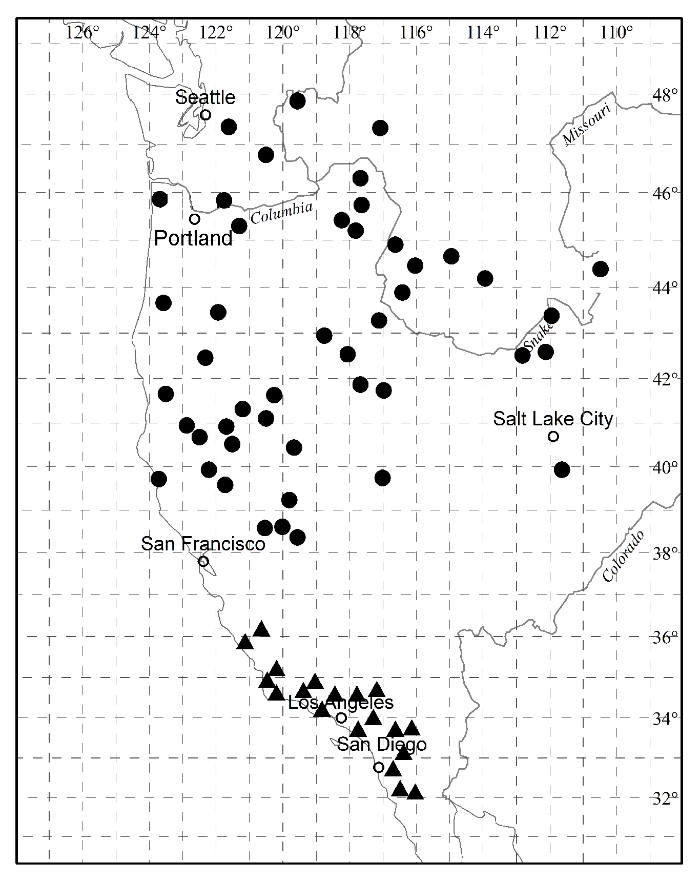
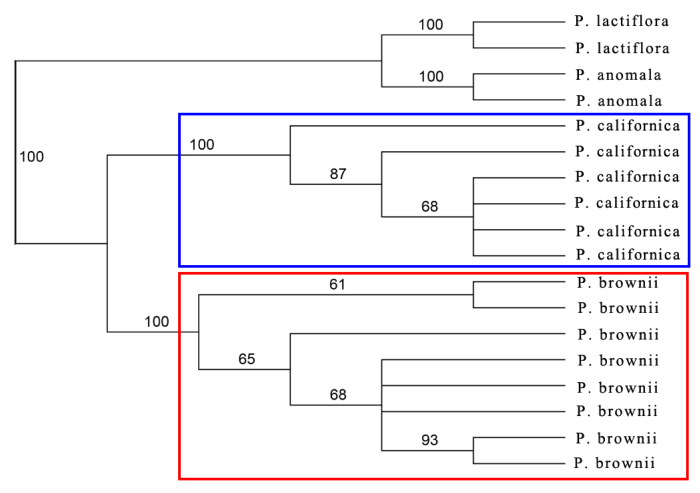
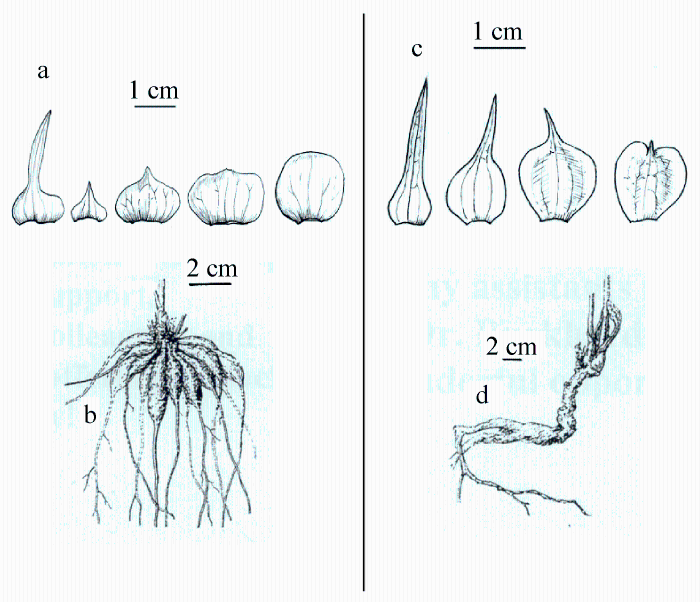
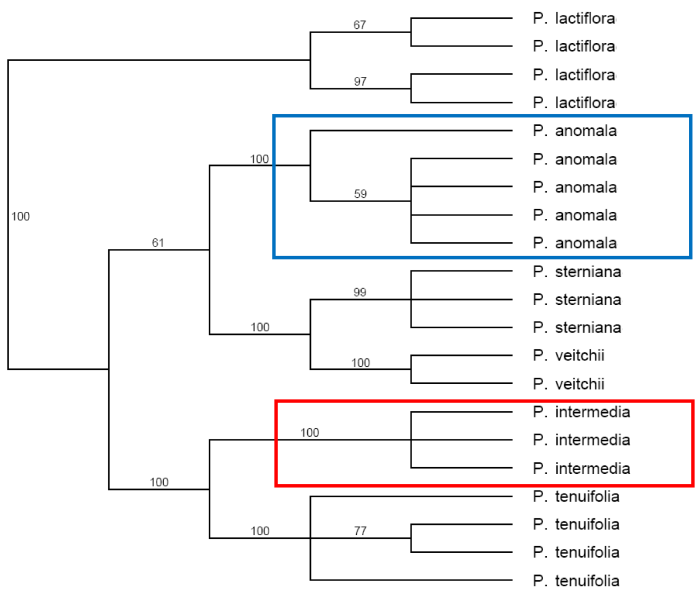
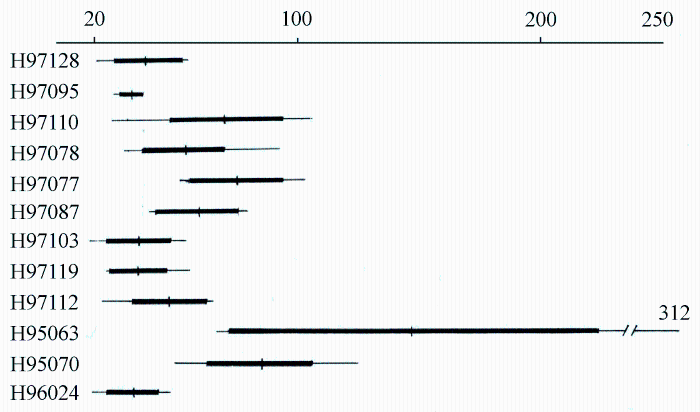
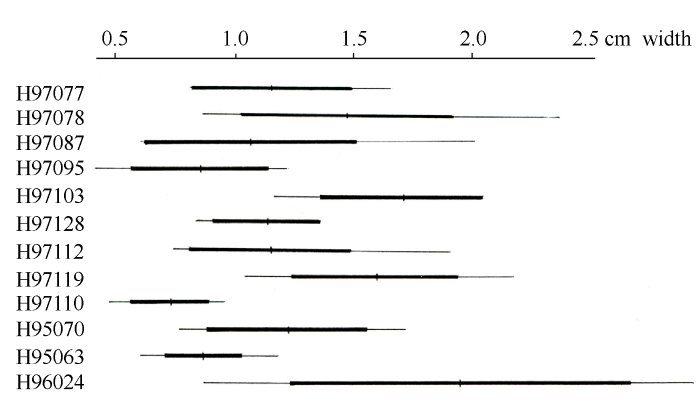
由图5-10可以看出, Paeonia californica和P. brownii之间至少有3个形态性状(下部叶一回三出vs. 二回三出、花瓣长于萼片vs. 短于萼片、下部叶末级裂片数目和心皮数目2 vs. 4)呈现不连续变异, 且有独立的分布区(图9), 因此它们是两个物种。基于25个单拷贝或寡拷贝核基因DNA序列的系统发生分析结果以100%的支持率支持它们是两个独立的谱系, 各自成单系(图10)。
图5 北美西部两种芍药的形态比较(
案例2: Paeonia intermedia C. A. Mey. (块根芍
药, 产中亚)的故事。
块根芍药(Paeonia intermedia C. A. Mey.)于1830年发表后, 亦曾有4种不同的分类处理: (1)作独立的物种, 即Paeonia intermedia C. A. Mey. (1830); (2)作亚种, 即P. anomala L. subsp. intermedia (C. A. Mey.) Trautv. (1904); (3)作变种, 即P. anomala L. var. intermedia (C. A. Mey.) B. Fedtsch. (1905); Pan (1979); (4)作变型, 即P. anomala L. var. hybrida Pall. f. intermedia (C. A. Mey.) Trautv. (1860)。这4种处理中哪一种是科学 的呢?
Paeonia intermedia C. A. Mey.与P. anomala L.在叶片和体态上不易区分, 仅知道前者的根像白薯, 而后者的根呈胡萝卜状, 但在标本馆里难见带根的标本, 因而对这两个类群在形态上的关系并未澄清, 于是造成: (1)不同作者对P. intermedia作了4种不同的处理; (2)标本馆中的标本鉴定常张冠 李戴。
由图11-14可以看出, Paeonia intermedia与P. anomala之间至少有两个形态性状呈现不连续变异, 各为单系, 即使偶尔杂交, 然杂种不育。它们各有独立的分布区(Hong, 2010), 即使有部分重叠, 也在不同的生境中, 前者在山坡灌丛或草地, 后者在沟谷林下或林窗中。因此得出结论: Paeonia intermedia C. A. Mey.和P. anomala L.是两个形态鲜明有别、存在生殖隔离、各呈单系的好种。
图11 Paeonia intermedia (a)和P. anomala (b)两个类群根形态的比较。持根者为1993年时的石河子大学李学禹教授。(洪德元摄)
案例3: Paeonia arietina G. Anderson(产欧洲和土耳其)是不是一个独立的物种?
Paeonia arietina G. Anderson自1818年发表后近150年中一直被作为物种对待(Lynch, 1890; Stern, 1943, 1946), 但却被《欧洲植物志》(Flora Europaea) (Cullen & Heywood, 1964; Akeroyd, 1993)、《土耳其植物志》(Flora of Turkey) (Davis & Cullen, 1965)以及Halda (2004)处理为P. mascula中的一个亚种, 即Paeonia mascula (L.) Mill. subsp. arietina (G. Anderson) Cullen & Heywood (1964)。这一处理是否合理?带着这一问题, 我们在欧洲南部和土耳其作了广泛考察, 并随后作了多方面观察和分析, 得出了以下结果:
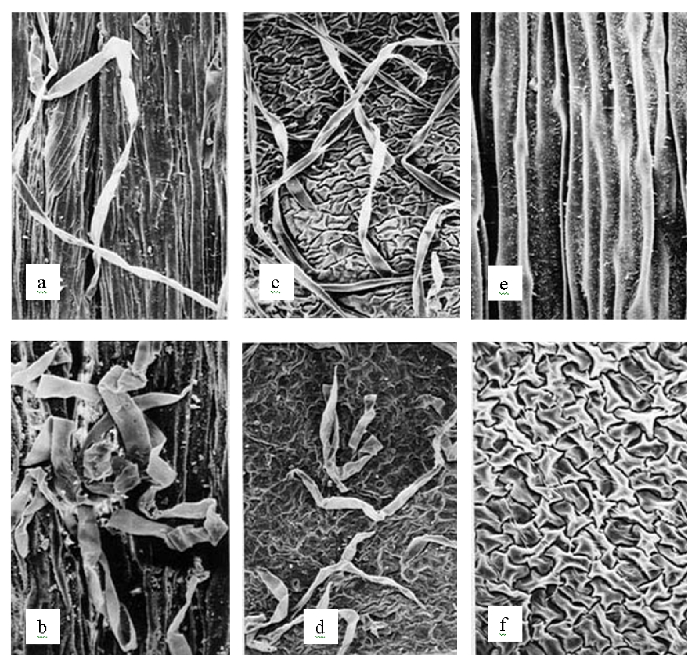
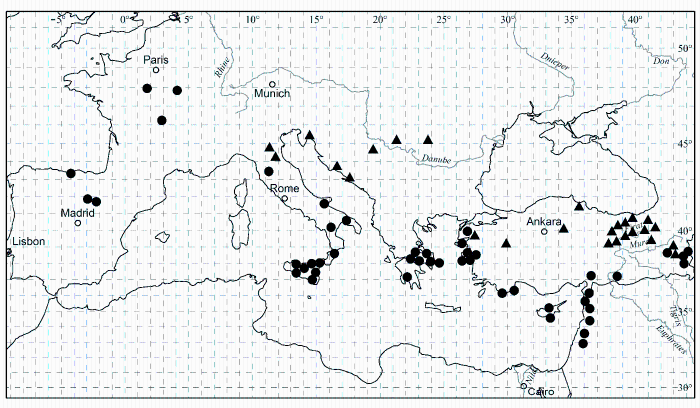
对形态性状的观察和分析显示, Paeonia arietina G. Anderson是一个独立于P. mascula (L.) Mill.的物种, 它们至少有两个形态性状呈不连续变异。前者茎、叶柄和花梗被长毛, 根块状, 而后者茎、叶柄和花梗无毛, 根胡萝卜状(图15, 16); 有着各自独立的地理分布(图17)。这一分类处理纠正了上述文献中的错误。P. arietina在形态性状上的独立性也在系统发生学分析中体现出来, 出现在两个分支上(图18)。
这一案例不仅说明我们的理念是正确、可行的, 而且也说明贯彻这一理念还需要对物种进行深入研究, 特别要对形态性状进行全面观察和分析。
图14 两个不孕的个体, 疑似Paeonia intermedia和P. anomala之间的杂种(PE标本)。左图: 顶端有一朵小而发育不全的花, 根细纺锤状; 右图: 无花, 根细纺锤状。(洪德元摄)
案例4: Paeonia japonica (Makino) Miyabe & Takeda (1910)的故事。
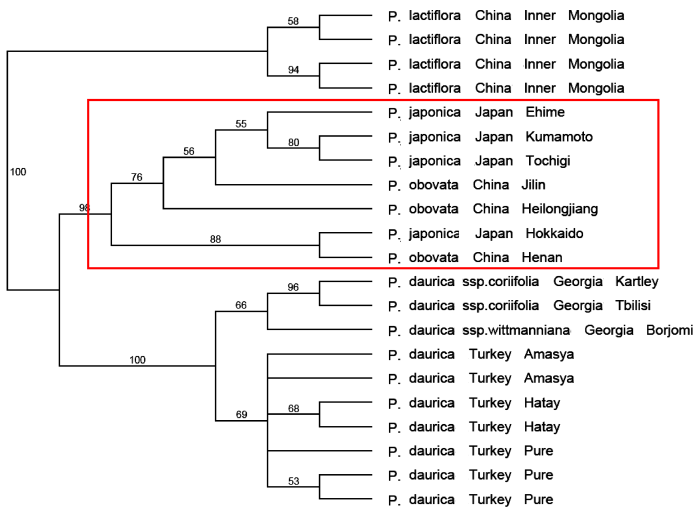
Paeonia japonica是由变种P. obovata Maxim. var. japonica Makino提升为物种等级的。它的身份几经变动: (1)作为独立物种, 且被广泛认同: Paeonia japonica (Makino) Miyabe & Takeda (1910); Schipczinsky (1937); Stern (1946); Ding & Liu (1991); Page & Sinnott (2001); Kadota (2006)在新版Flora of Japan中仍然坚持这一处理。(2)作为亚种: Paeonia obovata Maxim. subsp. japonica (Makino) J. J. Halda (1997); Halda (2004)。(3)作为变种: Paeonia obovata Maxim. japonica Makino (1898); Schi- pczinsky (1921)。(4)作为Paeonia obovata Maxim.的异名: Hong等(2001)和Hong (2010)认为P. japonica只是多变的P. obovata中的一些类型。那么它的身份究竟是什么?
图15 Paeonia arietina (a-d, 均被柔毛)和P. mascula (e, f,两处均无毛)的茎表面和萼片背面的扫描电镜照片, 示被毛的差异。a, b (H02216)和c, d (H02217)均采自土耳其的Sivas; e, f: H01004, 采自法国的Chadieu。a, c, e: 茎表面; b, d, f: 萼片背面。
图16 Paeonia arietina (P. mascula subsp. arietina) (a, b)和P. mascula (c, d)两个类群的根形态。a: H02204, 采自土耳其的Ida山; b: H02224, 采自土耳其的Giresun; c: H02226, 采自希腊的Euboea; d: H02203, 采自土耳其的Canakkale。(洪德元摄)
图18 芍药属芍药组内基于ITS和mat K DNA序列的系统发生关系(
我们在20世纪90年代对P. obovata和P. japonica分布的地区进行了广泛的野外考察, 几乎覆盖中国的所有省份, 以及日本的本州、北海道和四国。对过去学者提出的二者所有区别性状, 如花颜色, 心皮数目、柱头反卷程度、叶被毛、叶姿态等一一进行了观察和分析(Hong et al, 2001)。图19显示花颜色、叶姿态、花丝与花药颜色等的多变性, 图20显示叶被毛密度的连续变异(在中国的秦岭及周边地区, 被毛明显较密, 且为四倍体(依据我们的11个计数)。我们将它处理为亚种, 即P. obovata subsp. willmottiae (Stapf) D. Y. Hong & K. Y. Pan [= P. willmottiae Stapf = P. ovovata var. willmottiae (Stapf) F. C. Stern]。这一新的发现不在讨论范围。)。
图19 Paeonia obovata中花色、叶姿态、花丝和花药颜色的变异。a: 长白山, 花红色, 但叶展开; b: 河南卢氏县, 花瓣白色, 但周边粉色, 花丝下部紫色, 但上部白色; c: 日本爱媛县; d: 河南卢氏县; e: 四川卧龙; f: 吉林安图县。这里也包括了“P. japonica”的特征。
图20 Paeonia obovata中被毛密度和染色体倍性分布图(
我们在那篇文章(Hong et al, 2001)中详细描述了上述各个形态性状的变异, 没有发现一个形态性状呈现变异的间断。显然, P. japonica的身份是P. obovata中的红花类型, 而且在白花与红花之间有一系列过渡, P. japonica应是P. obovata的异名。这一结论也受到具有极高分辨率的系统发生分析的强有力支持(图21)。
案例5: 滇牡丹复合群(Paeonia delavayi Franch. complex)中物种划分的故事。
从表2可以看出滇牡丹复合群中物种划分的历史变动: 最初是4个种, Stern (1946)和方文培(1958)处理为3个种, 而《中国植物志》(潘开玉, 1979)则将它们归并为1个种, 下分3个变种。西藏东南部的 P. lutea var. ludlowii Stern & G. Taylor (1951)在后两项工作中都未提及(表2), 但其分布地点被包括在P. delavayi var. lutea的范围内。对此, 我们自然会问两个问题: (1)西藏东南部的P. lutea var. ludlowii Stern & G. Taylor是什么?(2)其余的分类群是3个种, 还是1个种的3个变种, 还是另有可能?
表2 滇牡丹复合群(Paeonia delavayi Franch. complex) 1980年以前的分类处理
| Stern (1946) | 方文培 (1958) | 《中国植物志》27卷 (潘开玉, 1979) |
|---|---|---|
| P. delavayi Franch.: 云南丽江等 P. lutea Delavay ex Franch.: 云南、西藏东南部 P. potaninii Kom. var. potaninii: 四川西部、云南永宁 var. trollioides (= P. trollioidess Stapf ex Stern): 云南德钦、西藏东南部 ……………………………… P. lutea var. ludlowii Stern & G. Taylor (1951): 西藏东南部 | P. delavayi: 四川木里、西藏察瓦龙、云南 P. lutea: 云南 P. potaninii var. potaninii: 四川木里、云南丽江 var. trollioides: 四川木里、云南丽江 ……………………………… | P. delavayi var. delavayi: 云南西北部、 四川西南部、西藏东南部 var. lutea: 云南、四川西南部、西藏东南部 var. angustiloba: 四川西部 ………………………………… |
图22 西藏大花黄牡丹(Paeonia ludlowii)与滇牡丹(P. delavayi)地下部分形态比较。a: 西藏的P. lutea var. ludlowii Stern & G. Taylor (1951) (P. delavayi var. lutea的一部分), 根渐细, 无地下走茎; b, c: 滇牡丹(P. delavayi) (包括“黄牡丹” P. delavayi var. lutea)根块状加粗, 具地下走茎。
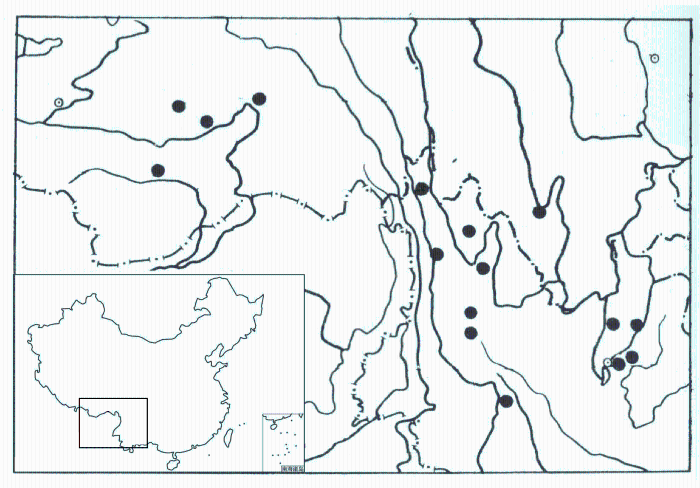
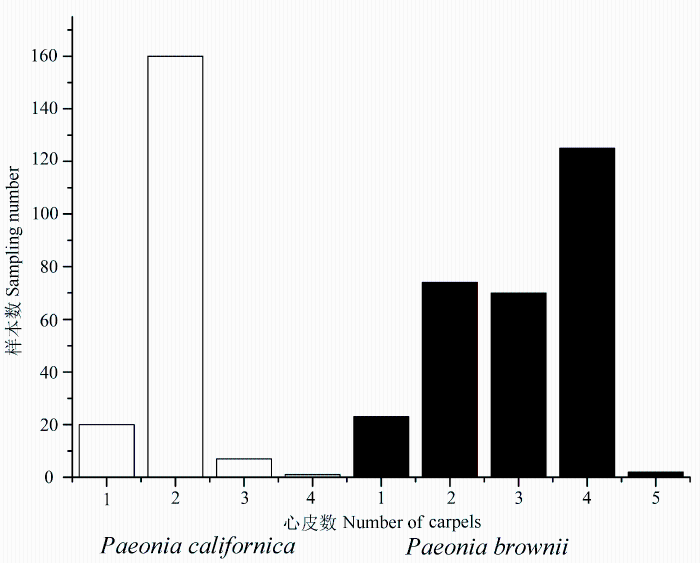
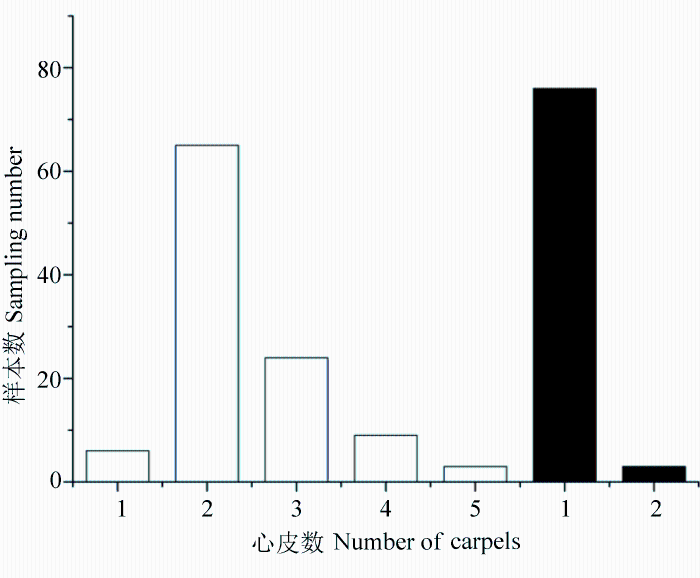
先回答第一个问题。图22展示的是P. lutea var. ludlowii (大花黄牡丹)和滇牡丹P. delavayi (包括“黄牡丹”P. delavayi var. lutea)的地下部分。前者像灌木的地下部分; 而后者有地下横走茎, 根纺锤状增粗, 像白薯。可见两者的地下部分截然不同。图23展示大花黄牡丹和滇牡丹(包括P. delavayi var. lutea)的习性, 前者靠种子繁殖, 成年植株下常见实生苗, 植株形成大丛; 后者常靠地下横走茎繁殖, 植株散生。图24显示两者心皮数目, 大花黄牡丹的花仅有1个心皮, 偶见两个, 而滇牡丹从1至5, 但以2-4为主, 1个心皮的极少见。图25展示蒴果长度的不同。虽然两者变异的极端值不重叠但相连, 然而标准偏差分析显示, 两者变异幅度的统计数据差异悬殊。这样, P. lutea var. ludlowii(大花黄牡丹)至少有4个性状与P. delavayi(滇牡丹)复合体(包括var. lutea)的对应性状呈现间断或统计上的间断。因此, 对第一个问题的回答很肯定, 西藏大花黄牡丹是一个独立的物种, 应该把Paeonia lutea var. ludlowii 提升为物种等级, 即P. ludlowii (Stern & G. Taylor) D. Y. Hong (大花黄牡丹), 它是西藏东南部特有的濒危物种。对于这样的分类处理, 英国学者S. G. Haw评述道: “Hong (1997)把P. ludlowii升级为物种, 提供了一个令人信服的案例。这一结论得到了Zou等(1999) RAPD分析结果的支持”(Haw, 2001)。
图23 西藏大花黄牡丹(Paeonia ludlowii)与滇牡丹(P. delavayi)习性的比较。a: 西藏的P. lutea var. ludlowii Stern & G. Taylor (1951) (P. delavayi var. lutea的一部分)种子繁殖, 茎丛生; b: 滇牡丹(P. delavayi)(包括“黄牡丹” P. delavayi var. lutea)克隆繁殖为主, 茎散生。
图25 西藏大花黄牡丹(Paeonia ludlowii) (a)与滇牡丹(P. delavayi, 包括“黄牡丹”P. delavayi var. lutea) (b)果实长度比较和果实长度标准差(standard deviation)分析
那么第二个问题呢?以前的学者区分这3个物种或变种所依据的是单个性状, 即花冠颜色或叶裂片数目与宽度(图26)。通过对这些性状进行观察和分析发现, 花冠颜色(图27-30)显示, 滇牡丹复合群花瓣颜色不管在居群间还是居群内都是极为多变的, 因此在这一复合群内依据花冠颜色划分物种或变种都是不科学的; 叶裂片数目和宽度的变异情况(图31, 32)则显示, 该性状在居群内和居群间的变异范围都很大, 不存在间断, 而连续变异的性状不能用作分类的依据。因此, 第二个问题的答案也是明确的, 未发现有两个独立形态性状呈现变异的间断, 结论只能是一个非常多变的种滇牡丹(Paeonia
delavayi Franch.), 亦不能划分种下等级(Hong et al, 1998)。对于这样的归并, Haw (2001)又发表了评论: “许多园艺工作者, 大概还有一些植物学家会难于接受这样一个事实, 以前承认的几个种和变种现在全都包括在单个种P. delavayi中, 且不分种下分类群。对此, Hong等(1998)作了认真研究和解释, 为这一做法提供了充分理由。他们的研究显示, 过去用来划分如今包括在这一物种中的那些分类群的性状, 如花颜色、明显总苞的存在与否、叶裂片的宽度全都非常多变, 且彼此之间以及与地理分布之间未表现出相关性。因此, 不能认为它们在分类上有什么意义, 而只是一个多变的物种”。
综上, 对滇牡丹复合群(Paeonia delavayi Franch. complex)分类修订的结果是确认有两个物种(图33):
(1) 大花黄牡丹P. ludlowii (Stern & G. Taylor) D. Y. Hong, Novon 7(2): 157. 1997.
syn.(异名): Paeonia lutea Delavay ex Franch. var. ludlowii Stern & G. Taylor (1951); Paeonia delavayi Franch. var. lutea (Delavay ex Franch.) Finet & Gagnep. 1904, Sensu Fl. Reipubl. Popular. Sin. 1979; China Plant Red Data Book, 1992, p.p.
(2) 滇牡丹P. delavayi Franch., Bull. Soc. Bot. France 33: 382. 1886.
syn. (异名): Paeonia lutea Delavay ex Franch., Bull. Soc. Bot. France 33: 382. 1886 [= P. delavayi var. lutea (Delavay ex Franch.) Finet & Gagnep., 1904]; Paeonia potaninii Kom., Bot. Mater. Gerb. Glavn. Bot. Sada RSFSR 2: 7. 1921; Paeonia potaninii var. trollioides (Stapf ex Stern) Stern, 1943 (= P. trollioides Stapf ex Stern, 1931)。
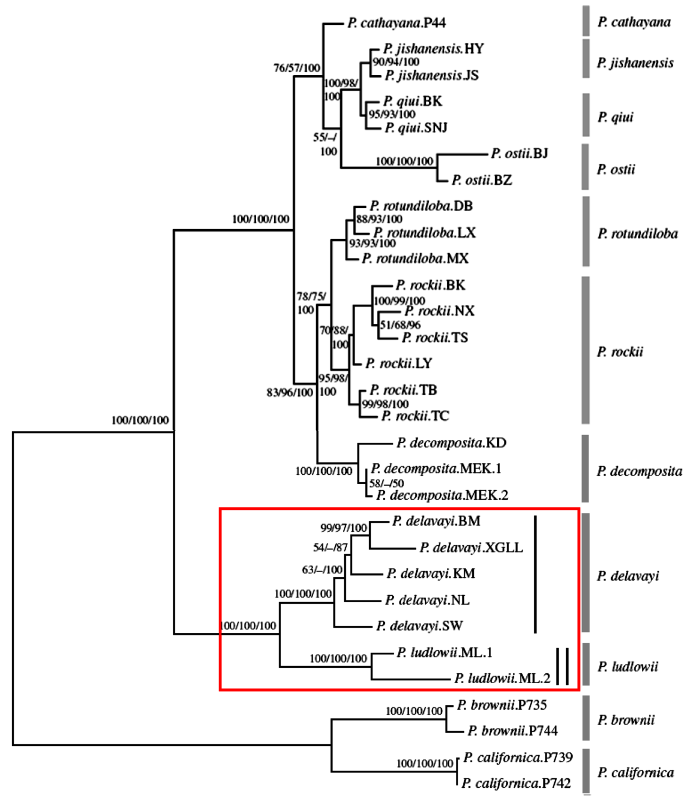
更进一步, 我们团队(Zhou et al, 2014)依据26个单拷贝或寡拷贝核基因DNA序列对牡丹组作了系统发生分析, 得出了分辨率极高的分子树(近乎物种树)。在这一分子树上P. ludlowii和P. delavayi各自成独立的单系, 得到100%的支持。过去承认的P. lutea和P. potaninii及一些变种和P. delayayi集成单一分支(图34), 充分说明滇牡丹复合群包含两个独立的物种, 其中滇牡丹是一个多变的物种, 物种下任何正式分类群的划分都是不科学的。
图26 滇牡丹复合群(Paeonia delavayi Franch. complex)中4个分类群的模式类型。a: 滇牡丹(P. delavayi Franch.); b: 黄牡丹(P. lutea Delavay ex Franch. = P. delavayi var. lutea); c: 银莲保氏牡丹(P. potaninii var. trollioides (Stapf ex Stern) Stern); d: 保氏牡丹(P. potaninii Kom., 1921)。(洪德元摄)
图33 滇牡丹复合群(Paeonia delavayi Franch. complex)两个物种的地理分布。● 滇牡丹(Paeonia delavayi Franch.); ▲ 大花黄牡丹(P. ludlowii (Stern & G. Taylor) D. Y. Hong)。
首先, 本文对现有较流行的6类物种概念作了回顾和述评, 发现生物学物种概念、遗传学物种概念和进化物种概念恰当地概括了物种的自然本质, 推动了人们对物种的认识。但可惜这些概念的可操作性不强。系统发生物种概念用共有衍征来确立单系类群, 无疑是高明的。但是对于物种一级没有指出用多少个共有衍征来限定“可鉴别的最小的簇”(the smallest diagnosable cluster)或“涵盖范围最小的分类群”(the least inclusive taxon)。若仅仅用一个共有衍征, 那得到的“簇”(cluster)或“分类群”(taxon)则不一定是物种, 而很可能是种下等级。因此, 它的可操作性和准确性也不很高。物种划分是由分类学家进行的, 可是所有流行的分类学物种概念都或多或少含有主观成分。Hedberg (1958)的方法虽不含主观因素, 但他未作为物种概念提出, 仅叙述了所用的方法, 而且叙述过于简单, 也未提数量性状。总之, 目前尚没有一个既科学又可操作的物种概念。造成这一状况的主要原因是过去研究遗传、进化等的学者不从事分类研究的实践, 而从事物种划分的学者一般不关心遗传与进化等领域的思想, 就是说, 学术沟通很不够。
其次, 形态性状有两类: 质量性状和数量性状。处理质量性状往往是回答有和无, 黑与白等不同状态的问题, 比较容易。数量性状是所有生物学家都必须面对的, 但是如何恰当地处理它却不那么容易。在遗传学和物种生物学(biosystematics)(早期称为实验分类学, experimental taxonomy)中, 统计学方法不可或缺。但是在分类学(至少是植物分类学)中, 应用统计学方法还不常见。遇到数量性状, 往往用“较长”或“较短”、“宽”或“窄”等含糊词语, 是否作为鉴别性状也会因人、因时而异。在我的实践中逐渐认识到, 统计学方法对于科学的分类处理是必不可少的。例如在滇牡丹复合群中, 叶裂片数目和它的宽度在我们研究之前一直被用作区分物种或变种的性状, 然而我们的统计分析否定了它们的分类价值(图31, 32)。我们经过统计分析后作的分类处理令人信服(Haw, 2001)。Paeonia lutea var. ludlowii Stern & G. Taylor在心皮数目和果实长度上就绝对值而言, 和该复合体的其他部分是连续的, 但心皮数目柱状图(图24)和果实长度统计数据(图25)告诉我们, 其间的性状变异是间断的。本文提出的物种概念把数量性状与质量性状分开, 并引入统计学方法处理数量性状, 这是它的一大特点。
第三, 应当充分认识单个形态性状反映的生物学本质。单个形态性状通常是由一个基因或一个基因家族决定的, 有的很显著, 如花的颜色、被毛的有无、被毛的类型等。它们通过基因突变、基因流和重组, 产生居群内的多态现象(polymorphism)。很显然, 这样的差异是不应该给予任何分类学等级的。然而很遗憾, 有不少分类学工作者把这样的差异描述为新变型、新变种, 甚至新物种。所以, 在新的物种概念中我们应用至少两个独立的形态性状来区分物种。
第四, 与上一论题有关, 持无性繁殖物种概念的学者把无性繁殖(营养繁殖、孤雌生殖、无融合结籽)生物的不同无性系(clone)区分为不同物种。自然界可能并没有真正的专性无性繁殖生物(Davis & Heywood, 1963)。无性系之间若干年后在一定条件下又可能进行有性生殖。竹子数十年至上百年开一次花, 而开花导致的后代分离, 产生更多的无性系。把不同无性系处理为物种的做法是不科学的。无性系之所以被一些人处理成物种是因为无性系之间的差异虽很微小, 但因克隆繁殖而很稳定, 看起来具有隐种(sibling species)的特点。岂不知, 真正的隐种之间存在着至少两个形态性状变异的间断, 果蝇中的Drosophila pseudoobscura和D. persimilis是隐种, 植物中的Gilia transmontana及另外4个物种是隐种, 这是隐种的经典例子。而无性系之间往往只有一个基因的差异, 而且它们很可能还是兄弟姊妹。现在有一些人把无性系、隐种以及多态现象混为一谈, 把依据多态现象和无性系描述的“新种”美其名为“隐种”。
第五, 本文提出的物种概念基于形态性状。人们会问, 基于形态性状的物种概念能否反映物种的生物学本质?人们观察到的形态性状是两种作用的产物, 即遗传与环境饰变, 而且环境饰变的幅度(即可塑性)也是遗传控制的。在本文的5个案例中,按新的物种概念划分的物种在高支持率的系统发生树上成为单系, 成为独立的谱系, 其内部不再有强有力支持的次级分支, 说明物种内是有基因交流的。这表明关于单一性状和连续性变异的数量性状不能作为划分物种依据的观点是科学的。这样划分的不同物种在系统发生树上均彼此独立, 互不混杂, 显示它们之间没有基因交换, 这应该是隔离(包括生殖隔离、遗传隔离、生态隔离和地理隔离)的结果。而且, 所划分的物种有独立的分布区, 与其他物种在生态位上有分化。可以说, 本文提出的物种概念与一些较为流行的物种概念高度相融, 更重要的是, 新的物种概念还有高度的可操作性。
第六, 在前文引述的分类学物种概念中, 除Hedberg (1958)所应用的方法外都含有主观成分, 充斥着“有经验的系统学家”、“判断”、“假定”、“通常手段”等带主观性的字眼。没有科学的理念和方法就难有科学的实践。对类群的处理常因人而异, 难见一致, 以至于连植物学权威Davis和Heywood (1963)都说“分类学的两重性: 科学和艺术交汇的场所”。真是这样吗?科学与非科学的分水岭是: 结论的可重复性和可预期性。在本文叙述的5个案例中划分的物种都为后来获得的高分辨率系统发生树所高度支持。这使我更加坚信, 分类学是一门科学, 当然亦是一门难于驾驭的科学。我承认, 现有的分类学研究结果中大多包含有程度不同的人为成分。这是因为: (1)学科本身的难度。生物学就是一门复杂的科学, 不像数学中的1 + 1 = 2, 物理学中的惯性定律, 化学中的元素周期表。分类学比一般生物学分支更难, 它研究的对象是生物多样性。可以说, 生物多样性就是“复杂”的代名词。(2)客观和历史条件的限制。生物多样性规模究竟有多大, 至今仍是一个谜, 然而投入的人力、经费和时间仍然是非常有限的, 尤其是在热带、亚热带地区困难更加突出。这些地区分类研究的深度可想而知。不能因客观和历史条件造成的现实分类处理中的人为成分而否定分类学本身的科学性。
最后, 应当说明, 运用这一新的物种概念的基础是要对所有形态性状进行观察和分析。不仅要观察所有形态性状, 还要使掌握的样本具有代表性, 为性状分析奠定扎实基础。在Paeonia arietina和P. intermedia的案例中, 如果过去的分类学家对这两个物种的地下部分和萼片进行了全面深入的观察和科学分析就不会造成分类处理历史中的混乱。P. lutea var. ludlowii Stern & G. Taylor被误定为P. delavayi var. lutea也是因为对前者的形态性状缺乏全面的观察和分析。
致谢: 首先我要感谢我们牡丹、芍药团队的周世良、葛颂、汪小全、张大明、潘开玉、邹新慧、桑涛、饶广远(北京大学)和周志钦(西南大学)等。没有这一团队的努力, 牡丹群和芍药群的系统发生分析不可能达到近乎物种树的水平。这使我有勇气构想我的物种概念。特别感谢组织我们1997年云南考察的虞泓教授(云南大学)和考察队成员顾红雅教授(北京大学); 组织我们1993年考察新疆的李学禹教授(石河子大学)和考察队成员张芝玉教授(第二军医大学)。我也十分感谢鼓励我在年迈之年还构思物种概念这一极其具挑战性课题的孔宏智研究员和王印政研究员(均属系统与进化植物学国家重点实验室)。很感谢四川大学刘建全教授与我分享他对物种概念的见解。非常感谢加拿大的Woodward女士以及美国的文军(J. Wen)、D. Boufford、B. Barthol- omew和I. Al-Shehbaz 4位博士在我研究北美西部两种芍药问题上的鼎力支持; 在日本我得到了Nakata博士, 在土耳其和地中海地区我得到了Kuruklu博士和Fridlender博士的无私帮助。感谢王强博士提供伦敦自然博物馆前大花黄牡丹(Paeonia ludlowii)的照片。很感谢宫晓林、彭丹博士和宣晶在稿件加工、图像处理中的大力协助。我十分感谢美国国家地理学会给我5个基金支持, 使我的足迹能够遍布牡丹、芍药的几乎所有分布地点; 十分感谢国家自然科学基金委提供重点项目支持, 使我的团队能够对牡丹和芍药两类群进行如此深入的系统发生分析。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
Speciation in mammals and the genetic species concept. |
| [3] |
Taxonomy: A Text and Reference Book. Wiley, New York.
|
| [4] |
|
| [5] |
Species: the concept, category and taxon.
|
| [6] |
|
| [7] |
Species concepts and speciation analysis.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
Principles of Angiosperm Taxonomy . |
| [12] |
Evolution. W. H. Freeman and Company , |
| [13] |
The fundamental units of biological taxonomy. |
| [14] |
Notes on Chinese Paeonies. 中国芍药属的研究 . |
| [15] |
|
| [16] |
Biosystematic studies on Adenophora potaninii Korsh. complex (Campanulaceae). II. Crossing experiment. |
| [17] |
|
| [18] |
Systematic treatment of the genus Paeonia L. with some nomenclatoric changes. |
| [19] |
|
| [20] |
Tree peonies: a review of their history and taxonomy.
|
| [21] |
The taxonomic treatment of vicarious taxa. |
| [22] |
Peaonia (Paeoniaceae) in Xizang (Tibet). |
| [23] |
|
| [24] |
Taxonomy of Paeonia delavayi complex. |
| [25] |
Cytogeography and taxonomy of the Paeonia obovata polyploid complex (Paeoniaceae).
|
| [26] |
Permanent splits as speciation events: a formal reconstruction of the internodal species concept. |
| [27] |
Chromosome numbers of central and northwest European plant species. |
| [28] |
A new classification of the genus Paeonia. |
| [29] |
|
| [30] |
On biological species, species concepts and individuation in the natural world. |
| [31] |
|
| [32] |
|
| [33] |
Individuality, pluralism, and the phylogenetic species concept. |
| [34] |
Species concept: a case for pluralism. |
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
The cladistic solution to the species problem. |
| [39] |
Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). |
| [40] |
Criteria for genera, species and subspecies in zoology and paleontology.
|
| [41] |
The species concept.
|
| [42] |
|
| [43] |
|
| [44] |
Genus Paeonia. |
| [45] |
A Study of the Genus Paeonia. Royal Horticultural Society, London. |
| [46] |
A new peony from SE Tibet. |
| [47] |
Ecological species, multispecies, and oaks. |
| [48] |
|
| [49] |
|
| [50] |
A list of 26 species “concepts” source. . |
| [51] |
|
| [52] |
The genic view of the process of speciation.
|
| [53] |
|
| [54] |
Multiple species of wild tree peonies gave rise to the “king of flowers” Paeonia suffruticosa Anderews. |

备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()
/
| 〈 |
|
〉 |















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}