物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 )。很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 )。其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 )。这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献。
基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾。一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 )。因此, 常见种决定丰富度格局的普适性仍需进一步验证。同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 )。但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究。事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视。稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题。
黄土高原是我国特有的地理区域, 该区环境恶劣, 资源贫乏。由于水土流失以及人为活动干扰, 其生态系统正处于极度退化的状态, 因此, 加速该地区退化生态系统的恢复与重建是当务之急(李裕元和邵明安, 2004 ), 而物种多样性的恢复与保护是该区生态系统恢复与重建的重要基础。因此, 在黄土高原进行物种多样性格局研究具有非常重要的理论和现实意义。本研究以该区的顶极群落辽东栎(Quercus wutaishanica )林为模式群落, 根据物种频度序列通过逐一添加(或去除)稀有种或常见种的方式, 探讨由此引起的群落总体物种多样性及其成分(α多样性和β多样性)的变化程度, 以度量稀有种和常见种对物种丰富度格局的相对贡献, 旨在验证常见种对物种多样性格局决定性假说在黄土高原的适用性, 比较稀有种和常见种对α多样性和β多样性这两种成分相对贡献的差异性。
1 研究方法
1.1 数据来源
在对子午岭植被类型进行全面考察的基础上, 选择具有代表性的辽东栎林为研究样地进行随机抽样调查。根据生长型和高度将样地中的植物划分成乔木层(高度> 3 m)、灌木层(高度1-3 m)和草本层(高度< 1 m)三层分别进行调查。共设置5个样地, 在每个样地设置5个20 m × 20 m的样方调查乔木层植物, 每个乔木层样方中分别设置5个4 m × 4 m和5个1 m × 1 m的小样方调查灌木层和草本层植物。共调查了乔木层样方25个, 灌木层和草本层样方各125个。乔木层主要由乔木物种组成, 灌木层由灌木种和少量乔木的幼树组成, 而草本层主要由草本植物组成, 还包括部分乔木和灌木的幼苗。记录每个样方的物种种类、盖度、数量、高度、乔木的胸径和冠幅等数量指标, 同时记录样地的经纬度、海拔、坡度、坡向、坡位以及林冠郁闭度等环境特征。
1.2 分析方法
稀有种和常见种的定义依赖于研究尺度和研究目的, 一般根据物种的分布范围来定义, 如多度和频度(Curnutt et al, 1994 ; 李智琦等, 2010 )。但是, 如果对群落中的物种进行直接分类, 可能会导致不同种类物种数量的不均衡性, 这不利于物种类别间的比较。为了研究稀有种和常见种对物种丰富度的相对贡献, 本研究采用频度来定义物种的稀有性和常见性, 按照频度由低到高或者由高到底的序列对物种依次排序(Lennon et al, 2004 ; Heegaard et al, 2013 ), 然后根据序列逐一添加(或去除)物种, 通过比较物种添加(或去除)前后所引起的多样性变化程度来确定该物种对物种丰富度格局的相对贡献。
1.2.1 稀有种和常见种对群落总体物种丰富度的相对贡献
此方法以相关系数为测度指标, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的贡献。其中, 群落由样方组成, 每个样方的物种数即为该群落的物种丰富度, n 个样方可以理解为群落的n 次重复, 添加或去除物种并不改变样方个数而只改变每个样方中的物种数。因而可以度量两个群落间的相关系数大小并通过相关系数的变化间接度量稀有种和常见种的相对贡献, 这也是研究该问题的经典方法(Lennon et al, 2004 )。
1.2.2 稀有种和常见种对物种丰富度成分的相对贡献
通过比较添加或去除处理前后所引起的物种丰富度成分(α多样性和β多样性)的直接变化来度量稀有种和常见种对物种多样性格局的相对贡献。其中, α多样性的值为各个样方中物种丰富度的平均值。由于物种分布范围的不同, 导致样方间α多样性的变化不同, 故α多样性可直接度量稀有种和常见种对α多样性格局的相对贡献。
同样地, 稀有种和常见种的不同分布也是导致群落异质性的重要原因。因此, 物种添加(或去除)前后群落异质性的变化也是度量稀有和常见种对物种丰富度格局相对贡献的有效指标。而β多样性既可以有效度量群落的异质性, 还是群落物种丰富度的重要组成成分(Beck et al, 2012 )。关于β多样性的度量方式很多, 但是, 绝大多数β多样性指数的大小常常依赖于α多样性的变化(Anderson et al, 2011 )。例如, 两个样方间的共有种常常随α多样性的增加而增加, 即β多样性随α多样性的增加而减小, 反之亦然(Chase et al, 2011 )。因此, 通过常规的β多样性指数不能有效比较稀有种和常见种二者的效应大小。
根据Vellend等(2007)的方法, 本研究引入一种基于概率的β多样性指数βRC , 该指数通过随机方式排除了不同样方间的α多样性差异, 可以反映β多样性的真实变化。其计算过程如下: 假设群落由n 个样方组成, 其中, 样方x 有i 个物种, 样方y 有j 个物种。首先计算两个样方的实际共有物种数, 然后每次从“区域种库”中随机抽取i 和j 个物种分别赋予x 和y 两个样方, 计算两个样方的期望共有物种数。重复此过程9,999次, 以这9,999次重复中期望共有物种数≥实际共有物种数的比例即βRC 值作为样方x 和样方y 间的真实β多样性度量。n 个样方两两间形成的βRC 多样性矩阵的平均值即为该群落的β多样性(Vellend et al, 2007 )。其中, 所有取样单元中的物种整合形成一个“区域种库”, 种库中物种被选择的机率依赖于物种频度大小。由于βRC 指数不依赖于α多样性的大小(Raup & Crick, 1979 ; Chase et al, 2011 ), 因此可 以有效度量稀有和常见种对β多样性格局的相对 贡献。
2 结果
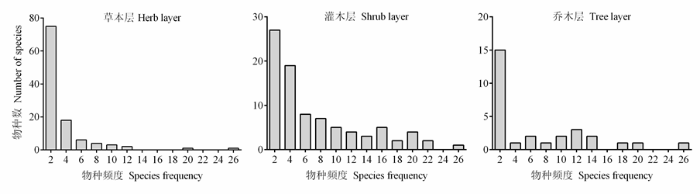
乔、灌、草三层所有样方共记录到191个物种, 其中, 乔木层物种29个, 灌木层物种87个, 草本层物种110个。由于群落中稀有种较多, 乔、灌、草三层都形成了明显右偏的物种频度分布格局(图1 )。其中, 频度≤ 2的物种占到各层物种总数的30%以上。草本层的常见种为细叶薹草(Carex duriuscula )和紫菀(Aster tataricus ), 灌木层的常见种为土庄绣线菊(Spiraea pubescens )、茶条槭(Acer ginnala )幼树、水栒子(Cotoneaster multiflorus )和辽东栎幼树, 乔木层的常见种为辽东栎、甘肃山楂(Crataegus kansuensis )和茶条槭等。
图1 子午岭辽东栎群落各层物种频度分布
Fig. 1 Species-frequency diagram for different layers of Quercus wutaishanica forests in the Ziwu Mountains of Loess Plateau
2.1 稀有种和常见种对群落总体物种丰富度的相对贡献
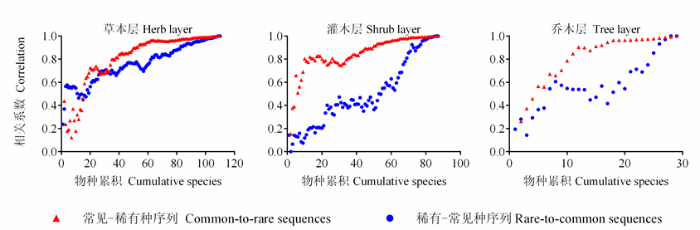
无论是稀有-常见种序列还是常见-稀有种序列, 随着物种加入形成的“新”群落与群落总体物种丰富度的相关性均逐渐增加。但是, 常见-稀有种序列与群落总体的物种丰富度的相关系数较稀有-常见种序列的相关性高, 且不同物种引起的相关系数的增加程度明显不同(图2 )。可以看出, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大。同样, 稀有种的效应在稀有-常见种序列上表现比较明显, 但其在该序列上的相关系数增加效应较常见种的增加效应仍显缓慢, 且相关系数均小于0.5, 在统计学上被认为是低度相关, 这一段的长度也明显依赖于稀有种的数量, 即稀有种越多, 相关系数缓慢增加的区域就越大。
图2 稀有种和常见种对总体物种丰富度格局的贡献。
Fig. 2 Contribution of rare and common species to overall pattern in species richness.
2.2 稀有种和常见种对物种丰富度成分的相对贡献
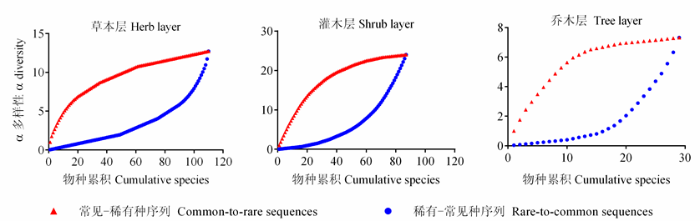
稀有种和常见种对物种丰富度成分的影响在两种物种频度序列中均呈现了明显不同的变化趋势, 这不仅表现在α多样性随物种的加入呈增长趋势而β多样性随物种的加入呈递减趋势, 同时还表现为稀有种和常见种对这两种丰富度成分的影响截然不同。其中, α多样性的变化与相关系数的度量结果相似, 在常见-稀有种序列中随物种的加入呈明显的对数变化趋势, 而在稀有-常见种序列中随物种的加入呈明显的指数增长趋势(图3 )。
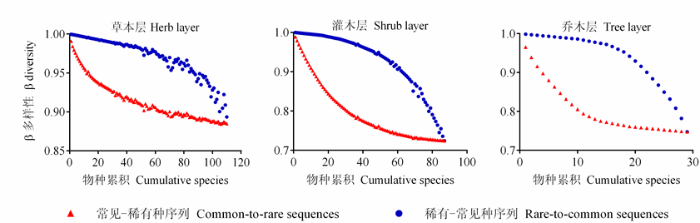
β多样性在两个物种序列中呈现与α多样性完全不同的变化趋势。在常见-稀有种序列中, β多样性先迅速降低后逐步稳定; 而在稀有-常见种序列中, β多样性先稳定后迅速降低(图4 )。可以看出, 无论是α多样性还是β多样性, 二者的急剧变化都表现为在常见种的加入阶段, 而稀有种对两种物种多样性成分的相对影响较小(图3 , 图4 )。
图3 稀有种和常见种对物种丰富度格局α多样性成分的贡献。
Fig. 3 Contribution of rare and common species to α diversity pattern in species richness.
图4 稀有和常见种对物种丰富度格局β多样性成分的贡献。
Fig. 4 Contribution of rare and common species to β diversity pattern in species richness.
3 讨论
尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少。与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献。研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大。同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段。这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 )。不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论。因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 )。
需要注意的是, 常见种对群落物种丰富度格局的主导性并不依赖于稀有种和常见种在群落中的相对比例。本研究中, 尽管稀有种的数量明显多于常见种(图1 ), 但是群落总体的物种丰富度及其成分的多样性格局仍然由常见种主导。这可能是因为, 尽管常见种的物种数较稀有种少, 但其分布范围广, 由此引起的多样性效应也就越大。一个常见种的增加或者减少可以引起整个或者多个样方内的物种变化, 这在α水平上十分明显。而稀有种则不然, 由于其仅局限于个别的样方, 它的增加或减少引起的整个群落物种丰富度的变化较小。尽管乔、灌、草三层中常见种的影响都表现了相似的变化趋势, 但是, 由于其在各层的比例不同, 趋势线也不尽相同。一般地, 常见种的数量越多, 其效应越明显, 对丰富度格局的贡献也就越大(图3 )。
群落的物种丰富度不仅依赖于α多样性, 而且依赖于β多样性。很多研究表明, β多样性也是很多群落物种丰富度的重要贡献者(Beck et al, 2012 )。本研究结果表明, 稀有种和常见种所引起的β多样性的变化与α多样性变化方向完全相反。同时, 稀有种的加入所引起的β多样性变化较小, 但随着常见种的进入, β多样性急剧下降(图4 )。这说明, 与α多样性一样, 常见种也是β多样性格局的重要贡献者, 这与人们的预期并不一致。群落中的稀有种数量较多且与群落总物种数显著正相关(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 ), 物种丰富度常常呈现随取样面积增加而增加的内嵌式分布格局(Wright et al, 1998 )。一般认为, 两个群落间物种丰富度的差异应该是β多样性间的差异。例如, 对北美树栖甲虫群落物种丰富度的多尺度研究结果表明, 大尺度上的β多样性强烈影响稀有种(物种多度< 0.05%)的丰富度, 稀有种甚至主导了整个群落的丰富度格局(Gering et al, 2003 )。对稀有种主导整个群落物种丰富度格局的一个可能解释是由于稀有种占较大的比例(36%-47%)所致。
与此研究相似, 本文中乔、灌、草三层的物种分布格局也均为稀有种占绝大比例的右偏分布, 但研究结论却不完全一致, 可能原因如下: (1)两个研究中定义稀有种和常见种的方式不同, 尽管分布范围广的物种多度也相对较大, 但是局域群落上的优势种与大尺度上的常见种是不同的; (2)物种多样性格局是尺度依赖的(Gering et al, 2003 ; Heegaard et al, 2013 ), 研究稀有种和常见种对多样性格局的相对贡献应该充分考虑空间尺度的影响, 这也是下一步应该亟需开展的重要工作; (3)两个研究中使用的β多样性度量指数不同, 如前所述, 使用依赖于α 多样性变化的β多样性指数不能真实度量β多样性 大小。
目前, 关于常见种和稀有种对物种丰富度格局贡献差异的原因仍不是很清楚, 一个可能的解释是常见种是群落物种丰富度的信号(signals)贡献者, 而稀有种是物种丰富度的噪音(noise)贡献者(Vázquez & Gaston, 2004 )。尽管βRC 多样性指数考虑了α多样性的变化对β多样性的影响, 但是该指数仍具一定的局限性(Chase et al, 2011 )。首先, 该指数来自于古生物学, 并没有很好地考虑群落内不同物种多度的差异, 这对于群落生态学来说是一种不足; 第二, 该方法没有考虑物种在大尺度上的出现概率(Raup & Crick, 1979 ; Vellend et al, 2007 )。一般来说, 分布范围广的物种出现的概率更高, 但是βRC 多样性指数的随机模型没有考虑到这个权重。另外, 种库大小也将明显影响βRC 多样性指数的大小(Gotelli & Colwell, 2001 )。
物种多样性格局的形成不仅依赖于群落的构建过程, 同样依赖于群落中的物种组成(如稀有种和常见种)。稀有种和常见种对物种多样性格局的相对贡献的研究表明, 常见种不仅主导群落的总体物种丰富度格局, 同时也是α多样性和β多样性格局的重要贡献者。尽管常见种对群落物种多样性格局具有主导性, 但是这不能否定稀有种对群落多样性格局的贡献, 很多研究表明, 稀有种也是群落功能的重要载体。这也暗示我们, 仅确定稀有种和常见种对群落多样性格局的相对贡献的价值可能有限, 要得出有意义的结论还应结合其他信息综合分析。总之, 稀有种和常见种对群落的贡献犹如一个硬币的两个面, 二者对物种多样性格局的贡献应该是共同的。在物种多样性的保护实践中, 应该根据物种的不同性状采取不同的保护措施。
参考文献
文献选项
[1]
Anderson MJ Crist TO Chase JM Vellend M Inouye BD Freestone AL Sanders NJ Cornell HV Comita LS Davie KF 2011 ) Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist.
Ecology Letters , 14 , 19 -28 .
[本文引用: 1]
[2]
Beck J Holloway JD Khen CV Kitching IJ 2012 ) Diversity partitioning confirms the importance of beta components in tropical rainforest Lepidoptera.
The American Naturalist , 180 , E64 -E74 .
[本文引用: 3]
[3]
Berg A Tjernberg M 1996 ) Common and rare Swedish vertebrates—distribution and habitat preferences.
Biodiversity and Conservation , 5 , 101 -128 .
[本文引用: 3]
[4]
Curnutt J Lockwood J Luh HK Nott P Russell G 1994 ) Hotspots and species diversity.
Nature , 367 , 326 -327 .
[本文引用: 1]
[5]
Chase JM Kraft NJ Smith KG Vellend M Inouye BD 2011 ) Using null models to disentangle variation in community dissimilarity from variation in α-diversity.
Ecosphere , 2 , 1 -11 .
[本文引用: 3]
[6]
Freestone AL Inouye BD 2006 ) Dispersal limitation and environmental heterogeneity shape scale-dependent diversity patterns in plant communities.
Ecology , 87 , 2425 -2432 .
[本文引用: 2]
[7]
Gaston KJ Blackburn TM 1996 ) Global scale macroecology: interactions between population size, geographic range size and body size in the Anseriformes.
Journal of Animal Ecology , 65 , 701 -714 .
[本文引用: 2]
[8]
Gaston KJ 2008 ) Biodiversity and extinction: the importance of being common.
Progress in Physical Geography , 32 , 73 .
[本文引用: 1]
[9]
Gering JC Crist TO Veech JA 2003 ) Additive partitioning of species diversity across multiple spatial scales: implications for regional conservation of biodiversity.
Conservation Biology , 17 , 488 -499 .
[本文引用: 3]
[10]
Gotelli N Colwell RK 2001 ) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness.
Ecology Letters , 4 , 379 -391 .
[本文引用: 1]
[11]
Heegaard E Gjerde I Sætersdal M 2013 ) Contribution of rare and common species to richness patterns at local scales.
Ecography , 36 , 937 -946 .
[本文引用: 3]
[12]
Hubbell SP 2001 ) The Unified Neutral Theory of Biodiversity and Biogeography . Princeton University Press , Princeton .
[本文引用: 1]
[13]
Kraft NJ Comita LS Chase JM Sanders NJ Swenson NG Crist TO Stegen JC Vellend M Boyle B Anderson MJ 2011 ) Disentangling the drivers of β diversity along latitudinal and elevational gradients.
Science , 333 , 1755 -1758 .
[本文引用: 1]
[14]
Landi S Chiarucci A 2014 ) Commonness and rarity of plants in a reserve network: just two faces of the same coin.
Rendiconti Lincei , 25 , 369 -380 .
[本文引用: 2]
[15]
Lennon JJ Koleff P Greenwood JJ Gaston KJ 2004 ) Contribution of rarity and commonness to patterns of species richness.
Ecology Letters , 7 , 81 -87 .
[本文引用: 6]
[16]
Li YY Shao MA 2004 ) The change of plant diversity during natural recovery process of vegetation in Ziwuling area.
Acta Ecologica Sinica , 24 , 252 -260 . (in Chinese with English abstract)
[李裕元 , 邵明安 (2004 ) 子午岭植被自然恢复过程中植物多样性的变化
. 生态学报 , 24 , 252 -260 . ]
[17]
Li ZQ Ouyang ZY Zeng HQ 2010 ) Assessment methods for territorial biodiversity hotspot based on species richness at broad scale.
Acta Ecologica Sinica , 30 , 1586 -1593 . (in Chinese with English abstract)
[本文引用: 1]
[李智琦 , 欧阳志云 , 曾慧卿 (2010 ) 基于物种的大尺度生物多样性热点研究方法
. 生态学报 , 30 , 1586 -1593 .]
[本文引用: 1]
[18]
Niu KC Liu YN Shen ZH He FL Fang JY 2009 ) Community assembly: the relative importance of neutral theory and niche theory.
Biodiversity Science , 17 , 579 -593 . (in Chinese with English abstract)
[牛克昌 , 刘怿宁 , 沈泽昊 , 何芳良 , 方精云 (2009 ) 群落构建的中性理论和生态位理论
. 生物多样性 , 17 , 579 -593 .]
[19]
Pearman PB Weber D 2007 ) Common species determine richness patterns in biodiversity indicator taxa.
Biological Conservation , 138 , 109 -119 .
[本文引用: 1]
[20]
Pool TK Grenouillet G Villéger S 2014 ) Species contribute differently to the taxonomic, functional, and phylogenetic alpha and beta diversity of freshwater fish communities.
Diversity and Distributions , 20 , 1235 -1244 .
[21]
Raup DM Crick RE 1979 ) Measurement of faunal similarity in paleontology.
Journal of Paleontology , 53 , 1213 -1227 .
[本文引用: 1]
[22]
Šizling AL Šizlingová E Storch D Reif J Gaston KJ 2009 ) Rarity, commonness, and the contribution of individual species to species richness patterns.
The American Naturalist , 174 , 82 -93 .
[本文引用: 2]
[23]
Vázquez LB Gaston KJ 2004 ) Rarity, commonness, and patterns of species richness: the mammals of Mexico.
Global Ecology and Biogeography , 13 , 535 -542 .
[本文引用: 1]
[24]
Vellend M Verheyen K Flinn KM Jacquemyn H Kolb A van Calster H Peterken G Graae BJ Bellemare J Honnay O 2007 ) Homogenization of forest plant communities and weakening of species-environment relationships via agricultural land use.
Journal of Ecology , 95 , 565 -573 .
[本文引用: 2]
[25]
Wright DH Patterson BD Mikkelson GM Cutler A Atmar W 1998 ) A comparative analysis of nested subset patterns of species composition.
Oecologia , 113 , 1 -20 .
[本文引用: 1]
Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist.
1
2011
... 同样地, 稀有种和常见种的不同分布也是导致群落异质性的重要原因.因此, 物种添加(或去除)前后群落异质性的变化也是度量稀有和常见种对物种丰富度格局相对贡献的有效指标.而β多样性既可以有效度量群落的异质性, 还是群落物种丰富度的重要组成成分(Beck et al, 2012 ).关于β多样性的度量方式很多, 但是, 绝大多数β多样性指数的大小常常依赖于α多样性的变化(Anderson et al, 2011 ).例如, 两个样方间的共有种常常随α多样性的增加而增加, 即β多样性随α多样性的增加而减小, 反之亦然(Chase et al, 2011 ).因此, 通过常规的β多样性指数不能有效比较稀有种和常见种二者的效应大小. ...
Diversity partitioning confirms the importance of beta components in tropical rainforest Lepidoptera.
3
2012
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 同样地, 稀有种和常见种的不同分布也是导致群落异质性的重要原因.因此, 物种添加(或去除)前后群落异质性的变化也是度量稀有和常见种对物种丰富度格局相对贡献的有效指标.而β多样性既可以有效度量群落的异质性, 还是群落物种丰富度的重要组成成分(Beck et al, 2012 ).关于β多样性的度量方式很多, 但是, 绝大多数β多样性指数的大小常常依赖于α多样性的变化(Anderson et al, 2011 ).例如, 两个样方间的共有种常常随α多样性的增加而增加, 即β多样性随α多样性的增加而减小, 反之亦然(Chase et al, 2011 ).因此, 通过常规的β多样性指数不能有效比较稀有种和常见种二者的效应大小. ...
... 群落的物种丰富度不仅依赖于α多样性, 而且依赖于β多样性.很多研究表明, β多样性也是很多群落物种丰富度的重要贡献者(Beck et al, 2012 ).本研究结果表明, 稀有种和常见种所引起的β多样性的变化与α多样性变化方向完全相反.同时, 稀有种的加入所引起的β多样性变化较小, 但随着常见种的进入, β多样性急剧下降(图4 ).这说明, 与α多样性一样, 常见种也是β多样性格局的重要贡献者, 这与人们的预期并不一致.群落中的稀有种数量较多且与群落总物种数显著正相关(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 ), 物种丰富度常常呈现随取样面积增加而增加的内嵌式分布格局(Wright et al, 1998 ).一般认为, 两个群落间物种丰富度的差异应该是β多样性间的差异.例如, 对北美树栖甲虫群落物种丰富度的多尺度研究结果表明, 大尺度上的β多样性强烈影响稀有种(物种多度< 0.05%)的丰富度, 稀有种甚至主导了整个群落的丰富度格局(Gering et al, 2003 ).对稀有种主导整个群落物种丰富度格局的一个可能解释是由于稀有种占较大的比例(36%-47%)所致. ...
Common and rare Swedish vertebrates—distribution and habitat preferences.
3
1996
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少.与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献.研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大.同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段.这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 ).不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
... 群落的物种丰富度不仅依赖于α多样性, 而且依赖于β多样性.很多研究表明, β多样性也是很多群落物种丰富度的重要贡献者(Beck et al, 2012 ).本研究结果表明, 稀有种和常见种所引起的β多样性的变化与α多样性变化方向完全相反.同时, 稀有种的加入所引起的β多样性变化较小, 但随着常见种的进入, β多样性急剧下降(图4 ).这说明, 与α多样性一样, 常见种也是β多样性格局的重要贡献者, 这与人们的预期并不一致.群落中的稀有种数量较多且与群落总物种数显著正相关(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 ), 物种丰富度常常呈现随取样面积增加而增加的内嵌式分布格局(Wright et al, 1998 ).一般认为, 两个群落间物种丰富度的差异应该是β多样性间的差异.例如, 对北美树栖甲虫群落物种丰富度的多尺度研究结果表明, 大尺度上的β多样性强烈影响稀有种(物种多度< 0.05%)的丰富度, 稀有种甚至主导了整个群落的丰富度格局(Gering et al, 2003 ).对稀有种主导整个群落物种丰富度格局的一个可能解释是由于稀有种占较大的比例(36%-47%)所致. ...
Hotspots and species diversity.
1
1994
... 稀有种和常见种的定义依赖于研究尺度和研究目的, 一般根据物种的分布范围来定义, 如多度和频度(Curnutt et al, 1994 ; 李智琦等, 2010 ).但是, 如果对群落中的物种进行直接分类, 可能会导致不同种类物种数量的不均衡性, 这不利于物种类别间的比较.为了研究稀有种和常见种对物种丰富度的相对贡献, 本研究采用频度来定义物种的稀有性和常见性, 按照频度由低到高或者由高到底的序列对物种依次排序(Lennon et al, 2004 ; Heegaard et al, 2013 ), 然后根据序列逐一添加(或去除)物种, 通过比较物种添加(或去除)前后所引起的多样性变化程度来确定该物种对物种丰富度格局的相对贡献. ...
Using null models to disentangle variation in community dissimilarity from variation in α-diversity.
3
2011
... 同样地, 稀有种和常见种的不同分布也是导致群落异质性的重要原因.因此, 物种添加(或去除)前后群落异质性的变化也是度量稀有和常见种对物种丰富度格局相对贡献的有效指标.而β多样性既可以有效度量群落的异质性, 还是群落物种丰富度的重要组成成分(Beck et al, 2012 ).关于β多样性的度量方式很多, 但是, 绝大多数β多样性指数的大小常常依赖于α多样性的变化(Anderson et al, 2011 ).例如, 两个样方间的共有种常常随α多样性的增加而增加, 即β多样性随α多样性的增加而减小, 反之亦然(Chase et al, 2011 ).因此, 通过常规的β多样性指数不能有效比较稀有种和常见种二者的效应大小. ...
... 根据Vellend等(2007)的方法, 本研究引入一种基于概率的β多样性指数βRC , 该指数通过随机方式排除了不同样方间的α多样性差异, 可以反映β多样性的真实变化.其计算过程如下: 假设群落由n 个样方组成, 其中, 样方x 有i 个物种, 样方y 有j 个物种.首先计算两个样方的实际共有物种数, 然后每次从“区域种库”中随机抽取i 和j 个物种分别赋予x 和y 两个样方, 计算两个样方的期望共有物种数.重复此过程9,999次, 以这9,999次重复中期望共有物种数≥实际共有物种数的比例即βRC 值作为样方x 和样方y 间的真实β多样性度量.n 个样方两两间形成的βRC 多样性矩阵的平均值即为该群落的β多样性(Vellend et al, 2007 ).其中, 所有取样单元中的物种整合形成一个“区域种库”, 种库中物种被选择的机率依赖于物种频度大小.由于βRC 指数不依赖于α多样性的大小(Raup & Crick, 1979 ; Chase et al, 2011 ), 因此可 以有效度量稀有和常见种对β多样性格局的相对 贡献. ...
... 目前, 关于常见种和稀有种对物种丰富度格局贡献差异的原因仍不是很清楚, 一个可能的解释是常见种是群落物种丰富度的信号(signals)贡献者, 而稀有种是物种丰富度的噪音(noise)贡献者(Vázquez & Gaston, 2004 ).尽管βRC 多样性指数考虑了α多样性的变化对β多样性的影响, 但是该指数仍具一定的局限性(Chase et al, 2011 ).首先, 该指数来自于古生物学, 并没有很好地考虑群落内不同物种多度的差异, 这对于群落生态学来说是一种不足; 第二, 该方法没有考虑物种在大尺度上的出现概率(Raup & Crick, 1979 ; Vellend et al, 2007 ).一般来说, 分布范围广的物种出现的概率更高, 但是βRC 多样性指数的随机模型没有考虑到这个权重.另外, 种库大小也将明显影响βRC 多样性指数的大小(Gotelli & Colwell, 2001 ). ...
Dispersal limitation and environmental heterogeneity shape scale-dependent diversity patterns in plant communities.
2
2006
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
... ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Global scale macroecology: interactions between population size, geographic range size and body size in the Anseriformes.
2
1996
... 尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少.与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献.研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大.同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段.这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 ).不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
... 群落的物种丰富度不仅依赖于α多样性, 而且依赖于β多样性.很多研究表明, β多样性也是很多群落物种丰富度的重要贡献者(Beck et al, 2012 ).本研究结果表明, 稀有种和常见种所引起的β多样性的变化与α多样性变化方向完全相反.同时, 稀有种的加入所引起的β多样性变化较小, 但随着常见种的进入, β多样性急剧下降(图4 ).这说明, 与α多样性一样, 常见种也是β多样性格局的重要贡献者, 这与人们的预期并不一致.群落中的稀有种数量较多且与群落总物种数显著正相关(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 ), 物种丰富度常常呈现随取样面积增加而增加的内嵌式分布格局(Wright et al, 1998 ).一般认为, 两个群落间物种丰富度的差异应该是β多样性间的差异.例如, 对北美树栖甲虫群落物种丰富度的多尺度研究结果表明, 大尺度上的β多样性强烈影响稀有种(物种多度< 0.05%)的丰富度, 稀有种甚至主导了整个群落的丰富度格局(Gering et al, 2003 ).对稀有种主导整个群落物种丰富度格局的一个可能解释是由于稀有种占较大的比例(36%-47%)所致. ...
Biodiversity and extinction: the importance of being common.
1
2008
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Additive partitioning of species diversity across multiple spatial scales: implications for regional conservation of biodiversity.
3
2003
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 群落的物种丰富度不仅依赖于α多样性, 而且依赖于β多样性.很多研究表明, β多样性也是很多群落物种丰富度的重要贡献者(Beck et al, 2012 ).本研究结果表明, 稀有种和常见种所引起的β多样性的变化与α多样性变化方向完全相反.同时, 稀有种的加入所引起的β多样性变化较小, 但随着常见种的进入, β多样性急剧下降(图4 ).这说明, 与α多样性一样, 常见种也是β多样性格局的重要贡献者, 这与人们的预期并不一致.群落中的稀有种数量较多且与群落总物种数显著正相关(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 ), 物种丰富度常常呈现随取样面积增加而增加的内嵌式分布格局(Wright et al, 1998 ).一般认为, 两个群落间物种丰富度的差异应该是β多样性间的差异.例如, 对北美树栖甲虫群落物种丰富度的多尺度研究结果表明, 大尺度上的β多样性强烈影响稀有种(物种多度< 0.05%)的丰富度, 稀有种甚至主导了整个群落的丰富度格局(Gering et al, 2003 ).对稀有种主导整个群落物种丰富度格局的一个可能解释是由于稀有种占较大的比例(36%-47%)所致. ...
... 与此研究相似, 本文中乔、灌、草三层的物种分布格局也均为稀有种占绝大比例的右偏分布, 但研究结论却不完全一致, 可能原因如下: (1)两个研究中定义稀有种和常见种的方式不同, 尽管分布范围广的物种多度也相对较大, 但是局域群落上的优势种与大尺度上的常见种是不同的; (2)物种多样性格局是尺度依赖的(Gering et al, 2003 ; Heegaard et al, 2013 ), 研究稀有种和常见种对多样性格局的相对贡献应该充分考虑空间尺度的影响, 这也是下一步应该亟需开展的重要工作; (3)两个研究中使用的β多样性度量指数不同, 如前所述, 使用依赖于α 多样性变化的β多样性指数不能真实度量β多样性 大小. ...
Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness.
1
2001
... 目前, 关于常见种和稀有种对物种丰富度格局贡献差异的原因仍不是很清楚, 一个可能的解释是常见种是群落物种丰富度的信号(signals)贡献者, 而稀有种是物种丰富度的噪音(noise)贡献者(Vázquez & Gaston, 2004 ).尽管βRC 多样性指数考虑了α多样性的变化对β多样性的影响, 但是该指数仍具一定的局限性(Chase et al, 2011 ).首先, 该指数来自于古生物学, 并没有很好地考虑群落内不同物种多度的差异, 这对于群落生态学来说是一种不足; 第二, 该方法没有考虑物种在大尺度上的出现概率(Raup & Crick, 1979 ; Vellend et al, 2007 ).一般来说, 分布范围广的物种出现的概率更高, 但是βRC 多样性指数的随机模型没有考虑到这个权重.另外, 种库大小也将明显影响βRC 多样性指数的大小(Gotelli & Colwell, 2001 ). ...
Contribution of rare and common species to richness patterns at local scales.
3
2013
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 稀有种和常见种的定义依赖于研究尺度和研究目的, 一般根据物种的分布范围来定义, 如多度和频度(Curnutt et al, 1994 ; 李智琦等, 2010 ).但是, 如果对群落中的物种进行直接分类, 可能会导致不同种类物种数量的不均衡性, 这不利于物种类别间的比较.为了研究稀有种和常见种对物种丰富度的相对贡献, 本研究采用频度来定义物种的稀有性和常见性, 按照频度由低到高或者由高到底的序列对物种依次排序(Lennon et al, 2004 ; Heegaard et al, 2013 ), 然后根据序列逐一添加(或去除)物种, 通过比较物种添加(或去除)前后所引起的多样性变化程度来确定该物种对物种丰富度格局的相对贡献. ...
... 与此研究相似, 本文中乔、灌、草三层的物种分布格局也均为稀有种占绝大比例的右偏分布, 但研究结论却不完全一致, 可能原因如下: (1)两个研究中定义稀有种和常见种的方式不同, 尽管分布范围广的物种多度也相对较大, 但是局域群落上的优势种与大尺度上的常见种是不同的; (2)物种多样性格局是尺度依赖的(Gering et al, 2003 ; Heegaard et al, 2013 ), 研究稀有种和常见种对多样性格局的相对贡献应该充分考虑空间尺度的影响, 这也是下一步应该亟需开展的重要工作; (3)两个研究中使用的β多样性度量指数不同, 如前所述, 使用依赖于α 多样性变化的β多样性指数不能真实度量β多样性 大小. ...
1
2001
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Disentangling the drivers of β diversity along latitudinal and elevational gradients.
1
2011
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Commonness and rarity of plants in a reserve network: just two faces of the same coin.
2
2014
... 尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少.与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献.研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大.同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段.这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 ).不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
... )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
Contribution of rarity and commonness to patterns of species richness.
6
2004
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
... ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 稀有种和常见种的定义依赖于研究尺度和研究目的, 一般根据物种的分布范围来定义, 如多度和频度(Curnutt et al, 1994 ; 李智琦等, 2010 ).但是, 如果对群落中的物种进行直接分类, 可能会导致不同种类物种数量的不均衡性, 这不利于物种类别间的比较.为了研究稀有种和常见种对物种丰富度的相对贡献, 本研究采用频度来定义物种的稀有性和常见性, 按照频度由低到高或者由高到底的序列对物种依次排序(Lennon et al, 2004 ; Heegaard et al, 2013 ), 然后根据序列逐一添加(或去除)物种, 通过比较物种添加(或去除)前后所引起的多样性变化程度来确定该物种对物种丰富度格局的相对贡献. ...
... 此方法以相关系数为测度指标, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的贡献.其中, 群落由样方组成, 每个样方的物种数即为该群落的物种丰富度, n 个样方可以理解为群落的n 次重复, 添加或去除物种并不改变样方个数而只改变每个样方中的物种数.因而可以度量两个群落间的相关系数大小并通过相关系数的变化间接度量稀有种和常见种的相对贡献, 这也是研究该问题的经典方法(Lennon et al, 2004 ). ...
... 尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少.与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献.研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大.同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段.这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 ).不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
子午岭植被自然恢复过程中植物多样性的变化
2004
子午岭植被自然恢复过程中植物多样性的变化
2004
基于物种的大尺度生物多样性热点研究方法
1
2010
... 黄土高原是我国特有的地理区域, 该区环境恶劣, 资源贫乏.由于水土流失以及人为活动干扰, 其生态系统正处于极度退化的状态, 因此, 加速该地区退化生态系统的恢复与重建是当务之急(李裕元和邵明安, 2004 ), 而物种多样性的恢复与保护是该区生态系统恢复与重建的重要基础.因此, 在黄土高原进行物种多样性格局研究具有非常重要的理论和现实意义.本研究以该区的顶极群落辽东栎(Quercus wutaishanica )林为模式群落, 根据物种频度序列通过逐一添加(或去除)稀有种或常见种的方式, 探讨由此引起的群落总体物种多样性及其成分(α多样性和β多样性)的变化程度, 以度量稀有种和常见种对物种丰富度格局的相对贡献, 旨在验证常见种对物种多样性格局决定性假说在黄土高原的适用性, 比较稀有种和常见种对α多样性和β多样性这两种成分相对贡献的差异性. ...
基于物种的大尺度生物多样性热点研究方法
1
2010
... 黄土高原是我国特有的地理区域, 该区环境恶劣, 资源贫乏.由于水土流失以及人为活动干扰, 其生态系统正处于极度退化的状态, 因此, 加速该地区退化生态系统的恢复与重建是当务之急(李裕元和邵明安, 2004 ), 而物种多样性的恢复与保护是该区生态系统恢复与重建的重要基础.因此, 在黄土高原进行物种多样性格局研究具有非常重要的理论和现实意义.本研究以该区的顶极群落辽东栎(Quercus wutaishanica )林为模式群落, 根据物种频度序列通过逐一添加(或去除)稀有种或常见种的方式, 探讨由此引起的群落总体物种多样性及其成分(α多样性和β多样性)的变化程度, 以度量稀有种和常见种对物种丰富度格局的相对贡献, 旨在验证常见种对物种多样性格局决定性假说在黄土高原的适用性, 比较稀有种和常见种对α多样性和β多样性这两种成分相对贡献的差异性. ...
Common species determine richness patterns in biodiversity indicator taxa.
1
2007
... 稀有种和常见种的定义依赖于研究尺度和研究目的, 一般根据物种的分布范围来定义, 如多度和频度(Curnutt et al, 1994 ; 李智琦等, 2010 ).但是, 如果对群落中的物种进行直接分类, 可能会导致不同种类物种数量的不均衡性, 这不利于物种类别间的比较.为了研究稀有种和常见种对物种丰富度的相对贡献, 本研究采用频度来定义物种的稀有性和常见性, 按照频度由低到高或者由高到底的序列对物种依次排序(Lennon et al, 2004 ; Heegaard et al, 2013 ), 然后根据序列逐一添加(或去除)物种, 通过比较物种添加(或去除)前后所引起的多样性变化程度来确定该物种对物种丰富度格局的相对贡献. ...
Species contribute differently to the taxonomic, functional, and phylogenetic alpha and beta diversity of freshwater fish communities.
2014
Measurement of faunal similarity in paleontology.
1
1979
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Rarity, commonness, and the contribution of individual species to species richness patterns.
2
2009
... 基于相关系数法的很多研究发现, 添加或去除常见种对相关系数的影响较大, 而添加或去除稀有种对相关系数的影响相对较小, 表明常见种决定丰富度格局(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Heegaard et al, 2013 ), 这似乎与人们的长期认识相矛盾.一般认为, 物种丰富度格局主要由大量分布范围较窄且物种频度(多度)较低的稀有种决定, 而不是由少数分布范围广且物种频度(多度)大的常见种决定(Berg & Tjernberg, 1996 ).因此, 常见种决定丰富度格局的普适性仍需进一步验证.同时, 区域上的物种丰富度(γ多样性)由局域样方内的α多样性和样方间的β多样性两种成分共同组成(Gering et al, 2003 ).但是, 很多类似的研究都集中在α多样性水平上而忽略了β多样性水平的研究.事实上, β多样性也是很多生态系统物种多样性维持的重要成分(Beck et al, 2012 ), 特别是在研究物种多样性格局时应引起人们的足够重视.稀有种和常见种对α多样性和β多样性这两种成分的相对贡献的大小仍是一个悬而未决的问题. ...
... 尽管关于物种丰富度格局的研究较多, 但是关于群落内物种组成对丰富度格局影响的研究仍然相对较少.与以往的研究不同, 本研究将总体物种丰富度分解为α多样性和β多样性两种成分, 进一步度量了稀有种和常见种对这两种多样性成分的相对贡献.研究结果表明, 无论是稀有-常见种序列还是常见-稀有种序列, “新”群落与原群落间物种多样性相关性的急剧变化都表现在常见种逐渐加入的阶段, 这一段的长度也明显依赖于常见种的数量, 即常见种越多, 相关系数急剧增加的区域就越大.同时, 无论是稀有-常见种序列还是常见-稀有种序列, 由于物种的加入而引起的“新”群落与原群落间物种多样性成分(α多样性和β多样性)的变化也均表现在常见种逐渐加入的阶段.这与很多研究结果相似, 说明常见种的确是植物群落物种丰富度格局的重要贡献者(Lennon et al, 2004 ; Pearman & Weber, 2007 ; Landi & Chiarucci, 2014 ).不支持群落物种丰富度格局主要由大量分布范围较窄且物种多度(频度)较低的稀有种决定, 而不是由少数分布范围广的常见种决定(Berg & Tjernberg, 1996 ; Gaston & Blackburn, 1996 )的结论.因此, 常见种是群落物种丰富度格局的指示者, 也应该是优先保护的物种(Landi & Chiarucci, 2014 ). ...
Rarity, commonness, and patterns of species richness: the mammals of Mexico.
1
2004
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
Homogenization of forest plant communities and weakening of species-environment relationships via agricultural land use.
2
2007
... 根据Vellend等(2007)的方法, 本研究引入一种基于概率的β多样性指数βRC , 该指数通过随机方式排除了不同样方间的α多样性差异, 可以反映β多样性的真实变化.其计算过程如下: 假设群落由n 个样方组成, 其中, 样方x 有i 个物种, 样方y 有j 个物种.首先计算两个样方的实际共有物种数, 然后每次从“区域种库”中随机抽取i 和j 个物种分别赋予x 和y 两个样方, 计算两个样方的期望共有物种数.重复此过程9,999次, 以这9,999次重复中期望共有物种数≥实际共有物种数的比例即βRC 值作为样方x 和样方y 间的真实β多样性度量.n 个样方两两间形成的βRC 多样性矩阵的平均值即为该群落的β多样性(Vellend et al, 2007 ).其中, 所有取样单元中的物种整合形成一个“区域种库”, 种库中物种被选择的机率依赖于物种频度大小.由于βRC 指数不依赖于α多样性的大小(Raup & Crick, 1979 ; Chase et al, 2011 ), 因此可 以有效度量稀有和常见种对β多样性格局的相对 贡献. ...
... 目前, 关于常见种和稀有种对物种丰富度格局贡献差异的原因仍不是很清楚, 一个可能的解释是常见种是群落物种丰富度的信号(signals)贡献者, 而稀有种是物种丰富度的噪音(noise)贡献者(Vázquez & Gaston, 2004 ).尽管βRC 多样性指数考虑了α多样性的变化对β多样性的影响, 但是该指数仍具一定的局限性(Chase et al, 2011 ).首先, 该指数来自于古生物学, 并没有很好地考虑群落内不同物种多度的差异, 这对于群落生态学来说是一种不足; 第二, 该方法没有考虑物种在大尺度上的出现概率(Raup & Crick, 1979 ; Vellend et al, 2007 ).一般来说, 分布范围广的物种出现的概率更高, 但是βRC 多样性指数的随机模型没有考虑到这个权重.另外, 种库大小也将明显影响βRC 多样性指数的大小(Gotelli & Colwell, 2001 ). ...
A comparative analysis of nested subset patterns of species composition.
1
1998
... 物种多样性格局及其形成机制一直是植物生态学家关注的焦点(Lennon et al, 2004 ; Freestone & Inouye, 2006 ; Kraft et al, 2011 ).很多研究表明, 群落物种多样性格局的形成不仅取决于生态过程, 如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程(Hubbell, 2001 ; Freestone & Inouye, 2006 ; 牛克昌等, 2009 ), 还依赖于群落中的物种组成, 如不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献等(Lennon et al, 2004 ; Šizling et al, 2009 ; Pool et al, 2014 ).其中, 有关稀有种和常见种对物种多样性格局相对贡献的研究较多, 但也颇具争议(Gaston, 2008 ).这些研究主要采用相关性分析法, 即通过添加或去除稀有种(或常见种)形成一个“新”群落, 然后比较“新”群落物种丰富度与原群落物种丰富度间相关系数的变化程度来间接度量稀有种或常见种对群落总体物种丰富度的相对贡献. ...
 , 李娜
, 李娜

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}