欧阳明, 杨清培 , 陈昕, 杨光耀, 施建敏, 方向民
, 陈昕, 杨光耀, 施建敏, 方向民
江西农业大学江西省竹子种质资源与利用重点实验室, 南昌 330045
Ming Ouyang, Qingpei Yang, Xin Chen, Guangyao Yang, Jianmin Shi, Xiangmin Fang
通讯作者:
收稿日期: 2015-10-25
接受日期: 2016-04-19
网络出版日期: 2016-06-20
版权声明: 2016 《生物多样性》编辑部 版权所有.《生物多样性》编辑部
基金资助:
展开
摘要
毛竹(Phyllostachys edulis)向邻近次生常绿阔叶林扩张现象明显, 极大地影响了常绿阔叶林的生态功能, 但关于其扩张对常绿阔叶林的群落结构与生物多样性影响的后效研究较少。本文采用时空替代法, 在江西井冈山国家级自然保护区沿毛竹扩张方向, 依次设置毛竹林、竹阔混交林和常绿阔叶林样地, 比较分析了扩张前后群落物种相似性、群落结构和多样性指数等特征。结果表明: (1)毛竹林与常绿阔叶林乔木层、灌木层和草本层的Bray-Curtis相似性指数很小, 分别为0.003、0.046和0.030。(2)毛竹林的垂直结构呈“>”型, 高度12-14 m区间的多度百分比达33.3%, 径级结构集中分布于5-10 cm区间, 达总数的90.0%; 常绿阔叶林的垂直结构为“L”型, 高度2-4 m的物种数占54.3%, 径级分布范围较广, 4个较大径级区间的平均百分比为10.3%。(3)乔木层的Shannon-Wiener指数由常绿阔叶林的2.56降至毛竹林的0.06, 降幅高达98%; 灌木层也由2.58降至2.03, 降幅21%。以上结果说明, 毛竹扩张会导致次生常绿阔叶林群落组成和结构简化、物种多样性下降, 对森林生态系统功能产生负面影响。
关键词:
Abstract
The expansion of Phyllostachys edulis into the adjacent secondary evergreen broad-leaved forest (EBF) is obvious and greatly affects its ecological function. Little research has examined its effects on community structure and biodiversity. We comparatively analyzed the characteristics of species composition, community structure and diversity before and after the expansion of P. edulis forest (PEF), P. edulis-broad-leaved mixed forest (PBMF) and EBF along a gradient of P. edulis expansion in the Jinggangshan National Nature Reserve in Jiangxi Province using a space for time substitution method. Results indicated that: (1) The Bray-Curtis similarity index values of the tree layer, shrub layer and herb layer between PEF and EBF were 0.003, 0.046 and 0.030, respectively. (2) The PEF vertical structure showed a “>” type and the abundance percentage was 33.3% in 12-14 m interval, its diameter at breast height (DBH) class structure concentrated distribution in 5-10 cm interval, whose percentage was as high as 90.0%; while the EBF vertical structure showed a “L” type and the abundance percentages was 54.3% in 2-4 m interval, its DBH class distribution range was relatively wide, the average percentage of four larger diameter grades was 10.3%. (3) The Shannon-Wiener index value in the tree layer declined from 2.56 in EBF to 0.06 in PEF, with a reduction of 98%. In the shrub layer, the index value dropped from 2.58 to 2.03, declining 21%. We suggest that the expansion of P. edulis simplified the community composition and structure of the secondary evergreen broad-leaved forest and reduced species diversity, which can cause adverse impacts on forest ecosystem functioning.
Keywords:
竹子扩张引起的森林生态系统功能退化与景观变化正逐渐受到生态学家的关注(Lima et al, 2012; 杨清培等, 2015)。我国毛竹(Phyllostachys edulis)林面积已达400万ha以上, 且呈持续增长的趋势(Jiang, 2007)。毛竹是我国特有的乔木状散生竹种, 广泛分布于亚热带地区, 其株高可达20 m, 胸径可达20 cm, 具有生长繁殖速度快、适应改造能力强的特点(耿伯介和王正平, 1996)。虽然毛竹作为重要的经济竹种, 在我国林农增收和竹产业经济发展中起着重要作用, 但作为一类入侵性很强的本土植物, 它自身的生物学特性使其扩张潜力巨大(白尚斌等, 2013)。许多研究发现, 毛竹正不断向邻近群落扩张, 引发了许多生态环境问题(Isagi & Torii, 1997; 丁丽霞等, 2006; 吴家森等, 2008)。然而, 因其较高的经济价值, 人们忽视了毛竹扩张造成的生态影响。
常绿阔叶林是亚热带地区的地带性植被, 集中分布在我国南方, 适宜生境与毛竹林相似(吴征镒, 1980)。常绿阔叶林物种组成多样、群落结构复杂, 在调节气候、涵养水源和保持水土等环境效益方面发挥着重要作用(宋永昌等, 2013)。然而, 毛竹扩张对该区常绿阔叶林的严重威胁使其生态功能下降, 毛竹扩张引发的生态负面效应逐渐受到关注(杨怀等, 2010; Fukushima et al, 2015; Shinohara & Otsuki, 2015)。目前已有研究分别从土壤理化性质、凋落物水文特征、植物多样性和生态景观格局等方面进行了初步分析(白尚斌等, 2013; 宋庆妮等, 2015; 杨清培等, 2015), 但关于毛竹扩张对常绿阔叶林群落特征影响的生态效应评价仍重视不够。物种组成、空间结构和多样性作为群落基本特征, 共同决定着森林的生态系统功能(Chapin III et al, 2000; Huang et al, 2003)。然而, 自然条件下以上特征很容易受侵入物种的种群发展影响, 尤其是高大侵入种的种群扩张改造作用更强(Hejda et al, 2009)。
为此, 本文采用时空替代法, 在江西井冈山国家级自然保护区选择典型的毛竹向常绿阔叶林扩张的样地, 对群落特征进行全面调查, 并比较分析了毛竹林、竹阔混交林和次生常绿阔叶林乔木层、灌木层和草本层的物种组成、径级结构和物种多样性等特征, 以期为客观评价毛竹扩张对常绿阔叶林的生态后效提供参考。
江西井冈山国家级自然保护区(26°13'-26°52' N, 113°59'-114°18' E)始建于1981年, 2000年批准为国家级自然保护区, 主要保护对象是中亚热带湿润常绿阔叶林生态系统及其生物多样性。该保护区位于东南丘陵盆地区, 属中亚热带季风气候(张继平等, 2014)。该地四季分明, 水热条件充沛, 平均气温14.2℃, 最热月平均气温23.9℃ (7月), 极端最高温36.7℃; 最冷月平均气温3.4℃ (1月), 极端最低温-11.0℃; 年平均降水量1,889.8 mm; 海拔202-2,120.4 m。土壤以山地黄壤为主, 土层厚度一般在50-80 cm, 土质疏松、肥沃、湿润(邓贤兰等, 2003; 张继平等, 2014)。该区地带性植被为常绿阔叶林, 同时也是毛竹适生区。保护区建立以前, 由于长期的人类采伐和自然干扰, 原始常绿阔叶林大面积退化为次生林, 自然保护区建立后, 采取封山育林的植被恢复措施, 次生常绿阔叶林处于向原始林正向演替阶段, 但同时也因遭受邻近毛竹自然扩张的严重干扰, 形成了大量竹阔混交林和毛竹林。
2014年8月, 在保护区内选择3个典型的毛竹向次生常绿阔叶林扩张的界面, 每个界面宽20 m, 长90 m, 根据群落特征依次划分为毛竹林、竹阔混交林和阔叶林。每个群落设置1个20 m × 20 m的样方用于乔木层树种监测, 在样地的对角线位置设置2个5 m × 5 m的样方用于灌木层物种监测, 在样地中心和4角设置5个1 m × 1 m的样方用于草本层物种监测。样地海拔高度为850-950 m, 阔叶林为正在恢复中的次生常绿阔叶林, 乔木层优势种主要为红楠(Machilus thunbergii), 林龄40-50年, 伴生树种主要有交让木(Daphniphyllum macropodum)、赤杨叶(Alniphyllum fortunei)等。乔木层高度17.0 m, 平均胸径15.8 cm, 密度1,000株/ha, 郁闭度0.85。竹阔混交林为毛竹向阔叶林扩张6-7年后形成, 竹木数量比为8:1, 其中毛竹高约15 m, 密度约4,200株/ha。毛竹林为约30年前自然扩张形成的纯林, 密度约6,200株/ha, 平均胸径8.0 cm, 平均高度12.5 m。
乔木层: 起测胸径5 cm, 记录物种、胸径、高度和多度等; 灌木层: 测定胸径小于5 cm的木本植物, 包括乔木的幼树和灌木, 记录种名、地径、高度和多度等; 草本层: 记录种名、高度和盖度等。
1.3.1 群落物种组成测度
重要值: IV = (相对多度+相对频度+相对显著度+相对高度)/4 (1)
群落相似性的Bray-Curtis指数(Bray & Curtis, 1957; 马克平等, 1995):
Cn = 2jN/(aN + bN) (2)
jN = 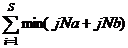
式中, aN为样地A的物种数目, bN为样地B的物种数目, jN为样地A(jNa)和样地B(jNb)共有种中个体数目较小者之和。
1.3.2 群落结构测度
群落垂直结构: 将群落乔木层和灌木层所有植株的高度按2 m的组间距划分成11个组, 分别计算每个组内植株多度占总多度的百分比, 绘制频率分布图。
群落径级结构: 将群落乔木层所有植株的胸径按5 cm的组间距分成8个组, 分别计算每个组内植株多度占总多度的百分比, 绘制频率分布图。
1.3.3 物种多样性测度(马克平和刘玉明, 1994)
物种丰富度: R = S (4)
物种丰富度的Margalef指数: 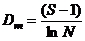
Simpson指数: 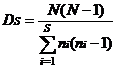
Shannon-Wiener指数: 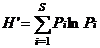
Pielou均匀度指数: 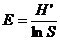
生态优势度指数: 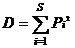
式中, S为物种数目, N为所有物种的个体数之和, ni为第i个种的个体数, Pi为第i种的个体数占所有个体总数N的比例。
采用单因素方差分析(One-Way ANOVA)和最小显著差异法(LSD)比较各多样性指数在毛竹林、竹阔混交林和常绿阔叶林间的差异显著性。分别通过SPSS 19.0和Origin 9.0进行数据分析和绘图。
由表1可知, 3个群落的立竹度和立木数差别较大。毛竹扩张前后, 群落立竹度和总立木树明显上升, 每公顷分别增加为6,233和4,466株; 而其他树种立木数则显著下降, 单位面积减少了1,767株/ha, 降幅高达94.6%。说明毛竹扩张过程中其增加的个体数量显著大于减少的阔叶树种株数, 导致群落总立木数明显增加。
由表2可知, 3个群落的物种组成和数量特征均差异明显。对于乔木层来说, 毛竹林的优势种毛竹的重要值高达87.02; 竹阔混交林的优势种也是毛竹, 重要值达57.28, 伴生种红楠、杉木(Cunning- hamia lanceolata)和青榨槭(Acer davidii)的重要值分别是11.51、9.98和3.83; 常绿阔叶林中红楠占优势, 重要值为30.22, 伴生树种主要包括杉木、赤杨叶、青榨槭、黄丹木姜子(Litsea elongata)和交让木等。对于灌木层而言, 毛竹林的优势种仍为毛竹, 重要值达51.24, 伴生种主要有赤杨叶、青榨槭和香港四照花(Dendrobenthamia hongkongensis)等; 竹阔混交林的物种分布较均匀, 重要值前4位的分别是鹿角杜鹃(Rhododendron latoucheae)、油茶(Camellia oleifera)、红楠和香港四照花; 常绿阔叶林中鹿角杜鹃略占优, 重要值为19.86, 其他物种主要包括黄丹木姜子、油茶和红楠等。最后, 毛竹林、竹阔混交林和常绿阔叶林的草本层优势植物分别为翠云草(Selaginella uncinata)和麦冬(Ophiopogon japonicus)、星蕨(Microsorum punctatum)、短毛熊巴掌(Phyllagathis cavaleriei)。说明毛竹扩张不仅改变了常绿阔叶林的物种组成, 而且还影响了物种在群落中的地位。
由表3可知, 不同群落间的相似性系数差别较大。常绿阔叶林与竹阔混交林3个层次的Bray-Curtis相似性指数范围为0.219-0.463, 而与毛竹林的相似性系数波动在0.003-0.046。说明常绿阔叶林与竹阔混交林群落物种组成较相似, 而与毛竹林差异较大。可见, 毛竹扩张改变了常绿阔叶林的物种组成及数量特征。
表1 毛竹扩张对次生常绿阔叶林群落立木数的影响(株/ha)
Table 1 Effect of the expansion of Phyllostachys edulis on stem number in the secondary evergreen broad-leaved forest (ind./ha)
| 扩张阶段 Expansion phase | 群落类型 Community type | 毛竹立竹度 Density of bamboo stand | 其他树种立木数 Stem number of other trees | 总立木数 Total stem number |
|---|---|---|---|---|
| 前期 Early stage | 常绿阔叶林 EBF | 0 | 1,867 | 1,867 |
| 中期 Middle stage | 竹阔混交林 PBMF | 3,667 | 1,017 | 4,684 |
| 后期 Later stage | 毛竹林 PEF | 6,233 | 100 | 6,333 |
| 扩张前后的变化 Δ | 6,233 | -1,767 | 4,466 |
表2 毛竹扩张对次生常绿阔叶林主要物种组成与重要值的影响
Table 2 Effects of the expansion of Phyllostachys edulis on main species composition and importance value in the secondary evergreen broad-leaved forest
| 层次 Layer | 群落 Community | 物种 Species | 重要值 Importance value |
|---|---|---|---|
| 乔木层 Tree | 毛竹林 Phyllostachys edulis forest | 毛竹 Phyllostachys edulis | 87.02 |
| 杉木 Cunninghamia lanceolata | 8.73 | ||
| 黄丹木姜子 Litsea elongata | 2.37 | ||
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest | 毛竹 Phyllostachys edulis | 57.28 | |
| 红楠 Machilus thunbergii | 11.51 | ||
| 杉木 Cunninghamia lanceolata | 9.98 | ||
| 青榨槭 Acer davidii | 3.83 | ||
| 常绿阔叶林 Evergreen broad-leaved forest | 红楠 Machilus thunbergii | 30.22 | |
| 杉木 Cunninghamia lanceolata | 14.06 | ||
| 赤杨叶 Alniphyllum fortunei | 8.33 | ||
| 青榨槭 Acer davidii | 7.65 | ||
| 黄丹木姜子 Litsea elongata | 4.22 | ||
| 交让木 Daphniphyllum macropodum | 3.23 | ||
| 灌木层 Shrub | 毛竹林 Phyllostachys edulis forest | 毛竹 Phyllostachys edulis | 51.24 |
| 赤杨叶 Alniphyllum fortunei | 8.98 | ||
| 青榨槭 Acer davidii | 6.43 | ||
| 香港四照花 Dendrobenthamia hongkongensis | 5.64 | ||
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest | 鹿角杜鹃 Rhododendron latoucheae | 15.21 | |
| 油茶 Camellia oleifera | 8.44 | ||
| 红楠 Machilus thunbergii | 7.99 | ||
| 香港四照花 Dendrobenthamia hongkongensis | 7.27 | ||
| 常绿阔叶林 Evergreen broad-leaved forest | 鹿角杜鹃 Rhododendron latoucheae | 19.86 | |
| 黄丹木姜子 Litsea elongata | 8.84 | ||
| 油茶 Camellia oleifera | 7.92 | ||
| 红楠 Machilus thunbergii | 7.42 | ||
| 草本层 Herb | 毛竹林 Phyllostachys edulis forest | 翠云草 Selaginella uncinata | 19.99 |
| 麦冬 Ophiopogon japonicus | 15.39 | ||
| 星蕨 Microsorum punctatum | 8.54 | ||
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest | 星蕨 Microsorum punctatum | 28.54 | |
| 翠云草 Selaginella uncinata | 8.53 | ||
| 短毛熊巴掌 Phyllagathis cavaleriei | 6.58 | ||
| 常绿阔叶林 Evergreen broad-leaved forest | 短毛熊巴掌 Phyllagathis cavaleriei | 25.83 | |
| 淡竹叶 Lophatherum gracile | 11.13 | ||
| 薹草 Carex tristachya | 10.79 |
表3 3个群落乔木层、灌木层和草本层的Bray-Curtis相似性系数比较
Table 3 Comparison of Bray-Curtis similarity coefficient between three community types in tree, shrub and herb layers.
| 层次 Layer | 群落 Community | 竹阔混交林 PBMF | 毛竹林 Phyllostachys edulis forest |
|---|---|---|---|
| 乔木层 Tree | 常绿阔叶林 Evergreen broad-leaved forest | 0.219 | 0.003 |
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest (PBMF) | 0.626 | ||
| 灌木层 Shrub | 常绿阔叶林 Evergreen broad-leaved forest | 0.383 | 0.046 |
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest | 0.202 | ||
| 草本层 Herb | 常绿阔叶林 Evergreen broad-leaved forest | 0.463 | 0.030 |
| 竹阔混交林 Phyllostachys edulis-broad-leaved mixed forest | 0.319 |
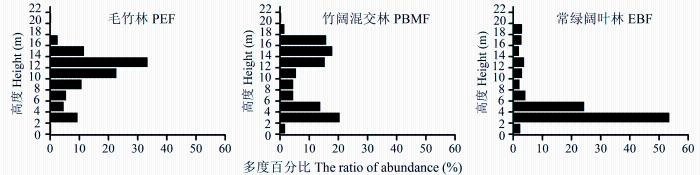
2.2.1 对群落垂直结构的影响
由图1可知, 3种群落的垂直结构差异明显, 毛竹林呈“>”型, 竹阔混交林呈“<”型, 而常绿阔叶林则为“L”型。其中毛竹林的物种高度主要集中于8-10、10-12、12-14和14-16 m区间, 多度百分比分别为10.7%、22.6%、33.3%和11.5%; 竹阔混交林的物种高度主要分布在2-4、4-6、12-14、14-16和16-18 m区间, 多度百分比分别为20.3%、13.6%、15.2%、17.8%和15.7%, 共占82.6%; 常绿阔叶林的物种高度在2-4 m和4-6 m区间的百分比分别达53.4%和24.2%, 而上层7个区间的高度分布百分比则差别不大, 平均为2.9%。说明毛竹向常绿阔叶林扩张过程中, 会导致群落的分层现象逐渐减弱, 使群落垂直结构单一, 主要集中在12-14 m。
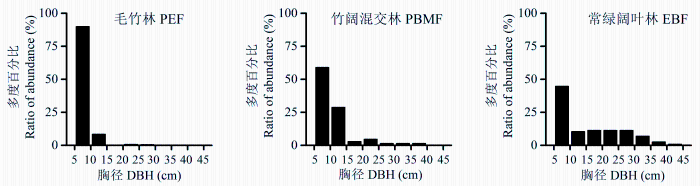
2.2.2 对群落径级结构的影响
由图2可知, 3种群落的径级结构明显不同, 毛竹林呈“L”型, 竹阔混交林和常绿阔叶林则呈倒“J”型。其中毛竹林物种径级主要集中于5-10 cm和10-15 cm区间, 多度百分比分别达90.0%和8.4%; 竹阔混交林的物种径级主要分布在5-10、10-15、15-20和20-25 cm区间, 百分比分别为59.1%、28.8%、2.8%和4.6%, 共占95.4%; 而常绿阔叶林物种径级分布范围较广, 5-10、10-15、15-20、20-25、25-30和30-35 cm区间的多度百分比分别占44.7%、10.5%、11.4%、11.4%、11.4%和7.0%, 总和为96.5%。说明毛竹在向邻近阔叶林扩张的过程中, 大径级的个体数量会逐渐减少, 导致群落径级结构变窄, 主要分布于小径级(如5-10 cm)区间。
图1 毛竹扩张对次生常绿阔叶林群落垂直结构的影响。三种群落类型见
Fig. 1 Effect of the expansion of Phyllostachys edulis on vertical structure in the secondary evergreen broad-leaved forest. The three community types see
图2 毛竹扩张对次生常绿阔叶林群落径级结构的影响。三种群落类型见
Fig. 2 Effect of the expansion of Phyllostachys edulis on diameter at breast height (DBH) class structure in the secondary evergreen broad-leaved forest. The three community types see
2.3.1 对乔木层的影响
由表4可知, 不同群落乔木层的物种多样性指数差异较大。除生态优势度外, 其余5种多样性指数均在毛竹林中最低, 竹阔混交林居中, 常绿阔叶林最高, 且毛竹林的多样性指数均显著小于常绿阔叶林(P < 0.05), 其中Shannon-Wiener指数由常绿阔叶林的2.56降至毛竹林的0.06, 降幅达98%; 而生态优势度指数则为毛竹林>竹阔混交林>常绿阔叶林, 且差异均显著(P < 0.05)。说明随着毛竹扩张, 常绿阔叶林乔木层的物种种类逐渐减少、物种分布均匀度降低, 即毛竹扩张显著降低了常绿阔叶林乔木层的物种多样性。
表4 毛竹扩张对次生常绿阔叶林乔木层物种多样性的影响(平均值±标准误)
Table 4 Effect of the expansion of Phyllostachys edulis on species diversity of tree layer in the secondary evergreen broad-leaved forest (mean ± SE)
| 群落 Community | 物种丰富度 Species richness | Margalef指数 Margalef index | Simpson指数 Simpson index | Shannon-Wiener指数 Shannon-Wiener index | Pielou均匀度 Pielou eveness | 生态优势度 Ecological dominance |
|---|---|---|---|---|---|---|
| PEF | 3.00 ± 1.41b | 0.38 ± 0.26b | 1.03 ± 0.00b | 0.06 ± 0.03c | 0.08 ± 0.07c | 0.97 ± 0.00a |
| PBMF | 11.50 ± 4.95ab | 2.12 ± 0.98ab | 1.62 ± 0.21b | 0.93 ± 0.25b | 0.39 ± 0.03b | 0.63 ± 0.08b |
| EBF | 20.00 ± 4.24a | 4.70 ± 1.08a | 10.72 ± 0.74a | 2.56 ± 0.12a | 0.86 ± 0.02a | 0.11 ± 0.01c |
2.3.2 对灌木层的影响
由表5可知, 物种多样性指数在3种群落间表现不一致。其中物种丰富度在常绿阔叶林中最高, 竹阔混交林次之, 毛竹林最低, 且阔叶林与毛竹林间差异显著(P < 0.05); Simpson指数表现为竹阔混交林>阔叶林>毛竹林, 且差异显著(P < 0.05); Margalef指数、Shannon-Wiener指数、Pielou均匀度和生态优势度指数在3种群落中均差异不显著, 但Shannon-Wiener指数由阔叶林的2.58降至毛竹林的2.03, 降幅21%。这说明毛竹向阔叶林扩张过程中, 灌木层的物种种类呈减少的趋势, 而物种分布均匀度变化不大。换言之, 毛竹扩张一定程度上降低了灌木层的物种多样性。
表5 毛竹扩张对次生常绿阔叶林灌木层物种多样性的影响(平均值±标准误)
Table 5 Effect of the expansion of Phyllostachys edulis on species diversity of shrub layer in the secondary evergreen broad-leaved forest (mean ± SE)
| 群落 Community | 物种丰富度 Species richness | Margalef指数 Margalef index | Simpson指数 Simpson index | Shannon-Wiener指数 Shannon-Wiener index | Pielou均匀度 Pielou eveness | 生态优势度 Ecological dominance |
|---|---|---|---|---|---|---|
| PEF | 15.50 ± 6.36b | 3.66 ± 1.66a | 4.65 ± 1.93c | 2.01 ± 0.47a | 0.74 ± 0.06a | 0.25 ± 0.09a |
| PBMF | 27.50 ± 4.95ab | 6.13 ± 0.66a | 17.13 ± 1.31a | 2.95 ± 0.12a | 0.89 ± 0.01a | 0.07 ± 0.01a |
| EBF | 33.50 ± 3.54a | 6.42 ± 0.16a | 11.07 ± 2.00b | 2.58 ± 0.33a | 0.74 ± 0.12a | 0.10 ± 0.02a |
2.3.3 对草本层的影响
由表6可知, 不同群落间草本层的物种多样性指数变化程度不同。物种丰富度和Margalef丰富度指数发生了显著变化, 表现为毛竹林和竹阔混交林显著高于常绿阔叶林(P < 0.05); 而Simpson、Shannon-Wiener、Pielou均匀度和生态优势度指数在3个群落中差异较小。说明毛竹向常绿阔叶林扩张过程中, 草本层的物种数逐渐增加, 而物种分布均匀度基本不变。即毛竹扩张一定程度上增加了草本层的物种多样性。
表6 毛竹扩张对次生常绿阔叶林草本层物种多样性的影响
Table 6 Effect of the expansion of Phyllostachys edulis on species diversity of herb layer in the secondary evergreen broad-leaved forest
| 群落 Community | 物种丰富度 Species richness | Margalef指数 Margalef index | Simpson指数 Simpson index | Shannon-Wiener指数 Shannon-Wiener index | Pielou均匀度 Pielou eveness | 生态优势度 Ecological dominance |
|---|---|---|---|---|---|---|
| PEF | 31.00a | 6.32a | 15.00a | 2.95a | 0.86a | 0.07a |
| PBMF | 28.00a | 5.40a | 16.48a | 2.95a | 0.89a | 0.07a |
| EBF | 18.00b | 3.93b | 13.77a | 2.64a | 0.91a | 0.08a |
物种组成是构成群落结构和生物多样性的基础, 也是影响森林生态系统过程的根源(Tilman et al, 1997)。Bray-Curtis指数能准确衡量群落间的物种组成差异, 也可以反映沿环境梯度变化的物种替代程度(马克平等, 1995)。群落间共有种越多且个体数量分布越相近, Bray-Curtis指数值越大, 群落间物种组成相似性就越高。本研究中, 毛竹林与常绿阔叶林乔木层、灌木层和草本层的Bray-Curtis相似性指数均很小(表3), 说明毛竹林与常绿阔叶林物种组成差异较大。毛竹扩张使阔叶树种类和数量明显减少, 其中建群种红楠的重要值由扩张前的30.22最终减少为0.00, 这与许多研究结果相符。如杨怀等(2010)研究发现, 毛竹扩张使常绿阔叶林植物种类减少近30余种; 白尚斌等(2013)通过毛竹扩张的动态定位监测发现, 扩张过程中毛竹数量逐渐增加, 其他树种种类和数量明显减少; 祁红艳(2014)研究也发现, 丝栗栲(Castanopsis fargesii)、苦槠(C. sclerophylla)和赤杨叶等阔叶树受毛竹扩张影响很大①(① 祁红艳 (2014) 氮磷根际效应:毛竹扩张的潜在策略. 硕士学位论文, 江西农业大学, 南昌.)竹扩张显著减少了常绿阔叶林中阔叶树的种类及数量。
虽然群落物种组成变化与森林生态系统功能的响应关系较复杂, 但许多研究表明, 群落物种丧失和数量减少会引起森林生产力和抵抗力下降, 增加系统的不稳定性(Lavorel & Garnier, 2002), 尤其是建群种生活型的更替, 会影响其他树种赖以生存的群落环境, 导致植物功能群的转变(Hooper et al, 2005), 从而影响整个生态系统的组成、结构与功能(Ellison et al, 2005)。因此, 毛竹向阔叶林扩张并最终成为群落建群种, 会从根本上影响常绿阔叶林的生态功能属性, 如凋落物分解、土壤养分矿化、碳流通与固定等特征。今后应加强竹子的生物生态学习性研究, 从本质上认识竹子扩张的生态后效。
本研究发现, 毛竹扩张减少了阔叶林群落的分层, 导致群落垂直结构单一, 高度主要集中在12-14 m区间(图1); 同时也减少了大径级个体数量, 导致群落径级结构变窄, 主要分布于5-10 cm (图2), 这与前人的研究相符。杨清培等(2015)认为, 竹子扩张会影响邻近群落结构, 尤其对高大竹种而言, 其影响效应更为明显, 如毛竹向邻近森林成功扩张后, 会在乔木层占绝对优势。本研究中, 扩张后期毛竹的立竹度和其他树种的立木数比例为62:1, 毛竹平均高为12.22 m, 平均胸径为7.95 cm, 导致了扩张后期的群落结构较单一。
此外, 径级分布还可以反映种群年龄结构, 一般小径级代表幼年阶段, 较大径级则表示成年时期(祝燕等, 2008)。毛竹扩张使大径级个体逐渐被淘汰, 小径级个体幸存下来, 影响了种群的年龄结构, 使常绿阔叶林中许多物种的种群趋于幼龄化。虽然幼龄个体相对增多, 种群发展潜力似乎较大, 但植物在幼龄阶段对外界环境变化的适应能力弱, 死亡率往往很高, 而具有繁殖能力的成年个体又少, 易导致种群衰退(张文辉等, 2004)。如华西箭竹(Fargesia nitida)明显改变了岷江冷杉(Abies faxoniana)种群的年龄结构, 同时严重抑制了幼苗更新和生长② (② 李媛 (2007) 卧龙自然保护区亚高山暗针叶林下华西箭竹(Fargesia nitida)对岷江冷杉(Abies faxoniana)幼苗定居的影响. 硕士学位论文, 西南大学, 重庆.)竹扩张简化了常绿阔叶林的群落结构, 阻碍了群落正常更新。
另外, 复杂且合理的群落结构存在充分的空间异质生态位, 为物种多样性的形成与维持提供了基础生境(Pommerening, 2002), 有利于建立生物与生物、生物与环境的适应关系, 可促进生态系统生产的高效流通(Peet, 1992)。因此, 群落结构简化会改变植物竞争关系、植物与微生物的互利共生以及适宜的环境因子, 进而影响森林生态系统的物质循环、能量流动和信息传递等过程, 这也是毛竹扩张引起常绿阔叶林生态功能退化的重要原因。
群落丰富的物种多样性有利于增强植物抵抗入侵的能力与减缓外界环境变化的影响, 是维持森林生态系统动态平衡的关键(Chapin III et al, 2000; Cardinale et al, 2012)。物种多样性是物种丰富度和分布均匀性的综合体现, 物种种类越丰富, 多度分布越平均, 物种多样性就越高(茹文明等, 2006), 其中Shannon-Wiener指数是广泛用于评价物种多样性水平的一个经典指标(马克平和刘玉明, 1994; 王永繁和余世孝, 2002)。本研究发现, 随着毛竹扩张, 乔木层的Shannon-Wiener指数由常绿阔叶林的2.56降至毛竹林的0.06, 降幅高达98%; 灌木层也由2.58降至2.03, 降幅21% (表4和表5), 这一结果与许多研究相符。杨怀等(2010)探讨毛竹扩张时发现, 乔木层的Shannon-Wiener指数由常绿阔叶林中的2.68降至毛竹林的0.49, 降幅82%, 灌木层也从2.59降至2.07, 降幅20%。林倩倩等(2014)研究也指出, 毛竹扩张导致常绿阔叶林乔木层的Shannon-Wiener指数由2.35降为0.00, 降幅达100%。以上结果均说明, 毛竹扩张严重威胁到乔木层的物种多样性, 这主要是因为毛竹通过地上遮阴、机械损伤等物理作用和地下根系、养分竞争等生物化学过程联合淘汰了乔木层其他阔叶树, 逐步占据其空间生态位(杨清培等, 2015)。因此, 毛竹扩张降低了常绿阔叶林的物种多样性, 这很可能又会加剧毛竹扩张形势, 严重影响常绿阔叶林生态功能的持续稳定。
总之, 毛竹向次生常绿阔叶林扩张, 导致其群落组成结构简化、物种多样性下降。而物种组成结构决定生态系统功能, 多样性维持系统稳定, 那么毛竹扩张引起的群落特征改变会对森林生态系统功能及生态服务造成怎样的后续影响, 有待进一步研究。
| [1] |
Plant species diversity and dynamics in forests invaded by moso bamboo (Phyllostachys edulis) in Tianmu Mountain Nature Reserve.
天目山保护区森林群落植物多样性对毛竹入侵的响应及动态变化 .
|
| [2] |
An ordination of the upland forest communities of southern Wisconsin. |
| [3] |
Biodiversity loss and its impact on humanity. |
| [4] |
Consequences of changing biodiversity. |
| [5] |
Interconnection among dominant plant populations of Castanopsis community in Jinggang Mountain Nature Reserve. 井冈山自然保护区栲属群落优势种群的种间联结关系研究 . |
| [6] |
Monitoring Phyllostachys pubescens stands expansion in National Nature Reserve of Mount Tianmu by remote sensing.
天目山国家级自然保护区毛竹林扩张遥感监测 .
|
| [7] |
Loss of foundation species: consequences for the structure and dynamics of forested ecosystems. |
| [8] |
Impacts of moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in Kyoto, western Japan.
|
| [9] |
|
| [10] |
Impact of invasive plants on the species richness, diversity and composition of invaded communities. |
| [11] |
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. |
| [12] |
Species diversity, forest structure and species composition in Tanzanian tropical forests.
|
| [13] |
Range expansion and its mechanisms in a naturalized bamboo species, Phyllostachys pubescens, in Japan. |
| [14] |
|
| [15] |
Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. |
| [16] |
Bamboo overabundance alters forest structure and dynamics in the Atlantic forest hotspot. |
| [17] |
Effects of Phyllostachys pubescens forest expansion on biodiversity in Tianmu Mountain National Nature Reserve. 天目山国家级自然保护区毛竹林扩张对生物多样性的影响 . |
| [18] |
Measurement of biotic community diversity. II. β diversity. 生物群落多样性的测度方法.II. β多样性的测度方法 . |
| [19] |
Measurement of biotic community diversity. I. α diversity (Part 2). 生物群落多样性的测度方法. I. α多样性的测度方法(下) . |
| [20] |
|
| [21] |
Approaches to quantifying forest structures.
|
| [22] |
Species diversity and community structure of forest communities in Lishan Mountain.
历山森林群落物种多样性与群落结构研究 .
|
| [23] |
Comparisons of soil-water content between a moso bamboo (Phyllostachys pubescens) forest and an evergreen broadleaved forest in western Japan.
|
| [24] |
Changes in the hydrological functions of litter layer following Phyllostachys edulis expansion into evergreen broadleaved forest. 毛竹扩张的生态后效: 凋落物水文功能评价 . |
| [25] |
|
| [26] |
The influence of functional diversity and composition on ecosystem processes. |
| [27] |
A new species diversity index and its fractal analysis. |
| [28] |
物种多样性指数及其分形分析 . |
| [28] |
The effects of Phyllostachys pubescens expansion on soil fertility in National Nature Reserve of Mount Tianmu. 天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响 . |
| [29] |
|
| [30] |
Effects of Phyllostachys pubescens expansion on plant species diversity in Jigong Mountain and discussion of control measures. 鸡公山毛竹扩张对植物多样性的影响及控制措施 . |
| [31] |
Ecological studies on bamboo expansion: process, consequence and mechanism. 竹子扩张生态学研究: 过程、后效与机制 . |
| [32] |
Spatial variation of soil nutrient contents in the Jinggangshan National Nature Reserve.
井冈山国家级自然保护区森林土壤养分含量的空间变化 .
|
| [33] |
Age structure and time sequence prediction of populations of an endangered plant, Larix potaninii var. chinensis. 濒危植物太白红杉种群年龄结构及其时间序列预测分析 . |
| [34] |
Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, East China.
古田山中亚热带常绿阔叶林动态监测样地—群落组成与结构 .
|

备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}