生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 )。过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 )。
海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves)。迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 )。最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 )。然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 )。
在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度。从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 )。横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 )。本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说。Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别。纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 )。但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及。
本研究沿着海拔梯度分别在金沙江、澜沧江和怒江的干旱河谷区域设置样地, 对于3个河谷垂直方向均完全覆盖, 且不同河谷、不同经纬度之间形成海拔梯度的对比。研究三江并流地区干旱河谷的群落类型结构和物种多样性的垂直变化格局及其水平对比(McCain, 2010 ), 旨在探讨以下3个方面的问题: (1)该地区植物多样性的海拔格局及不同山地之间的差异; (2)影响三江并流地区植物群落α和β多样性格局的空间和地形因子; (3)探讨平行的纵向岭谷地形对植物群落物种构成所产生的隔离效应。通过探寻该区域植物多样性的垂直分布规律及区域差异, 本研究可为该区域生物多样性保护重点的选择提供依据。
1 研究区域与方法
1.1 研究区概况
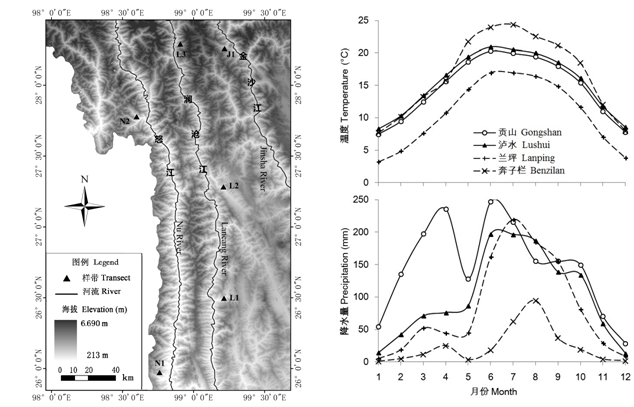
研究区域位于滇西北三江并流地区, 地理坐标范围为26°25′-28°24′ N, 97°55′-100°22′ E, 在北、东方向分别与西藏的林芝地区、四川甘孜藏族自治州和凉山彝族自治州毗邻, 西面与缅甸接壤。行政上这一区域包含怒江傈僳族自治州、迪庆藏族自治州和丽江市的11个县市。发源于青藏高原的怒江、澜沧江、金沙江3条大河南北贯穿本区域, 与自西向东平行排列的高黎贡山、怒山-碧罗雪山、云岭-老君山、雪山4条山脉, 形成“三江并流”的纵向平行岭谷地貌。境内最高点为海拔6,740 m的梅里雪山, 位于怒山-碧罗雪山北端, 亦是云南省最高峰; 最低点位于泸水县怒江河谷, 海拔600 m以下(何永彬等, 2000 ; 明庆忠和史庆涛, 2007 )。
三江并流区的气候受西南季风、东南季风和青藏高原大陆性高寒气候的交互影响, 加之地表相对高差显著, 因而气候具有强烈的季节性变化, 且垂直分化十分明显。高海拔地带冬春季长、夏秋季短, 而河谷低地则相反。该区域日温差和年温差大。普遍干湿季分明, 降雨集中于夏季, 暴雨多, 立体气候显著。据区内贡山(海拔1,583.3 m)、泸水(1,804.7 m)、奔子栏(2,005 m)和兰坪(2,344.9 m)四地气象台站的长期观测数据, 年均温度10.7-16.6℃, 月均温年较差12.5-17.8℃; 年平均降雨量自西向东降低, 且沿海拔梯度自下而上增加, 其中, 怒江流域年降水量>1,000 mm (贡山县1,772.4 mm), 澜沧江、金沙江流域依次减少(兰坪1,008.2 mm, 香格里拉619.9 mm, 金沙江河谷奔子栏仅285.6 mm)(图1 )。
区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱。各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带。区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m。怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 )。由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 )。
1.2 野外调查
根据野外调查条件的不同, 在怒江、澜沧江和金沙江河谷的不同地点和坡向共设置了6条沿海拔梯度的非连续样带(图1 , 表1 )。其中, 金沙江河谷样带(J1)在白马雪山东坡、金沙江西岸奔子栏镇附近; 澜沧江河谷样带分别设置在兰坪营盘镇(L1)、维西白济汛乡(L2)和德钦县云岭乡(L3); 怒江河谷样带设置在泸水县片马(N1)和贡山县(N2)。每条样带依托一条翻山公路, 沿公路从江边开始布置样方。一般情况下海拔每上升100 m, 在公路两侧100 m以外选择人为干扰不明显的生境, 布置2-5个面积100 m2 的样方。有些区域由于地形陡峭或人为干扰较大等因素, 样方间的海拔梯度间隔较大。
图1 研究区域内河流、地貌及植被调查的海拔样带位置图。N1: 片马; N2: 贡山; L1: 兰坪; L2: 维西; L3: 德钦; J1: 奔子栏。
Fig. 1 The geomorphology, rivers and the locations of altitudinal transects of vegetation survey in the study area. N1, Pianma; N2, Gongshan; L1, Lanping; L2, Weixi; L3, Deqin; J1, Benzilan.
以10 m×10 m的样方为调查单元, 清查每个样方中出现的物种。对于乔木, 记录每株的种名, 并测量其胸径和高度; 对于草本和灌木, 记录每个物种的种名、个体数或多度, 并目测其百分比盖度。对于每个调查样方, 用GPS测定地理坐标和海拔, 罗盘测量地形的坡向和坡度。
由于河谷干旱气候与山地湿润气候的差异, 加上低海拔地区长期的人为活动影响, 各条海拔样带上的植物群落样方都可以分为2个植被类型, 即低海拔的灌草丛和中山以上的森林群落。各样带因基带气候和人为干扰强度的差异, 2种植被类型的分界海拔不同。
1.3 数据分析
1.3.1 多样性指标计算
α多样性采用单位面积样方中的物种丰富度值。分别统计各调查样方中全部维管束植物以及乔木、灌木和草本植物(含蕨类)的物种丰富度。每条样带上某一海拔的植物群落物种丰富度及草本、灌木、乔木物种丰富度分别为该海拔全部样方物种丰富度的平均值。
β多样性包含两个样本之间由于物种数量减少(nestedness)和物种更替(turnover)两个可加性成分(Koleffet al, 2003 )。在诸多β多样性指数中, Sim- pson相异性指数(βsim )反映了样本之间物种构成的更替速率, 且不受样方物种丰富度的影响(Baselga, 2010 ; Baselga & Leprieur, 2015 ), 故用来描述同一样带内不同海拔样方群落之间的物种更替。
其中, a 表示两个样方间共有的物种数, b 表示在第一个样方中出现但是在第二个样方中不出现的物种数, c 表示在第二个样方中出现但是在第一个样方中不出现的物种数。
计算同一样带相邻海拔间的物种更替速率时, 由于每个海拔有2-5个样方不等, 故计算两个海拔之间所有成对样方组合之间的βsim 并求平均值。如A、B是同一样带内的相邻海拔, A处有2个样方a1、a2, B处有3个样方b1、b2、b3 , 则海拔A、B之间共6种a、b样方组合, 求此6种样方对之间的βsim 平均值, 得到A、B海拔之间的β多样性。
根据野外对调查样带的植被类型划分, 用βsim 计算6条样带和2种植被类型(即低海拔的灌草丛和中山森林), 总共12个样带段之间的β多样性。由于不同样带段内的样方数量不同, 为了减少基数不同导致的误差, 此处使用重复随机抽样, 每次从每条样带段中各抽出5个样方。而后以各条样带段的5个样方中出现的物种计算12个样带段之间的βsim 。重复抽样1,000次并计算结果的平均值, 作为样带段间的β多样性指数。
1.3.2 物种多样性与构成变化的环境解释和方差分析
运用通用线性回归模型, 分析草本、灌木、乔木和全部物种丰富度的海拔梯度格局受相关环境因子的影响及其区域差异。解释变量包括: (1)定量变量: 经纬度、海拔、坡向、坡度; (2)类型变量: 流域(金沙江、澜沧江、怒江)、植被类型(灌草丛、森林)。其中坡向δ作三角函数变换为cos (δ ), 代表光照影响, 值域为[-1, 1], 正北为-1, 正南为1; 作三角函数变换为sin(δ ), 代表东西坡, 值域为[-1, 1], 正西为-1, 正东为1。
分析β多样性时, 针对3个类型变量:
(1)植被类型差别: 0 = 相同, 1 = 不同; (2)空间隔离: 1 = 同一样带, 2 = 同流域不同样带, 3 = 相邻流域样带, 4 = 间隔流域样带; (3)样带植被类型: 1 = 灌草丛带, 2 = 森林带。
运用方差分析, 解析前述12个样带段之间的β多样性来源。
文中所有的计算都在R软件包中进行, βsim 指数的计算使用软件中的vegan包完成, 重复抽样的实现则是通过在R语言中编写循环程序完成。
2 结果
2.1 植物物种丰富度的基本信息
L1 (兰坪)的灌草丛带是以清香木(Pistacia wei- nmannifolia )、马棘(Indigofera pseudotinctoria )、胡枝子(Lespedeza spp.) + 荩草(Arthraxon hispidus )、白茅(Imperata cylindrica )、野艾蒿(Artemisia lavandulaefolia )为优势种的干旱河谷植被类型, 森林带是以云南松(Pinus yunnanensis )、华山松(Pinus armandii )、滇杨(Populus yunnanensis ) + 野艾蒿、白车轴草(Trifolium repens )、画眉草(Eragrostis pilosa )为优势种的温带针阔叶混交林。
L2 (维西)的灌草丛带是以截叶铁扫帚(Les- pedeza cuneata )、马棘、地菍(Melastoma dodecandrum ) + 荩草、鬼针草(Bidens spp.)、野艾蒿为优势种的干旱河谷植被类型, 森林带是以云南松、槲树(Quercus dentata ) + 野艾蒿、白茅为优势种的温带针阔叶混交林。
L3 (德钦)的灌草丛带是以川西白刺花(Sophora davidii var. chuansiensis )、小叶灰毛莸(Caryopteris forrestii var. minor )、华西小石积(Osteomeles schwerinae ) + 荩草、画眉草、阿尔泰狗娃花(Heteropappus altaicus )为优势种的干旱河谷植被类型, 森林带是以高山松(Pinus densata )、华山松、黄背栎(Quercus pannosa ) + 画眉草、槲蕨(Drynaria fortunei )、火绒草(Leontopodium spp.)为优势种的温带针阔叶混交林。
N1 (片马)的灌草丛带是以地菍、地桃花(Urena lobata )、山柳(Clethra spp.) + 荩草、白茅、鬼针草为优势种的干旱河谷植被类型, 森林带是以尼泊尔桤木(Alnus nepalensis )、高山桦(Betula delavayi )、云南铁杉(Tsuga dumosa ) + 常山(Dichroa febrifuga )、箭竹(Fargesia spp.)、鼠麯草(Gnaphalium spp.)为优势种的温带针阔叶混交林。
N2 (贡山)的灌草丛带是以地菍、地桃花、饿蚂蝗(Desmodium multiflorum ) + 荩草、五月艾(Artemisia indica )、鬼针草为优势种的干旱河谷植被类型, 森林带是以云南松、桤木、润楠(Machilus spp.) + 常山、滇北悬钩子(Rubus bonatianus )、竹叶茅(Microstegium nudum )为优势种的温带针阔叶混交林。
J1 (奔子栏)的灌草丛带是以川滇野丁香(Lept- odermis pilosa )、川西白刺花、马棘+荩草、草沙蚕(Tripogon spp.)、垫状卷柏(Selaginella pulvinata )为优势种的干旱河谷植被类型, 森林带是以高山松、华山松、山杨(Populus spp.) + 槲蕨、菝葜(Smilax spp.)、苔草(Carex spp.)为优势种的温带针阔叶混交林。
按流域来看, 怒江两条样带100 m2 样方面积上的平均植物物种丰富度分别为N1 (片马) 36.0 ± 13.8和N2 (贡山) 46.4 ± 15.4种, 高于澜沧江L1 (兰坪) 34.7 ± 9.1种、L2 (维西) 32.7 ± 14.2种和L3 (德钦) 18.1 ± 6.2种, 其中怒江贡山样带的平均物种丰富度最高。白马雪山东坡的J1样带平均物种丰富度18.7 ± 8.1种, 与其西坡相对的L3样带的物种丰富度值彼此接近且最低。同一流域内不同样带相比, 怒江北段的样方物种平均丰富度高于南段, 而在澜沧江是南段的该值高于北段。
按植物生长型比较, 草本植物物种丰富度普遍较高, 怒江两条样带样方平均的草本物种丰富度大约为灌木种的2.5倍(附录1); 在澜沧江中、南部的2条样带, 样方草本种与灌木种的平均丰富度约2:1; 但在澜沧江北部样带和金沙江样带中, 样方草本物种与灌木物种的丰富度接近1:1。草本种与灌木种丰富度的比值从西向东沿着怒江、澜沧江、金沙江逐渐下降, 主要是草本种丰富度显著降低。乔木种丰富度则在各个流域较为相近, 在6条样带均处于0.5-2.8种/100 m2 范围。
按植被带比较, 怒江流域森林带与灌草丛带草本物种数目的差异最大, 澜沧江和金沙江流域森林带与灌草丛带草本物种数目较为相近; 所有流域的森林带灌木物种数目基本都要大于灌草丛带, 但数目差异与草本数目相比差异不大; 森林带和灌草丛带的乔木物种数目差异显著, 灌草丛带乔木物种数很少乃至为零。
2.2 植物物种丰富度的海拔梯度格局
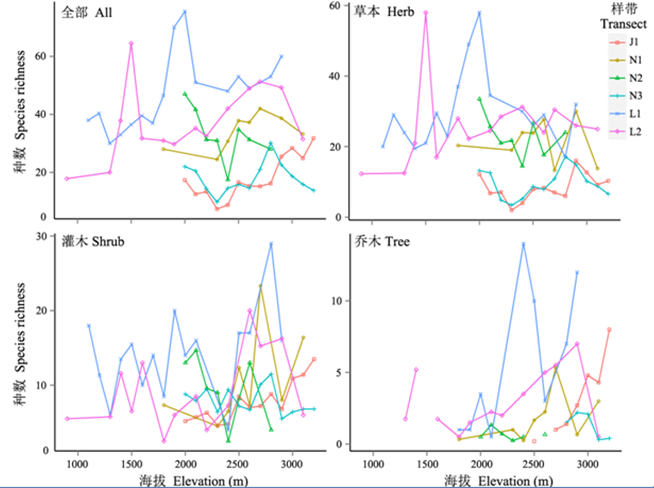
6条样带植物物种丰富度在海拔梯度上具有不太一致的格局(图2 ), 其中怒江两条样带都分别在2,000 m及以下和在2,800 m附近出现峰值; 澜沧江南部和北部样带的丰富度峰值出现在海拔2,700- 2,800 m, 而中部样带的峰值在海拔2,000 m以下; 金沙江样带的物种丰富度从2,300 m以上随海拔上升而增加, 在3,300 m处达到峰值。
图2 三个流域6条植被样带中全部植物和不同生长型物种丰富度的海拔梯度分布。N1: 怒江片马; N2: 怒江贡山; L1: 澜沧江兰坪; L2: 澜沧江维西; L3: 澜沧江德钦; J1: 金沙江奔子栏。
Fig. 2 Elevational gradients of species richness of all plants and different growth-forms for the six vegetation transects in the three watersheds. N1, Nu River Pianma; N2, Nu River Gongshan; L1, Lancang River Lanping; L2, Lancang River Weixi; L3, Lancang River Deqin; J1, Jinsha River Benzilan.
从不同的植物生长型来看, 对于草本植物, 金沙江样带和澜沧江北部样带丰富度最低, 澜沧江中、南部两条样带丰富度与怒江样带接近, 但怒江2条样带在海拔2,000 m以下分别出现1个显著的高峰值。除此之外, 其余样带的海拔梯度格局不明显。对于灌木种, 海拔梯度格局同样不明显, 但各条样带都在海拔2,300-2,400 m出现较低值, 怒江2条样带灌木种丰富度高峰值出现在海拔2,600-2,800 m, 澜沧江样带的灌木种丰富度海拔梯度格局均不明显, 而金沙江样带灌木种丰富度随海拔上升而增加。乔木种丰富度除金沙江样带呈现随海拔上升而增加的趋势外, 其余样带峰值都在海拔2,300-2,900 m出现。
2.3 植物物种丰富度格局的地理环境解释
区内植物物种分布格局受到地理环境因子的影响, 如表2 所示。总体来讲, 各层植物物种均沿着纬度和海拔梯度升高而丰富度显著增加, 其中草本层和灌木层增加更为显著, 但乔木种基本不随着地理梯度的变化而变化; 相比草本层和乔木层, 灌木物种丰富度沿着经度自西向东显著增加。从流域方面分析, 澜沧江样带样方的物种丰富度整体上与金沙江样带没有显著区别, 尽管草本多样性偏高, 灌木多样性偏低, 但只有乔木种丰富度显著低于金沙江样带。尽管怒江样带的样方中全部种、灌木和草本种丰富度显著高于金沙江和澜沧江样带, 但乔木种丰富度的差异并不显著。从植被类型方面来分析, 森林带内样方中的草本物种丰富度显著低于灌草丛带样方, 而且拥有后者不具备的乔木种, 但灌木种丰富度的多样性差别并不显著, 而森林带与灌草丛带的物种丰富度在整体上也没有显著差异。
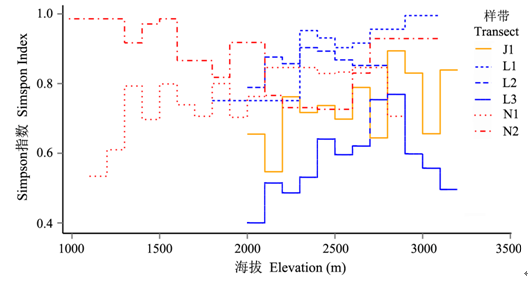
图3 不同样带植被沿海拔梯度的物种更替率(以Simpson指数表示)
Fig. 3 The species turnover rate (Simpson index) of plant community along the elevational transects of vegetation
在样方尺度上, 地形特征影响物种丰富度的分布。坡向与样方草本层物种和物种总数显著负相关, 即北坡草本物种数高于南坡。坡度对灌木和草本物种数都有接近显著的影响, 坡度越大, 草本物种多样性越低, 而灌木种多样性越高, 但对乔木种和总种数影响不显著。以上模型变量总共解释了草本生物多样性67.13%的变异, 解释了灌木生物多样性的34.10%, 解释了乔木生物多样性的40.65%, 解释了总体植物生物多样性的71.26%。
2.4 植物β多样性的空间格局
总体来看, 三个流域的6条样带中物种更替的平均水平存在明显的差异。对于怒江西坡, 北部贡山样带的βsim (0.870±0.095)大于南部的片马样带; 而对于澜沧江东坡的3条样带, 海拔梯度上βsim 平均值从南向北有降低趋势, 分别为0.915 ± 0.033 (兰坪), 0.863 ± 0.037 (维西)和0.581 ± 0.108 (德钦); 金沙江样带处于白马雪山东坡, 与澜沧江的德钦样带(L3)处在同一纬度, 但βsim (0.731 ± 0.098)高于后者。
6条样带的物种更替速率也呈现了不一致的海拔梯度格局(图3 )。位于白马雪山东、西坡的金沙江样带J1和澜沧江样带L3, 其βsim 大致呈现了单峰格局, 且两者的最高值都出现在海拔2,800 m; 澜沧江样带L2的βsim 峰值下降到海拔2,300-2,400 m处, 而澜沧江样带L1的βsim 却在此海拔位置以上继续增大。怒江南北两条样带的物种更替速率呈现相反的海拔梯度格局。在南部片马的样带N1, βsim 有大致向上的单峰格局, 高值区在海拔2,100-2,800 m之间; 而北部贡山的样带N2, βsim 却表现了一个明显的倒单峰格局, 在低海拔段数值最高, 而在海拔2,400-2,600 m段最低。
2.5 样带间植物β多样性的格局
12个样带段的随机抽样(重复1,000次)统计表明(附录2), 群落间物种构成的变异性很大, βsim 值为0.845 ± 0.118。总体上, 样带森林段之间的物种更替率与低海拔灌草丛段相近, βsim 值分别为0.844 ± 0.105和0.850 ± 0.121, 并且单一因素的差异在统计上不显著。
所有样带的森林段或灌草丛段, 都是与同一样带的另一部分(灌草丛段或森林段)之间的物种更替率最小, βsim 均值为0.687, 小于同一流域不同样带相同植被段之间物种更替率的均值(灌草丛段之间 = 0.797; 森林段之间 = 0.776), 更小于所有样带相同植被段之间的更替率均值(灌草丛段之间 = 0.850; 森林段之间 = 0.844)。
金沙江样带两段与相邻的澜沧江6个样带段之间的βsim (0.905 ± 0.055)小于其与怒江4个样带段βsim 的均值(0.958 ± 0.015); 而所有澜沧江样带段与怒江样带段之间βsim 值0.846 ± 0.086, 小于其与金沙江样带段之间的指数均值0.905 ± 0.055。
方差分析结果表明, 以样带植被带段为统计单位的物种更替的所有变化中, 空间隔离因素可以解释34.2%, 而植被类型差异仅能解释不到0.5%。
3 讨论
3.1 植物α多样性海拔梯度的比较和影响因素
作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 )。此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局。Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势。但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致。其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物。
这两种α多样性海拔梯度模式的存在与区域气候的整体格局应有密切联系。由于印度洋季风对区域气候的主导性影响, 区域内降水整体上呈现西高东低、南高北低的趋势(吴绍洪等, 2009 ), 高黎贡山北段西坡和白马雪山北段东坡分别成为区域的“湿极”和“旱极”, 因此在海拔梯度的下部, 流域内从西向东、从南向北所受的干旱胁迫渐趋严重, 而山地中上部的森林带内湿度差逐渐减小。这种环境变化可以解释诸样带之间灌木种丰富度自西向东显著增加, 而草本种和乔木种丰富度(不显著)降低的格局(表2 ); 而灌木和草本种丰富度随海拔上升而显著增加的趋势也应与水分条件同方向改善有直接的联系。同时, 水分条件的限制也强化了局部地形特征对物种丰富度的影响, 如草本和乔木种的丰富度从北坡向南坡降低, 灌木种则相反; 草本和乔木种丰富度随坡度增大而减少, 灌木种则增加。这3种生长型的种丰富度在局部和区域尺度上的差异性变化具有一致性的格局, 显示水分条件在不同尺度上对区内植物物种多样性分布具有突出的限制作用, 以及灌木生长型对干旱气候的适应。灌草丛带的植被极为特殊, 与非洲和印度等地大量分布的稀树干草原(Savanna)植被的外貌、结构和区系等相似(欧晓昆和金振洲, 1996 )。灌木生长型主要存在于灌草丛带, 其优势种以马棘、清香木、地菍、地桃花、川西白刺花等适应干旱气候的灌木为主, 自西向东不同流域水分减少, 使得适应湿润气候的草本和乔木物种减少, 适应干旱气候的灌木物种优势突出, 稀树干草原植被更为典型, 从而造成了灌木生长型与木本和草本的分布差异。
3.2 物种更替的区域格局与主导因素
不同植被类型之间的交错带通常导致海拔梯度上β多样性的峰值(沈泽昊等, 2004 ; Rossatto, 2009 )。在研究区内三个流域的6条植被样带上, β多样性同样显示了多种海拔梯度格局, 包括单峰、上升梯度、平台甚至中部低谷等(图3 )。不同位置的样带所显示出的β多样性海拔梯度变化均有不同, 这主要与样带每个海拔梯度上所取样方较少从而导致物种随机变化较大有关。但总体来看, 这些起伏之中存在的峰值都与样带上不同植被类型之间的过渡带海拔密切相关。白马雪山东、西坡样带(J1和L3)处于同一纬度, 灌丛带过渡到森林带的海拔都在2,700-2,900 m。澜沧江中、南段的两条样带L1和L2上这一植被交错带在2,300-2,400 m之间也对应于β多样性峰值。怒江北部N2样带的β多样性海拔梯度在1,900-2,000 m出现峰值, 南部N1样带的β多样性海拔梯度在2,100-2,200 m出现峰值, 均对应于两条样带的植被交错带。β多样性主要受到空间距离和环境距离的影响, 本研究中样带内样方之间的空间距离基本相近, 差别在于环境距离不同。环境距离是由环境因子差异所产生, 各海拔梯度之间海拔距离相近, 依赖于海拔变化所产生的温度和降水变化也相近, 因此植被类型的变化便成为了影响环境变化的主要因素。在剧烈的环境变化条件下, 植物物种的β多样性也会显示出峰值。也正是这些原因形成了β多样性随海拔呈波浪式变化, 且在植被类型过渡带处呈现峰值的特点。
群落物种构成的更替主要受环境差异和距离产生的扩散限制影响(Graham & Fine, 2008 ; Novillo & Ojeda, 2014 ); 且在区域尺度上, 扩散限制的成因往往不仅仅是空间几何距离(Qian et al, 2009 ; Svenning et al, 2010 )。分隔在3条河流中的6条样带12个植被带段之间的物种更替显示了3方面因素的影响, 但最为突出的显然是地形隔离对物种分布的影响(附录2)。一方面, 因为不同河流样带段之间的几何距离常常小于同一河流不同样带段之间的距离, 如N1-N2的距离就大于N1-L1、N1-L2、N2-L1、N2- L2、N2-L3以及N2-J1, 而L3-J1的距离小于L2-L3和L1-L3, 但不同河流之间样带段的物种更替率明显较大; 而同一样带上、下两段之间主要由于气候差异产生的物种更替率效应远低于区域隔离的效应。另一方面, 位于海拔梯度下部的干旱河谷灌草丛植被之间比中部森林带之间的隔离程度应该更强。可见在研究区域尺度上, 岭谷地形隔离对植物群落的物种更替具有最重要的影响。当然, 由于经、纬度方向上明显的气温和降水梯度的存在, 区域环境(特别是气候)梯度可能是未能解释的物种更替格局变异性的另一重要原因。
怒江、澜沧江和金沙江三个流域6条样带植物种α和β多样性海拔梯度的变化格局, 反映了它们与区域内显著的地形起伏隔离、气候垂直梯度与地理分异之间的联系, 这也许正是区内生物多样性丰富的重要成因。大量的生物多样性区域研究也都证实了气候和环境隔离因素的贡献(Peixoto et al, 2014 ; Yang et al, 2016 )。本研究没有直接分析物种丰富度和物种更替的空间格局与气候因子的定量关系, 主要是因为在这一地形起伏剧烈的区域, 缺少高空间分辨率的气候数据来准确描述气温和降水的地理变异, 而基于少数气象台站的观测数据难以胜任相关的分析。随着更多观测数据的积累, 未来有望更细致地分析和理解本区生物多样性及其分异格局的环境机制。
(责任编委: 王国宏 责任编辑: 时意专)
附录 Supplementary Material
附录1 怒江、澜沧江和金沙江6条样带的植物物种丰富度的均值(100 m2 样方内)和变化范围(括号内), 包括全部种、乔木、灌木和草本(含蕨类)种类。
Appendix 1 The mean values and value ranges (in parenses) of species richness within the plot area 100 m2 of the six altitudinal transects in Nu River, Lancang River and Jinsha River, including total species richness and those of trees, shrubs, herbs (ferns).
http://www.biodiversity-science.net/fileup/PDF/2015361-1.pdf
附录2 不同样带段之间的物种更替率(Simpson指数)。样地代号见表1 。s: 灌草丛段; f: 森林段。
Appendix 2 Species turnover rate (Simpson index) between sections of the sampled transects. The plot codes refer to Table 1 . S, Shrubs and herbs section of the transect; f, Forest section of the transect.
http://www.biodiversity-science.net/fileup/PDF/2015361-2.pdf
参考文献
文献选项
[1]
Alicia MY Moreno LA Piñero D Jorgensen TH Emerson BC 2015 ) Biodiversity in the mexican highlands and the interaction of geology, geography and climate within the trans-mexican volcanic belt
. Journal of Biogeography , 42 , 1586 -1600 .
[2]
Barthlott W Hostert A Kier G Kueper W Kreft H Mutke J 2007 ) Geographic patterns of vascular plant diversity at continental to global scales
. Erdkunde , 61 , 305 -315 .
[本文引用: 1]
[3]
Baselga A 2010 ) Partitioning the turnover and nestedness components of beta diversity
. Archives Danatomie Dhistologie Et Dembryologie Normales Et Expérimentales , 19 , 134 -143 .
[本文引用: 1]
[4]
Baselga A Leprieur F 2015 ) Comparing methods to separate components of beta diversity
. Methods in Ecology and Evolution , 6 , 1069 -1079 .
[本文引用: 1]
[5]
Beck J Chey VK 2008 ) Explaining the elevational diversity pattern of geometrid moths from borneo: a test of five hypotheses
. Journal of Biogeography , 35 , 1452 -1464 .
[本文引用: 1]
[6]
Beck J Kitching IJ 2009 ) Drivers of moth species richness on tropical altitudinal gradients: a cross-regional comparison
. Global Ecology & Biogeography , 18 , 361 -371 .
[本文引用: 1]
[7]
Bram V Falko B Luc B 2013 ) Disturbance regime alters the impact of dispersal on alpha and beta diversity in a natural metacommunity
. Ecology , 11 , 2547 -2557 .
[8]
Colwell RK Rahbek C Gotelli N 2004 ) The mid-domain effect and species richness patterns: what have we learned so far?
The American Naturalist , 163 , E1 -E23 .
[本文引用: 1]
[9]
Donzelli D DeMichele C Scholes RJ 2013 ) Competition between trees and grasses for both soil water and mineral nitrogen in dry savannas
. Journal of Theoretical Biology , 332 , 181 -190 .
[10]
Fang JY 2004 ) Exploring altitudinal patterns of plant diversity of China’s mountains
. Biodiversity Science , 12 , 1 -4 .(in Chinese)
[本文引用: 1]
[方精云 (2004 ) 探索中国山地植物多样性的分布规律
. 生物多样性 , 12 , 1 -4 .]
[本文引用: 1]
[11]
Feng JM Wang XP Xu CD Yang YH Fang JY 2006 ) Altitudinal patterns of plant species diversity and community structure on Yulong Mountains, Yunnan, China
. Journal of Mountain Science , 24 , 110 -116 .(in Chinese with English abstract)
[本文引用: 2]
[冯建孟 , 王襄平 , 徐成东 , 杨元合 , 方精云 (2006 ) 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
. 山地学报, 24 , 110 -116 .]
[本文引用: 2]
[12]
Graham CH Fine PVA 2008 ) Phylogenetic beta diversity: linking ecological and evolutionary processes across space in time
. Ecology Letters , 11 , 1265 -1277 .
[本文引用: 1]
[13]
Grytnes JA 2003 ) Species-richness patterns of vascular plants along seven altitudinal transects in Norway
. Ecography , 26 , 291 -300 .
[本文引用: 1]
[14]
Grytnes JA Heegaard E Romdal TS 2008 ) Can the mass effect explain the mid-altitudinal peak in vascular plant species richness?
Basic and Applied Ecology , 9 , 373 -382 .
[本文引用: 1]
[15]
He JS Chen WL 1997 ) A review of gradient changes in spec ies diversity of land plant communities
. Acta ecologica sinica , 17 , 91 -99 .(in Chinese with English abstract)
[本文引用: 1]
[贺金生 , 陈伟烈 (1997 ) 陆地植物群落物种多样性的梯度变化特征. 生态学报,
17 , 91 -99 .]
[本文引用: 1]
[16]
He YB Lu PZ Zhu T 2000 ) Causes for the formation of dry-hot valleys in Hengduan Mountain, Yunnan Plateau
. Resources Science , 22 (5 ), 69 -72 .(in Chinese with English abstract)
[本文引用: 1]
[何永彬 , 卢培泽 , 朱彤 (2000 ) 横断山——云南高原干热河谷形成原因研究. 资源科学,
22(5) , 69 -72 .]
[本文引用: 1]
[17]
Hubbell SP 1979 ) Tree dispersion, abundance, and diversity in a tropical dry forest
. Science , 203 , 1299 -1309 .
[18]
Jiang LG Liu XN Feng ZM 2014 ) Spatial-temporal pattern analysis of land use and land cover change in Xishuangbanna
. Resources Science , 36 (2 ), 259 -266 .(in Chinese with English abstract)
[姜鲁光 , 刘晓娜 , 封志明 (2014 ) 西双版纳土地利用/土地覆被变化时空格局分析
. 资源科学, 36(2) , 259 -266 .]
[19]
Jiang Y Kang MY Zhu Y Xu GC 2007 ) Plant biodiversity patterns on Helan Mountain, China
. Acta Oecologica , 32 , 125 -133 .
[20]
Kessler M 2000 ) Altitudinal zonation of andean cryptogam communities
. Journal of Biogeography , 27 , 275 -282 .
[本文引用: 1]
[21]
Kraaij T Ward D 2006 ) Effects of rain, nitrogen, fire and grazing on tree recruitment and early survival in bush-encroached savanna, South Africa
. Plant Ecology , 186 , 235 -246 .
[22]
Li K Liu FY Yang ZY Sun YY 2011 ) Study status and trends of vegetation restoration of dry-hot valley in Southwest China
. World Forestry Research , 24 (4 ), 55 -60 .(in Chinese with English abstract)
[李昆 , 刘方炎 , 杨振寅 , 孙永玉 (2011 ) 中国西南干热河谷植被恢复研究现状与发展趋势
. 世界林业研究, 24(4) , 55 -60 .]
[23]
Li XW 1995 ) A floristic study on the seed plants from the region of Yunnan Plateau
. Acta Botanica Yunnanica , 17 (1 ), 1 -14 .
[本文引用: 1]
(in Chinese with English abstract) [李锡文 (1995 ) 云南高原地区种子植物区系. 云南植物研究,
17(1) , 1 -14 .]
[本文引用: 1]
[24]
Liu FY Li K Sun YY Tang GY Zhang CH 2010 ) Effects of climate on vegetation recovery in dry-hot valleys of Heng- duan Mountainous region in Southwest China
. Resources and Environment in the Yangtze Basin , 19 , 1386 -1391 .(in Chinese with English abstract)
[本文引用: 1]
[刘方炎 , 李昆 , 孙永玉 , 唐国勇 , 张春华 (2010 ) 横断山区干热河谷气候及其对植被恢复的影响
.长江流域资源与环境, 19 , 1386 -1391 .]
[本文引用: 1]
[25]
MacArthur RH 1972 ) Mathematical ecology and its place among the sciences
. Science , 178 , 389 -394 .
[本文引用: 1]
[26]
Ma K Li YN Jin H Long ZH Lu YP 2011 ) A comparison of the species diversity of Trollius chinensis community in different habitats
. Pratacultural Science , 28 , 1467 -1472 .(in Chinese with English abstract)
[马凯 , 李永宁 , 金辉 , 龙在海 , 卢银平 (2011 ) 不同生境类型金莲花群落物种多样性比较
. 草业科学, 28 , 1467 -1472 .]
[27]
McCain CM Sanders NJ 2010 ) Metabolic theory and elevational diversity of vertebrate ectotherms
. Ecology , 91 , 601 -609 .
[本文引用: 1]
[28]
Ming QZ Shi ZT 2007 ) New discussion on dry valley formation in the Three Parallel Rivers Region
. Journal of Desert Research , 27 (1 ), 99 -104 .(in Chinese with English abstract)
[本文引用: 1]
[明庆忠 , 史正涛 (2007 ) 三江并流区干热河谷成因新探析
. 中国沙漠, 27 , 99 -104 .]
[本文引用: 1]
[29]
Myers N Mittermeier RA Mittermeier CG Da FG Kent J 2000 ) Biodiversity hotspots for conservation priorities
. Nature , 403 , 853 -858 .
[本文引用: 1]
[30]
Nong Y Zheng L Jia HY Lu LH Huang DW Huang BH Lei LQ 2015 ) Community characteristics and spatial distribution of dominant tree species in a secondary forest of Daqing Mountains, southwestern Guangxi, China
. Biodiversity Science , 23 , 321 -331 .(in Chinese with English abstract)
[农友 , 郑璐 , 贾宏炎 , 卢立华 , 黄德卫 , 黄柏华 , 雷丽群 (2015 ) 广西大青山次生林的群落特征及主要乔木种群的空间分布格局
. 生物多样性, 23 , 321 -331 .]
[31]
Novillo A Ojeda RA 2014 ) Elevation patterns in rodent diversity in the dry Andes: disentangling the role of environmental factors
. Journal of Mammalogy , 95 , 99 -107 .
[本文引用: 1]
[32]
O’Brien EM Field R Whittaker RJ 2000 ) Climatic gradients in woody plant (tree and shrub) diversity: water-energy dynamics, residual variation, and topography
. Oikos , 89 , 588 -600 .
[33]
Ou XK Jin ZZ 1996 ) A preliminary study on the flora and ecological diversity in Jinsha River dry-hot Valley
. Journal of Wuhan Botanical Research , 14 , 318 -322 .(in Chinese with English abstract)
[本文引用: 2]
[欧晓昆 , 金振洲 (1996 ) 金沙江干热河谷植物区系和生态多样性的初步研究
. 武汉植物研究, 14 , 318 -322 .]
[本文引用: 2]
[34]
Peixoto FP Braga PHP Cianciaruso MV Diniz-Filho JAF Brito D 2014 ) Global patterns of phylogenetic beta diversity components in bats
. Journal of Biogeography , 41 , 762 -772 .
[本文引用: 1]
[35]
Qian H Badgley C Fox DL 2009 ) The latitudinal gradient of beta diversity in relation to climate and topography for mammals in North America
. Global Ecology and Biogeography , 18 , 111 -122 .
[本文引用: 1]
[36]
Rahbek , C. (2005 ). The role of spatial scale and the perception of large-scale species-richness patterns
. Ecology Letters , 8 , 224 -239 .
[37]
Rossatto DR Hoffmann WA Franco AC 2009 ) Differences in growth patterns between co-occurring forest and savanna trees affect the forest-savanna boundary
. Functional Ecology , 23 , 689 -698 .
[本文引用: 1]
[38]
Ruifrok JL Postma F Olff H Smit C 2014 ) Scale-dependent effects of grazing and topographic heterogeneity on plant species richness in a Dutch salt marsh ecosystem
. Applied Vegetation Science , 17 , 615 -624 .
[39]
Salazar A Goldstein G Franco AC 2012 ) Differential seedling establishment of woody plants along a tree density gradient in Neotropical savannas
. Journal of Ecology , 100 , 1411 -1421 .
[40]
Sankaran M Ratnam J Hanan N 2008 ) Woody cover in African savannas: the role of resources, fire and herbivory
. Global Ecology and Biogeography , 17 , 236 -245 .
[41]
Shen R Zhang JL He B Li F Zhang ZM Zhou R Ou XK 2010 ) The structure characteristic and analysis on similarity of grassland community in dry-hot valley of Yuanjiang River
. Ecology and Environmental Sciences , 19 , 2821 -2825 .(in Chinese with English abstract)
[沈蕊 , 张建利 , 何彪 , 李峰 , 张志明 , 周睿 , 欧晓昆 (2010 ) 元江流域干热河谷草地植物群落结构特征与相似性分析. 生态环境学报,
19 , 2821 -2825 .]
[42]
Shen ZH Hu HF Zhou Y Fang JY 2004 ) Altitudinal patterns of plant species diversity on the southern slope of Mt. Shennongjia, Hubei, China
. Biodiversity Science , 12 , 99 -107 .(in Chinese with English abstract)
[本文引用: 1]
[沈泽昊 , 胡会峰 , 周宇 , 方精云 (2004 ) 神农架南坡植物群落多样性的海拔梯度格局
. 生物多样性, 12 , 99 -107 .]
[本文引用: 1]
[43]
Shen ZH Zhang XS 2000 ) A quantitative analysis on the floristic elements of the Chinese subtropical region and their spatial patterns
. Acta phytotaxonomica Sinica , 38 , 366 -380 .
[本文引用: 1]
[沈泽昊 , 张新时 (2000 )中国亚热带地区植物区系地理成分及其空间格局的数量分析
. 植物分类学报 , 38 , 366 -380 .]
[本文引用: 1]
[44]
Slik JWF Raes N Aiba SI Brearley FQ Cannon CH 2009 ) Environmental correlates for tropical tree diversity and distribution patterns in Borneo
. Diversity and Distributions , 19 , 523 -532 .
[45]
Stevens G 1996 ) Extending rapoport's rule to pacific marine fishes
. Journal of Biogeography , 23 , 149 -154 .
[本文引用: 1]
[46]
Sun ZH Peng SJ Ou XK 2007 ) A quick assessment and environmental interpretation for the tree species richness in the Gaoligong Mountain Range
. Chinese Science Bulletin , 52 (S2 ), 195 -200 .
[本文引用: 1]
[孙振华 , 彭声静 , 欧晓昆 (2007 ) 高黎贡山乔木树种丰富度快速评估及其环境解释
. 科学通报 , 52 (S2 ), 195 -200 .]
[本文引用: 1]
[47]
Svenning JC Fitzpatrick MC Normand S Graham CH Pear- man PB Iverson LR Skov F 2010 ) Geography, topogra- phy, and history affect realized-to-potential tree species richness patterns in Europe
. Ecography , 33 , 1070 -1080 .
[本文引用: 1]
[48]
Tao J Zang RG Yu CY 2011 ) Altitudinal patterns of plant communities and species diversity in the Habaxueshan Mountains, Yunnan, China
. Scientia Silvae Sinicae , 47 (7 ), 1 -6 .(in Chinese with English abstract)
[陶晶 , 臧润国 , 余昌元 (2011 ) 云南哈巴雪山植物群落和植物多样性海拔梯度分布格局
. 林业科学, 47(7) , 1 -6 .]
[49]
Tang ZY Fang JY 2004 ) A review on the elevational patterns of plant species diversity
. Biodiversity Sciences , 2004 , 12 , 20 -28 .(in Chinese with English abstract)
[本文引用: 1]
[唐志尧 , 方精云 (2004 ) 植物物种多样性的垂直分布格局
. 生物多样性, 12 , 20 -28 .]
[本文引用: 1]
[50]
Valdés A Lenoir J Emilie GM Andrieu E Brunet J Chabrerie O Déborah CK Cousins SA Deconchat M Frenne PD Smedt PD Diekmann M Hansen K Hermy M Kolb A Liira J Lindgren J Naaf T Paal T Prokofieva I Michael SL Wulf M Verheyen K Decocq G 2015 ) The contribution of patch-scale conditions is greater than that of macroclimate in explaining local plant diversity in fragmented forests across Europe
. Global Ecology and Biogeography , 24 , 1094 -1105 .
[51]
Vega RV Salazar A Wilhelm FM 2014 ) Do groundwater dynamics drive spatial patterns of tree density and diversity in Neotropical savannas
. Journal of Vegetation Science , 25 , 1465 -1473 .
[52]
Wang KQ Shen YX Chen QB Wang ZH 2004 ) Soil water environment of artificial vegetation in Jinshajiang dry-hot valley
. Chinese Journal of Applied Ecology , 15 , 809 -813 .(in Chinese with English abstract)
[本文引用: 2]
[王克勤 , 沈有信 , 陈奇伯 , 王震洪 (2004 ) 金沙江干热河谷人工植被土壤水环境
. 应用生态学报, 15 , 809 -813 .]
[本文引用: 2]
[53]
Wang ZH Chen AP Piao SL Fang JY 2004 ) Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China
. Biodiversity Science , 12 , 82 -88 .(in Chinese with English abstract)
[本文引用: 1]
[王志恒 , 陈安平 , 朴世龙 , 方精云 (2004 ) 高黎贡山种子植物物种丰富度沿海拔梯度的变化
. 生物多样性, 12 , 82 -88 .]
[本文引用: 1]
[54]
Wang ZH Tang ZY Fang JY 2007 ) Altitudinal patterns of seed plant richness in the Gaoligong Mountains, southeast Tibet, China
. Diversity and Distributions , 13 , 845 -854 .
[本文引用: 1]
[55]
Whittaker RH 1960 ) Vegetation of the Siskiyou Mountains, Oregon and California
. Ecological Monographs , 30 , 279 -338 .
[本文引用: 1]
[56]
Wu SH Zhang YP 2009 ) The Special Environmental Patterns and Ecological Impacts in the Longitudinal Range-Gorge Region . Science Press, Beijing .(in Chinese with English abstract)
[本文引用: 1]
[吴绍洪 , 张一平 (2009 ) 纵向岭谷区特殊环境格局与生态效应. 科学出版社 , 北京.]
[本文引用: 1]
[57]
Wu Y Colwell RK Han N Zhang R Wang W Quan Q Zhang C Song G Qu Y Lei F 2014 ) Understanding historical and current patterns of species richness of babblers along a 5000 m subtropical elevational gradient
. Global Ecology and Biogeography , 23 , 1167 -1176 .
[本文引用: 1]
[58]
Xu J Zhang BP Zhu YH Sun RH 2006 ) Distribution and geographical analysis of altitudinal belts in the Altun-Qilian Mountains
. Geographical Research , 25 , 977 -984 .(in Chinese with English abstract)
[许娟 , 张百平 , 朱运海 , 孙然好 (2006 ) 阿尔金山-祁连山山地植被垂直带谱分布及地学分析
. 地理研究, 25 , 977 -984 .]
[59]
Yang D Xiong DH Zhai J Li JJ Su ZA Dong YF 2012 ) Morphological characteristics and causes of gullies in Yuanmou dry-hot valley region
. Science of Soil and Water Conservation , 10 , 38 -45 .(in Chinese with English abstract)
[杨丹 , 熊东红 , 翟娟 , 李佳佳 , 苏正安 , 董一帆 (2012 ) 元谋干热河谷冲沟形态特征及其成因
. 中国水土保持科学, 10 , 38 -45 .]
[60]
Yang R Zhang BR Wang LL Yang JJ Su WH Zhang GF Yang B Zhou R 2010 ) The response of plant functional traits’ group to gradients of altitude in dry-hot valley of Yuan-Mou
. Ecology and Environmental Sciences , 24 , 49 -56 .(in Chinese with English abstract)
[杨锐 , 张博睿 , 王玲玲 , 杨建军 , 苏文华 , 张光飞 , 杨波 , 周睿 (2010 ) 元谋干热河谷植物功能性状组合的海拔梯度响应
. 生态环境学报, 24 , 49 -56 .]
[61]
Yang Y Shen Z Han J Zhongyong C 2016 ) Elevational gradients of plant species diversity along east and west slopes of Baima Snow Mountain in Southwest China
. Forests , 7 , 89 -105 .
[本文引用: 2]
[62]
Zhang JL Liu XK Shen R Shi W Zhang ZM Zhou R Ou XK 2010 ) The species quantitative & diversity characteristic of grassland community in hot-dry valley of Jinsha River
. Ecology and Environmental Sciences , 19 , 1519 -1524 .(in Chinese with English abstract)
[张建利 , 柳小康 , 沈蕊 , 施雯 , 张志明 , 周睿 , 欧晓昆 (2010 ) 金沙江流域干热河谷草地群落物种数量及多样性特征
. 生态环境学报, 19 , 1519 -1524 .]
[63]
Zhang YP Duan ZX Dou JX 2005 ) Comparison of climate characteristics between a dry-warm valley in upper reaches of Min River and a dry-hot valley of Yuanjiang River
. Resources and Environment in the Yangtze Basin , 14 (1 ), 76 -82 .(in Chinese with English abstract)
[张一平 , 段泽新 , 窦军霞 (2005 ) 岷江上游干暖河谷与元江干热河谷的气候特征比较研究
. 长江流域资源与环境, 14(1) , 76 -82 .]
[64]
Zhang ZM Wang WL Ou XK Wu WQ 2009 ) The correlation between vegetation spatial pattern and environmental factors
. Journal of Yunnan University (Natural Sciences Edition) , 31 , 311 -315 .(in Chinese with English abstract)
[本文引用: 1]
[张志明 , 王文礼 , 欧晓昆 , 吴伟强 (2009 ) 梅里雪山植被空间格局与环境因子关系分析
. 云南大学学报(自然科学版),31 , 311 -315 .]
[本文引用: 1]
[65]
Zhao L Lang NJ Zheng K Peng MJ 2006 ) A study on eco-environmental characteristics of dry and hot valley of Yunnan Province
. Forest Inventory and Planning , 31 (3 ), 114 -117 .(in Chinese with English abstract)
[赵琳 , 郎南军 , 郑科 , 彭明俊 (2006 ) 云南干热河谷生态环境特性研究
. 林业调查规划 ,31 (3 ), 114 -117 .]
Biodiversity in the mexican highlands and the interaction of geology, geography and climate within the trans-mexican volcanic belt
2015
Geographic patterns of vascular plant diversity at continental to global scales
1
2007
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
Partitioning the turnover and nestedness components of beta diversity
1
2010
... β多样性包含两个样本之间由于物种数量减少(nestedness)和物种更替(turnover)两个可加性成分(Koleffet al, 2003 ).在诸多β多样性指数中, Sim- pson相异性指数(βsim )反映了样本之间物种构成的更替速率, 且不受样方物种丰富度的影响(Baselga, 2010 ; Baselga & Leprieur, 2015 ), 故用来描述同一样带内不同海拔样方群落之间的物种更替. ...
Comparing methods to separate components of beta diversity
1
2015
... β多样性包含两个样本之间由于物种数量减少(nestedness)和物种更替(turnover)两个可加性成分(Koleffet al, 2003 ).在诸多β多样性指数中, Sim- pson相异性指数(βsim )反映了样本之间物种构成的更替速率, 且不受样方物种丰富度的影响(Baselga, 2010 ; Baselga & Leprieur, 2015 ), 故用来描述同一样带内不同海拔样方群落之间的物种更替. ...
Explaining the elevational diversity pattern of geometrid moths from borneo: a test of five hypotheses
1
2008
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
Drivers of moth species richness on tropical altitudinal gradients: a cross-regional comparison
1
2009
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
Disturbance regime alters the impact of dispersal on alpha and beta diversity in a natural metacommunity
2013
The mid-domain effect and species richness patterns: what have we learned so far?
1
2004
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
Competition between trees and grasses for both soil water and mineral nitrogen in dry savannas
2013
探索中国山地植物多样性的分布规律
1
2004
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
探索中国山地植物多样性的分布规律
1
2004
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
2
2006
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
2
2006
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
Phylogenetic beta diversity: linking ecological and evolutionary processes across space in time
1
2008
... 群落物种构成的更替主要受环境差异和距离产生的扩散限制影响(Graham & Fine, 2008 ; Novillo & Ojeda, 2014 ); 且在区域尺度上, 扩散限制的成因往往不仅仅是空间几何距离(Qian et al, 2009 ; Svenning et al, 2010 ).分隔在3条河流中的6条样带12个植被带段之间的物种更替显示了3方面因素的影响, 但最为突出的显然是地形隔离对物种分布的影响(附录2).一方面, 因为不同河流样带段之间的几何距离常常小于同一河流不同样带段之间的距离, 如N1-N2的距离就大于N1-L1、N1-L2、N2-L1、N2- L2、N2-L3以及N2-J1, 而L3-J1的距离小于L2-L3和L1-L3, 但不同河流之间样带段的物种更替率明显较大; 而同一样带上、下两段之间主要由于气候差异产生的物种更替率效应远低于区域隔离的效应.另一方面, 位于海拔梯度下部的干旱河谷灌草丛植被之间比中部森林带之间的隔离程度应该更强.可见在研究区域尺度上, 岭谷地形隔离对植物群落的物种更替具有最重要的影响.当然, 由于经、纬度方向上明显的气温和降水梯度的存在, 区域环境(特别是气候)梯度可能是未能解释的物种更替格局变异性的另一重要原因. ...
Species-richness patterns of vascular plants along seven altitudinal transects in Norway
1
2003
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
Can the mass effect explain the mid-altitudinal peak in vascular plant species richness?
1
2008
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
陆地植物群落物种多样性的梯度变化特征. 生态学报,
1
1997
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
陆地植物群落物种多样性的梯度变化特征. 生态学报,
1
1997
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
横断山——云南高原干热河谷形成原因研究. 资源科学,
1
2000
... 研究区域位于滇西北三江并流地区, 地理坐标范围为26°25′-28°24′ N, 97°55′-100°22′ E, 在北、东方向分别与西藏的林芝地区、四川甘孜藏族自治州和凉山彝族自治州毗邻, 西面与缅甸接壤.行政上这一区域包含怒江傈僳族自治州、迪庆藏族自治州和丽江市的11个县市.发源于青藏高原的怒江、澜沧江、金沙江3条大河南北贯穿本区域, 与自西向东平行排列的高黎贡山、怒山-碧罗雪山、云岭-老君山、雪山4条山脉, 形成“三江并流”的纵向平行岭谷地貌.境内最高点为海拔6,740 m的梅里雪山, 位于怒山-碧罗雪山北端, 亦是云南省最高峰; 最低点位于泸水县怒江河谷, 海拔600 m以下(何永彬等, 2000 ; 明庆忠和史庆涛, 2007 ). ...
横断山——云南高原干热河谷形成原因研究. 资源科学,
1
2000
... 研究区域位于滇西北三江并流地区, 地理坐标范围为26°25′-28°24′ N, 97°55′-100°22′ E, 在北、东方向分别与西藏的林芝地区、四川甘孜藏族自治州和凉山彝族自治州毗邻, 西面与缅甸接壤.行政上这一区域包含怒江傈僳族自治州、迪庆藏族自治州和丽江市的11个县市.发源于青藏高原的怒江、澜沧江、金沙江3条大河南北贯穿本区域, 与自西向东平行排列的高黎贡山、怒山-碧罗雪山、云岭-老君山、雪山4条山脉, 形成“三江并流”的纵向平行岭谷地貌.境内最高点为海拔6,740 m的梅里雪山, 位于怒山-碧罗雪山北端, 亦是云南省最高峰; 最低点位于泸水县怒江河谷, 海拔600 m以下(何永彬等, 2000 ; 明庆忠和史庆涛, 2007 ). ...
Tree dispersion, abundance, and diversity in a tropical dry forest
1979
西双版纳土地利用/土地覆被变化时空格局分析
2014
西双版纳土地利用/土地覆被变化时空格局分析
2014
Plant biodiversity patterns on Helan Mountain, China
2007
Altitudinal zonation of andean cryptogam communities
1
2000
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
Effects of rain, nitrogen, fire and grazing on tree recruitment and early survival in bush-encroached savanna, South Africa
2006
中国西南干热河谷植被恢复研究现状与发展趋势
2011
中国西南干热河谷植被恢复研究现状与发展趋势
2011
云南高原地区种子植物区系. 云南植物研究,
1
1995
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
云南高原地区种子植物区系. 云南植物研究,
1
1995
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
横断山区干热河谷气候及其对植被恢复的影响
1
2010
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
横断山区干热河谷气候及其对植被恢复的影响
1
2010
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
Mathematical ecology and its place among the sciences
1
1972
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
Metabolic theory and elevational diversity of vertebrate ectotherms
1
2010
... 本研究沿着海拔梯度分别在金沙江、澜沧江和怒江的干旱河谷区域设置样地, 对于3个河谷垂直方向均完全覆盖, 且不同河谷、不同经纬度之间形成海拔梯度的对比.研究三江并流地区干旱河谷的群落类型结构和物种多样性的垂直变化格局及其水平对比(McCain, 2010 ), 旨在探讨以下3个方面的问题: (1)该地区植物多样性的海拔格局及不同山地之间的差异; (2)影响三江并流地区植物群落α和β多样性格局的空间和地形因子; (3)探讨平行的纵向岭谷地形对植物群落物种构成所产生的隔离效应.通过探寻该区域植物多样性的垂直分布规律及区域差异, 本研究可为该区域生物多样性保护重点的选择提供依据. ...
三江并流区干热河谷成因新探析
1
2007
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
三江并流区干热河谷成因新探析
1
2007
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
Biodiversity hotspots for conservation priorities
1
2000
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
广西大青山次生林的群落特征及主要乔木种群的空间分布格局
2015
广西大青山次生林的群落特征及主要乔木种群的空间分布格局
2015
Elevation patterns in rodent diversity in the dry Andes: disentangling the role of environmental factors
1
2014
... 群落物种构成的更替主要受环境差异和距离产生的扩散限制影响(Graham & Fine, 2008 ; Novillo & Ojeda, 2014 ); 且在区域尺度上, 扩散限制的成因往往不仅仅是空间几何距离(Qian et al, 2009 ; Svenning et al, 2010 ).分隔在3条河流中的6条样带12个植被带段之间的物种更替显示了3方面因素的影响, 但最为突出的显然是地形隔离对物种分布的影响(附录2).一方面, 因为不同河流样带段之间的几何距离常常小于同一河流不同样带段之间的距离, 如N1-N2的距离就大于N1-L1、N1-L2、N2-L1、N2- L2、N2-L3以及N2-J1, 而L3-J1的距离小于L2-L3和L1-L3, 但不同河流之间样带段的物种更替率明显较大; 而同一样带上、下两段之间主要由于气候差异产生的物种更替率效应远低于区域隔离的效应.另一方面, 位于海拔梯度下部的干旱河谷灌草丛植被之间比中部森林带之间的隔离程度应该更强.可见在研究区域尺度上, 岭谷地形隔离对植物群落的物种更替具有最重要的影响.当然, 由于经、纬度方向上明显的气温和降水梯度的存在, 区域环境(特别是气候)梯度可能是未能解释的物种更替格局变异性的另一重要原因. ...
Climatic gradients in woody plant (tree and shrub) diversity: water-energy dynamics, residual variation, and topography
2000
金沙江干热河谷植物区系和生态多样性的初步研究
2
1996
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
... 这两种α多样性海拔梯度模式的存在与区域气候的整体格局应有密切联系.由于印度洋季风对区域气候的主导性影响, 区域内降水整体上呈现西高东低、南高北低的趋势(吴绍洪等, 2009 ), 高黎贡山北段西坡和白马雪山北段东坡分别成为区域的“湿极”和“旱极”, 因此在海拔梯度的下部, 流域内从西向东、从南向北所受的干旱胁迫渐趋严重, 而山地中上部的森林带内湿度差逐渐减小.这种环境变化可以解释诸样带之间灌木种丰富度自西向东显著增加, 而草本种和乔木种丰富度(不显著)降低的格局(表2 ); 而灌木和草本种丰富度随海拔上升而显著增加的趋势也应与水分条件同方向改善有直接的联系.同时, 水分条件的限制也强化了局部地形特征对物种丰富度的影响, 如草本和乔木种的丰富度从北坡向南坡降低, 灌木种则相反; 草本和乔木种丰富度随坡度增大而减少, 灌木种则增加.这3种生长型的种丰富度在局部和区域尺度上的差异性变化具有一致性的格局, 显示水分条件在不同尺度上对区内植物物种多样性分布具有突出的限制作用, 以及灌木生长型对干旱气候的适应.灌草丛带的植被极为特殊, 与非洲和印度等地大量分布的稀树干草原(Savanna)植被的外貌、结构和区系等相似(欧晓昆和金振洲, 1996 ).灌木生长型主要存在于灌草丛带, 其优势种以马棘、清香木、地菍、地桃花、川西白刺花等适应干旱气候的灌木为主, 自西向东不同流域水分减少, 使得适应湿润气候的草本和乔木物种减少, 适应干旱气候的灌木物种优势突出, 稀树干草原植被更为典型, 从而造成了灌木生长型与木本和草本的分布差异. ...
金沙江干热河谷植物区系和生态多样性的初步研究
2
1996
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
... 这两种α多样性海拔梯度模式的存在与区域气候的整体格局应有密切联系.由于印度洋季风对区域气候的主导性影响, 区域内降水整体上呈现西高东低、南高北低的趋势(吴绍洪等, 2009 ), 高黎贡山北段西坡和白马雪山北段东坡分别成为区域的“湿极”和“旱极”, 因此在海拔梯度的下部, 流域内从西向东、从南向北所受的干旱胁迫渐趋严重, 而山地中上部的森林带内湿度差逐渐减小.这种环境变化可以解释诸样带之间灌木种丰富度自西向东显著增加, 而草本种和乔木种丰富度(不显著)降低的格局(表2 ); 而灌木和草本种丰富度随海拔上升而显著增加的趋势也应与水分条件同方向改善有直接的联系.同时, 水分条件的限制也强化了局部地形特征对物种丰富度的影响, 如草本和乔木种的丰富度从北坡向南坡降低, 灌木种则相反; 草本和乔木种丰富度随坡度增大而减少, 灌木种则增加.这3种生长型的种丰富度在局部和区域尺度上的差异性变化具有一致性的格局, 显示水分条件在不同尺度上对区内植物物种多样性分布具有突出的限制作用, 以及灌木生长型对干旱气候的适应.灌草丛带的植被极为特殊, 与非洲和印度等地大量分布的稀树干草原(Savanna)植被的外貌、结构和区系等相似(欧晓昆和金振洲, 1996 ).灌木生长型主要存在于灌草丛带, 其优势种以马棘、清香木、地菍、地桃花、川西白刺花等适应干旱气候的灌木为主, 自西向东不同流域水分减少, 使得适应湿润气候的草本和乔木物种减少, 适应干旱气候的灌木物种优势突出, 稀树干草原植被更为典型, 从而造成了灌木生长型与木本和草本的分布差异. ...
Global patterns of phylogenetic beta diversity components in bats
1
2014
... 怒江、澜沧江和金沙江三个流域6条样带植物种α和β多样性海拔梯度的变化格局, 反映了它们与区域内显著的地形起伏隔离、气候垂直梯度与地理分异之间的联系, 这也许正是区内生物多样性丰富的重要成因.大量的生物多样性区域研究也都证实了气候和环境隔离因素的贡献(Peixoto et al, 2014 ; Yang et al, 2016 ).本研究没有直接分析物种丰富度和物种更替的空间格局与气候因子的定量关系, 主要是因为在这一地形起伏剧烈的区域, 缺少高空间分辨率的气候数据来准确描述气温和降水的地理变异, 而基于少数气象台站的观测数据难以胜任相关的分析.随着更多观测数据的积累, 未来有望更细致地分析和理解本区生物多样性及其分异格局的环境机制. ...
The latitudinal gradient of beta diversity in relation to climate and topography for mammals in North America
1
2009
... 群落物种构成的更替主要受环境差异和距离产生的扩散限制影响(Graham & Fine, 2008 ; Novillo & Ojeda, 2014 ); 且在区域尺度上, 扩散限制的成因往往不仅仅是空间几何距离(Qian et al, 2009 ; Svenning et al, 2010 ).分隔在3条河流中的6条样带12个植被带段之间的物种更替显示了3方面因素的影响, 但最为突出的显然是地形隔离对物种分布的影响(附录2).一方面, 因为不同河流样带段之间的几何距离常常小于同一河流不同样带段之间的距离, 如N1-N2的距离就大于N1-L1、N1-L2、N2-L1、N2- L2、N2-L3以及N2-J1, 而L3-J1的距离小于L2-L3和L1-L3, 但不同河流之间样带段的物种更替率明显较大; 而同一样带上、下两段之间主要由于气候差异产生的物种更替率效应远低于区域隔离的效应.另一方面, 位于海拔梯度下部的干旱河谷灌草丛植被之间比中部森林带之间的隔离程度应该更强.可见在研究区域尺度上, 岭谷地形隔离对植物群落的物种更替具有最重要的影响.当然, 由于经、纬度方向上明显的气温和降水梯度的存在, 区域环境(特别是气候)梯度可能是未能解释的物种更替格局变异性的另一重要原因. ...
The role of spatial scale and the perception of large-scale species-richness patterns
2005
Differences in growth patterns between co-occurring forest and savanna trees affect the forest-savanna boundary
1
2009
... 不同植被类型之间的交错带通常导致海拔梯度上β多样性的峰值(沈泽昊等, 2004 ; Rossatto, 2009 ).在研究区内三个流域的6条植被样带上, β多样性同样显示了多种海拔梯度格局, 包括单峰、上升梯度、平台甚至中部低谷等(图3 ).不同位置的样带所显示出的β多样性海拔梯度变化均有不同, 这主要与样带每个海拔梯度上所取样方较少从而导致物种随机变化较大有关.但总体来看, 这些起伏之中存在的峰值都与样带上不同植被类型之间的过渡带海拔密切相关.白马雪山东、西坡样带(J1和L3)处于同一纬度, 灌丛带过渡到森林带的海拔都在2,700-2,900 m.澜沧江中、南段的两条样带L1和L2上这一植被交错带在2,300-2,400 m之间也对应于β多样性峰值.怒江北部N2样带的β多样性海拔梯度在1,900-2,000 m出现峰值, 南部N1样带的β多样性海拔梯度在2,100-2,200 m出现峰值, 均对应于两条样带的植被交错带.β多样性主要受到空间距离和环境距离的影响, 本研究中样带内样方之间的空间距离基本相近, 差别在于环境距离不同.环境距离是由环境因子差异所产生, 各海拔梯度之间海拔距离相近, 依赖于海拔变化所产生的温度和降水变化也相近, 因此植被类型的变化便成为了影响环境变化的主要因素.在剧烈的环境变化条件下, 植物物种的β多样性也会显示出峰值.也正是这些原因形成了β多样性随海拔呈波浪式变化, 且在植被类型过渡带处呈现峰值的特点. ...
Scale-dependent effects of grazing and topographic heterogeneity on plant species richness in a Dutch salt marsh ecosystem
2014
Differential seedling establishment of woody plants along a tree density gradient in Neotropical savannas
2012
Woody cover in African savannas: the role of resources, fire and herbivory
2008
元江流域干热河谷草地植物群落结构特征与相似性分析. 生态环境学报,
2010
元江流域干热河谷草地植物群落结构特征与相似性分析. 生态环境学报,
2010
神农架南坡植物群落多样性的海拔梯度格局
1
2004
... 不同植被类型之间的交错带通常导致海拔梯度上β多样性的峰值(沈泽昊等, 2004 ; Rossatto, 2009 ).在研究区内三个流域的6条植被样带上, β多样性同样显示了多种海拔梯度格局, 包括单峰、上升梯度、平台甚至中部低谷等(图3 ).不同位置的样带所显示出的β多样性海拔梯度变化均有不同, 这主要与样带每个海拔梯度上所取样方较少从而导致物种随机变化较大有关.但总体来看, 这些起伏之中存在的峰值都与样带上不同植被类型之间的过渡带海拔密切相关.白马雪山东、西坡样带(J1和L3)处于同一纬度, 灌丛带过渡到森林带的海拔都在2,700-2,900 m.澜沧江中、南段的两条样带L1和L2上这一植被交错带在2,300-2,400 m之间也对应于β多样性峰值.怒江北部N2样带的β多样性海拔梯度在1,900-2,000 m出现峰值, 南部N1样带的β多样性海拔梯度在2,100-2,200 m出现峰值, 均对应于两条样带的植被交错带.β多样性主要受到空间距离和环境距离的影响, 本研究中样带内样方之间的空间距离基本相近, 差别在于环境距离不同.环境距离是由环境因子差异所产生, 各海拔梯度之间海拔距离相近, 依赖于海拔变化所产生的温度和降水变化也相近, 因此植被类型的变化便成为了影响环境变化的主要因素.在剧烈的环境变化条件下, 植物物种的β多样性也会显示出峰值.也正是这些原因形成了β多样性随海拔呈波浪式变化, 且在植被类型过渡带处呈现峰值的特点. ...
神农架南坡植物群落多样性的海拔梯度格局
1
2004
... 不同植被类型之间的交错带通常导致海拔梯度上β多样性的峰值(沈泽昊等, 2004 ; Rossatto, 2009 ).在研究区内三个流域的6条植被样带上, β多样性同样显示了多种海拔梯度格局, 包括单峰、上升梯度、平台甚至中部低谷等(图3 ).不同位置的样带所显示出的β多样性海拔梯度变化均有不同, 这主要与样带每个海拔梯度上所取样方较少从而导致物种随机变化较大有关.但总体来看, 这些起伏之中存在的峰值都与样带上不同植被类型之间的过渡带海拔密切相关.白马雪山东、西坡样带(J1和L3)处于同一纬度, 灌丛带过渡到森林带的海拔都在2,700-2,900 m.澜沧江中、南段的两条样带L1和L2上这一植被交错带在2,300-2,400 m之间也对应于β多样性峰值.怒江北部N2样带的β多样性海拔梯度在1,900-2,000 m出现峰值, 南部N1样带的β多样性海拔梯度在2,100-2,200 m出现峰值, 均对应于两条样带的植被交错带.β多样性主要受到空间距离和环境距离的影响, 本研究中样带内样方之间的空间距离基本相近, 差别在于环境距离不同.环境距离是由环境因子差异所产生, 各海拔梯度之间海拔距离相近, 依赖于海拔变化所产生的温度和降水变化也相近, 因此植被类型的变化便成为了影响环境变化的主要因素.在剧烈的环境变化条件下, 植物物种的β多样性也会显示出峰值.也正是这些原因形成了β多样性随海拔呈波浪式变化, 且在植被类型过渡带处呈现峰值的特点. ...
中国亚热带地区植物区系地理成分及其空间格局的数量分析
1
2000
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
中国亚热带地区植物区系地理成分及其空间格局的数量分析
1
2000
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
Environmental correlates for tropical tree diversity and distribution patterns in Borneo
2009
Extending rapoport's rule to pacific marine fishes
1
1996
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
高黎贡山乔木树种丰富度快速评估及其环境解释
1
2007
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
高黎贡山乔木树种丰富度快速评估及其环境解释
1
2007
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
Geography, topogra- phy, and history affect realized-to-potential tree species richness patterns in Europe
1
2010
... 群落物种构成的更替主要受环境差异和距离产生的扩散限制影响(Graham & Fine, 2008 ; Novillo & Ojeda, 2014 ); 且在区域尺度上, 扩散限制的成因往往不仅仅是空间几何距离(Qian et al, 2009 ; Svenning et al, 2010 ).分隔在3条河流中的6条样带12个植被带段之间的物种更替显示了3方面因素的影响, 但最为突出的显然是地形隔离对物种分布的影响(附录2).一方面, 因为不同河流样带段之间的几何距离常常小于同一河流不同样带段之间的距离, 如N1-N2的距离就大于N1-L1、N1-L2、N2-L1、N2- L2、N2-L3以及N2-J1, 而L3-J1的距离小于L2-L3和L1-L3, 但不同河流之间样带段的物种更替率明显较大; 而同一样带上、下两段之间主要由于气候差异产生的物种更替率效应远低于区域隔离的效应.另一方面, 位于海拔梯度下部的干旱河谷灌草丛植被之间比中部森林带之间的隔离程度应该更强.可见在研究区域尺度上, 岭谷地形隔离对植物群落的物种更替具有最重要的影响.当然, 由于经、纬度方向上明显的气温和降水梯度的存在, 区域环境(特别是气候)梯度可能是未能解释的物种更替格局变异性的另一重要原因. ...
云南哈巴雪山植物群落和植物多样性海拔梯度分布格局
2011
云南哈巴雪山植物群落和植物多样性海拔梯度分布格局
2011
植物物种多样性的垂直分布格局
1
2004
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
植物物种多样性的垂直分布格局
1
2004
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
The contribution of patch-scale conditions is greater than that of macroclimate in explaining local plant diversity in fragmented forests across Europe
2015
Do groundwater dynamics drive spatial patterns of tree density and diversity in Neotropical savannas
2014
金沙江干热河谷人工植被土壤水环境
2
2004
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
金沙江干热河谷人工植被土壤水环境
2
2004
... 在我国滇西北三江并流地区具有平行岭谷地貌格局, 大致南北走向、自西向东平行排列的山脉与镶嵌其中的江河谷底之间形成悬殊的海拔梯度.从河谷到山顶显著的气候垂直分异, 主导了包含热带、亚热带、温带到高山寒带等多个植被类型的植被垂直带谱(Wang et al, 2004 ; 冯建孟等, 2006 ), 并普遍为海拔基带干旱炎热、西南季风影响下干湿季明显的气候特征(张荣祖, 1992 ).横断山脉是我国和全球生物多样性的热点区域之一(Myers et al, 2000 ; Barthlott et al, 2007 ), 滇西北地区位于其东南端, 这一地区也是云南省两大生物多样性中心之一(李锡文, 1995 ).本区域已有不少关于生物多样性海拔梯度格局的研究, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度的分布格局研究, 张志明等(2009)对梅里雪山植被空间格局与环境因子关系的分析, 冯建孟等(2006)对玉龙雪山植物物种多样性沿海拔梯度的分布格局研究, Yang等(2016)用白马雪山东、西坡植物多样性梯度格局的比较验证了不同机制性假说.Alicia等(2015)曾指出, 空间异质性对于局地物种多样性的解释高于宏观气候因子, Bram等(2013)也指出, 空间隔离通过阻止物种的扩散形成了物种间的较大差别.纵向岭谷区域具有峡谷和山峰交错分布的特点, 为生物提供南北通道的同时对其东西分布形成了空间隔离(欧晓昆, 2009 ).但是, 对于区域内植物多样性海拔梯度格局沿南北纬度梯度的变化和自西向东不同流域之间的差异, 以及平行岭谷的地貌隔离对植物群落物种多样性的影响, 现有研究还尚未涉及. ...
... 区域在高山深谷地貌和立体气候的影响下, 植被沿海拔显示了层次分明的垂直带谱.各河流两侧沿海拔梯度自下而上分布着河谷(疏林)灌丛带、森林带、高山灌丛草甸带和冰雪带.区内三条江河谷底部的高度差不同, 怒江河谷水面海拔700-1,700 m, 澜沧江河谷海拔1,200-2,000 m, 金沙江河谷海拔1,600-2,000 m.怒江河谷低地以云南松(Pinus yunnanensis )次生林和半湿润的常绿阔叶林为主, 而澜沧江和金沙江河谷植被群落外貌多为灌丛或稀树草地, 无明显乔木层(欧晓昆和金振洲, 1996 ).由于气候普遍干热, 河谷低地主要的土壤类型有燥红土、褐红壤、赤红壤、紫色土等(王克勤等, 2004 ;刘方炎, 2010 ). ...
高黎贡山种子植物物种丰富度沿海拔梯度的变化
1
2004
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
高黎贡山种子植物物种丰富度沿海拔梯度的变化
1
2004
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
Altitudinal patterns of seed plant richness in the Gaoligong Mountains, southeast Tibet, China
1
2007
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
Vegetation of the Siskiyou Mountains, Oregon and California
1
1960
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
1
2009
... 这两种α多样性海拔梯度模式的存在与区域气候的整体格局应有密切联系.由于印度洋季风对区域气候的主导性影响, 区域内降水整体上呈现西高东低、南高北低的趋势(吴绍洪等, 2009 ), 高黎贡山北段西坡和白马雪山北段东坡分别成为区域的“湿极”和“旱极”, 因此在海拔梯度的下部, 流域内从西向东、从南向北所受的干旱胁迫渐趋严重, 而山地中上部的森林带内湿度差逐渐减小.这种环境变化可以解释诸样带之间灌木种丰富度自西向东显著增加, 而草本种和乔木种丰富度(不显著)降低的格局(表2 ); 而灌木和草本种丰富度随海拔上升而显著增加的趋势也应与水分条件同方向改善有直接的联系.同时, 水分条件的限制也强化了局部地形特征对物种丰富度的影响, 如草本和乔木种的丰富度从北坡向南坡降低, 灌木种则相反; 草本和乔木种丰富度随坡度增大而减少, 灌木种则增加.这3种生长型的种丰富度在局部和区域尺度上的差异性变化具有一致性的格局, 显示水分条件在不同尺度上对区内植物物种多样性分布具有突出的限制作用, 以及灌木生长型对干旱气候的适应.灌草丛带的植被极为特殊, 与非洲和印度等地大量分布的稀树干草原(Savanna)植被的外貌、结构和区系等相似(欧晓昆和金振洲, 1996 ).灌木生长型主要存在于灌草丛带, 其优势种以马棘、清香木、地菍、地桃花、川西白刺花等适应干旱气候的灌木为主, 自西向东不同流域水分减少, 使得适应湿润气候的草本和乔木物种减少, 适应干旱气候的灌木物种优势突出, 稀树干草原植被更为典型, 从而造成了灌木生长型与木本和草本的分布差异. ...
1
2009
... 这两种α多样性海拔梯度模式的存在与区域气候的整体格局应有密切联系.由于印度洋季风对区域气候的主导性影响, 区域内降水整体上呈现西高东低、南高北低的趋势(吴绍洪等, 2009 ), 高黎贡山北段西坡和白马雪山北段东坡分别成为区域的“湿极”和“旱极”, 因此在海拔梯度的下部, 流域内从西向东、从南向北所受的干旱胁迫渐趋严重, 而山地中上部的森林带内湿度差逐渐减小.这种环境变化可以解释诸样带之间灌木种丰富度自西向东显著增加, 而草本种和乔木种丰富度(不显著)降低的格局(表2 ); 而灌木和草本种丰富度随海拔上升而显著增加的趋势也应与水分条件同方向改善有直接的联系.同时, 水分条件的限制也强化了局部地形特征对物种丰富度的影响, 如草本和乔木种的丰富度从北坡向南坡降低, 灌木种则相反; 草本和乔木种丰富度随坡度增大而减少, 灌木种则增加.这3种生长型的种丰富度在局部和区域尺度上的差异性变化具有一致性的格局, 显示水分条件在不同尺度上对区内植物物种多样性分布具有突出的限制作用, 以及灌木生长型对干旱气候的适应.灌草丛带的植被极为特殊, 与非洲和印度等地大量分布的稀树干草原(Savanna)植被的外貌、结构和区系等相似(欧晓昆和金振洲, 1996 ).灌木生长型主要存在于灌草丛带, 其优势种以马棘、清香木、地菍、地桃花、川西白刺花等适应干旱气候的灌木为主, 自西向东不同流域水分减少, 使得适应湿润气候的草本和乔木物种减少, 适应干旱气候的灌木物种优势突出, 稀树干草原植被更为典型, 从而造成了灌木生长型与木本和草本的分布差异. ...
Understanding historical and current patterns of species richness of babblers along a 5000 m subtropical elevational gradient
1
2014
... 海拔对物种丰富度垂直格局的影响随着区域位置和植物类群的不同而存在较大差异(Kessler, 2000 ; 唐志尧和方精云, 2004 ), 主要有4种形式: 单调下降、先平台后下降、中部隆起(hump-shaped)以及先下降后升高(inverse hump-shaped curves).迄今为止, 大多数研究的结果表明, 山地气温和降水梯度的综合效应决定最优生境在海拔梯度上的分布, 这对应于物种丰富度的峰值海拔; 地形海拔上下限制所造成的“中域效应”决定物种丰富度在海拔中部达到最大, 但其影响大小尚不确定(Grytnes et al, 2008 ; Beck & Chey, 2008 ).最近研究发现进化因素也是物种丰富度的高峰值出现的机制之一(Wu et al, 2014 ).然而目前诸多成因对物种丰富度垂直格局的影响大小还没有形成一致结论, 因此不同山体、不同区域生物多样性的比较研究对于阐明物种丰富度和海拔关系十分必要(Grytnes, 2003 ; Beck & Kitching, 2009 ). ...
阿尔金山-祁连山山地植被垂直带谱分布及地学分析
2006
阿尔金山-祁连山山地植被垂直带谱分布及地学分析
2006
元谋干热河谷植物功能性状组合的海拔梯度响应
2010
元谋干热河谷植物功能性状组合的海拔梯度响应
2010
Elevational gradients of plant species diversity along east and west slopes of Baima Snow Mountain in Southwest China
2
2016
... 生物多样性的空间分布格局及其潜在机制是生态学和生物地理学领域的重要研究内容(Whittaker, 1960 ; MacArthur, 1972 ).过去30年来, 山地生物多样性随海拔梯度的变化规律及其影响因素在国内外生态学界受到广泛关注(贺金生和陈伟烈, 1997 ; 沈泽昊和张新时, 2000 ; 方精云, 2004 ; Rahbek, 2005 ), 并成为检验各种生物多样性机制假说的“模式对象”(Stevens, 1996 ; Colwell et al, 2004 ; Yang et al, 2016 ). ...
... 怒江、澜沧江和金沙江三个流域6条样带植物种α和β多样性海拔梯度的变化格局, 反映了它们与区域内显著的地形起伏隔离、气候垂直梯度与地理分异之间的联系, 这也许正是区内生物多样性丰富的重要成因.大量的生物多样性区域研究也都证实了气候和环境隔离因素的贡献(Peixoto et al, 2014 ; Yang et al, 2016 ).本研究没有直接分析物种丰富度和物种更替的空间格局与气候因子的定量关系, 主要是因为在这一地形起伏剧烈的区域, 缺少高空间分辨率的气候数据来准确描述气温和降水的地理变异, 而基于少数气象台站的观测数据难以胜任相关的分析.随着更多观测数据的积累, 未来有望更细致地分析和理解本区生物多样性及其分异格局的环境机制. ...
金沙江流域干热河谷草地群落物种数量及多样性特征
2010
金沙江流域干热河谷草地群落物种数量及多样性特征
2010
岷江上游干暖河谷与元江干热河谷的气候特征比较研究
2005
岷江上游干暖河谷与元江干热河谷的气候特征比较研究
2005
梅里雪山植被空间格局与环境因子关系分析
1
2009
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...
梅里雪山植被空间格局与环境因子关系分析
1
2009
... 作为生物多样性的全球热点区域, 滇西北三江并流地区的植被和生物多样性分布格局引人注目, 尤其是独具特色的平行岭谷地貌带来的显著海拔梯度分异(明庆忠和史正涛, 2007 ).此前, 对于境内的高黎贡山(王志恒等, 2004 ; 孙振华等, 2007 ; Wang et al, 2007 )、梅里雪山(张志明等, 2009 )、玉龙雪山(冯建孟等, 2006 )等已有不少相关的研究, 但都是基于区域物种分布海拔范围数据推算的结果, 均发现这些山地植物物种丰富度的海拔分布呈现单峰格局.Yang等(2016)基于1,000 m2 样带对白马雪山东西坡物种丰富度进行的对比分析也大致反映了类似的趋势.但是, 本文比较3个流域中分布在南北不同纬度的6条海拔样带的α多样性格局, 发现局部尺度的物种丰富度海拔格局并不一致.其中怒江2条样带和澜沧江中、南部的2条样带都显示了在海拔2,000 m左右的高峰值, 并特别表现在灌木种和草本种, 而位于区域东北部白马雪山东西坡的金沙江样带(J1)和澜沧江样带(L3)都显示α多样性峰值在海拔3,000 m以上, 特别是木本植物. ...

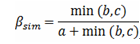


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}