1 Key Laboratory of Forest Ecology and Environment of State Forestry Administration, Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 1000912 Co-Innovation Center for Sustainable Forestry in Sounthern China, Nanjing Forestry University, Nanjing 210037
通讯作者: * 通讯作者 Author for correspondence. E-mail: ydingcaf@163.com* 通讯作者 Author for correspondence. E-mail: ydingcaf@163.com
Interspecific variation in plant functional traits is the basis of species coexistence in natural ecosystems. However, intraspecific variation is also extremely important for community assemblage and distribution. Here, we sampled 28 dominant tree species with two different leaf forms (14 evergreen species and 14 deciduous species, respectively, obtained by species abundance ranking) in a subtropical evergreen and deciduous broad-leaved mixed forest in Hubei Xingdoushan National Nature Reserve. Differences in interspecific and intraspecific variations of four functional traits were explored, including specific leaf area (SLA), leaf dry matter content (LDMC), leaf area (LA), and stem specific density (SSD). Results demonstrated that: (1) There were significant differences in the four functional traits between evergreen and deciduous trees. The SLA and LA of evergreen species were significantly lower than those of deciduous species, but LDMC and SSD showed the opposite pattern; (2) Leaf habit was the main source (57.49%) for variation of SLA. Interspecific variability (66.80%) played an important role in LA than intraspecific variability (27.52%). LDMC variation was contributed relatively evenly by interspecific (38.12%) and intraspecific (33.88%) variability. On the contrary, the variation in SSD was explained more by intraspecific (51.50%) than interspecific (32.52%) variability; (3) Correlations of different functional traits on intraspecific levels of both evergreen and deciduous trees were higher than those found for interspecific levels. Our results showed that the variation of functional traits between communities could be influenced significantly by intraspecific variability. However, there were differences in the degree of variation of different functional traits.
Keywords:functional trait
;
coefficient of variation
;
interspecific variability
;
intraspecific variability
;
evergreen and deciduous plants
;
leaf habit
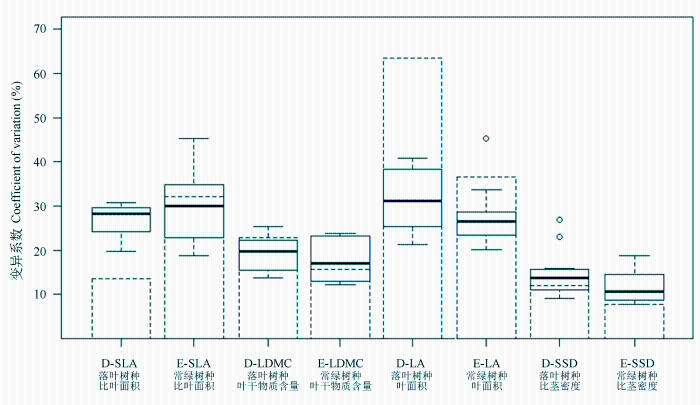
Fig. 2 Coefficient of variation of plant functional traits for plant species with different leaf habits of evergreen and deciduous broad-leaved mixed forest in Xingdoushan. The circle represents the average trait value of the species. Histograms represent interspecific coefficient of variation and box figures represent intraspecific coefficient of variation. D: Deciduous trees; E: Evergreen trees; SLA, Specific leaf area; LDMC, Leaf dry matter content; LA, Lamina area; SSD, Stem specific density.
Quantitative classification of the subtropical evergreen-deciduous broad- leaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China.
Chinese Journal of Plant Ecology, 39, 990-1002.(in Chinese with English abstract)
Relationship be- tween hydraulic resistance and leaf morphology in broadleaf Quercus species: a new interpretation of leaf lobation. Trees- Structure and Function
, 15, 341-345.
[39]
SwensonNG, EnquistBJ (2009)
Opposing assembly mechanisms in a Neotropical dry forest: implications for phylogenetic and functional community ecology.
A trait-based approach to community assembly: partitioning of species trait values into within- and among-community components.
2
2007
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific vari- ability in plant traits.
2010
Intraspecific functional variability: extent, structure and sources of variation.
2010
Interspecific and intraspecific trait variation along short environmental gradients in an old-growth temperate forest.
2
2013
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
东灵山地区不同森林群落叶功能性状比较
1
2009
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
东灵山地区不同森林群落叶功能性状比较
1
2009
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
Functional trait variation and sampling strategies in species-rich plant communities.
1
2010
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
Consequences of changing biodiversity.
1
2000
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
Ecological implications of changes in drought patterns: shifts in forest composition in Panama.
1
1998
... 2013年7-8月, 在湖北星斗山国家级自然保护区的亚热带常绿落叶阔叶混交林内设置48个空间上不连续的20 m × 20 m的森林动态样地.按照史密森研究院热带林业科学中心(CTFS)的样地建设方法(Condit, 1998), 对样地中所有胸径(DBH) ≥ 1 cm的木本植物进行标记、编号、胸径测定和坐标测量.2014年6-8月, 依照最新的植物功能性状测定方法(Perez-Harguindeguy et al, 2013), 根据森林动态样地群落调查资料, 对样地内的树木进行叶片和枝条采样.首先对每个样地内所有胸径≥ 10 cm的个体全部采样.然后根据每个样地内的物种调查数据, 对样地内胸径< 10 cm的个体按照胸径从大到小依次选择5个个体进行采样.如果每个样地中物种的个体数不足5个则全部进行采样.利用高枝剪在每个植株的林冠采集5-10枚完整无病虫害的叶片.由于森林动态样地中的树木为定期监测对象, 考虑到采样数量较多, 我们采集了2-3年生枝条进行植物功能性状测定.每个枝条样品长约10 cm, 直径约 1 cm. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.
2
2003
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
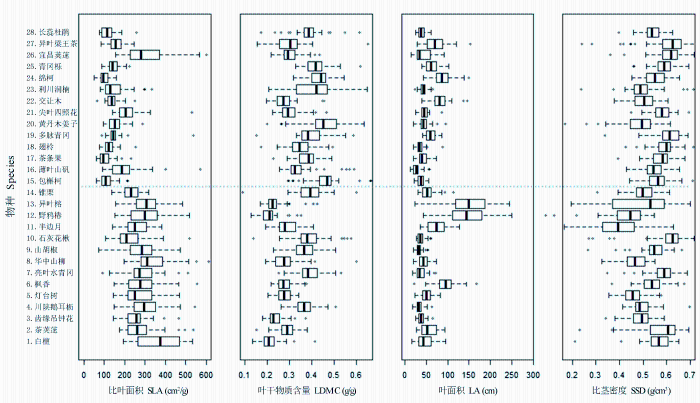
... 本研究结果表明, 4个功能性状(比叶面积、叶干物质含量、叶面积和比茎密度)在常绿和落叶树种之间均存在显著差异, 这种差异反映了不同叶习性树种形态和功能的内在区别(Tomlinson et al, 2014).对植物来说, 叶片与光照的截获和二氧化碳的同化有关, 是植物光合作用和蒸腾作用的场所; 树枝和树干等主要负责运输水分和营养物质, 并提供高度和机械支持, 从而提高植物截取光照的竞争力(Cornelissen et al, 2003).植物叶片和木材密度的差异能够反映树种在生态策略上的差异, 通常高比叶面积表明树种采用资源获取性策略, 而高叶干物质含量和比茎密度表明树种采用保守性策略.我们的结果也进一步证明了亚热带常绿落叶阔叶混交林内常绿和落叶树种的生态学差异. ...
Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California.
1
2009
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
The plant traits that drive ecosystems: evidence from three continents.
1
2004
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests.
1
2012
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
2000
2000
Environmental factors predict community functional composition in Amazonian forests.
1
2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Plant functional markers capture ecosystem properties during secondary succession.
1
2004
... 叶片是植物光合作用的最主要场所, 其功能性状或经济谱(economics spectrum)能够很好地表征养分循环、生产力等生态系统功能特征(Garnier et al, 2004; Wright et al, 2004).植物叶习性的差异是光合能力和叶寿命之间的权衡, 在一定程度上决定了植物的不同生态学策略.在森林生态系统中, 目前开展的基于个体水平的性状变异研究主要针对同一叶习性植物, 而对不同叶习性物种种内个体之间的功能性状差异的研究还比较有限.了解不同叶习性树木个体功能性状的差异不仅能够扩展基于个体水平的群落功能生态学研究(Violle et al, 2012), 而且有助于探究不同尺度下落叶和常绿树种共存的生态学机理(Quigley & Platt, 2003). ...
Structural and functional plasticity of Quercus ilex seedlings of different provenances in Italy. Trees-Structure and Function
1
2003
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
Evidence for existence of three primary strategies in plants and its relevance to ecological and evolu- tionary theory.
The withinspecies leaf economic spectrum does not predict leaf litter decomposability at either the within-species or whole community levels.
2013
Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events.
1
2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Intraspecific variability and trait-based community assembly.
2
2010
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
Trait variability differs between leaf and wood tissues across ecological scales in subtropical forests.
2
2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees.
Functional traits and niche-based tree community assembly in an Amazonian forest.
1
2008
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
Intraspecific variability in functional traits matters: case study of Scots pine.
2
2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
Applying trait-based models to achieve functional targets for theory-driven ecological restoration.
1
2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail.
1
2002
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
植物功能性状研究进展
1
2015
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
植物功能性状研究进展
1
2015
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
How do traits vary across ecological scales? A case for trait-based ecology.
1
2010
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
A functional approach reveals community responses to disturbances.
3
2013
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
... )及对干扰的响应(Mouillot et al, 2013). ...
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
New handbook for standardised measurement of plant functional traits worldwide.
2
2013
... 2013年7-8月, 在湖北星斗山国家级自然保护区的亚热带常绿落叶阔叶混交林内设置48个空间上不连续的20 m × 20 m的森林动态样地.按照史密森研究院热带林业科学中心(CTFS)的样地建设方法(Condit, 1998), 对样地中所有胸径(DBH) ≥ 1 cm的木本植物进行标记、编号、胸径测定和坐标测量.2014年6-8月, 依照最新的植物功能性状测定方法(Perez-Harguindeguy et al, 2013), 根据森林动态样地群落调查资料, 对样地内的树木进行叶片和枝条采样.首先对每个样地内所有胸径≥ 10 cm的个体全部采样.然后根据每个样地内的物种调查数据, 对样地内胸径< 10 cm的个体按照胸径从大到小依次选择5个个体进行采样.如果每个样地中物种的个体数不足5个则全部进行采样.利用高枝剪在每个植株的林冠采集5-10枚完整无病虫害的叶片.由于森林动态样地中的树木为定期监测对象, 考虑到采样数量较多, 我们采集了2-3年生枝条进行植物功能性状测定.每个枝条样品长约10 cm, 直径约 1 cm. ...
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
Radial changes in wood specific gravity of tropical trees: inter‐ and intraspecific variation during secondary succession.
1
2015
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Carbohydrate storage and light requirements of tropical moist and dry forest tree species.
Composition and structure of seasonally deciduous forests in the Americas.
1
2003
... 叶片是植物光合作用的最主要场所, 其功能性状或经济谱(economics spectrum)能够很好地表征养分循环、生产力等生态系统功能特征(Garnier et al, 2004; Wright et al, 2004).植物叶习性的差异是光合能力和叶寿命之间的权衡, 在一定程度上决定了植物的不同生态学策略.在森林生态系统中, 目前开展的基于个体水平的性状变异研究主要针对同一叶习性植物, 而对不同叶习性物种种内个体之间的功能性状差异的研究还比较有限.了解不同叶习性树木个体功能性状的差异不仅能够扩展基于个体水平的群落功能生态学研究(Violle et al, 2012), 而且有助于探究不同尺度下落叶和常绿树种共存的生态学机理(Quigley & Platt, 2003). ...
Convergent effects of elevation on functional leaf traits within and among species.
1
2014
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems.
A global meta-analysis of the relative extent of intraspecific trait variation in plant communities.
1
2015
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
Relationship be- tween hydraulic resistance and leaf morphology in broadleaf Quercus species: a new interpretation of leaf lobation. Trees- Structure and Function
2001
Opposing assembly mechanisms in a Neotropical dry forest: implications for phylogenetic and functional community ecology.
1
2009
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
Relative growth rate variation of evergreen and deciduous savanna tree species is driven by different traits.
1
2014
... 本研究结果表明, 4个功能性状(比叶面积、叶干物质含量、叶面积和比茎密度)在常绿和落叶树种之间均存在显著差异, 这种差异反映了不同叶习性树种形态和功能的内在区别(Tomlinson et al, 2014).对植物来说, 叶片与光照的截获和二氧化碳的同化有关, 是植物光合作用和蒸腾作用的场所; 树枝和树干等主要负责运输水分和营养物质, 并提供高度和机械支持, 从而提高植物截取光照的竞争力(Cornelissen et al, 2003).植物叶片和木材密度的差异能够反映树种在生态策略上的差异, 通常高比叶面积表明树种采用资源获取性策略, 而高叶干物质含量和比茎密度表明树种采用保守性策略.我们的结果也进一步证明了亚热带常绿落叶阔叶混交林内常绿和落叶树种的生态学差异. ...
The distribution of below-ground traits is explained by intrinsic species differences and intraspecific plasticity in response to root neighbours.
1
2013
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
The return of the variance: intraspecific variability in community ecology.
6
2012
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... ).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 叶片是植物光合作用的最主要场所, 其功能性状或经济谱(economics spectrum)能够很好地表征养分循环、生产力等生态系统功能特征(Garnier et al, 2004; Wright et al, 2004).植物叶习性的差异是光合能力和叶寿命之间的权衡, 在一定程度上决定了植物的不同生态学策略.在森林生态系统中, 目前开展的基于个体水平的性状变异研究主要针对同一叶习性植物, 而对不同叶习性物种种内个体之间的功能性状差异的研究还比较有限.了解不同叶习性树木个体功能性状的差异不仅能够扩展基于个体水平的群落功能生态学研究(Violle et al, 2012), 而且有助于探究不同尺度下落叶和常绿树种共存的生态学机理(Quigley & Platt, 2003). ...
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
... 本研究结果表明, 与种间变异对性状的平均贡献(37.4%)相比, 植物性状的种内平均变异贡献率(34.6%)同样不能忽略.种内变异是植物应对环境变化的重要响应机制, 是预测植物动态变化的重要内容(宝乐和刘艳红, 2009; Violle et al, 2012; Laforest-Lapointe et al, 2014).相关性分析结果表明性状间种内水平的相关性较种间水平显著.如忽略种内变异, 只考虑共存物种的平均性状值之间的相关性, 将会低估物种间生态位和性状的重叠程度(Albert et al, 2010a), 最终会导致低估物种在竞争中的相对作用(Mouillot et al, 2013).通过种内性状变异研究能更好地表现植物对特定外部环境的适应性(Gratani et al, 2003).Jung等(2014)研究表明种内变异比物种更替的相对贡献更大, 群落中的种内差异随处可见, 这也是达尔文自然选择理论的基础(Violle et al, 2012).基于种间的性状值仅仅反映不同生境下的物种组成变化情况, 而如果考虑了种内水平的功能性状变异则能够揭示物种更替、种内基因型差异及环境条件导致的形态可塑性(Auger & Shipley, 2013). ...
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提, 因此一直是植物功能性状变异研究的主要内容(Díaz et al, 2004).物种内的不同个体是构建群落结构的核心, 故植物功能性状的研究也应以物种的个体为基础(Violle et al, 2007, 2012).然而由于取样、研究尺度等原因, 许多植物功能性状研究中对种内个体性状的变异考虑较少, 通常采用植物性状平均值来反映不同环境梯度或者干扰状况下的功能性群落结构和群落构建规则.Jackson等(2013)认为种间变异对于群落构建的贡献更大, 种内变异既不能预测植物功能性状的变化, 也不能提高预测的准确性.但越来越多的证据表明, 物种间的性状变异研究具有一定的局限性, 只有结合种内和种间性状变异才可能真实反映在群落构建过程中, 种内水平上物种对环境变化和资源竞争的响应(Albert et al, 2010a, b; Jung et al, 2014; Plourde et al, 2015).Albert等(2010a)研究发现30%的功能性状变异来源于种内个体差异, 而且在不同环境和群落内, 不同功能性状的种间和种内差异也存在显著不同(Siefert et al, 2015).植物通过种内变异能很好地调节生态过程如分解作用和抵抗植食性动物(Laforest-Lapointe et al, 2014), 而且个体变异可使物种通过非生物和生物筛选过程进入局域群落(Jung et al, 2010).植物种内性状变异表征了植物沿生物和非生物梯度上的最大适应能力, 从根本上决定了植物的生态位宽度(Ackerly & Cornwell, 2007; Albert et al, 2010b).Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素.多数物种在不同环境下通过功能性状的可塑性或遗传多样性来适应不同的生境.因此探讨种内个体变异能够更好地理解群落中物种多样性维持机制, 并有助于预测不同环境条件下的群落组成(Valverde et al, 2013; Fortunel et al, 2014; Laughlin et al, 2014).在局域尺度上的群落生态学研究中, 只有考虑物种内个体水平的功能性状差异才能深入理解多物种共存机制及其群落构建规则(Violle et al, 2012; Kang et al, 2014) . ...
... 生物相互作用和生态位互补最终发生于群落内相邻的树种间, 并且可通过改变个体水平上的功能性状来适应相邻物种的生长和存活, 今后的研究中, 应该关注这种微尺度下种间和种内变异对于群落构建和物种共存的影响(Ackerly et al, 2007).基于种内水平的研究可能是群落生态学新的突破口, 也是目前群落生态学普遍关注的一个热点(Violle et al, 2007; Auger & Shipley, 2013; Kang et al, 2014).尽管在研究过程中根据实际情况会分别采用基于物种水平和种内水平的功能性状分析, 但是在分析不同生境条件时必须考虑不同生境或者样地内物种个体之间的差异(Baraloto et al, 2010).本研究中采用的方法建立在以往标准采样方法(Perez- Harguindeguy et al, 2013)基础之上, 因此与分析种间和种内性状差异的研究相比(Messier et al, 2010; Read et al, 2014), 还存在个体层次上的样本数偏小的缺陷.在局域尺度上个体变异的研究中, 进一步考虑不同林冠、个体、微环境等层次上功能性状的变化将能够进一步解释群落物种共存的重要生态学机理. ...
A structured and dynamic framework to advance traits-based theory and prediction in ecology.
1
2010
... 植物功能性状(plant functional trait)是影响植物存活、生长、繁殖速率和最终适合度的生物特征(Cornelissen et al, 2003).它是植物在漫长的进化过程中, 采用特定生态策略或属性适应周围环境的结果(刘晓娟和马克平, 2015).其中某些性状能在一定程度上反映物种对环境的适应以及对生态系统功能的效应, 因而这些植物功能性状对研究物种共存、群落构建和生态系统功能具有重要作用(Chapin et al, 2000; Lavorel & Garnier, 2002; Mouillot et al, 2013).植物功能性状的研究方法则有效地将植物个体的生理、形态和生活史特征与环境变化联系起来, 因而能够预测不同时间和空间尺度下物种组成变化对生态系统功能的影响(Webb et al, 2010)及对干扰的响应(Mouillot et al, 2013). ...
Advances, challenges and a developing synthesis of ecological community assembly theory.
1
2011
... 植物功能性状在物种间和物种内个体间的差异对于群落构建具有重要的生态学意义(Violle et al, 2012),群落构建理论表明生境和生物竞争影响性状值的分布(Weiher et al, 2011).生境过滤作用能够减少性状值的变化范围, 然而竞争又导致共存生物的生态位分化(Kraft et al, 2008; Cornwell & Ackerly, 2009; Jung et al, 2010).一方面, 栖息地通过生境过滤筛选出一组与环境条件匹配的具有相似功能性状的物种, 另一方面, 资源竞争可通过个体功能性状变异限制具有相似性状的物种, 从而形成生态位的分化.植物的性状是遗传因素、生境过滤和资源竞争共同作用的结果.本文变异分解结果表明, 叶面积的变异主要来源于种间, 受到遗传因素限制, 具有稳定的变化特性, 而比叶面积和比茎密度种内变异的贡献均大于种间变异.比叶面积和比茎密度反映的是不同植物个体的投入和产出策略, 实际上反映了群落减少竞争的部分策略.本研究结果显示比叶面积和比茎密度的种内变异均大于种间变异, 这表明环境过滤作用会导致具有相似功能性状的物种在局域生境中共存, 而这些共存物种会通过种内变异呈现某些生态策略的趋异性, 从而减少生物竞争强度, 即生境过滤和限制相似性在群落构建过程中同时发挥了重要作用(Swenson & Enquist, 2009; Ding et al, 2012). ...
The worldwide leaf economics spectrum.
1
2004
... 叶片是植物光合作用的最主要场所, 其功能性状或经济谱(economics spectrum)能够很好地表征养分循环、生产力等生态系统功能特征(Garnier et al, 2004; Wright et al, 2004).植物叶习性的差异是光合能力和叶寿命之间的权衡, 在一定程度上决定了植物的不同生态学策略.在森林生态系统中, 目前开展的基于个体水平的性状变异研究主要针对同一叶习性植物, 而对不同叶习性物种种内个体之间的功能性状差异的研究还比较有限.了解不同叶习性树木个体功能性状的差异不仅能够扩展基于个体水平的群落功能生态学研究(Violle et al, 2012), 而且有助于探究不同尺度下落叶和常绿树种共存的生态学机理(Quigley & Platt, 2003). ...
湖北星斗山天然次生林的群落类型、结构与物种多样性特征
1
2015
... 研究地点位于湖北省恩施市星斗山国家级自然保护区(29°57′-30°10′ N, 108°57′-109°27′ E).保护区总面积68,339 ha, 主峰星斗山海拔1,751.2 m.该地处于北亚热带与暖温带的过渡性地带, 属热带大陆性季风气候.年平均日照时数1,298.9 h, 年平均气温12°C, 无霜期255 d, 年平均降水量1,471.7 mm, 年平均蒸发量1,109.2 mm.降水年内季节分配不均匀, 5-9月间的降水量占全年的71%.土壤类型主要有黄壤(海拔800 m以下)、黄棕壤(海拔800- 1,500 m)和棕壤(海拔1,500 m以上).目前亚热带常绿落叶阔叶混交林为该地区的主要植被类型(姚兰等, 2015).该区域有维管束植物200科843属2,033种, 包括水杉(Metasequoia glyptostroboides)、珙桐(Davidia involucrata)、莼菜(Brasenia schreberi)、南方红豆杉(Taxus chinensis var. mairei)等8种国家I级保护植物和台湾杉(Taiwania cryptomerioides)、黄杉(Pseudotsuga sinensis)、榧树(Torreya grandis)、水青树(Tetracentron sinense)等29种国家II级保护植物. ...
湖北星斗山天然次生林的群落类型、结构与物种多样性特征
1
2015
... 研究地点位于湖北省恩施市星斗山国家级自然保护区(29°57′-30°10′ N, 108°57′-109°27′ E).保护区总面积68,339 ha, 主峰星斗山海拔1,751.2 m.该地处于北亚热带与暖温带的过渡性地带, 属热带大陆性季风气候.年平均日照时数1,298.9 h, 年平均气温12°C, 无霜期255 d, 年平均降水量1,471.7 mm, 年平均蒸发量1,109.2 mm.降水年内季节分配不均匀, 5-9月间的降水量占全年的71%.土壤类型主要有黄壤(海拔800 m以下)、黄棕壤(海拔800- 1,500 m)和棕壤(海拔1,500 m以上).目前亚热带常绿落叶阔叶混交林为该地区的主要植被类型(姚兰等, 2015).该区域有维管束植物200科843属2,033种, 包括水杉(Metasequoia glyptostroboides)、珙桐(Davidia involucrata)、莼菜(Brasenia schreberi)、南方红豆杉(Taxus chinensis var. mairei)等8种国家I级保护植物和台湾杉(Taiwania cryptomerioides)、黄杉(Pseudotsuga sinensis)、榧树(Torreya grandis)、水青树(Tetracentron sinense)等29种国家II级保护植物. ...
 , 臧润国
, 臧润国

{kind=link}
{kind=link}
{kind=link}
{kind=link}