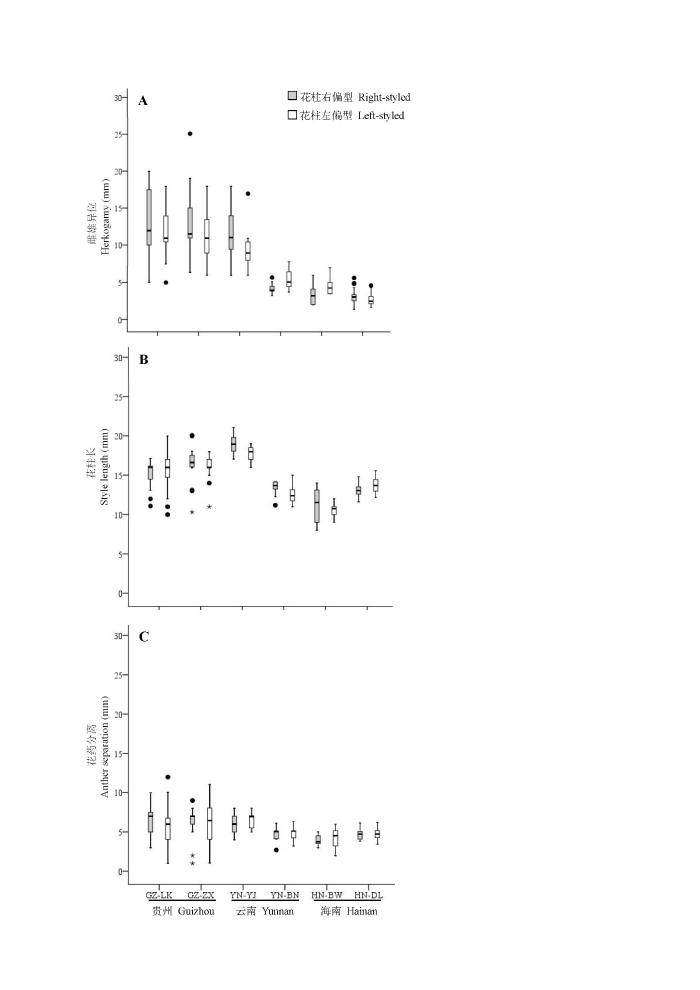
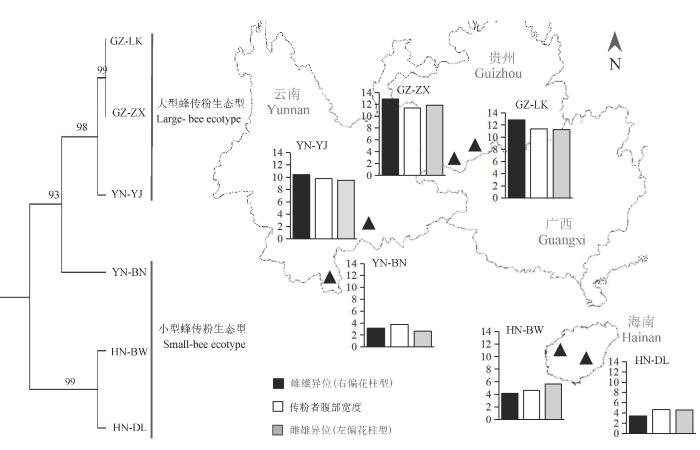
Herkogamy, the spatial separation of sexual organs within flowers, has long been thought to be a floral adaptation of reducing self pollination in hermaphrodites. Herkogamy of mirror-image flowers in the Asian endemic Hiptage benghalensis (Malpighiaceae) varies greatly between populations on Hainan Island (smaller than 4 mm) and Guizhou Province (larger than 10 mm). Such divergence in herkogamy could be adaptive to pollinator body size, which relates to pollen placement and stigma contact position. Recipient selection between local pollinators and plants may facilitate pollinator assembly in isolated populations, even forming pollination ecotypes. Pollinators were investigated in six geographic populations of H. benghalensis to study whether variations in herkogamy correspond to pollinator size. Molecular markers with nuclear ITS 1/4, chloroplast ycf1b and rbcL were also used to detect genetic differentiation between populations with large or small herkogamy. Our data showed small herkogamy in two populations from Hainan Island and one population from Xishuangbanna (southern Yunnan Province) as they were pollinated by small bees including Apis dorsata and Apis mellifera, with an abdomen width varying from 2 to 4 mm. Two populations from Guizhou Province and one population from Southeast Yunnan with larger herkogamy were pollinated by carpenter bees Xylocopa nasalis and bumble bees (Bombus sp.), with an obvious larger body size with a mean abdomen width of 11 mm. The style length (F = 12.5, P < 0.05) was also significantly different between large versus small herkogamous populations. Genetic data based on nuclear (ITS 1/4) and chloroplast (rbcL and ycf1) DNA also supported the differentiation between small and large herkogamous populations. These results indicated associations between flower and pollinator size, suggesting pollination ecotypes in H. benghalensis. This study indicated that the pollination ecotype in mirror-image flowers was associated with variations in herkogamy.
Fig. 2 Boxplot of variations in herkogamy (A), style length (B) and anther separation (C) of mirror-image flowers among Hiptage benghalensis populations
Fig. 3 Pollinators in different geographic populations of Hiptage benghalensis. (A) and (B), Xylocopa nasalis visiting GZ-LK and GZ-ZX populations in Guizhou Province; (C) Bombus sp. visiting YN-YJ Population in Yunnan Province; (D) Pollen grains (red circles) on the thorax and abdomen of a carpenter bee; (E) Apis mellifera in YN-BN Population in Yunnan Province; (F) Apis dorsata in HN-BW Population on Hainan Island.
Fig. 4 Pollination ecotypes of Hiptage benghalensis. Neighbor-joining phylogenetic tree was drawn based on ITS1/4 and chloroplast (rbcLand ycf1b) DNA sequences. The histograms on the map refer to the mean values of herkogamy, and the unit of ordinate is millimeter (mm). ■ Herkogamy (righ-styled flower); □ Pollinator abdomen width; Herkogamy (left-styled flower).
传粉生态型在兰科、杜鹃花科、鸢尾科等植物中已有报道, 往往也与花部关键性状如花距长、花冠管长的适应性变化息息相关。Robertson和Wyatt (1990)发现, 高海拔地区和低海拔地区分布的兰科植物Platantha ciliaris具有不同的矩长, 吸引不同吻长的蝴蝶为其传粉。南非开普敦地区的兰科植物Satyrium hallackii与鸢尾科植物Gladiolus longicollis的种群间也出现了不同长度花冠管适应不同吻长蝇类传粉者的传粉生态型(Johnson, 1997; Anderson et al, 2009)。杜鹃花科的Erica plukenetii也在适应鸟媒和蛾媒传粉者的过程中出现了3种传粉生态型, 花冠管长度与形态在其中扮演着关键的角色(Van der Niet et al, 2014)。风筝果的情况与此类似, 镜像花的雌雄异位作为关键性状在适应不同体型传粉者的过程中分化出传粉生态型, 也证实了镜像花这类特化传粉系统对传粉环境变化有着一定的适应灵活性。
虽然镜像花一直被认为是一种非常特化、复杂的传粉机制, 但大多数研究也发现镜像花的访花昆虫专一性并不特别强, 体型大小与访花行为近似的多种昆虫都可以是其有效的传粉者(Jesson & Barrett, 2003; Gao et al, 2006; Ren et al, 2013)。本文发现的风筝果镜像花出现了适应不同类型传粉者的两种生态型这一现象, 进一步证实了镜像花这类特化传粉系统可能具有较多样的传粉者。风筝果大蜂传粉生态型的传粉昆虫包括了体型较大的木蜂和熊蜂, 这类传粉昆虫有着体型大、动作粗暴、以花粉和花蜜为回报物的共同特点(吴燕如, 2000)。小蜂传粉生态型的传粉者则是以体型较小的西方蜜蜂与大蜜蜂为主, 这类蜜蜂属典型的热带分布昆虫, 体型偏小、访花时间较短, 对花气味有超强的记忆识别能力(卿卓等, 2014), 回报物也是花蜜与花粉(Ren et al, 2013)。
The upper reaches of the largest river in Southern China as an ‘evolutionary front’ of tropical plants: evidences from Asia-endemic genus Hiptage (Malpighiaceae).
WhiteTJ, BrunsT, LeeS, TaylorJ (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR Protocols: A Guide to Methods and Applications (eds Innis MA, Gelfand DH, Sninsky JJ, White TJ ), pp. 315-322. Academic Press, New York.
[41]
WhittallJB, HodgesSA (2007)
Pollinator shifts drive increasingly long nectar spurs in columbine flowers.
Evolution and coexistence of pollination ecotypes in an African Gladiolus (Iridaceae).
2
2009
... 由于环境因子与资源的斑块状分布, 同种植物的不同地理种群往往处于水热条件与伴生生物大不相同的微生境中.在局域不同环境条件的选择下, 不同地理种群可能在个体形态与结构、遗传背景方面都呈现出显著的不同, 形成不同的“生态型”(ecotype).生态型是植物同一种内表现出的具有遗传基础的生态分化(Parisod & Joost, 2010), 是物种形成的一个初级阶段(Johnson & Steiner, 1997; Whittall & Hodges, 2007).对于以异交传粉为主的植物而言, 不同地理种群还可能面临不同的传粉者类群与组成, 特别是异交物种拓殖到一个新分布区的先锋种群必然经历着适应局域“更有效传粉者”的强烈选择(Grant & Grant, 1965; Stebbins, 1970; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
... 传粉生态型在兰科、杜鹃花科、鸢尾科等植物中已有报道, 往往也与花部关键性状如花距长、花冠管长的适应性变化息息相关.Robertson和Wyatt (1990)发现, 高海拔地区和低海拔地区分布的兰科植物Platantha ciliaris具有不同的矩长, 吸引不同吻长的蝴蝶为其传粉.南非开普敦地区的兰科植物Satyrium hallackii与鸢尾科植物Gladiolus longicollis的种群间也出现了不同长度花冠管适应不同吻长蝇类传粉者的传粉生态型(Johnson, 1997; Anderson et al, 2009).杜鹃花科的Erica plukenetii也在适应鸟媒和蛾媒传粉者的过程中出现了3种传粉生态型, 花冠管长度与形态在其中扮演着关键的角色(Van der Niet et al, 2014).风筝果的情况与此类似, 镜像花的雌雄异位作为关键性状在适应不同体型传粉者的过程中分化出传粉生态型, 也证实了镜像花这类特化传粉系统对传粉环境变化有着一定的适应灵活性. ...
The evolution and function of stylar polymorphisms in ?owering plants.
... 虽然镜像花一直被认为是一种非常特化、复杂的传粉机制, 但大多数研究也发现镜像花的访花昆虫专一性并不特别强, 体型大小与访花行为近似的多种昆虫都可以是其有效的传粉者(Jesson & Barrett, 2003; Gao et al, 2006; Ren et al, 2013).本文发现的风筝果镜像花出现了适应不同类型传粉者的两种生态型这一现象, 进一步证实了镜像花这类特化传粉系统可能具有较多样的传粉者.风筝果大蜂传粉生态型的传粉昆虫包括了体型较大的木蜂和熊蜂, 这类传粉昆虫有着体型大、动作粗暴、以花粉和花蜜为回报物的共同特点(吴燕如, 2000).小蜂传粉生态型的传粉者则是以体型较小的西方蜜蜂与大蜜蜂为主, 这类蜜蜂属典型的热带分布昆虫, 体型偏小、访花时间较短, 对花气味有超强的记忆识别能力(卿卓等, 2014), 回报物也是花蜜与花粉(Ren et al, 2013). ...
Pollination ecotypes of Satyrium hallackii (Orchidaceae) in South Africa.
2
1997
... 由于环境因子与资源的斑块状分布, 同种植物的不同地理种群往往处于水热条件与伴生生物大不相同的微生境中.在局域不同环境条件的选择下, 不同地理种群可能在个体形态与结构、遗传背景方面都呈现出显著的不同, 形成不同的“生态型”(ecotype).生态型是植物同一种内表现出的具有遗传基础的生态分化(Parisod & Joost, 2010), 是物种形成的一个初级阶段(Johnson & Steiner, 1997; Whittall & Hodges, 2007).对于以异交传粉为主的植物而言, 不同地理种群还可能面临不同的传粉者类群与组成, 特别是异交物种拓殖到一个新分布区的先锋种群必然经历着适应局域“更有效传粉者”的强烈选择(Grant & Grant, 1965; Stebbins, 1970; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
... 传粉生态型在兰科、杜鹃花科、鸢尾科等植物中已有报道, 往往也与花部关键性状如花距长、花冠管长的适应性变化息息相关.Robertson和Wyatt (1990)发现, 高海拔地区和低海拔地区分布的兰科植物Platantha ciliaris具有不同的矩长, 吸引不同吻长的蝴蝶为其传粉.南非开普敦地区的兰科植物Satyrium hallackii与鸢尾科植物Gladiolus longicollis的种群间也出现了不同长度花冠管适应不同吻长蝇类传粉者的传粉生态型(Johnson, 1997; Anderson et al, 2009).杜鹃花科的Erica plukenetii也在适应鸟媒和蛾媒传粉者的过程中出现了3种传粉生态型, 花冠管长度与形态在其中扮演着关键的角色(Van der Niet et al, 2014).风筝果的情况与此类似, 镜像花的雌雄异位作为关键性状在适应不同体型传粉者的过程中分化出传粉生态型, 也证实了镜像花这类特化传粉系统对传粉环境变化有着一定的适应灵活性. ...
Long-tongued fly pollination and evolution of floral spur length in the Disa draconis Complex (Orchidaceae).
3
1997
... 由于环境因子与资源的斑块状分布, 同种植物的不同地理种群往往处于水热条件与伴生生物大不相同的微生境中.在局域不同环境条件的选择下, 不同地理种群可能在个体形态与结构、遗传背景方面都呈现出显著的不同, 形成不同的“生态型”(ecotype).生态型是植物同一种内表现出的具有遗传基础的生态分化(Parisod & Joost, 2010), 是物种形成的一个初级阶段(Johnson & Steiner, 1997; Whittall & Hodges, 2007).对于以异交传粉为主的植物而言, 不同地理种群还可能面临不同的传粉者类群与组成, 特别是异交物种拓殖到一个新分布区的先锋种群必然经历着适应局域“更有效传粉者”的强烈选择(Grant & Grant, 1965; Stebbins, 1970; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
... ; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
... ; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
Generalization vs. specialization in plant pollination systems.
1
2000
... 由于环境因子与资源的斑块状分布, 同种植物的不同地理种群往往处于水热条件与伴生生物大不相同的微生境中.在局域不同环境条件的选择下, 不同地理种群可能在个体形态与结构、遗传背景方面都呈现出显著的不同, 形成不同的“生态型”(ecotype).生态型是植物同一种内表现出的具有遗传基础的生态分化(Parisod & Joost, 2010), 是物种形成的一个初级阶段(Johnson & Steiner, 1997; Whittall & Hodges, 2007).对于以异交传粉为主的植物而言, 不同地理种群还可能面临不同的传粉者类群与组成, 特别是异交物种拓殖到一个新分布区的先锋种群必然经历着适应局域“更有效传粉者”的强烈选择(Grant & Grant, 1965; Stebbins, 1970; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
Specialized pollination systems in southern Africa.
... 虽然镜像花一直被认为是一种非常特化、复杂的传粉机制, 但大多数研究也发现镜像花的访花昆虫专一性并不特别强, 体型大小与访花行为近似的多种昆虫都可以是其有效的传粉者(Jesson & Barrett, 2003; Gao et al, 2006; Ren et al, 2013).本文发现的风筝果镜像花出现了适应不同类型传粉者的两种生态型这一现象, 进一步证实了镜像花这类特化传粉系统可能具有较多样的传粉者.风筝果大蜂传粉生态型的传粉昆虫包括了体型较大的木蜂和熊蜂, 这类传粉昆虫有着体型大、动作粗暴、以花粉和花蜜为回报物的共同特点(吴燕如, 2000).小蜂传粉生态型的传粉者则是以体型较小的西方蜜蜂与大蜜蜂为主, 这类蜜蜂属典型的热带分布昆虫, 体型偏小、访花时间较短, 对花气味有超强的记忆识别能力(卿卓等, 2014), 回报物也是花蜜与花粉(Ren et al, 2013). ...
花蜜化学成分及其生态功能研究进展
1
2014
... 虽然镜像花一直被认为是一种非常特化、复杂的传粉机制, 但大多数研究也发现镜像花的访花昆虫专一性并不特别强, 体型大小与访花行为近似的多种昆虫都可以是其有效的传粉者(Jesson & Barrett, 2003; Gao et al, 2006; Ren et al, 2013).本文发现的风筝果镜像花出现了适应不同类型传粉者的两种生态型这一现象, 进一步证实了镜像花这类特化传粉系统可能具有较多样的传粉者.风筝果大蜂传粉生态型的传粉昆虫包括了体型较大的木蜂和熊蜂, 这类传粉昆虫有着体型大、动作粗暴、以花粉和花蜜为回报物的共同特点(吴燕如, 2000).小蜂传粉生态型的传粉者则是以体型较小的西方蜜蜂与大蜜蜂为主, 这类蜜蜂属典型的热带分布昆虫, 体型偏小、访花时间较短, 对花气味有超强的记忆识别能力(卿卓等, 2014), 回报物也是花蜜与花粉(Ren et al, 2013). ...
The upper reaches of the largest river in Southern China as an ‘evolutionary front’ of tropical plants: evidences from Asia-endemic genus Hiptage (Malpighiaceae).
Do pollinator distributions underlie the evolution of pollination ecotypes in the Cape shrub Erica plukenetii?
2
2014
... 由于环境因子与资源的斑块状分布, 同种植物的不同地理种群往往处于水热条件与伴生生物大不相同的微生境中.在局域不同环境条件的选择下, 不同地理种群可能在个体形态与结构、遗传背景方面都呈现出显著的不同, 形成不同的“生态型”(ecotype).生态型是植物同一种内表现出的具有遗传基础的生态分化(Parisod & Joost, 2010), 是物种形成的一个初级阶段(Johnson & Steiner, 1997; Whittall & Hodges, 2007).对于以异交传粉为主的植物而言, 不同地理种群还可能面临不同的传粉者类群与组成, 特别是异交物种拓殖到一个新分布区的先锋种群必然经历着适应局域“更有效传粉者”的强烈选择(Grant & Grant, 1965; Stebbins, 1970; Johnson & Steiner, 1997; Fortuna et al, 2008).为适应最有效传粉者、促进繁殖成功, 不同地理种群的个体可能出现适应局域传粉者的花部特征调整(Robertson & Wyatt, 1990), 形成在形态特征与遗传背景都出现分化的“传粉生态型”(pollination ecotype) (Johnson, 1997; Johnson & Steiner, 1997, 2000, 2003; Anderson et al, 2009; Van der Niet et al, 2014).与其他类型的生态型相比, 传粉生态型可直接影响到传粉隔离从而更易导致物种分化(Grant & Grant, 1965), 是认识植物物种形成、花-传粉昆虫交互选择的一个重要方面. ...
... 传粉生态型在兰科、杜鹃花科、鸢尾科等植物中已有报道, 往往也与花部关键性状如花距长、花冠管长的适应性变化息息相关.Robertson和Wyatt (1990)发现, 高海拔地区和低海拔地区分布的兰科植物Platantha ciliaris具有不同的矩长, 吸引不同吻长的蝴蝶为其传粉.南非开普敦地区的兰科植物Satyrium hallackii与鸢尾科植物Gladiolus longicollis的种群间也出现了不同长度花冠管适应不同吻长蝇类传粉者的传粉生态型(Johnson, 1997; Anderson et al, 2009).杜鹃花科的Erica plukenetii也在适应鸟媒和蛾媒传粉者的过程中出现了3种传粉生态型, 花冠管长度与形态在其中扮演着关键的角色(Van der Niet et al, 2014).风筝果的情况与此类似, 镜像花的雌雄异位作为关键性状在适应不同体型传粉者的过程中分化出传粉生态型, 也证实了镜像花这类特化传粉系统对传粉环境变化有着一定的适应灵活性. ...
Sequence Matrix, concatenation software for the fast assembly of multi-gene datasets with character set and codon information.
1
2011
... 花部关键性状的数据用SPSS 13.0 for Windows统计软件分析, 与传粉昆虫体宽的关系采用单因素方差分析(One-Way ANOVA).对不同地理种群DNA测序结果用MEGA 6.0 (Tamura et al, 2013)自行比对, 将叶绿体基因序列和核基因序列用Sequence Matrix (Vaidya et al, 2011)合并, 再使用MEGA 6.0的邻接法(neighbor-joining method)对6个地理种群进行遗传关系的聚类分析, 并将种群遗传关系与雌雄异位程度的关系进行比对, 判断雌雄异位程度相差较大的种群是否也有着较远的遗传关系.使用GenAlEx 6.501 (Excoffier et al, 1992)对遗传结构进行AMOVA分析, 区分种群间与种群内的遗传变异相对比例. ...
Control of the invasive liana, Hiptage benghalensis.
1
2009
... 风筝果是多年生木质藤本植物, 主要分布于亚洲热带地区的森林林缘、河谷滩地或石灰岩裸岩, 常攀爬于高大树木, 或铺散于石上.花期3-4月, 总状花序, 每花序约有花20朵.花大, 极芳香, 花瓣5枚、白色且向后方反折, 最上方的旗瓣基部具黄色斑点(图1).风筝果除了具有特化的传粉系统“镜像花”之外, 雄蕊还分化成1大9小的异型雄蕊(陈书坤, 1977; Ren et al, 2013) (图1), 较大的1枚雄蕊可育、花粉活性高, 而9枚小雄蕊的花粉活性极低, 可能是提供花粉诱物吸引传粉昆虫的不育雄蕊(Ren et al, 2013).萼片5, 上方1枚具一分泌花蜜的大腺体(Ren et al, 2013).果实是典型的翅果, 具窄翅3.叶对生、革质, 幼叶呈现闪亮的红色.风筝果适应性较强, 花、果、叶均有较高的观赏价值, 已在国内外一些植物园和庭院引种(Vitelli et al, 2009; 任明迅, 个人观察). ...
The avoidance of interference between the presentation of pollen and stigmas in angiosperms. II. Herkogamy.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}