近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 )。一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 )。实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution)。与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用。与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 )。
生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用。前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 )。在以现实分布为目的时, 生态位模型有时也被称为物种分布模型。目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间。一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 )。Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 )。
在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 )。如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 )。Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 )。随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 )。当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释。
在本研究中, 以入侵害虫茶翅蝽(Halyomorpha halys )为例, 以预测入侵物种的潜在分布为目的, 探讨Maxent模型的复杂度对模型转移能力的影响。本研究采用经典生态模型方案, 通过构建本土模型, 然后将其转移至入侵地来模拟茶翅蝽的潜在分布, 同时采用地理空间分层的方法测试Maxent模型对本土分布数据是否过度拟合。通过调用ENMeval数据包(Muscarella et al, 2014 )来调整Maxent模型参数和分析各种参数条件下的模型复杂度, 并选取最低复杂度的模型参数, 综合比较Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响以及Maxent模型构建时的注意事项。本文结合模型的构建材料及模型的构建区域全面阐述提高模型转移能力的策略, 以期对物种潜在分布进行合理的预测, 促进Maxent模型在我国的合理运用和发展。
1 材料与方法
1.1 数据来源及处理
茶翅蝽原产于亚洲东部(中国、日本、韩国和朝鲜), 现已在北美洲和欧洲建立种群, 危害比较严重。物种分布点数据和环境变量源自Zhu等(2012, 2016)中, 已去除采样偏差对模型的影响。环境变量的选取主要考虑其对物种分布的限制作用和变量间的空间相关性(Peterson et al, 2011 ), 选取了年平均气温(bio1)、最热月份最高气温(bio5)、最冷月份最低气温(bio6)、年降雨量(bio12)和年平均辐射量(bio20)等5个环境变量(Hijmans et al, 2005 ; Kriticos et al, 2011 )。研究区域分为茶翅蝽的本土种群生存地区以及入侵地, 其中本土地区包括中国、朝鲜、韩国和日本, 入侵地以北美洲为例(Zhu et al, 2016 )。
1.2 模型构建及评价
首先采用Maxent默认参数在茶翅蝽本土地区构建模型, 然后将其转移至入侵地来检验和评价模型; 随后通过调用ENMeval数据包来调整Maxent模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数, 分析各种参数条件下模型的复杂度, 选取最低复杂度的模型参数; 最后综合比较分析Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响。Maxent模型的复杂度与其RM和FC参数密切相关。目前Maxent中有5种特征, 即: 线性(linear - L), 二次型(quadratic - Q), 片段化(hinge - H), 乘积型(product - P)和阈值性(threshold - T)。在其默认设置中, RM的值为1, 具体特征组合的选择和使用与物种分布点数量有关, 通常情况下linear feature一直在运行, quadratic feature在物种分布点>10时使用, hinge feature在分布点>15时使用, threshold和product feature在分布点>80时使用(Elith et al, 2010 )。ENMeval数据包通过测试不同参数条件下Maxent模型修正的AIC值(即AICc)来评价模型的复杂度。AIC信息量准则(Akaike information criterion correction, AIC)是衡量统计模型拟合优良性的一种标准, 可以权衡所估计模型的复杂度和此模型拟合数据的优良性, AIC信息量准则优先考虑AIC值最小的模型(Akaike, 1973 )。AIC值可通过Maxent模型运行后产生的lambdas文件计算获得(Warren & Seifert, 2011 )。在本研究中, 我们将RM设置为0.5-4, 每次增加0.5, 采用6个特征组合(FC), 即: L, LQ, H, LQH, LQHP和LQHPT。为测试Maxent模型对本土分布数据拟合的效果, 在本土区域将物种分布点进行空间分割成4等份, 随机采用其中的3份构建模型, 剩余的1份用于检验模型(Muscarella et al, 2014 )。
在本研究中, 所有物种分布数据分成3份, 其中随机选取70%的本土物种分布点用于模型构建和优化, 剩下30%的本土物种分布点用于模型内部检验, 北美入侵地物种分布点用于检验模型转移能力。Maxent模型的对比和评价主要采用响应曲线和遗漏率曲线, 同时采用Partial ROC方案测试模型的本土预测能力和转移能力(Peterson et al, 2008 )。在ENMenv数据包中, 采用AUC来测试模型在本土4份地理空间之间的相互预测的效果, AUC.diff (等于AUCtrain-AUCtest)和OR10用来测试模型对本土物种分布点的拟合程度, delta.AICc用来测试模型的复杂度和拟合程度(详细参见Muscarella et al, 2014 )。
2 结果
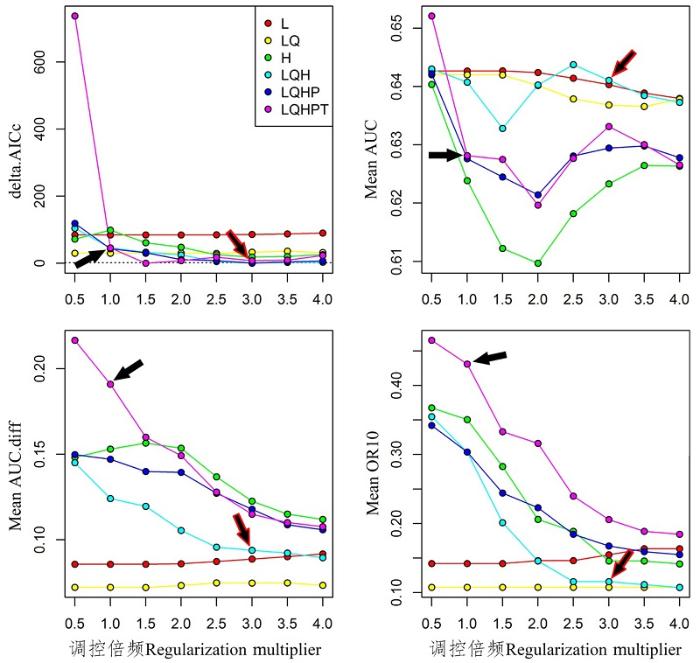
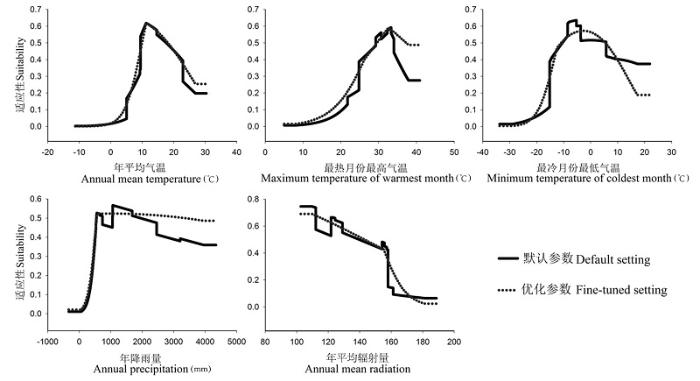
基于茶翅蝽的物种分布点数目(234个), Maxent模型在默认参数设置时, 其RM = 1, 运行的Feature有L, Q, H, P和T。基于AIC信息准则, 在Maxent的运行参数为RM = 3和运行Feature为L, Q和H时, 其AIC值最小(图1 )。在茶翅蝽本土范围内, Maxent模型进行地理空间之间相互预测时, 基于优化模型的AUC值(0.641)高于默认参数下模型的AUC值(0.628) (图1 ), 二者总体上都不高, 说明在东亚本土分布范围内, 茶翅蝽分布的生境异质性较大。基于优化模型的AUCdiff和OR10值明显低于默认参数下的Maxent模型(图1 ), 表明优化后的模型明显降低了对本土分布数据的过度拟合。在响应曲线中, 基于默认参数的Maxent模型表现出明显曲折性, 表明Maxent模型对模型构建区的物种分布数据存在过度拟合现象。相反, 当采用最优模型参数时, 响应曲线变得相对平滑(图2 ), 亦表明优化后的模型降低了Maxent模型对本土分布数据的过度拟合, 从而更接近茶翅蝽对环境因子的生理响应。
图1 不同参数下的茶翅蝽本土模型表现。黑色箭头表示Maxent默认参数, 红边箭头表示AIC值最小优化模型参数。
Fig. 1 Performances of native niche model of Halyomorpha halys under different settings. Black arrow indicates default setting, rededge arrow indicates the AICc-chosen setting.
图2 基于默认参数和优化参数的本土Maxent模型中茶翅蝽对5个气候变量的响应曲线
Fig. 2 Comparison of response curves of Halyomorpha halys to five bioclimatic variables based on the default and fine-tuned Maxent settings
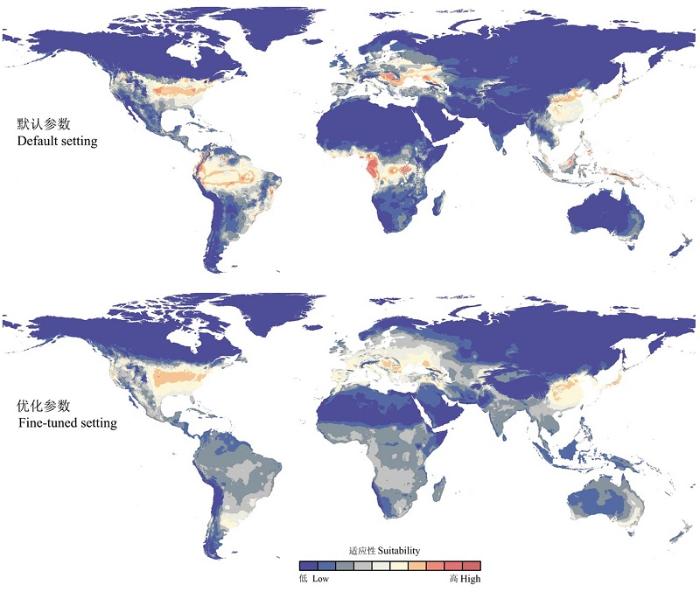
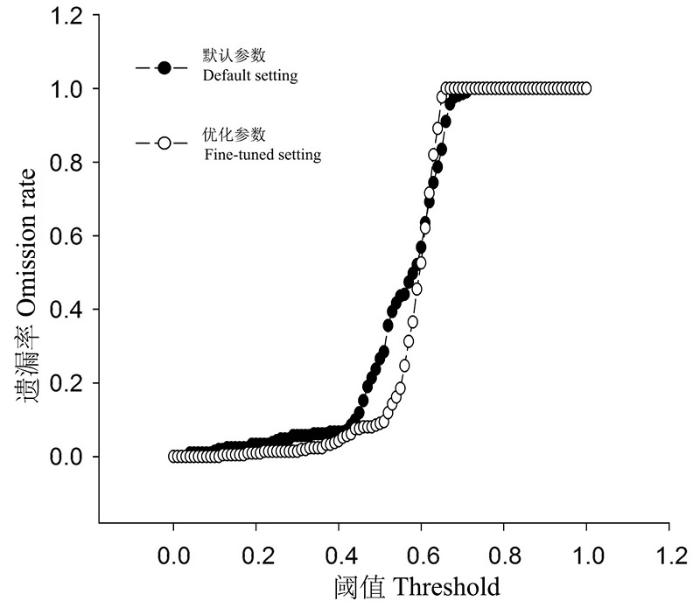
在本土模型构建区, 基于默认参数构建的Maxent模型其AUCratio值为1.526, 基于最小AICc值校正参数后的Maxent模型其AUCratio值为1.539。将Maxent模型转移至全球范围后, 基于默认参数和优化参数后模型的预测差别较大(图3 ), 特别是在南半球, 如南美洲北部和非洲的中部地区(图3 )。与南半球的预测相比, 二者在北美洲和欧洲的预测效果也有一定的差别。在北美洲, 基于默认参数和优化参数后的Maxent模型其AUCratio值分别为1.430和1.452。在遗漏率曲线中, 在不同的阈值下, 基于优化参数后的Maxent模型对入侵地分布数据的遗漏率相对较低, 表明优化后的模型转移能力较强, 在北美洲预测茶翅蝽潜在分布的效果较好(图4 )。在本土和入侵地的预测中, 优化参数后的Maxent模型均优于默认参数的模型, 对本土亚洲东部和入侵地北美洲分布的茶翅蝽种群均能够有相对较好的预测。
图3 基于默认参数和优化参数后的Maxent模型转移后对茶翅蝽的潜在分布预测
Fig. 3 Predictions of Halyomorpha halys based on its native Maxent models calibrated on the default and fine-tuned settings
图4 基于默认参数和优化后参数的Maxent模型转移后对北美分布茶翅蝽预测的遗漏率
Fig. 4 Omission rates of North American records of Halyomorpha halys based on its native Maxent models predictions using the default and fine-tuned settings.
3 讨论
生态位模型是利用物种已知分布点所关联的环境变量去推算物种的生态需求, 模拟物种的分布。在模拟入侵物种分布时, 经典生态位模型通过模型构建物种本土分布地模型, 然后将其转移并投射至另一地理区域, 来模拟入侵物种的潜在分布。然而在模型运用时, 出现了模型转移能力较低、模拟结果与物种的现实分布不相符的情况, 由此得出了生态位漂移等不恰当的结论。提高生态位模型的转移能力, 不仅可以准确地模拟物种的潜在分布, 同时对生态位保守性等理论问题具有重要的参考价值(朱耿平等, 2014 )。
3.1 Maxent模型默认参数的局限性
Maxent默认参数的设置源自早期模型开发者对6个不同地理区域的266个物种数据的测试(Phillips & Dudík, 2008 ), 他们采用海量物种的分布数据和多种实验方案, 以期得到一个最优的模型参数作为默认参数来推广和简化Maxent模型的应用, 这些物种涵盖了鸟类、哺乳类、爬行类以及植物等类群, 分布在澳洲、新西兰、欧洲和南美洲等地区。然而, 这些模型的应用均是以预测现实分布为目的, 模型构建后不需转移, 预测模型需要能够很好地辨别物种的分布与否(即区分物种的分布和不分布, Jiménez-Valverde et al, 2008 ; Phillips & Dudík, 2008 )。在生物入侵、全球变化对物种分布影响和谱系生物地理学中, 生态位模型的应用是以模拟物种的潜在分布为目的, Maxent模型构建以后需要转移至不同地理空间中去预测物种的潜在分布, 需要降低模型预测的遗漏率。虽然Maxent模型所模拟的物种分布介于潜在分布和现实分布之间(Jiménez- Valverde et al, 2011 ), 但从研究者目的出发有时需要调整模拟方案来实现模拟需求。在以模拟物种潜在分布为目的时, Maxent模型默认参数可能不再适用, 如本研究中, 茶翅蝽的最优模型参数不同于默认参数(图1 ), 如果继续采用默认参数, Maxent模型虽能够很好地拟合模型构建区的物种分布, 但由于过度拟合会造成模型转移能力降低(图4 )。此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 )。
3.2 提高模型转移能力的策略——物种分布点和环境变量
相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 )。在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意。生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 )。在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差。环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 )。
3.3 提高模型转移能力的策略——模型复杂度
Maxent模型的复杂度对模型的转移能力影响较大, 基于默认参数和优化参数后模型的预测结果差异显著(图3 )。在本研究中, 经过调整参数后的Maxent模型, 其响应曲线较为平滑, 能够较为实际地反映物种对环境因子的响应。相反采用默认参数所构建的模型, 其响应曲线曲折较多。简单模型和复杂模型各有利弊, 简单的模型对模型构建区物种分布数据拟合效果较差, 而复杂的模型虽能较好地拟合构建区物种分布数据, 但其所反映出的物种和环境变量之间的响应关系偏离实际(乔慧捷等, 2013 )。在以模拟潜在分布为目的的生态位模型应用中, Maxent模型复杂度对预测结果的不利影响更为严重(Warren & Seifert, 2011 ; Warren et al, 2014 ), 这是因为模型构建后需要将其转移投射至其他地理空间来预测该物种的分布。复杂的模型由于其过度拟合模型构建区的物种分布数据, 导致模型转移至其他地理空间后预测能力降低, 所模拟的结果不可靠或与实际偏差较大。因此在构建模型之前, 建议对物种分布数据和环境变量进行预处理; 在构建Maxent生态位模型时, 建议采用ENMeval数据包等方法, 对Maxent模型的复杂度进行评估, 选择复杂度较低的模型来预测物种的潜在分布, 同时采用空间分割的方法对模型构建区的拟合度进行评估。
致谢: 感谢美国Kansas大学A. Townsend Peterson教授和Jorge Soberón教授在生态位模型学习中所给予的指导和帮助。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ahmed SE McInerny G O’Hara K Harper R Salido L Emmott S Joppa LN 2015 ) Scientists and software - surveying the species distribution modelling community
. Diversity and Distributions , 21 , 258 -267 .
[本文引用: 1]
[2]
Akaike H 1973 ) Information theory and an extension of the maximum likelihood principle. In: 2nd International Symposium on Information Theory (eds Petrov BN, Csáki F) , pp. 267 -281. Akadémiai Kiadó, Budapest.
[本文引用: 1]
[3]
Barbosa FG Schneck F 2015 ) Characteristics of the top-cited papers in species distribution predictive models
. Ecological Modelling , 313 , 77 -83 .
[本文引用: 1]
[4]
Elith J Phillips SJ Hastie T Dudík D Chee YE Yates CJ 2010 ) A statistical explanation of MaxEnt for ecologists
. Diversity and Distributions , 17 , 43 -57 .
[本文引用: 1]
[5]
Hijmans RJ Cameron SE Parra JL Jones PG Jarvis A 2005 ) Very high resolution interpolated climate surfaces for global land areas
. International Journal of Climatology , 25 , 1965 -1978 .
[本文引用: 1]
[6]
Jiménez-Valverde A Peterson AT Soberón J Overton JM Aragón P Lobo JM 2011 ) Use of niche models in invasive species risk assessments
. Biological Invasions , 13 , 2785 -2797 .
[本文引用: 5]
[7]
Kearney MR Wintle BA Porter WP 2010 ) Correlative and mechanistic models of species distribution provide congruent forecasts under climate change
. Conservation Letters , 3 , 203 -213 .
[本文引用: 1]
[8]
Kriticos DJ Webber BL Leriche A Ota N Macadam I Bathols J Scott JK 2011 ) CliMond: global high resolution historical and future scenario climate surfaces for bioclimatic modeling
. Methods in Ecology and Evolution , 3 , 53 -64 .
[本文引用: 1]
[9]
Muscarella R Galante PJ Soley-Guardia M Boria RA Kass JM Uriarte M Anderson RP 2014 ) ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models
. Methods in Ecology and Evolution , 5 , 1198 -1205 .
[本文引用: 3]
[10]
Peterson AT Papeş M Soberón J 2008 ) Rethinking receiver operating characteristic analysis applications in ecological niche modeling
. Ecological Modelling , 213 , 63 -72 .
[本文引用: 2]
[11]
Peterson AT Soberón J 2012 ) Species distribution modeling and ecological niche modeling: getting the concepts right
. Natureza & Conservacao , 10 , 102 -107 .
[本文引用: 1]
[12]
Peterson AT Soberón J Pearson RG Anderson RP Nakamura M Martínez-Meyer E Araújo MB 2011 ) Ecological Niches and Geographical Distributions . Princeton University Press , New Jersey .
[本文引用: 3]
[13]
Phillips SJ Anderson RP Schapire RE 2006 ) Maximum entropy modeling of species geographic distributions
. Ecological Modelling , 190 , 231 -259 .
[本文引用: 1]
[14]
Phillips SJ Dudík MM 2008 ) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation
. Ecography , 31 , 161 -175 .
[本文引用: 4]
[15]
Qiao HJ Hu JH Huang JH 2013 ) Theoretical basis, future directions, and challenges for ecological niche models
. Scientia Sinica Vitae , 43 , 915 -927 . (in Chinese with English abstract)
[本文引用: 3]
[乔慧捷 , 胡军华 , 黄继红 (2013 ) 生态位模型的理论基础、发展方向与挑战
. 中国科学: 生命科学 , 43 , 915 -927 .]
[本文引用: 3]
[16]
Qiao HJ Soberón J Peterson AT 2015 ) No silver bullets in correlative ecological niche modeling: insights from testing among many potential algorithms for niche estimation
. Methods in Ecology and Evolution , 6 , 1126 -1136 .
[本文引用: 1]
[17]
Soberón J Peterson AT 2005 ) Interpretation of models of fundamental ecological niches and species’ distributional areas
. Biodiversity Informatics , 2 , 1 -10 .
[本文引用: 1]
[18]
Vaz UL Cunha HF Nabout JC 2015 ) Trends and biases in global scientific literature about ecological niche models
. Brazilian Journal of Biology , 75 , 17 -24 .
[本文引用: 1]
[19]
Warren DL Seifert SN 2011 ) Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria
. Ecology Applications , 21 , 335 -342 .
[本文引用: 3]
[20]
Warren DL Wright AN Seifert SN Shaffer HB 2014 ) Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern
. Diversity and Distributions , 20 , 334 -343 .
[本文引用: 2]
[21]
Zhu GP Bu WJ Gao YB Liu GQ 2012 ) Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys )
. PLoS ONE , 7 , e31246 .
[22]
Zhu GP Gao YB Zhu L 2013 ) Delimiting the coastal geographic background to predict potential distribution of Spartina alterniflora
. Hydrobiologia , 717 , 177 -187 .
[本文引用: 2]
[23]
Zhu GP Redei D Kment P Bu WJ 2014 ) Effect of geographic background and equilibrium state on niche model transferability: predicting areas of invasion of Leptoglossus occidentalis
. Biological Invasions , 16 , 1069 -1081 .
[本文引用: 1]
[24]
Zhu GP Gariepy TD Haye T Bu WJ 2016 ) Patterns of niche filling and expansion across the invaded ranges of Halyomorpha halys in North America and Europe
. Journal of Pest Science , doi:10.1007/s10340-016-0786-z .
[本文引用: 1]
[25]
Zhu GP Liu GQ Bu WJ Gao YB 2013 ) Ecological niche modeling and its applications in biodiversity conservation
. Biodiversity Science , 21 , 90 -98 . (in Chinese with English abstract)
[本文引用: 2]
[朱耿平 , 刘国卿 , 卜文俊 , 高玉葆 (2013 ) 生态位模型的基本原理及其在生物多样性保护中的应用
. 生物多样性 , 21 , 90 -98 .]
[本文引用: 2]
[26]
Zhu GP Liu Q Gao YB 2014 ) Improving ecological niche model transferability to predict the potential distribution of invasive exotic species
. Biodiversity Science , 22 , 223 -230 . (in Chinese with English abstract)
[本文引用: 2]
[朱耿平 , 刘强 , 高玉葆 (2014 ) 提高生态位模型转移能力来模拟入侵物种的潜在分布
. 生物多样性 , 22 , 223 -230 .]
[本文引用: 2]
Scientists and software - surveying the species distribution modelling community
1
2015
... 生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用.前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
1
1973
... 首先采用Maxent默认参数在茶翅蝽本土地区构建模型, 然后将其转移至入侵地来检验和评价模型; 随后通过调用ENMeval数据包来调整Maxent模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数, 分析各种参数条件下模型的复杂度, 选取最低复杂度的模型参数; 最后综合比较分析Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响.Maxent模型的复杂度与其RM和FC参数密切相关.目前Maxent中有5种特征, 即: 线性(linear - L), 二次型(quadratic - Q), 片段化(hinge - H), 乘积型(product - P)和阈值性(threshold - T).在其默认设置中, RM的值为1, 具体特征组合的选择和使用与物种分布点数量有关, 通常情况下linear feature一直在运行, quadratic feature在物种分布点>10时使用, hinge feature在分布点>15时使用, threshold和product feature在分布点>80时使用(Elith et al, 2010 ).ENMeval数据包通过测试不同参数条件下Maxent模型修正的AIC值(即AICc)来评价模型的复杂度.AIC信息量准则(Akaike information criterion correction, AIC)是衡量统计模型拟合优良性的一种标准, 可以权衡所估计模型的复杂度和此模型拟合数据的优良性, AIC信息量准则优先考虑AIC值最小的模型(Akaike, 1973 ).AIC值可通过Maxent模型运行后产生的lambdas文件计算获得(Warren & Seifert, 2011 ).在本研究中, 我们将RM设置为0.5-4, 每次增加0.5, 采用6个特征组合(FC), 即: L, LQ, H, LQH, LQHP和LQHPT.为测试Maxent模型对本土分布数据拟合的效果, 在本土区域将物种分布点进行空间分割成4等份, 随机采用其中的3份构建模型, 剩余的1份用于检验模型(Muscarella et al, 2014 ). ...
Characteristics of the top-cited papers in species distribution predictive models
1
2015
... 生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用.前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
A statistical explanation of MaxEnt for ecologists
1
2010
... 首先采用Maxent默认参数在茶翅蝽本土地区构建模型, 然后将其转移至入侵地来检验和评价模型; 随后通过调用ENMeval数据包来调整Maxent模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数, 分析各种参数条件下模型的复杂度, 选取最低复杂度的模型参数; 最后综合比较分析Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响.Maxent模型的复杂度与其RM和FC参数密切相关.目前Maxent中有5种特征, 即: 线性(linear - L), 二次型(quadratic - Q), 片段化(hinge - H), 乘积型(product - P)和阈值性(threshold - T).在其默认设置中, RM的值为1, 具体特征组合的选择和使用与物种分布点数量有关, 通常情况下linear feature一直在运行, quadratic feature在物种分布点>10时使用, hinge feature在分布点>15时使用, threshold和product feature在分布点>80时使用(Elith et al, 2010 ).ENMeval数据包通过测试不同参数条件下Maxent模型修正的AIC值(即AICc)来评价模型的复杂度.AIC信息量准则(Akaike information criterion correction, AIC)是衡量统计模型拟合优良性的一种标准, 可以权衡所估计模型的复杂度和此模型拟合数据的优良性, AIC信息量准则优先考虑AIC值最小的模型(Akaike, 1973 ).AIC值可通过Maxent模型运行后产生的lambdas文件计算获得(Warren & Seifert, 2011 ).在本研究中, 我们将RM设置为0.5-4, 每次增加0.5, 采用6个特征组合(FC), 即: L, LQ, H, LQH, LQHP和LQHPT.为测试Maxent模型对本土分布数据拟合的效果, 在本土区域将物种分布点进行空间分割成4等份, 随机采用其中的3份构建模型, 剩余的1份用于检验模型(Muscarella et al, 2014 ). ...
Very high resolution interpolated climate surfaces for global land areas
1
2005
... 茶翅蝽原产于亚洲东部(中国、日本、韩国和朝鲜), 现已在北美洲和欧洲建立种群, 危害比较严重.物种分布点数据和环境变量源自Zhu等(2012, 2016)中, 已去除采样偏差对模型的影响.环境变量的选取主要考虑其对物种分布的限制作用和变量间的空间相关性(Peterson et al, 2011 ), 选取了年平均气温(bio1)、最热月份最高气温(bio5)、最冷月份最低气温(bio6)、年降雨量(bio12)和年平均辐射量(bio20)等5个环境变量(Hijmans et al, 2005 ; Kriticos et al, 2011 ).研究区域分为茶翅蝽的本土种群生存地区以及入侵地, 其中本土地区包括中国、朝鲜、韩国和日本, 入侵地以北美洲为例(Zhu et al, 2016 ). ...
Use of niche models in invasive species risk assessments
5
2011
... 生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用.前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
... ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
... ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
... Maxent默认参数的设置源自早期模型开发者对6个不同地理区域的266个物种数据的测试(Phillips & Dudík, 2008 ), 他们采用海量物种的分布数据和多种实验方案, 以期得到一个最优的模型参数作为默认参数来推广和简化Maxent模型的应用, 这些物种涵盖了鸟类、哺乳类、爬行类以及植物等类群, 分布在澳洲、新西兰、欧洲和南美洲等地区.然而, 这些模型的应用均是以预测现实分布为目的, 模型构建后不需转移, 预测模型需要能够很好地辨别物种的分布与否(即区分物种的分布和不分布, Jiménez-Valverde et al, 2008 ; Phillips & Dudík, 2008 ).在生物入侵、全球变化对物种分布影响和谱系生物地理学中, 生态位模型的应用是以模拟物种的潜在分布为目的, Maxent模型构建以后需要转移至不同地理空间中去预测物种的潜在分布, 需要降低模型预测的遗漏率.虽然Maxent模型所模拟的物种分布介于潜在分布和现实分布之间(Jiménez- Valverde et al, 2011 ), 但从研究者目的出发有时需要调整模拟方案来实现模拟需求.在以模拟物种潜在分布为目的时, Maxent模型默认参数可能不再适用, 如本研究中, 茶翅蝽的最优模型参数不同于默认参数(图1 ), 如果继续采用默认参数, Maxent模型虽能够很好地拟合模型构建区的物种分布, 但由于过度拟合会造成模型转移能力降低(图4 ).此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 ). ...
... ).在生物入侵、全球变化对物种分布影响和谱系生物地理学中, 生态位模型的应用是以模拟物种的潜在分布为目的, Maxent模型构建以后需要转移至不同地理空间中去预测物种的潜在分布, 需要降低模型预测的遗漏率.虽然Maxent模型所模拟的物种分布介于潜在分布和现实分布之间(Jiménez- Valverde et al, 2011 ), 但从研究者目的出发有时需要调整模拟方案来实现模拟需求.在以模拟物种潜在分布为目的时, Maxent模型默认参数可能不再适用, 如本研究中, 茶翅蝽的最优模型参数不同于默认参数(图1 ), 如果继续采用默认参数, Maxent模型虽能够很好地拟合模型构建区的物种分布, 但由于过度拟合会造成模型转移能力降低(图4 ).此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 ). ...
Correlative and mechanistic models of species distribution provide congruent forecasts under climate change
1
2010
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
CliMond: global high resolution historical and future scenario climate surfaces for bioclimatic modeling
1
2011
... 茶翅蝽原产于亚洲东部(中国、日本、韩国和朝鲜), 现已在北美洲和欧洲建立种群, 危害比较严重.物种分布点数据和环境变量源自Zhu等(2012, 2016)中, 已去除采样偏差对模型的影响.环境变量的选取主要考虑其对物种分布的限制作用和变量间的空间相关性(Peterson et al, 2011 ), 选取了年平均气温(bio1)、最热月份最高气温(bio5)、最冷月份最低气温(bio6)、年降雨量(bio12)和年平均辐射量(bio20)等5个环境变量(Hijmans et al, 2005 ; Kriticos et al, 2011 ).研究区域分为茶翅蝽的本土种群生存地区以及入侵地, 其中本土地区包括中国、朝鲜、韩国和日本, 入侵地以北美洲为例(Zhu et al, 2016 ). ...
ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models
3
2014
... 在本研究中, 以入侵害虫茶翅蝽(Halyomorpha halys )为例, 以预测入侵物种的潜在分布为目的, 探讨Maxent模型的复杂度对模型转移能力的影响.本研究采用经典生态模型方案, 通过构建本土模型, 然后将其转移至入侵地来模拟茶翅蝽的潜在分布, 同时采用地理空间分层的方法测试Maxent模型对本土分布数据是否过度拟合.通过调用ENMeval数据包(Muscarella et al, 2014 )来调整Maxent模型参数和分析各种参数条件下的模型复杂度, 并选取最低复杂度的模型参数, 综合比较Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响以及Maxent模型构建时的注意事项.本文结合模型的构建材料及模型的构建区域全面阐述提高模型转移能力的策略, 以期对物种潜在分布进行合理的预测, 促进Maxent模型在我国的合理运用和发展. ...
... 首先采用Maxent默认参数在茶翅蝽本土地区构建模型, 然后将其转移至入侵地来检验和评价模型; 随后通过调用ENMeval数据包来调整Maxent模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数, 分析各种参数条件下模型的复杂度, 选取最低复杂度的模型参数; 最后综合比较分析Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响.Maxent模型的复杂度与其RM和FC参数密切相关.目前Maxent中有5种特征, 即: 线性(linear - L), 二次型(quadratic - Q), 片段化(hinge - H), 乘积型(product - P)和阈值性(threshold - T).在其默认设置中, RM的值为1, 具体特征组合的选择和使用与物种分布点数量有关, 通常情况下linear feature一直在运行, quadratic feature在物种分布点>10时使用, hinge feature在分布点>15时使用, threshold和product feature在分布点>80时使用(Elith et al, 2010 ).ENMeval数据包通过测试不同参数条件下Maxent模型修正的AIC值(即AICc)来评价模型的复杂度.AIC信息量准则(Akaike information criterion correction, AIC)是衡量统计模型拟合优良性的一种标准, 可以权衡所估计模型的复杂度和此模型拟合数据的优良性, AIC信息量准则优先考虑AIC值最小的模型(Akaike, 1973 ).AIC值可通过Maxent模型运行后产生的lambdas文件计算获得(Warren & Seifert, 2011 ).在本研究中, 我们将RM设置为0.5-4, 每次增加0.5, 采用6个特征组合(FC), 即: L, LQ, H, LQH, LQHP和LQHPT.为测试Maxent模型对本土分布数据拟合的效果, 在本土区域将物种分布点进行空间分割成4等份, 随机采用其中的3份构建模型, 剩余的1份用于检验模型(Muscarella et al, 2014 ). ...
... 在本研究中, 所有物种分布数据分成3份, 其中随机选取70%的本土物种分布点用于模型构建和优化, 剩下30%的本土物种分布点用于模型内部检验, 北美入侵地物种分布点用于检验模型转移能力.Maxent模型的对比和评价主要采用响应曲线和遗漏率曲线, 同时采用Partial ROC方案测试模型的本土预测能力和转移能力(Peterson et al, 2008 ).在ENMenv数据包中, 采用AUC来测试模型在本土4份地理空间之间的相互预测的效果, AUC.diff (等于AUCtrain-AUCtest)和OR10用来测试模型对本土物种分布点的拟合程度, delta.AICc用来测试模型的复杂度和拟合程度(详细参见Muscarella et al, 2014 ). ...
Rethinking receiver operating characteristic analysis applications in ecological niche modeling
2
2008
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... 在本研究中, 所有物种分布数据分成3份, 其中随机选取70%的本土物种分布点用于模型构建和优化, 剩下30%的本土物种分布点用于模型内部检验, 北美入侵地物种分布点用于检验模型转移能力.Maxent模型的对比和评价主要采用响应曲线和遗漏率曲线, 同时采用Partial ROC方案测试模型的本土预测能力和转移能力(Peterson et al, 2008 ).在ENMenv数据包中, 采用AUC来测试模型在本土4份地理空间之间的相互预测的效果, AUC.diff (等于AUCtrain-AUCtest)和OR10用来测试模型对本土物种分布点的拟合程度, delta.AICc用来测试模型的复杂度和拟合程度(详细参见Muscarella et al, 2014 ). ...
Species distribution modeling and ecological niche modeling: getting the concepts right
1
2012
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
3
2011
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
... 茶翅蝽原产于亚洲东部(中国、日本、韩国和朝鲜), 现已在北美洲和欧洲建立种群, 危害比较严重.物种分布点数据和环境变量源自Zhu等(2012, 2016)中, 已去除采样偏差对模型的影响.环境变量的选取主要考虑其对物种分布的限制作用和变量间的空间相关性(Peterson et al, 2011 ), 选取了年平均气温(bio1)、最热月份最高气温(bio5)、最冷月份最低气温(bio6)、年降雨量(bio12)和年平均辐射量(bio20)等5个环境变量(Hijmans et al, 2005 ; Kriticos et al, 2011 ).研究区域分为茶翅蝽的本土种群生存地区以及入侵地, 其中本土地区包括中国、朝鲜、韩国和日本, 入侵地以北美洲为例(Zhu et al, 2016 ). ...
... 相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 ).在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意.生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 ).在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差.环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
Maximum entropy modeling of species geographic distributions
1
2006
... 生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用.前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation
4
2008
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... Maxent默认参数的设置源自早期模型开发者对6个不同地理区域的266个物种数据的测试(Phillips & Dudík, 2008 ), 他们采用海量物种的分布数据和多种实验方案, 以期得到一个最优的模型参数作为默认参数来推广和简化Maxent模型的应用, 这些物种涵盖了鸟类、哺乳类、爬行类以及植物等类群, 分布在澳洲、新西兰、欧洲和南美洲等地区.然而, 这些模型的应用均是以预测现实分布为目的, 模型构建后不需转移, 预测模型需要能够很好地辨别物种的分布与否(即区分物种的分布和不分布, Jiménez-Valverde et al, 2008 ; Phillips & Dudík, 2008 ).在生物入侵、全球变化对物种分布影响和谱系生物地理学中, 生态位模型的应用是以模拟物种的潜在分布为目的, Maxent模型构建以后需要转移至不同地理空间中去预测物种的潜在分布, 需要降低模型预测的遗漏率.虽然Maxent模型所模拟的物种分布介于潜在分布和现实分布之间(Jiménez- Valverde et al, 2011 ), 但从研究者目的出发有时需要调整模拟方案来实现模拟需求.在以模拟物种潜在分布为目的时, Maxent模型默认参数可能不再适用, 如本研究中, 茶翅蝽的最优模型参数不同于默认参数(图1 ), 如果继续采用默认参数, Maxent模型虽能够很好地拟合模型构建区的物种分布, 但由于过度拟合会造成模型转移能力降低(图4 ).此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 ). ...
... ; Phillips & Dudík, 2008 ).在生物入侵、全球变化对物种分布影响和谱系生物地理学中, 生态位模型的应用是以模拟物种的潜在分布为目的, Maxent模型构建以后需要转移至不同地理空间中去预测物种的潜在分布, 需要降低模型预测的遗漏率.虽然Maxent模型所模拟的物种分布介于潜在分布和现实分布之间(Jiménez- Valverde et al, 2011 ), 但从研究者目的出发有时需要调整模拟方案来实现模拟需求.在以模拟物种潜在分布为目的时, Maxent模型默认参数可能不再适用, 如本研究中, 茶翅蝽的最优模型参数不同于默认参数(图1 ), 如果继续采用默认参数, Maxent模型虽能够很好地拟合模型构建区的物种分布, 但由于过度拟合会造成模型转移能力降低(图4 ).此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 ). ...
... ).此外, Maxent模型默认参数的测试数据中所采用的物种数据和地理区域亦具有局限性, 如所测试数据未包括昆虫类群, 地理区域亦未包括亚洲和北美洲等地区(Phillips & Dudík, 2008 ). ...
生态位模型的理论基础、发展方向与挑战
3
2013
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... Maxent模型的复杂度对模型的转移能力影响较大, 基于默认参数和优化参数后模型的预测结果差异显著(图3 ).在本研究中, 经过调整参数后的Maxent模型, 其响应曲线较为平滑, 能够较为实际地反映物种对环境因子的响应.相反采用默认参数所构建的模型, 其响应曲线曲折较多.简单模型和复杂模型各有利弊, 简单的模型对模型构建区物种分布数据拟合效果较差, 而复杂的模型虽能较好地拟合构建区物种分布数据, 但其所反映出的物种和环境变量之间的响应关系偏离实际(乔慧捷等, 2013 ).在以模拟潜在分布为目的的生态位模型应用中, Maxent模型复杂度对预测结果的不利影响更为严重(Warren & Seifert, 2011 ; Warren et al, 2014 ), 这是因为模型构建后需要将其转移投射至其他地理空间来预测该物种的分布.复杂的模型由于其过度拟合模型构建区的物种分布数据, 导致模型转移至其他地理空间后预测能力降低, 所模拟的结果不可靠或与实际偏差较大.因此在构建模型之前, 建议对物种分布数据和环境变量进行预处理; 在构建Maxent生态位模型时, 建议采用ENMeval数据包等方法, 对Maxent模型的复杂度进行评估, 选择复杂度较低的模型来预测物种的潜在分布, 同时采用空间分割的方法对模型构建区的拟合度进行评估. ...
生态位模型的理论基础、发展方向与挑战
3
2013
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... Maxent模型的复杂度对模型的转移能力影响较大, 基于默认参数和优化参数后模型的预测结果差异显著(图3 ).在本研究中, 经过调整参数后的Maxent模型, 其响应曲线较为平滑, 能够较为实际地反映物种对环境因子的响应.相反采用默认参数所构建的模型, 其响应曲线曲折较多.简单模型和复杂模型各有利弊, 简单的模型对模型构建区物种分布数据拟合效果较差, 而复杂的模型虽能较好地拟合构建区物种分布数据, 但其所反映出的物种和环境变量之间的响应关系偏离实际(乔慧捷等, 2013 ).在以模拟潜在分布为目的的生态位模型应用中, Maxent模型复杂度对预测结果的不利影响更为严重(Warren & Seifert, 2011 ; Warren et al, 2014 ), 这是因为模型构建后需要将其转移投射至其他地理空间来预测该物种的分布.复杂的模型由于其过度拟合模型构建区的物种分布数据, 导致模型转移至其他地理空间后预测能力降低, 所模拟的结果不可靠或与实际偏差较大.因此在构建模型之前, 建议对物种分布数据和环境变量进行预处理; 在构建Maxent生态位模型时, 建议采用ENMeval数据包等方法, 对Maxent模型的复杂度进行评估, 选择复杂度较低的模型来预测物种的潜在分布, 同时采用空间分割的方法对模型构建区的拟合度进行评估. ...
No silver bullets in correlative ecological niche modeling: insights from testing among many potential algorithms for niche estimation
1
2015
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
Interpretation of models of fundamental ecological niches and species’ distributional areas
1
2005
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
Trends and biases in global scientific literature about ecological niche models
1
2015
... 生态位模型所模拟的物种分布朝着两个方向, 即现实分布和潜在分布, 它们在不同领域分别有着广泛的应用.前者以模拟物种现实分布为目的, 模型构建后不需要转移, 主要应用于保护区的界定和濒危物种迁地保护研究中; 后者以模拟物种的潜在分布为目的, 模型构建后需要转移, 这种转移以生态位的保守性为基础, 主要应用于入侵生物学、全球变化对物种分布的影响、以及谱系生物地理学中(Jiménez-Valverde et al, 2011 ).在以现实分布为目的时, 生态位模型有时也被称为物种分布模型.目前常用的生态位模型有约20种, 各自有不同的理论基础、分析方式和数据需求, 这些模型所模拟的物种分布分别处于现实分布和潜在分布之间.一般认为同时采用物种存在和不存在分布数据的模型所模拟的物种分布倾向于反映物种的现实分布, 仅采用物种存在分布点的模型所模拟的物种分布倾向于反映物种的潜在分布(Jiménez-Valverde et al, 2011 ).Maxent模型是目前使用最为广泛的生态位模型(Ahmed et al, 2015 ; Barbosa & Schneck, 2015 ; Vaz et al, 2015 ), 它以概率论和机器学习理论为基础(Phillips et al, 2006 ), 采用物种存在分布点和背景环境变量来构建模型, 所模拟的物种分布介于潜在分布和现实分布之间(Jiménez-Valverde et al, 2011 ). ...
Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria
3
2011
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... 首先采用Maxent默认参数在茶翅蝽本土地区构建模型, 然后将其转移至入侵地来检验和评价模型; 随后通过调用ENMeval数据包来调整Maxent模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数, 分析各种参数条件下模型的复杂度, 选取最低复杂度的模型参数; 最后综合比较分析Maxent模型默认参数和调整参数后的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响.Maxent模型的复杂度与其RM和FC参数密切相关.目前Maxent中有5种特征, 即: 线性(linear - L), 二次型(quadratic - Q), 片段化(hinge - H), 乘积型(product - P)和阈值性(threshold - T).在其默认设置中, RM的值为1, 具体特征组合的选择和使用与物种分布点数量有关, 通常情况下linear feature一直在运行, quadratic feature在物种分布点>10时使用, hinge feature在分布点>15时使用, threshold和product feature在分布点>80时使用(Elith et al, 2010 ).ENMeval数据包通过测试不同参数条件下Maxent模型修正的AIC值(即AICc)来评价模型的复杂度.AIC信息量准则(Akaike information criterion correction, AIC)是衡量统计模型拟合优良性的一种标准, 可以权衡所估计模型的复杂度和此模型拟合数据的优良性, AIC信息量准则优先考虑AIC值最小的模型(Akaike, 1973 ).AIC值可通过Maxent模型运行后产生的lambdas文件计算获得(Warren & Seifert, 2011 ).在本研究中, 我们将RM设置为0.5-4, 每次增加0.5, 采用6个特征组合(FC), 即: L, LQ, H, LQH, LQHP和LQHPT.为测试Maxent模型对本土分布数据拟合的效果, 在本土区域将物种分布点进行空间分割成4等份, 随机采用其中的3份构建模型, 剩余的1份用于检验模型(Muscarella et al, 2014 ). ...
... Maxent模型的复杂度对模型的转移能力影响较大, 基于默认参数和优化参数后模型的预测结果差异显著(图3 ).在本研究中, 经过调整参数后的Maxent模型, 其响应曲线较为平滑, 能够较为实际地反映物种对环境因子的响应.相反采用默认参数所构建的模型, 其响应曲线曲折较多.简单模型和复杂模型各有利弊, 简单的模型对模型构建区物种分布数据拟合效果较差, 而复杂的模型虽能较好地拟合构建区物种分布数据, 但其所反映出的物种和环境变量之间的响应关系偏离实际(乔慧捷等, 2013 ).在以模拟潜在分布为目的的生态位模型应用中, Maxent模型复杂度对预测结果的不利影响更为严重(Warren & Seifert, 2011 ; Warren et al, 2014 ), 这是因为模型构建后需要将其转移投射至其他地理空间来预测该物种的分布.复杂的模型由于其过度拟合模型构建区的物种分布数据, 导致模型转移至其他地理空间后预测能力降低, 所模拟的结果不可靠或与实际偏差较大.因此在构建模型之前, 建议对物种分布数据和环境变量进行预处理; 在构建Maxent生态位模型时, 建议采用ENMeval数据包等方法, 对Maxent模型的复杂度进行评估, 选择复杂度较低的模型来预测物种的潜在分布, 同时采用空间分割的方法对模型构建区的拟合度进行评估. ...
Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern
2
2014
... 在生态位模型中, 简单模型准确率常较低、可转移性高; 而复杂模型恰好相反, 准确率高、可转移性差(Qiao et al, 2015 ).如何在模型的可转移性和准确率之间找到平衡点, 是生态位模型的一个重要研究方向(乔慧捷等, 2013 ).Maxent模型属于机械学习式的复杂模型, 多数研究者采用Maxent默认参数来构建模型, 这些默认参数的设置源自早期Maxent模型开发者对不同地理区域的266个物种数据的测试(主要是鸟类、哺乳类、爬行类以及植物), 所测试物种以模拟其现实分布为目的(Phillips & Dudík, 2008 ).随后的研究发现, 在以模拟物种潜在分布为目的时, Maxent模型对采样偏差敏感, 容易产生过度拟合(overfitting)的问题, 从而影响模型的转移能力, 模型转移能力仅在低阈值情况下较好(Peterson et al, 2008 ; Warren & Seifert, 2011 ; Warren et al, 2014 ).当以模拟现实分布为目的时, Maxent模型的这种特性对预测结果影响不明显; 但当以模拟物种潜在分布为目的时, 由于Maxent模型容易过度拟合导致模型转移能力较低, 严重影响了其在入侵生物学和全球变化生物学等研究中的应用, 这种模拟不仅预测结果不可靠, 而且预测结果很难解释. ...
... Maxent模型的复杂度对模型的转移能力影响较大, 基于默认参数和优化参数后模型的预测结果差异显著(图3 ).在本研究中, 经过调整参数后的Maxent模型, 其响应曲线较为平滑, 能够较为实际地反映物种对环境因子的响应.相反采用默认参数所构建的模型, 其响应曲线曲折较多.简单模型和复杂模型各有利弊, 简单的模型对模型构建区物种分布数据拟合效果较差, 而复杂的模型虽能较好地拟合构建区物种分布数据, 但其所反映出的物种和环境变量之间的响应关系偏离实际(乔慧捷等, 2013 ).在以模拟潜在分布为目的的生态位模型应用中, Maxent模型复杂度对预测结果的不利影响更为严重(Warren & Seifert, 2011 ; Warren et al, 2014 ), 这是因为模型构建后需要将其转移投射至其他地理空间来预测该物种的分布.复杂的模型由于其过度拟合模型构建区的物种分布数据, 导致模型转移至其他地理空间后预测能力降低, 所模拟的结果不可靠或与实际偏差较大.因此在构建模型之前, 建议对物种分布数据和环境变量进行预处理; 在构建Maxent生态位模型时, 建议采用ENMeval数据包等方法, 对Maxent模型的复杂度进行评估, 选择复杂度较低的模型来预测物种的潜在分布, 同时采用空间分割的方法对模型构建区的拟合度进行评估. ...
Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys )
2012
Delimiting the coastal geographic background to predict potential distribution of Spartina alterniflora
2
2013
... 相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 ).在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意.生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 ).在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差.环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
... ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
Effect of geographic background and equilibrium state on niche model transferability: predicting areas of invasion of Leptoglossus occidentalis
1
2014
... 相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 ).在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意.生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 ).在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差.环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
Patterns of niche filling and expansion across the invaded ranges of Halyomorpha halys in North America and Europe
1
2016
... 茶翅蝽原产于亚洲东部(中国、日本、韩国和朝鲜), 现已在北美洲和欧洲建立种群, 危害比较严重.物种分布点数据和环境变量源自Zhu等(2012, 2016)中, 已去除采样偏差对模型的影响.环境变量的选取主要考虑其对物种分布的限制作用和变量间的空间相关性(Peterson et al, 2011 ), 选取了年平均气温(bio1)、最热月份最高气温(bio5)、最冷月份最低气温(bio6)、年降雨量(bio12)和年平均辐射量(bio20)等5个环境变量(Hijmans et al, 2005 ; Kriticos et al, 2011 ).研究区域分为茶翅蝽的本土种群生存地区以及入侵地, 其中本土地区包括中国、朝鲜、韩国和日本, 入侵地以北美洲为例(Zhu et al, 2016 ). ...
生态位模型的基本原理及其在生物多样性保护中的应用
2
2013
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
... ; 朱耿平等, 2013 ). ...
生态位模型的基本原理及其在生物多样性保护中的应用
2
2013
... 近年来, 生态位模型在生物多样性保护的多个领域得到应用, 如入侵生物学、保护生物学、全球气候变化对物种分布的影响、谱系生物地理学及传染病空间传播研究(Peterson et al, 2011 ; 朱耿平等, 2013 ).一般来说, 生态位模型可分为实验机理性方案(mechanistic approach)模型和相关性方案(correlative approach)模型两种(Soberón & Peterson, 2005 ; Kearney et al, 2010 ).实验机理性方案指的是通过实验测试物种的生理耐受性来推断物种在地理空间中的分布; 相关性方案生态位模型将地理空间和生态空间联系起来, 利用物种已知的分布数据和相关环境变量, 根据一定的算法来构建模型, 判断物种的生态需求, 并将运算结果投射到不同的时间或地理空间中来预测物种的现实分布(realized distribution)和潜在分布(potential distribution).与实验机理性方案生态位模型相比, 相关性方案的生态位模型具有开放性、构建相对简单、所需参数较少等特点, 从而被越来越多的学者所采用.与分子系统学的发展历程不同, 生态位模型的广泛应用推动着其理论基础不断向前发展, 其中生态位概念、生态位与物种分布的关系、生态位模型与生态位的关系等理论的丰富和发展是构建合理模型的重要基础(Peterson & Soberón, 2012 ; 乔慧捷等, 2013 ; 朱耿平等, 2013 ). ...
... ; 朱耿平等, 2013 ). ...
提高生态位模型转移能力来模拟入侵物种的潜在分布
2
2014
... 生态位模型是利用物种已知分布点所关联的环境变量去推算物种的生态需求, 模拟物种的分布.在模拟入侵物种分布时, 经典生态位模型通过模型构建物种本土分布地模型, 然后将其转移并投射至另一地理区域, 来模拟入侵物种的潜在分布.然而在模型运用时, 出现了模型转移能力较低、模拟结果与物种的现实分布不相符的情况, 由此得出了生态位漂移等不恰当的结论.提高生态位模型的转移能力, 不仅可以准确地模拟物种的潜在分布, 同时对生态位保守性等理论问题具有重要的参考价值(朱耿平等, 2014 ). ...
... 相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 ).在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意.生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 ).在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差.环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
提高生态位模型转移能力来模拟入侵物种的潜在分布
2
2014
... 生态位模型是利用物种已知分布点所关联的环境变量去推算物种的生态需求, 模拟物种的分布.在模拟入侵物种分布时, 经典生态位模型通过模型构建物种本土分布地模型, 然后将其转移并投射至另一地理区域, 来模拟入侵物种的潜在分布.然而在模型运用时, 出现了模型转移能力较低、模拟结果与物种的现实分布不相符的情况, 由此得出了生态位漂移等不恰当的结论.提高生态位模型的转移能力, 不仅可以准确地模拟物种的潜在分布, 同时对生态位保守性等理论问题具有重要的参考价值(朱耿平等, 2014 ). ...
... 相关性方案生态位模型所需设置的参数较少, 构建材料比较简单(即物种的分布点和环境变量), 随着全球物种分布数据的共享和GIS技术的快速发展, 这些数据的获取变得相对容易, 但构建一个合理的模型还需要充分了解所模拟物种的生物学特性、种群平衡状态、本土地理分布范围及种群历史等方面的知识(朱耿平等, 2014 ).在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意.生态位的概念、生态位与物种分布的关系等理论是构建合理模型的重要理论基础(Peterson et al, 2011 ).在准备模型构建材料时, 一般认为物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差.环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度(Zhu et al, 2013 ); 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态(Zhu et al, 2013 , 2014 ). ...
 , 乔慧捷
, 乔慧捷

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}