1 Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 1008712 Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008
通讯作者: * 通讯作者 Author for correspondence. E-mail: jshe@pku.edu.cn
As global biodiversity losses accelerate, there is increasing evidence shows that there may be negative impacts on ecosystem functioning, such as declines in plant primary productivity and imbalances in nutrient cycling. Thus, it is critical to understand the relationship between biodiversity and ecosystem functioning (BEF). However, ecosystems can provide multiple functions simultaneously (ecosystem multifunctionality, EMF). Since 2007, the quantification of relationships between biodiversity and ecosystem multifunctionality (BEMF) has generated additional questions and controversies, such as the lack of consensus in appropriate multifunctionality indices and uncertain trade-offs among ecosystem functions. In this review, we briefly summarize the history of BEMF studies and the methods of EMF quantification, then outline the mechanisms of EMF maintenance and current research progress. We emphasize the importance of optimizing EMF quantifications and investigating the relationship between different dimensions of biodiversity and EMF. We also provide suggestions and directions for future research on BEMF.
Keywords:ecosystem functioning
;
species loss
;
multifunctionality indices
;
multiple functions
Table 1 Key references on biodiversity and ecosystem multifunctionality (BEMF)
序号 No.
文献 References
备注 Remarks
1
Sanderson et al, 2004
首次提出多功能性的概念。This paper firstly defined multifunctionality.
2
Hector & Bagchi, 2007
第一次定量分析了生物多样性同时对多个生态系统过程的作用, 引发BEMF研究热潮。This paper first quantified the effects of biodiversity on multiple ecosystem processes, and evoked research climax of BEMF.
3
Gamfeldt et al, 2008
提出一个概念模型来探讨物种丧失对生态系统多个功能的影响; 证明多功能冗余比单功能冗余程度低。This paper proposed a conceptual model to address how species loss affects overall ecosystem functioning and demonstrated that multifunctional redundancy was generally lower than single functional redundancy.
4
Zavaleta et al, 2010
第一次在多个时间尺度上探讨生物多样性与生态系统多功能性的关系。This paper studied the relationships between biodiversity and ecosystem multifunctionality at multiple temporal scales.
5
Isbell et al, 2011
评估生物多样性与生态系统多功能性的关系时, 第一次同时考虑了不同的时间、地点、功能和气候变化等因素。This study first considered multiple times, places, functions or climate change scenarios in the evaluations of the relationships between biodiversity and ecosystem services.
6
Maestre et al, 2012b
第一次研究全球干旱地的植物物种丰富度和生态系统多功能性的关系。This paper first investigated the relationships between plant species richness and ecosystem multifunctionality in drylands at global scale.
7
Maestre et al, 2012a
第一次通过实验探讨关键群落属性(物种丰富度、群落物种组成、均匀度和空间格局)同时改变对生态系统多功能性的影响。This paper first investigated the effects of simultaneously changes in key community traits (e.g., species richness, community components, evenness, and spatial patterns) on ecosystem multifucatioality.
8
Pasari et al, 2013
第一次研究了多个尺度的生物多样性对生态系统多功能性的影响。This paper first studied the relationships between several scales of BEMF.
9
Byrnes et al, 2014a
利用BIODEPTH实验数据和R软件的multifunc程序包系统综述了当前BEMF研究中存在的问题, 首次提出多阈值法。This paper systematically reviewed the challenges and solutions in current BEMF studies and first quantified EMF with multiple threshold approach using data from the BIODEPTH experiment and the multifunc packages in R.
10
Wagg et al, 2014
通过改变土壤群落多样性实验, 提出土壤生物多样性和土壤群落组成决定生态系统多功能性。This study demonstrated that soil biodiversity and soil community composition determine ecosystem multifunctionality by manipulating soil community biodiversity.
11
Perkins et al, 2015
第一次研究了变化环境下生物多样性与生态系统多功能性的关系。This paper first studied the relationships between biodiversity and ecosystem multifunctionality in a changing environmental scenario.
12
Valencia et al, 2015
首次研究了功能多样性与生态系统多功能性的关系。This paper first studied the relationship between functional diversity and ecosystem multifunctionality.
13
Lefcheck et al, 2015
第一次系统分析了不同分类群、营养级和生境条件下生物多样性对生态系统多功能性的作用。This study presented the first systematic investigation of biodiversity’s effect on ecosystem multifunctionality across multiple taxonomy, trophic levels, and habitats.
图1 物种丰富度与生态系统功能的关系随冗余度的变化。 (a)物种丧失对假想群落的影响。实线表示当丧失的每个物种都是群落中存在的最有效率的物种时, 功能丧失最大化(物种丰富度由高转低时)。箭头和虚线表示随着冗余的变化, 物种丧失对生态系统功能影响的变化。(b)此图表示概念上维持整体功能的概率如何依赖于功能的数量和物种间多功能冗余的程度。随着某些功能的特化, 多个功能将比单个功能更易受到物种丧失的影响, 这种敏感性将随着冗余度的降低而增加(修改自Gamfeldt et al, 2008)。
Fig. 1 The relationship between species richness and ecosystem functioning changes with redundancy. (a) The effect of species loss on hypothetical communities. The solid line represents maximum loss in function if each species lost is the most efficient species of the ones remaining in the community (as we move from high to low species richness). The arrow and the dashed line indicate how the influence of species loss on ecosystem function shifts with changing redundancy. (b) A conceptual model of how the probability of maintaining overall functioning depends on both the number of functions and the degree of multifunctional redundancy across species. With some functional specialization, multiple functions will always be more susceptible to species loss than single functions. This susceptibility will increase with decreasing redundancy (Adopted from Gamfeldt et al, 2008).
Peter等(2011)通过控制生物膜上的微生物群落结构, 用分子指纹法来识别群落的组成, 以单阈值法量化多功能性, 发现微生物物种对环境的适应能力会影响多功能性冗余, 微生物群落多样性的减少对多功能性的影响比对单个功能的影响程度要大得多。可见, 微生物群落的多功能冗余比过去基于单个功能的实验和观测(Wohl et al, 2004)所得出的结果要低得多。
上述争论是针对多功能性指数的应用和开发进行的, 但关于土壤群落生物多样性的减少是否会对一个生态系统的综合表现造成影响仍然没有明确的答案。为此, Wagg等(2014)通过微宇宙实验, 在典型草原群落中建立了物种组成和多样性不同的土壤动物、微生物群落, 并检验它们对8个生态系统功能的作用。该研究以平均值法量化生态系统多功能性。实验结果表明, 生物多样性丧失和土壤群落组成的简单化会降低生态系统的多功能性, 并且随时间的推移会表现出越来越强的抑制作用; 所测得的生态系统多功能性与土壤生物多样性指数有很强的正向线性关系, 表明土壤群落组成是调节生态系统功能的关键因子。此外, 该实验还发现植物多样性随着土壤生物多样性的降低和土壤微生物群落的简单化而降低。而20年来的生物多样性研究已经证明, 在大部分生态系统中, 地上植物多样性是生态系统功能的关键驱动者(Tilman et al, 1996; Balvanera et al, 2006; Hector & Bagchi, 2007; Zavaleta et al, 2010), 因此, 土壤生物多样性下降和土壤群落组分的变化对维持生态系统多功能性的一系列生态系统过程均有影响。
van der Heijden等(2015)根据Hector和Bagchi (2007)的功能-物种替代法对早期实验数据进行分析发现: 丛枝菌根真菌和外生菌根真菌与多个生态系统功能如植物生产力、凋落物分解等相关, 且对很多生态系统过程(如植物对P的摄取)的贡献很大, 对某些过程的贡献甚至可达90%。因此, 菌根真菌的存在能显著提高生态系统的多功能性。此研究也验证了Wagg等(2014)的实验结果, 即土壤生物多样性与生态系统多功能性之间存在正相关关系。
Fig. 2 Structural equation models (SEM) of climate, soil and biodiversity as predictors of ecosystem multifuncitonality (EMF), represented by two hypotheses: (a) plant species richness drives belowground biodiversity (soil biodiversity), and conversely, (b) belowground biodiversity drives plant species richness. Solid black arrows represent positive paths (P < 0.05, piecewise SEM), solid gray arrows represent negative paths (P < 0.05, piecewise SEM), and dotted gray arrows represent non-significant paths (P > 0.05, piecewise SEM). We report the path coefficients as standardized effect sizes. Overall fit of piecewise SEM is evaluated using Shipley’s test of d-separation: Fisher’s C statistic (if P > 0.05, then no paths are missing and the model is a good fit) and Akaike Information Criterion (AIC). MAP: Mean annual precipitation; MAT: Mean annual temperature; SM: Soil moisture; Plant SR: Plant species richness (Adopted from Jing et al, 2015).
然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多。另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012)。当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性。功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系。例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015)。缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3)。希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性。
Fig. 3 The impacts of multiple biodiversity dimensionality on ecosystem multifunctionality under global change. The purple dashed box indicates current research focus while the red dashed box represents proposed future directions.
a) Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proceedings of the National Academy of Sciences,
Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences,
Domesticated nature: shaping landscapes and ecosystems for human welfare.
Science, 316, 1866-1869.
[54]
KinzigAP, PacalaSW, TilmanD (2001) The Functional Consequences of Biodiversity: Empirical Progress and Theoretical Extensions. Princeton University Press, Princeton.
Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: results from Inner Mongolia steppe ecosystem of north China.
Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences,
High productivity in grassland ecosystems: effected by species diversity or productive species.
1997
Quantifying the evidence for biodiversity effects on ecosystem functioning and services.
2
2006
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
Structural Equations with Latent Variables. John Wiley & Sons, New York.
1
2014
... 由于功能间的相关性除受多样性或物种组成的影响外, 还受微生物驱动的养分循环的影响, 所以有必要弄清各功能间是否具有相互作用(Byrnes et al, 2014a).常用的对单个功能的研究方法往往会忽视生态系统过程之间的权衡, 导致研究者对多样性如何影响生态系统功能的理解受到限制甚至产生偏见(Byrnes et al, 2014a).与单作的物种相比, 不同功能之间的权衡会导致生态系统提供多个功能的能力降低(Zavaleta et al, 2010; Gamfeldt et al, 2013), 这会使得多样性对多功能性的作用小于其对单个功能的作用.Byrnes等(2014a)认为, 可以用一些系统模型方法例如结构方程模型(structural equation modelling)来解决功能之间的相互作用问题; 为了得到完全正交的过程或将一系列的功能降低到几个综合成分, 可以通过验证性因子分析(confirmatory factor analysis) (Bollen, 2014)建立一个模型来分析观测的功能与背后隐藏的未测量的功能间有何关系, 用因子得分来代替所测量的功能进行后续分析, 或者也可以用主成分分析(principle components analysis)或其他降维的方法来处理. ...
Diversity and patch-size distributions of biological soil crusts regulate dryland ecosystem multifunctionality.
1
2013
... (3)对单个生态系统过程、功能或尺度分析的结果, 往往会低估生物多样性对生态系统功能的作用(Eisenhauer et al, 2012; Maestre et al, 2012b; Bowker et al, 2013; Pasari et al, 2013; Wagg et al, 2014; Lefcheck et al, 2015).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
Impacts of soil faunal community composition on model grassland ecosystems.
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
a) Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proceedings of the National Academy of Sciences,
2014
b) Reply to Byrnes et al.: aggregation can obscure understanding of ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
2014
a) Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions.
2014
b) Multifunctionality does not imply that all functions are positively correlated. Proceedings of the National Academy of Sciences,
2014
Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity.
1
2009
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
Biodiversity improves water quality through niche partitioning.
3
2011
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
... ).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
... 在过去10年间, 国内外已有很多关于BEF研究的综述文章(如刘峰等, 1999; Loreau, 2000; Loreau et al, 2001; 张全国和张大勇, 2003; Hooper et al, 2005; 江小雷等, 2010; Cardinale et al, 2012), Tilman等(2014)的综述再次对BEF过去的研究历程和未来的发展方向作了全面的展望. ...
The functional role of producer diversity in ecosystems.
2011
Effects of biodiversity on the functioning of trophic groups and ecosystems.
4
2006
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... ; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
... ).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences,
2
2007
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
... ), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
The standing crop of fish in lakes.
1
1955
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Exponential decline of deep-sea ecosystem functioning linked to benthic biodiversity loss.
2008
Soil invertebrate fauna enhances grassland succession and diversity.
1
2003
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Soil food web properties explain ecosystem services across European land use systems. Proceedings of the National Academy of Sciences,
1
2013
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
The plant traits that drive ecosystems: evidence from three continents.
... 有研究认为, 生物多样性与生态系统多功能性的关系至少受到两种重要权衡的影响(Gamfeldt et al, 2008): 一是由于某些功能间的排斥作用, 单一群落很难同时维持多个特定功能, 例如生产力和抗逆性常常是负相关的, 因此很难或不可能同时最大化(Grime, 1974); 二是不同的功能需要不同物种丰富度或组成的群落来使其最大化(Vinebrooke et al, 2004), 如不同乡土物种的组合能使其对入侵物种的抵抗力最大化(Zavaleta et al, 2010).Kareiva等(2007)指出, 重要的生态系统功能间常常存在权衡关系, 即一个功能(如生物量)的增加可能会伴随另一个功能(如对疾病的抵抗力)的降低.研究表明, 权衡作用限制了大多数物种组合的某些功能达到较高水平, 如高生产力与高抗旱性很难同时达到最高水平(Grime, 1974; Díaz et al, 2004).此外, 群落中可能有一些功能相似的物种, 它们能提供少数较高水平的功能; 或者具有功能多样性的物种, 它们能同时提供较高水平的不同功能(Zavaleta et al, 2010). ...
The statistical inevitability of stability-diversity relationships in community ecology.
1998
Testing the effects of diversity on ecosystem multi-functionality using a multivariate model.
1
2015
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
Why biodiversity is important to the functioning of real-world ecosystems.
1
2008
... 在一个生态系统中, 究竟需要多少个物种来维持其功能和服务, 研究者们仍然有不同的观点(Duffy, 2008; Ridder et al, 2008).如果仅考虑某个因素如不同时间、地点、功能或环境的差异, 则大多数物种都能提高生态系统功能, 但以前的研究没有同时考虑这些因子的时空变异(Isbell et al, 2011).此外, BEF实验常常在小尺度下, 通过测定有限数量的功能来研究生物多样性与生态系统功能的关系, 但是这与实际存在的生物多样性或生态系统功能和服务的尺度并不匹配(Pasari et al, 2013).研究生物多样性如何在多个尺度下提高生态系统功能对保护、管理和恢复多功能的生态景观非常重要, 但目前相关研究还很少. ...
Grazer diversity effects on ecosystem functioning in seagrass beds.
1
2003
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
Decomposer diversity and identity influence plant diversity effects on ecosystem functioning.
1
2012
... (3)对单个生态系统过程、功能或尺度分析的结果, 往往会低估生物多样性对生态系统功能的作用(Eisenhauer et al, 2012; Maestre et al, 2012b; Bowker et al, 2013; Pasari et al, 2013; Wagg et al, 2014; Lefcheck et al, 2015).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis.
1
2011
... 对于干旱、半干旱、半湿润的生态系统来说, 它们不仅要经受全球变化的影响(Maestre et al, 2012b, c), 而且还伴随着沙漠化(Maestre et al, 2012b)和灌木入侵的影响(Eldridge et al, 2011), 尤其值得一提的是灌木入侵也是全球旱地面临的主要问题之一(Knapp et al, 2008; Maestre et al, 2009; Li et al, 2013).Maestre等(2012b)对全球干旱区的研究发现, 在更寒冷和含沙量大的地区, 生态系统多功能性更高; 随着平均温度的增加, 在旱地生态系统中, 与C、N、P循环相关的多个功能将会降低, 而植物物种丰富度可能对维持与C循环和N循环相关的生态系统功能非常重要.研究结果还表明, 除了气候和非生物因子的影响外, 植物多样性是干旱区生态系统多功能性非常重要的驱动因素, 故而保护植物 多样性对缓解气候变化和沙漠化的负面影响非常 重要. ...
The Ecology of Invasions by Animals and Plants. Methuen, London.
2
1958
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
The microbial engines that drive Earth’s biogeochemical cycles.
3
2008
... 阐明地下生物多样性丧失给生态系统功能可能带来的影响至关重要, 因为土壤分类单元(soil taxa)中的生物几乎参与了每一个生物地球化学循环并在其中扮演了关键的角色, 让地球成为一个适于居住的星球(Falkowski et al, 2008; Wall & Bardgett, 2013).但到目前为止, 大多数的研究集中于地上生物多样性丧失的生态学影响(Wagg et al, 2014), 对土壤BEMF的研究相对缺乏. ...
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
... ).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Plant functional composition influences rates of soil carbon and nitrogen accumulation.
1
2008
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Multiple functions increase the importance of biodiversity for overall ecosystem functioning.
15
2008
... BEF研究的深入使研究者逐步意识到生态系统能同时提供多项功能和服务的事实, 即生态系统多功能性(ecosystem multifunctionality, EMF) (Sanderson et al, 2004), 亦即生态系统同时维持多种生态系统功能和服务的能力(Hector & Bagchi, 2007; Maestre et al, 2012b), 或者说生态系统多个功能的同时表现(Byrnes et al, 2014a).与此同时, 如何量化多样性丧失对EMF的影响, 以及生物多样性对多个生态系统功能的响应与其对单个生态系统功能的响应是否一致等问题应运而生(Byrnes et al, 2014a).在2007年之前, 大部分BEF研究都只考虑了生物多样性对单一生态过程的影响(Hector & Bagchi, 2007), 即使有研究测定了多个生态系统功能, 但对每个功能仍是独立地进行分析(Gamfeldt et al, 2008).Hector和Bagchi (2007)首次定量分析了生物多样性对多个生态系统过程的影响, 发现维持生态系统多功能性比维持单个生态系统功能需要更多的物种.由此, 生物多样性与生态系统多功能性的研究才受到人们的关注, 逐渐成为当前生态学研究的热点. ...
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
... (2)多功能冗余(multifunctional redundancy)比单功能冗余(single functional redundancy)程度低(Gamfeldt et al, 2008; Peter et al, 2011; Miki et al, 2014).所谓功能冗余是指有些物种在群落或生态系统中具有相似的功能, 这些物种的替代对生态系统功能的影响很小(Lawton & Brown, 1994).单功能冗余即单个功能的冗余, 也就是常说的功能冗余; 多功能冗余即多个功能的冗余, 也就是不同物种具有一种以上相似功能的情况. ...
... (4)多功能性比单个功能更易受到物种丧失的影响(Hector & Bagchi, 2007; Gamfeldt et al, 2008). ...
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
... (4)多功能性比单个功能更易受到物种丧失的影响(Hector & Bagchi, 2007; Gamfeldt et al, 2008). ...
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
... 生物多样性提升了生态系统维持多个功能如碳汇、生产力、养分库的积累的能力, 但这些研究一般是基于少数生态系统的小尺度受控实验, 且主要集中在北美和欧洲(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Maestre et al, 2012b), 缺少在全球尺度下的自然生态系统中以及从多个时间尺度上评估生物多样性和多功能性关系的研究(Maestre et al, 2012b).此外, 生物多样性并非生态系统功能的唯一驱动因素, 它同时还受其他生物和非生物因素的影响(Godbold & Solan et al, 2009; Maestre et al, 2012a), 因此, 需要在不同时间对不同地点、不同尺度的生态系统进行整合研究, 这些研究地点的空间差异应能充分体现资源可利用性、非生物因子、物种丰富度和物种组成等因素(Wardle & Jonsson, 2010)的影响. ...
... 生物多样性与生态系统多功能性的研究表明: 以前的研究低估了多样性的作用, 维持生态系统多功能性所需的生物多样性比预期的更高(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a,b; Perkins et al, 2014); 多样性丧失对多功能性的影响比对单个功能的影响更大(Gamfeldt et al, 2008).在生物多样性丧失加速的大背景下, 如何通过生物多样性来定量预测生态系统多功能性, 从而定量预测生物多样性丧失对生态系统多个功能造成的影响, 是当前亟需解决的问题, 而这方面的研究很少有人涉及.人类活动的加剧使生态系统不断遭到干扰, 而其遭受干扰后的恢复能力也将遭到严重破坏.Villnäs等(2013)的研究表明, 干扰会降低生态系统的多个功能以及生态系统对环境的适应能力.但是, 目前很少有研究评估生态系统承受的重复干扰, 以及其在此条件下维持多功能性的能力.如何能在持续获得生态系统产品和服务的同时减少对生态系统的干扰? 这方面的研究亟须加强. ...
Darwin and the first ecological experiment.
1
2002
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Plant diversity and productivity experiments in European grasslands.
1
1999
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Biodiversity effects on soil processes explained by interspecific functional dissimilarity.
1
2004
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Ploughing up the wood-wide web?
1
1998
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Biodiversity in a complex world: consolidation and progress in functional biodiversity research.
1
2009
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge.
4
2005
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... ), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
... 在过去10年间, 国内外已有很多关于BEF研究的综述文章(如刘峰等, 1999; Loreau, 2000; Loreau et al, 2001; 张全国和张大勇, 2003; Hooper et al, 2005; 江小雷等, 2010; Cardinale et al, 2012), Tilman等(2014)的综述再次对BEF过去的研究历程和未来的发展方向作了全面的展望. ...
The effects of plant composi tion and diversity on ecosystem processes.
1
1997
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Effects of plant composition and diversity on nutrient cycling.
1
1998
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity.
1997
2014
High plant diversity is needed to maintain ecosystem services.
5
2011
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
... 在一个生态系统中, 究竟需要多少个物种来维持其功能和服务, 研究者们仍然有不同的观点(Duffy, 2008; Ridder et al, 2008).如果仅考虑某个因素如不同时间、地点、功能或环境的差异, 则大多数物种都能提高生态系统功能, 但以前的研究没有同时考虑这些因子的时空变异(Isbell et al, 2011).此外, BEF实验常常在小尺度下, 通过测定有限数量的功能来研究生物多样性与生态系统功能的关系, 但是这与实际存在的生物多样性或生态系统功能和服务的尺度并不匹配(Pasari et al, 2013).研究生物多样性如何在多个尺度下提高生态系统功能对保护、管理和恢复多功能的生态景观非常重要, 但目前相关研究还很少. ...
Domesticated nature: shaping landscapes and ecosystems for human welfare.
2007
1
2001
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
Shrub encroachment in North American grasslands: shifts in growth form dominance rapidly alters control of ecosystem carbon inputs.
1
2008
... 对于干旱、半干旱、半湿润的生态系统来说, 它们不仅要经受全球变化的影响(Maestre et al, 2012b, c), 而且还伴随着沙漠化(Maestre et al, 2012b)和灌木入侵的影响(Eldridge et al, 2011), 尤其值得一提的是灌木入侵也是全球旱地面临的主要问题之一(Knapp et al, 2008; Maestre et al, 2009; Li et al, 2013).Maestre等(2012b)对全球干旱区的研究发现, 在更寒冷和含沙量大的地区, 生态系统多功能性更高; 随着平均温度的增加, 在旱地生态系统中, 与C、N、P循环相关的多个功能将会降低, 而植物物种丰富度可能对维持与C循环和N循环相关的生态系统功能非常重要.研究结果还表明, 除了气候和非生物因子的影响外, 植物多样性是干旱区生态系统多功能性非常重要的驱动因素, 故而保护植物 多样性对缓解气候变化和沙漠化的负面影响非常 重要. ...
Effects of plant species richness on invasion dynamics, disease outbreaks, insect abundances and diversity.
1
1999
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Redundancy in Ecosystems. Springer, Berlin.
1
1994
... (2)多功能冗余(multifunctional redundancy)比单功能冗余(single functional redundancy)程度低(Gamfeldt et al, 2008; Peter et al, 2011; Miki et al, 2014).所谓功能冗余是指有些物种在群落或生态系统中具有相似的功能, 这些物种的替代对生态系统功能的影响很小(Lawton & Brown, 1994).单功能冗余即单个功能的冗余, 也就是常说的功能冗余; 多功能冗余即多个功能的冗余, 也就是不同物种具有一种以上相似功能的情况. ...
Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.
4
2015
... (3)对单个生态系统过程、功能或尺度分析的结果, 往往会低估生物多样性对生态系统功能的作用(Eisenhauer et al, 2012; Maestre et al, 2012b; Bowker et al, 2013; Pasari et al, 2013; Wagg et al, 2014; Lefcheck et al, 2015).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
... ).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
... (5)不同营养级的物种丰富度对多功能性的影响不同, 食草动物的物种丰富度比植物物种丰富度对多功能性的影响更大(Lefcheck et al, 2015). ...
Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: results from Inner Mongolia steppe ecosystem of north China.
1
2013
... 对于干旱、半干旱、半湿润的生态系统来说, 它们不仅要经受全球变化的影响(Maestre et al, 2012b, c), 而且还伴随着沙漠化(Maestre et al, 2012b)和灌木入侵的影响(Eldridge et al, 2011), 尤其值得一提的是灌木入侵也是全球旱地面临的主要问题之一(Knapp et al, 2008; Maestre et al, 2009; Li et al, 2013).Maestre等(2012b)对全球干旱区的研究发现, 在更寒冷和含沙量大的地区, 生态系统多功能性更高; 随着平均温度的增加, 在旱地生态系统中, 与C、N、P循环相关的多个功能将会降低, 而植物物种丰富度可能对维持与C循环和N循环相关的生态系统功能非常重要.研究结果还表明, 除了气候和非生物因子的影响外, 植物多样性是干旱区生态系统多功能性非常重要的驱动因素, 故而保护植物 多样性对缓解气候变化和沙漠化的负面影响非常 重要. ...
Biodiversity influences plant productivity through niche-efficiency. Proceedings of the National Academy of Sciences,
1
2015
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
生物多样性的生态系统功能. 植物学通报,
1999
生物多样性的生态系统功能. 植物学通报,
1999
Biodiversity and ecosystem functioning: a mechanistic model. Proceedings of the National Academy of Sciences,
1998
Biodiversity and ecosystem functioning: recent theoretical advances.
1
2000
... 在过去10年间, 国内外已有很多关于BEF研究的综述文章(如刘峰等, 1999; Loreau, 2000; Loreau et al, 2001; 张全国和张大勇, 2003; Hooper et al, 2005; 江小雷等, 2010; Cardinale et al, 2012), Tilman等(2014)的综述再次对BEF过去的研究历程和未来的发展方向作了全面的展望. ...
1
2010
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
Biodiversity and ecosystem functioning: current knowledge and future challenges.
3
2001
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
... 在过去10年间, 国内外已有很多关于BEF研究的综述文章(如刘峰等, 1999; Loreau, 2000; Loreau et al, 2001; 张全国和张大勇, 2003; Hooper et al, 2005; 江小雷等, 2010; Cardinale et al, 2012), Tilman等(2014)的综述再次对BEF过去的研究历程和未来的发展方向作了全面的展望. ...
Fluctuations of animal populations and a measure of community stability.
1
1955
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
a) Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.
2012
b) Plant species richness and ecosystem multifunctionality in global drylands.
2012
c) It is getting hotter in here: determining and projecting the impacts of global environmental change on drylands.
2012
Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands.
1
2009
... 对于干旱、半干旱、半湿润的生态系统来说, 它们不仅要经受全球变化的影响(Maestre et al, 2012b, c), 而且还伴随着沙漠化(Maestre et al, 2012b)和灌木入侵的影响(Eldridge et al, 2011), 尤其值得一提的是灌木入侵也是全球旱地面临的主要问题之一(Knapp et al, 2008; Maestre et al, 2009; Li et al, 2013).Maestre等(2012b)对全球干旱区的研究发现, 在更寒冷和含沙量大的地区, 生态系统多功能性更高; 随着平均温度的增加, 在旱地生态系统中, 与C、N、P循环相关的多个功能将会降低, 而植物物种丰富度可能对维持与C循环和N循环相关的生态系统功能非常重要.研究结果还表明, 除了气候和非生物因子的影响外, 植物多样性是干旱区生态系统多功能性非常重要的驱动因素, 故而保护植物 多样性对缓解气候变化和沙漠化的负面影响非常 重要. ...
2
1973
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Biodiversity and multifunctionality in a microbial community: a novel theoretical approach to quantify functional redundancy.
3
2014
... (2)多功能冗余(multifunctional redundancy)比单功能冗余(single functional redundancy)程度低(Gamfeldt et al, 2008; Peter et al, 2011; Miki et al, 2014).所谓功能冗余是指有些物种在群落或生态系统中具有相似的功能, 这些物种的替代对生态系统功能的影响很小(Lawton & Brown, 1994).单功能冗余即单个功能的冗余, 也就是常说的功能冗余; 多功能冗余即多个功能的冗余, 也就是不同物种具有一种以上相似功能的情况. ...
... 目前, 国际上常用的多功能性测度方法主要包括: 单功能法(single-function approach) (Duffy et al, 2003)、功能-物种替代法(turnover approach) (Hector & Bagchi, 2007)、平均值法(averaging approach) (Hooper & Vitousek, 1998)、单阈值法(single threshold approach) (Gamfeldt et al, 2008)和多阈值法(multiple-threshold approach) (Byrnes et al, 2014a); 此外, 还有一些比较新颖的方法, 如直系同源基因法(orthologous approach) (Miki et al, 2014)和多元模型法(multivariate model approach) (Dooley et al, 2015). ...
... 以上研究说明, 多功能冗余在陆地植物和微生物群落中普遍存在, 且多功能冗余往往是有限的, 比单功能冗余程度低.多功能冗余在不同的区域和生境中表现不同, 具有很强的环境依赖性(Miki et al, 2014).冗余度决定着生态系统整体功能对物种丧失的响应, 在生物多样性丧失的背景下, 为确保生态系统具有较高的综合功能, 需要考虑多功能冗余的程度(Gamfeldt et al, 2008).此外, 有研究表明, 即使在冗余程度很高的群落中, 某些稀有种提供的功能仍是十分脆弱的, 一旦这些物种消失, 它们提供的功能也将随之丧失(Mouillot et al, 2013).因此, 稀有种的物种丰富度对一个生态系统的多功能性也可能有很大影响, 未来应当加强这方面的研究. ...
Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland.
1
2008
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
The Millennium Ecosystem Assessment: what is it all about?
1
2004
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
Global human footprint on the linkage between biodiversity and ecosystem functioning in reef fishes.
1
2011
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
Rare species support vulnerable functions in high-diversity ecosystems.
1
2013
... 以上研究说明, 多功能冗余在陆地植物和微生物群落中普遍存在, 且多功能冗余往往是有限的, 比单功能冗余程度低.多功能冗余在不同的区域和生境中表现不同, 具有很强的环境依赖性(Miki et al, 2014).冗余度决定着生态系统整体功能对物种丧失的响应, 在生物多样性丧失的背景下, 为确保生态系统具有较高的综合功能, 需要考虑多功能冗余的程度(Gamfeldt et al, 2008).此外, 有研究表明, 即使在冗余程度很高的群落中, 某些稀有种提供的功能仍是十分脆弱的, 一旦这些物种消失, 它们提供的功能也将随之丧失(Mouillot et al, 2013).因此, 稀有种的物种丰富度对一个生态系统的多功能性也可能有很大影响, 未来应当加强这方面的研究. ...
Root depth distribution and the diversity-productivity relationship in a long-term grassland experiment.
1
2013
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Plant diversity increases resistance to invasion in the absence of covarying extrinsic factors.
1
2000
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Biodiversity enhances ecosystem reliability.
1
1997
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Declining biodiversity can affect the functioning of ecosystems.
1
1994
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Soil biodiversity and carbon cycling: a review and synthesis of studies examining diversity-function relationships.
1
2011
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
The effect of biodiversity on tree productivity: from temperate to boreal forests.
1
2011
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
Several scales of biodiversity affect ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
4
2013
... (3)对单个生态系统过程、功能或尺度分析的结果, 往往会低估生物多样性对生态系统功能的作用(Eisenhauer et al, 2012; Maestre et al, 2012b; Bowker et al, 2013; Pasari et al, 2013; Wagg et al, 2014; Lefcheck et al, 2015).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
... 然而, BEMF研究仍存在许多问题, 如缺少公认的测定多功能性指数的测度标准(Bradford et al, 2014b; Byrnes et al, 2014a); 生态系统不同功能之间的权衡(trade-off)制约着多功能性的客观评价(Vinebrooke et al, 2004; Zavaleta et al, 2010, Byrnes et al, 2014a); 缺少在不同时空尺度上的研究(Maestre et al, 2012b; Pasari et al, 2013); 有关地下BEMF的研究相对缺乏(Bradford et al, 2014a; Wagg et al, 2014)等. ...
... 在一个生态系统中, 究竟需要多少个物种来维持其功能和服务, 研究者们仍然有不同的观点(Duffy, 2008; Ridder et al, 2008).如果仅考虑某个因素如不同时间、地点、功能或环境的差异, 则大多数物种都能提高生态系统功能, 但以前的研究没有同时考虑这些因子的时空变异(Isbell et al, 2011).此外, BEF实验常常在小尺度下, 通过测定有限数量的功能来研究生物多样性与生态系统功能的关系, 但是这与实际存在的生物多样性或生态系统功能和服务的尺度并不匹配(Pasari et al, 2013).研究生物多样性如何在多个尺度下提高生态系统功能对保护、管理和恢复多功能的生态景观非常重要, 但目前相关研究还很少. ...
Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes.
2
2015
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
Multifunctionality and diversity in bacterial biofilms.
4
2011
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
... (2)多功能冗余(multifunctional redundancy)比单功能冗余(single functional redundancy)程度低(Gamfeldt et al, 2008; Peter et al, 2011; Miki et al, 2014).所谓功能冗余是指有些物种在群落或生态系统中具有相似的功能, 这些物种的替代对生态系统功能的影响很小(Lawton & Brown, 1994).单功能冗余即单个功能的冗余, 也就是常说的功能冗余; 多功能冗余即多个功能的冗余, 也就是不同物种具有一种以上相似功能的情况. ...
... Miki等(2014)开发了一种新颖的理论研究方法, 通过对比分析微生物基因组数据库(MBGD), 定量评估了微生物群落的多功能性.他们以直系同源基因丰富度作为量化微生物群落多功能性的指标, 通过群落模拟实验证明, 多功能冗余程度通常很低, 且比单功能冗余程度更低, 这与其他研究(Dan- ovaro et al, 2008; Gamfeldt et al, 2008; Peter et al, 2011)的结果一致. ...
... 生物多样性与生态系统多功能性的研究表明: 以前的研究低估了多样性的作用, 维持生态系统多功能性所需的生物多样性比预期的更高(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a,b; Perkins et al, 2014); 多样性丧失对多功能性的影响比对单个功能的影响更大(Gamfeldt et al, 2008).在生物多样性丧失加速的大背景下, 如何通过生物多样性来定量预测生态系统多功能性, 从而定量预测生物多样性丧失对生态系统多个功能造成的影响, 是当前亟需解决的问题, 而这方面的研究很少有人涉及.人类活动的加剧使生态系统不断遭到干扰, 而其遭受干扰后的恢复能力也将遭到严重破坏.Villnäs等(2013)的研究表明, 干扰会降低生态系统的多个功能以及生态系统对环境的适应能力.但是, 目前很少有研究评估生态系统承受的重复干扰, 以及其在此条件下维持多功能性的能力.如何能在持续获得生态系统产品和服务的同时减少对生态系统的干扰? 这方面的研究亟须加强. ...
Impacts of biodiversity loss escalate through time as redundancy fades.
1
2012
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Questioning the ecosystem services argument for biodiversity conservation.
1
2008
... 在一个生态系统中, 究竟需要多少个物种来维持其功能和服务, 研究者们仍然有不同的观点(Duffy, 2008; Ridder et al, 2008).如果仅考虑某个因素如不同时间、地点、功能或环境的差异, 则大多数物种都能提高生态系统功能, 但以前的研究没有同时考虑这些因子的时空变异(Isbell et al, 2011).此外, BEF实验常常在小尺度下, 通过测定有限数量的功能来研究生物多样性与生态系统功能的关系, 但是这与实际存在的生物多样性或生态系统功能和服务的尺度并不匹配(Pasari et al, 2013).研究生物多样性如何在多个尺度下提高生态系统功能对保护、管理和恢复多功能的生态景观非常重要, 但目前相关研究还很少. ...
Plant species diversity and management of temperate forage and grazing land ecosystems.
1
2004
... BEF研究的深入使研究者逐步意识到生态系统能同时提供多项功能和服务的事实, 即生态系统多功能性(ecosystem multifunctionality, EMF) (Sanderson et al, 2004), 亦即生态系统同时维持多种生态系统功能和服务的能力(Hector & Bagchi, 2007; Maestre et al, 2012b), 或者说生态系统多个功能的同时表现(Byrnes et al, 2014a).与此同时, 如何量化多样性丧失对EMF的影响, 以及生物多样性对多个生态系统功能的响应与其对单个生态系统功能的响应是否一致等问题应运而生(Byrnes et al, 2014a).在2007年之前, 大部分BEF研究都只考虑了生物多样性对单一生态过程的影响(Hector & Bagchi, 2007), 即使有研究测定了多个生态系统功能, 但对每个功能仍是独立地进行分析(Gamfeldt et al, 2008).Hector和Bagchi (2007)首次定量分析了生物多样性对多个生态系统过程的影响, 发现维持生态系统多功能性比维持单个生态系统功能需要更多的物种.由此, 生物多样性与生态系统多功能性的研究才受到人们的关注, 逐渐成为当前生态学研究的热点. ...
Biodiversity and Ecosystem Function. Springer-Verlag, Berlin.
1
1993
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Ecosystem Function of Biodiversity: A Summary. Springer, Berlin, Heidelberg.
1
1994
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
Influence of body size of soil fauna on litter decomposition and 15N uptake by poplar in a pot trial.
1
1996
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Phylogenetic diversity and the functioning of ecosystems.
1
2012
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
The ecological consequences of changes in biodiversity: a search for general principles.
Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences,
1
2004
... 随着实验数据的不断积累, 一系列的关键发现使BEF研究实现了里程碑式的发展, 许多之前的争论逐渐平息(Cardinale et al, 2012).截至2009年, 已开展了600多项相关实验, 研究对象包括500多种淡水、海洋和陆地生物(Loreau et al, 2002; Cardinale et al, 2011), 已有数百篇相关研究论文发表.除了实验数量的增加, BEF研究还提出了一些相关的理论模型(Tilman et al, 1997; Kinzig et al, 2001; Tilman, 2004; Loreau, 2010; Liang et al, 2015), 并将其范围扩展至全球自然生态系统(Mora et al, 2011; Paquette & Messier, 2011; Maestre et al, 2012b).人们对生物多样性与生态系统功能的关系有了一些共识(详见Cardinale et al, 2012). ...
Biodiversity and stability in grasslands.
2
1994
... 事实上, BEF研究最早可追溯到达尔文时代, 那时, 乔治•辛克莱尔(George Sinclair)在英国贝德福德郡(Bedfordshire)对比了单作(monoculture)和混种(mixture)对植物生产力的影响, 结果发现物种多样性越高, 干草的产量也越高(Hector & Hooper, 2002).之后, 研究者虽然意识到多样性会影响生态系统的过程或功能(Carlander, 1955), 但多数研究仍集中于多样性与生态系统稳定性的关系等方面(MacArthur, 1955; Elton, 1958; May, 1973).1975年, Goodman发表了关于“多样性与稳定性”的综述, 之后BEF研究淡出了人们的视野.1992年, 在德国举行的主题为“生物多样性与生态系统功能(Biodiversity and Ecosystem Function)”的会议上, 生态学家才对BEF进行了新一轮的研讨(Schulze & Mooney, 1994).同年6月, 在巴西举行了联合国世界环境与发展会议(United Nations Conference on Environment and Development), 研究者重新审视了生物多样性丧失对生态系统功能和动态及其提供的产品和服务产生的影响(Cardinale et al, 2012).而1993年出版的专著Biodiversity and Ecosystem Function (Schulze & Mooney, 1993)为现代BEF研究奠定了基础(贺金生等, 2003).随后, 出现了大量的BEF实验研究, 比较有代表性的有生态气候室(ecotron)实验(Naeem et al, 1994)、 Cedar Creek野外实验(Tilman & Downing, 1994; Tilman et al, 1996, 2001)、微宇宙(microcosm)实验(McGrady-Steed et al, 1997; Naeem & Li, 1997)、美国加州草地实验(Hooper & Vitousek, 1997)、欧洲草地BEF实验(Hector et al, 1999)等.其中多数实验结果认为, 植物多样性越高, 群落生产力越高、生态系统稳定性和抗入侵能力等也越强(张全国和张大勇, 2003; Tilman et al, 2014). ...
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
1
2008
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Mycorrhizal ecology and evolution: the past, the present, and the future.
2015
The interference production principle: an ecological theory for agriculture.
The role of recurrent disturbances for ecosystem multifunctionality.
2013
Impacts of multiple stressors on biodiversity and ecosystem functioning: the role of species co-tolerance.
2
2004
... 然而, BEMF研究仍存在许多问题, 如缺少公认的测定多功能性指数的测度标准(Bradford et al, 2014b; Byrnes et al, 2014a); 生态系统不同功能之间的权衡(trade-off)制约着多功能性的客观评价(Vinebrooke et al, 2004; Zavaleta et al, 2010, Byrnes et al, 2014a); 缺少在不同时空尺度上的研究(Maestre et al, 2012b; Pasari et al, 2013); 有关地下BEMF的研究相对缺乏(Bradford et al, 2014a; Wagg et al, 2014)等. ...
... 有研究认为, 生物多样性与生态系统多功能性的关系至少受到两种重要权衡的影响(Gamfeldt et al, 2008): 一是由于某些功能间的排斥作用, 单一群落很难同时维持多个特定功能, 例如生产力和抗逆性常常是负相关的, 因此很难或不可能同时最大化(Grime, 1974); 二是不同的功能需要不同物种丰富度或组成的群落来使其最大化(Vinebrooke et al, 2004), 如不同乡土物种的组合能使其对入侵物种的抵抗力最大化(Zavaleta et al, 2010).Kareiva等(2007)指出, 重要的生态系统功能间常常存在权衡关系, 即一个功能(如生物量)的增加可能会伴随另一个功能(如对疾病的抵抗力)的降低.研究表明, 权衡作用限制了大多数物种组合的某些功能达到较高水平, 如高生产力与高抗旱性很难同时达到最高水平(Grime, 1974; Díaz et al, 2004).此外, 群落中可能有一些功能相似的物种, 它们能提供少数较高水平的功能; 或者具有功能多样性的物种, 它们能同时提供较高水平的不同功能(Zavaleta et al, 2010). ...
Diversity effects in early- and mid-successional species pools along a nitrogen gradient.
1
2009
... 然而, 现阶段大量的研究关注生物多样性(主要是物种多样性)与生态系统功能的关系; 虽然新兴的生物多样性与生态系统多功能性的研究也在逐渐增加, 但同时考虑全球变化的研究却不多.另外, 需要强调的是生物多样性包括物种多样性、功能多样性和谱系多样性三个维度, 其中功能多样性指影响植物功能的形态、生理或物候性状的组成与变化(Hillebrand & Matthiessen, 2009), 谱系多样性指一个群落中物种谱系距离的总和, 它受平均种间亲缘关系和群落中物种数量的影响(Srivastava et al, 2012).当前的研究常常局限于物种多样性与生态系统多功能性之间的关系, 而忽略了功能多样性和谱系多样性.功能多样性和谱系多样性与物种多样性息息相关, 它们都与生态系统功能有着密不可分的关系.例如, 已经有许多研究指出功能多样性对群落的功能有重要影响(Mokany et al, 2008; Cadotte et al, 2009; Wacker et al, 2009), 最近关于功能多样性与多功能性的研究发现: 高的功能多样性能增加生态系统多功能性对干旱的抗性(Valencia et al, 2015).缺乏这两个多样性维度的研究是不全面的, 由此, 我们建立了BEF及未来应当考虑的不同维度多样性与多功能性关系的研究框架(图3).希望未来的研究能立足于全球变化的大背景, 在物种多样性、功能多样性和谱系多样性水平上, 研究它们如何影响生态系统多功能性. ...
Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
7
2014
... 地球的最独特之处是存在生命, 而生命最奇妙的特点是其多样性(Cardinale et al, 2012).然而, 在全球范围内, 随着人类社会活动的加剧和气候变化, 生物多样性正以惊人的速度丧失.不断积累的证据表明, 生物多样性丧失会降低生态系统的功能和服务(Loreau et al, 2001; Hooper et al, 2005; Balvanera et al, 2006; Cardinale et al, 2006, 2011; Wagg et al, 2014), 如生产力下降、养分循环失衡、传粉能力下降等(Hooper et al, 2005; Cardinale et al, 2006, 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... , 2012; Wagg et al, 2014).人类社会的幸福感依赖于生态系统提供的产品和服务, 而这些则直接来自于生态系统功能(ecosystem functioning) (Mooney et al, 2004).因此, 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)之间的关系是生态学研究关注的热点之一. ...
... (3)对单个生态系统过程、功能或尺度分析的结果, 往往会低估生物多样性对生态系统功能的作用(Eisenhauer et al, 2012; Maestre et al, 2012b; Bowker et al, 2013; Pasari et al, 2013; Wagg et al, 2014; Lefcheck et al, 2015).随着所考虑的功能数的增加, 多样性对多功能性的作用会变得越来越重要(Lefcheck et al, 2015). ...
... 然而, BEMF研究仍存在许多问题, 如缺少公认的测定多功能性指数的测度标准(Bradford et al, 2014b; Byrnes et al, 2014a); 生态系统不同功能之间的权衡(trade-off)制约着多功能性的客观评价(Vinebrooke et al, 2004; Zavaleta et al, 2010, Byrnes et al, 2014a); 缺少在不同时空尺度上的研究(Maestre et al, 2012b; Pasari et al, 2013); 有关地下BEMF的研究相对缺乏(Bradford et al, 2014a; Wagg et al, 2014)等. ...
... 阐明地下生物多样性丧失给生态系统功能可能带来的影响至关重要, 因为土壤分类单元(soil taxa)中的生物几乎参与了每一个生物地球化学循环并在其中扮演了关键的角色, 让地球成为一个适于居住的星球(Falkowski et al, 2008; Wall & Bardgett, 2013).但到目前为止, 大多数的研究集中于地上生物多样性丧失的生态学影响(Wagg et al, 2014), 对土壤BEMF的研究相对缺乏. ...
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
... ); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
2
2013
... 阐明地下生物多样性丧失给生态系统功能可能带来的影响至关重要, 因为土壤分类单元(soil taxa)中的生物几乎参与了每一个生物地球化学循环并在其中扮演了关键的角色, 让地球成为一个适于居住的星球(Falkowski et al, 2008; Wall & Bardgett, 2013).但到目前为止, 大多数的研究集中于地上生物多样性丧失的生态学影响(Wagg et al, 2014), 对土壤BEMF的研究相对缺乏. ...
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Biodiversity in the dark.
1
2010
... 到目前为止, 因地下生物多样性研究较为困难, 如大量微生物类群的功能并不明确; 高通量测序数据量大, 分析过程复杂等, 大多数的研究集中于地上生物多样性丧失的生态学影响.而地球上很大一部分生物隐藏在地下(Wagg et al, 2014), 土壤生物类群在几乎所有的生物地球化学过程中均有着至关重要的作用(Falkowski et al, 2008; Wall & Bardgett, 2013), 其多样性的丧失可能严重影响生态系统功能, 如植物的养分获取能力、地上-地下群落的养分循环等(Helgason et al, 1998; van der Heijden et al, 2008; Wall et al, 2010; de Vries et al, 2013).但现在的地下生物多样性研究主要集中在特定群落, 如土壤微生物、菌根真菌、土壤动物等, 以及野外大尺度的相关分析, 土壤生物多样性和生态系统功能的一般关系尚不明确.不同研究结果表明, 土壤生物多样性对生态系统过程具有正、负或中性效应(Setälä et al, 1996; Bradford et al, 2002; de Deyn et al, 2003; Heemsbergen et al, 2004; Nielsen et al, 2011); 然而, 土壤群落多样性的减少是否会影响生态系统的综合功能仍不得而知(Wagg et al, 2014).因此, 阐明地下生物多样性丧失对生态系统功能的潜在影响具有重要意义(Falkowski et al, 2008; Bradford et al, 2014a). ...
Biodiversity effects in real ecosystems: a response to Duffy.
1
2010
... 生物多样性提升了生态系统维持多个功能如碳汇、生产力、养分库的积累的能力, 但这些研究一般是基于少数生态系统的小尺度受控实验, 且主要集中在北美和欧洲(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Maestre et al, 2012b), 缺少在全球尺度下的自然生态系统中以及从多个时间尺度上评估生物多样性和多功能性关系的研究(Maestre et al, 2012b).此外, 生物多样性并非生态系统功能的唯一驱动因素, 它同时还受其他生物和非生物因素的影响(Godbold & Solan et al, 2009; Maestre et al, 2012a), 因此, 需要在不同时间对不同地点、不同尺度的生态系统进行整合研究, 这些研究地点的空间差异应能充分体现资源可利用性、非生物因子、物种丰富度和物种组成等因素(Wardle & Jonsson, 2010)的影响. ...
Functional redundancy supports biodiversity and ecosystem function in a closed and constant environment.
Realistic species losses disproportionately reduce grassland resistance to biological invaders.
1
2004
... 到2006年, 超过100个生物多样性实验的证据表明: 物种多样性与生产力呈正相关关系(Loreau et al, 2001; Tilman et al, 2001; Hooper et al, 2005; Cardinale et al, 2006).经过多点实验(multisite experiments) (如BIODEPTH和Agrodiversity实验网络)和整合分析(meta-analysis) (Cardinale et al, 2006, 2007, 2011, 2012; Gross et al, 2014), 发现多样性对生态系统功能的作用具有普遍性.与此同时, 研究者发现: 在自然生态系统中, 种间作用、群落和生态系统间的反馈都需要较长的时间, 而多数实验因其时间短, 往往无法捕捉到这些作用, 从而低估了生物多样性对生态系统功能的作用(Cardinale et al, 2007; Fornara & Tilman, 2008; Reich et al, 2012; Mueller et al, 2013).经过10年的Cedar Creek实验, 研究者发现多样性高的群落其时间稳定性显著高于多样性低的群落, 这种高的群落稳定性更多来自于群落生物量随多样性的增加而显著地增加(远远超过了稳定性在时间上的差异), 而不是统计平均; 而单个物种的稳定性在较高的多样性下反而较低(Tilman et al, 2006).多年的草地生物多样性实验提供了大量的证据: 当多样性逐渐增加时, 群落抵御入侵的能力会随之增强(Knops et al, 1999; Naeem et al, 2000; Zavaleta & Hulvey, 2004). ...
Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences,
8
2010
... (1)在不同的时空尺度、环境条件下, 维持生态系统多个功能比单个功能需要更多的物种(Hector & Bagchi, 2007; Gamfeldt et al, 2008; Zavaleta et al, 2010; Isbell et al, 2011; Peter et al, 2011; Maestre et al, 2012a, b; Perkins et al, 2015). ...
... 然而, BEMF研究仍存在许多问题, 如缺少公认的测定多功能性指数的测度标准(Bradford et al, 2014b; Byrnes et al, 2014a); 生态系统不同功能之间的权衡(trade-off)制约着多功能性的客观评价(Vinebrooke et al, 2004; Zavaleta et al, 2010, Byrnes et al, 2014a); 缺少在不同时空尺度上的研究(Maestre et al, 2012b; Pasari et al, 2013); 有关地下BEMF的研究相对缺乏(Bradford et al, 2014a; Wagg et al, 2014)等. ...


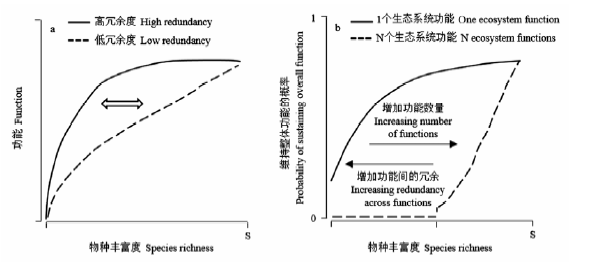
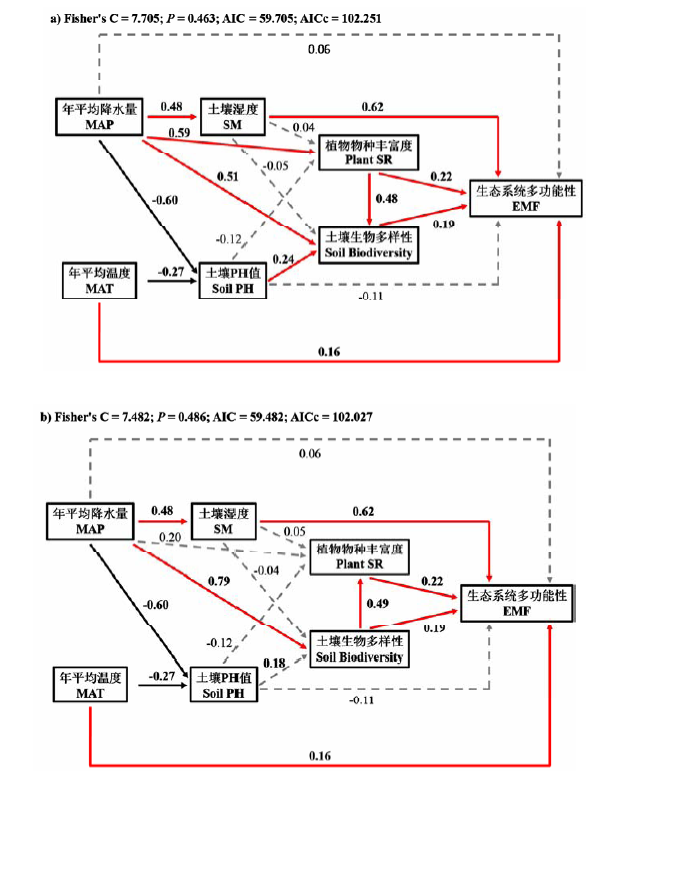
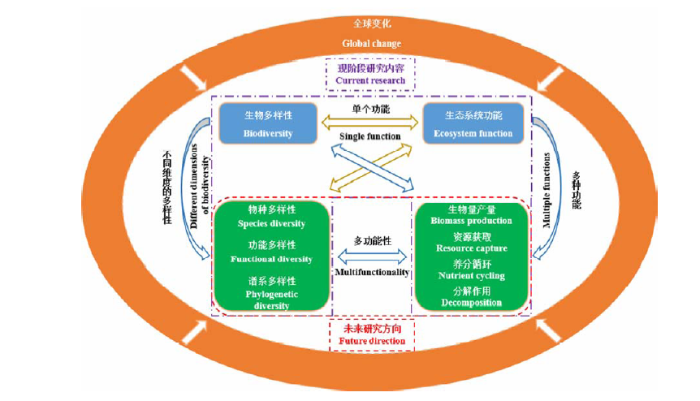
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}