物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 )。影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 )。由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 )。在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 )。影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 )。除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 )。一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 )。由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见。
1 研究地基本概况
雅鲁藏布江源自于喜马拉雅山北麓的杰马央宗冰川, 全长约3,350 km, 中国境内长达2,052 km(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 由西向东沿喜马拉雅山脉贯穿整个藏南河谷, 在东部绕过喜马拉雅山脉最东端的南迦巴瓦峰转向印度洋。从源头到东部大峡谷, 地质地貌复杂独特, 海拔高差明显, 气候变化多样。上游呈高原宽谷地貌, 谷底海拔约4,500-4,800 m; 中游河谷宽、狭相间, 谷底海拔为3,500-4,400 m; 而下游山高谷深, 水流湍急, 植物组成十分复杂多样。整个雅鲁藏布江流域包括了干旱、半干旱半湿润、湿润3种气候特征(赵鲁青, 2011)① 。流域内气温和降水量由西向东随海拔的降低而呈升高或增加趋势。本研究区内雅鲁藏布江下游地区年平均气温可达约11℃以上, 而在上游地区只有约-1℃左右(表1 )。整个流域内植被类型丰富, 上游谷底属于高寒草原和草甸, 而中游广大河谷区是喜温的半干旱草原和落叶阔叶灌丛草原, 下游逐渐向复杂的森林植被类型过渡(中国科学院青藏高原综合科学考察队, 1988 )。
2 方法
2.1 取样
于2011年8月至2012年8月分两次进行野外研究工作, 包括样方的调查、标本的采集和植物标本的室内鉴定等。从雅鲁藏布江的源头区, 即从仲巴县帕羊镇(30º01′ N, 83º28′ E)南侧开始, 沿着海拔梯度, 从西向东沿雅鲁藏布江河岸共设置了15个海拔段(附图1 ), 分别为4,593 m、4,580 m、4,490 m、4,010 m、3,892 m、3,789 m、3,716 m、3,590 m、3,620 m、3,570 m、3,580 m、3,550 m、3,220 m、3,170 m和2,950 m。样地既基本覆盖了河流经过的15个行政县区域, 又基本能保证在地理空间上具有一定的代表性。在每个海拔段河岸植物群落里, 随机设置6个5 m×5 m样方。每个样方设置时尽可能考虑生境相对较为均匀和自然, 而避开极端小生境(如人为活动干扰严重、遭受地质灾害的地方), 并保证各样方基本能代表每个海拔段内最典型、最常见的自然植被类型。这样, 15个海拔段共设置了90个样方, 详细样地信息见表1 。详细记录每个样方内所有维管束植物物种组成, 同时, 测定每个样方的海拔、经度、纬度和植被盖度。绝大多数植物物种通过野外识别和室内标本鉴定识别到了种水平, 少数植物因缺花少果而暂时不能鉴定到种, 以属的名称+数字加以区别和记录, 如棘豆属一种, 记成Oxytropis sp.1。
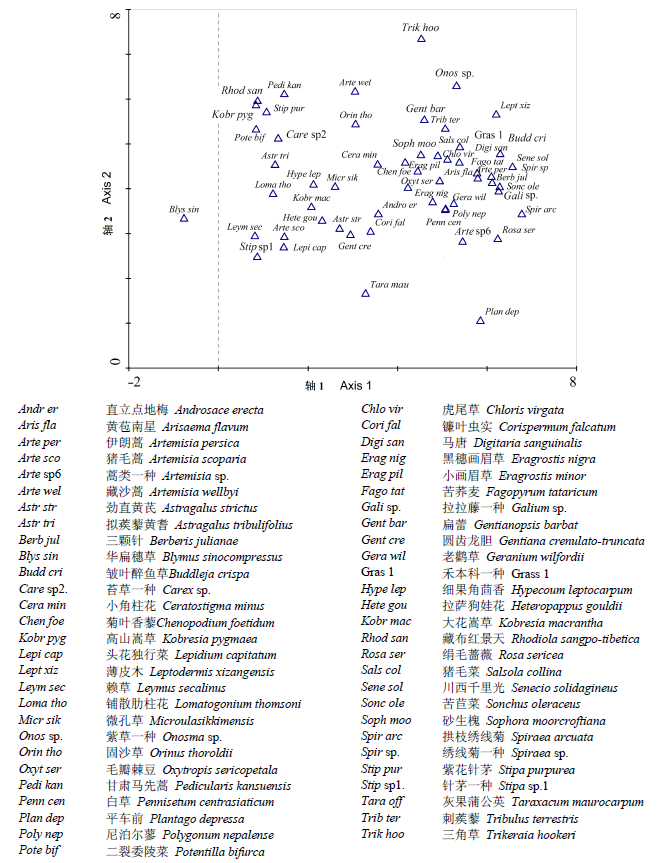
图1 雅鲁藏布江河岸植物物种DCA排序图。样方中出现频率很低的物种未在图中显示。
Fig. 1 DCA species ordination for the riparian plots along the Yarlung Zangbo River. Species with low frequency have been removed from the diagram.
各样方的气象数据如年均降水量、年均气温采用15个海拔段所属行政县气象观测站的近35年气象数据平均值。因为每个海拔样带设置的地点考虑到了尽可能靠近河流经过各行政县的位置(最远不超过15 km), 因此, 各县气象观测站的气象数据基本能够表征样方所处的局地气候。
2.2 数据分析
排序的方法能够比较客观地反映植物群落物种分布格局、群落类型及群落与生态环境之间的关系。排序一般分为间接排序和直接排序, 间接排序即在特定的环境轴上探讨物种的变化情况, 分析和推测其可能的环境梯度, 而直接排序用潜在的环境梯度或已测定环境因子数据来解释物种或植被类型变化(Lepš & Šmilauer, 2003 )。因此, 本研究先采用间接排序方法, 即除趋势对应分析方法(Detrended Correspondence Analysis, DCA)(Hill & Cauch, 1980 )对整个样方物种组成进行分析, 揭示所调查植物群落物种组成的空间变化, 并分析影响其格局的可能环境因子。在此基础上, 进一步采用直接排序方法, 即典范对应分析方法(Canonical Correspondence Analysis, CCA), 用本研究直接测得的环境数据, 进一步对物种数据进行分析, 确认植被类型空间分布格局与具体测定的生态环境因子之间是否存在显著相关性, 即海拔、经度、纬度、各海拔样带年均降雨量和年均温度和植被空间分布格局之间关系。CCA排序特别适合用来分析植物群落与具体环境因子之间关系(Braak, 1986 )。
为了分析物种丰富度分布格局与环境因子之间关系, 我们采用广义线性回归模型(Generalized Linear Model, GLM)(McCullagh & Nelder, 1989 )进行回归分析, 研究物种丰富度分布格局与海拔、经度、纬度、年均降雨量、年均温度和植被盖度之间的关系。
有关排序分析采用计算机软件CANOCO 4.5完成, 统计回归分析采用统计软件S-Plus 6.2(2003) 完成计算。
3 结果
3.1 物种组成
90个样方共记录维管束植物238种, 常见植物名录见附表1 。整个样方中出现频度最高的前三大科分别是菊科、禾本科和豆科。出现频度较高的物种有: 菊科的拉萨狗娃花(Heteropappus gouldii )(29次)和藏沙蒿(Artemisia wellbyi )(27次)、伊朗蒿(A. persica )(19次)等蒿属植物; 豆科的砂生槐(Sophora moorcroftiana )(55次)、劲直黄芪(Astragalus strictus ) (29次)、拟蒺藜黄芪(A. tribulifolius )(16次)、毛瓣棘豆(Oxytropis sericopetala )(10次); 禾本科的小画眉草(Eragrostis minor )(41次)、白草(Pennisetum flaccidum )(40次)、固沙草(Orinus thoroldii )(26次)和黑穗画眉草(E. nigra )(18次); 紫草科的微孔草(Microula sikkimensis )(29次)等。
3.2 物种和样方排序
3.2.1 DCA物种排序
从DCA分析总结表(表2 )中可看出, 排序轴1的特征值总和是0.65, 梯度长度为5.95, 物种与环境因子的相关系数约是0.94, 排序轴1解释了8%的物种变化。而排序轴2的特征值总和是0.47, 梯度长度为5.96, 物种与环境因子的相关系数是0.54, 与排序轴1一起累计解释了物种组成与环境因子之间关系的13.7%, 排序轴2单独解释了5.7%的物种变化。排序轴3和4的各项数据指标总体上都远低于前两者。通过综合分析这些数据信息, 表明第1轴和第2轴是最能反映群落物种组成变化的环境梯度。物种在DCA二维空间上的排序显示(图1 ), 像华扁穗草(Blymus sinocompressus )、赖草(Leymus secalinus )、针茅属一种(Stipa sp.1)、二裂委陵菜(Potentilla bifurca )、高山嵩草(Kobresia pygmaea )、藏布红景天(Rhodiola sangpo-tibetana )、铺散肋柱花(Lomatogonium thomsoni )等都显著出现在DCA第1轴的左侧, 这些物种都是高山草原群落的组成成分, 主要分布在高海拔(低经度)的雅鲁藏布江源头区。而薄皮木(Leptodermis xizangensis )、皱叶醉鱼草(Buddleja crispa )、川西千里光(Senecio solidagineus )、绣线菊(Spiraea sp.)、三颗针(Berberis julianae )、拉拉藤(Galium sp.)、拱枝绣线菊(Spiraea arcuata )、绢毛蔷薇 (Rosa sericea )等都出现在第1轴右侧, 这些物种大都来自于相对较低海拔(相对高经度)的雅鲁藏布江下游的林缘生境里, 表明第1轴是以海拔和经度作为主导因子的环境梯度, 经度梯度跟海拔梯度完全反向相偶联(图2 )。可见, 海拔和经度对于整个河岸植物物种的分布格局起重要作用。
然而, 第2轴的环境梯度不像第1轴那么清晰, 也很难看出是由哪个单一环境梯度所控制, 如平车前(Plantago depressa )、灰果蒲公英(Taraxacum maurocarpum )等物种被排在第2排序轴的下方, 这些物种往往出现在较为潮湿和人为干扰较大的生境里, 而像三角草(Trikeraia hookeri )、紫草一种(Onosma sp.)、藏沙蒿等被排在在第2排序轴的上方, 这些物种一般都出现在偏干沙性土壤生境里和人为干扰较少的草原自然生境内。因此, 第2排序轴很可能是跟湿度和干扰等多种环境因子相结合的较为复杂的一种复合梯度。
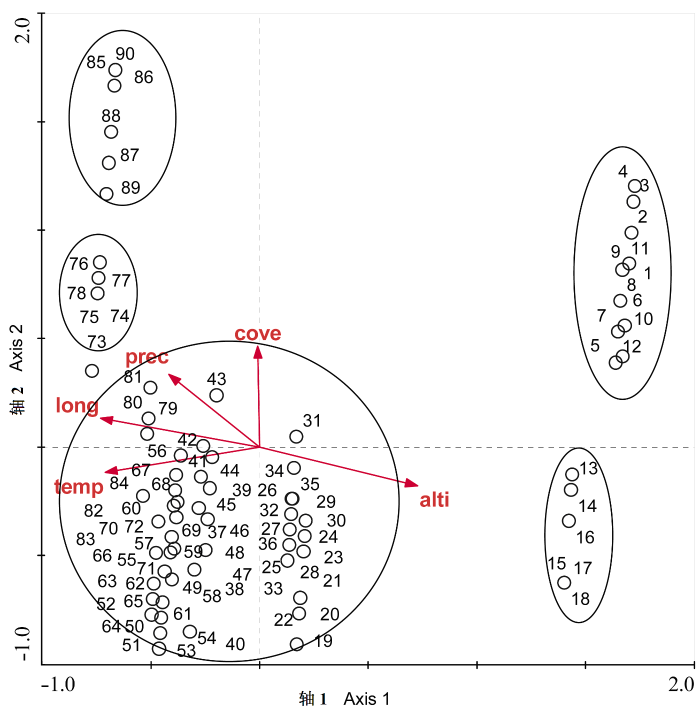
图2 雅鲁藏布江流域河岸植物群落调查样方CCA排序图。图中数字表示样方号(样方号的详细信息见表1 ), 为了使图更为清晰少数样方未在图中展示。
Fig. 2 CCA ordination for the surveyed riparian plots along the Yarlung Zangbo River. The numbers represent plot No. (detail information for each plot number see Table 1 ). Some plots have been removed from the diagram for clarity. alti = altitude, long = longitude, temp = temperature, prec = precipitation, cove = cover.
3.2.2 CCA样方排序
经CCA排序, 群落样方在二维排序图上可以明显地划分为5个大的类群, 即5种植被类型(图2 ), 其特征分别描述如下:
(1)分布在较高海拔的高寒草甸或高寒草原植被类型: 主要是由各种嵩草(Kobresia spp.)群落组成的草甸植被, 以及由三角草群落、紫花针茅(Stipa purpurea )群落等组成的草原植被, 包括被排在CCA第1轴右上侧的样方1-12.
(2)以青藏苔草(Carex moocroftii )、固沙草、藏沙蒿等为主形成的高山草原群落: 如被排在第1轴右下角的样方13-18。
(3)亚高山灌丛草原: 这一类型分布最广, 主要覆盖区包括拉孜、日喀则、尼木、曲水、乃东等县流域。整个调查样方的73%(样方19-84)属于这一植被类型。主要由砂生槐群落、白草群落和劲直黄芪群落等组成。局部还有藏沙蒿群落、固沙草和其他蒿属草原群落。这些样方占据着CCA第1排序轴的中稍偏左的大部分空间。
(4)伊朗蒿、薄皮木和皱叶醉鱼草组成的亚高山灌丛草原: 包括被排在第1轴左上角的样方73-78, 主要分布区在下游加查县一带河谷流域。
(5)分布于雅鲁藏布江下游段的次生林和林缘灌丛草原植被: 主要优势种有高山松(Pinus densata )、绢毛蔷薇、毛叶绣线菊(Spiraea mollifolia )和川西千里光等, 在CCA图上主要被排在第1轴最左上角的样方85-90。
从CCA排序结果(图2 和表3 )可以看出, CCA排序轴1的特征值总和是0.60, 物种与环境因子的相关系数约是0.97, 排序轴1解释了物种总数据的7.3%的变化。而排序轴2的特征值总和是0.44, 物种与环境因子的相关系数是0.90, 与排序轴1一起累计解释了物种组成与环境因子之间关系的12.7%。这些数据信息表明, CCA排序第1轴和第2轴是最能反映植被类型变化的环境梯度。第1排序轴与海拔、经度的相关性最大, 其相关系数分别为0.94和-0.94(表3 和图2 ), 第1轴与海拔显著正相关, 而与经度成显著负相关; 其次是温度, 相关系数为-0.92, 为显著负相关。而第2排序轴主要与植被盖度成显著正相关, 其相关系数为0.70; 其次, 降雨量也与第2轴正相关关系, 相关系数为0.51。
表4 列出了所调查的5个环境因子之间的相关系数, 其中海拔与经度、温度和降雨量都成显著负相关, 相关系数分别为-0.99、-0.87和-0.78, 而经度与降雨量和温度成显著正相关, 相关系数分别为0.75和0.89。这些数据分析说明海拔和经度梯度的变化是影响研究区内水、热环境条件的主要因素。另外, 降雨量与温度也成显著正相关(相关系数为0.59)。
3.2.3 物种丰富度分布格局
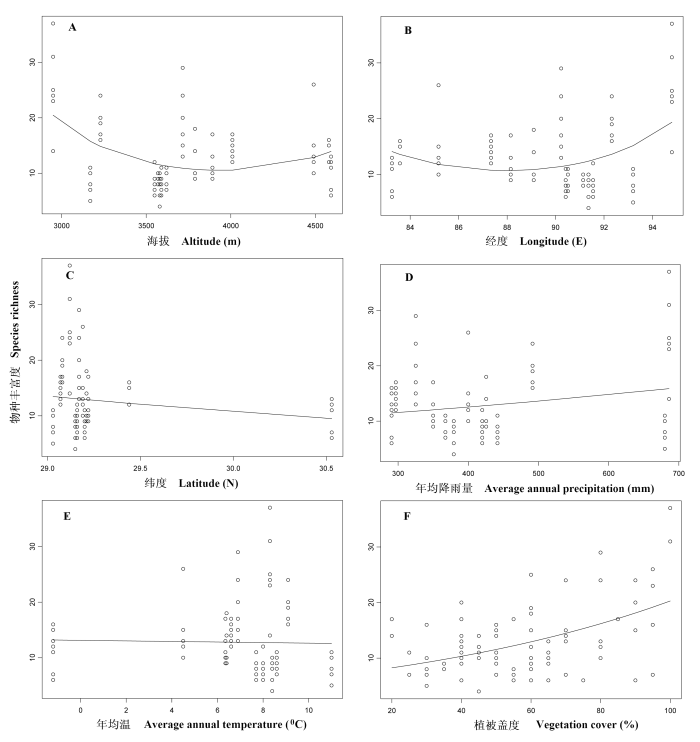
通过回归分析发现, 雅鲁藏布江河岸植物物种丰富度与海拔显著相关, 其分布格局大致呈浅“U”形分布(图3A , P < 0.001)。表现出物种丰富度在下游最高、中游偏低、上游略上升的趋势。物种丰富度在低海拔的米林地区(2,950 m)最高, 但在海拔3,170 m(朗县)处急剧降低, 后在海拔3,230 m一带(加查县)又有所升高, 在海拔3,550-3,590 m的中游区物种丰富度普遍偏低, 随后随海拔升高而又略有升高。物种丰富度与经度成极显著相关(图3B , P < 0.001), 其格局与海拔梯度相似, 也呈浅“U”型分布格局, 也表现出整个流域中间低、两头偏高的特点。物种丰富度与纬度相关不显著(图3C , P > 0.05)。物种丰富度与年均降雨量呈显著相关(图3D , P < 0.05), 但与年均温度的相关性不显著(图3E , P > 0.05)。另外, 物种丰富度与植被盖度极显著正相关(图3F , P < 0.001)。
图3 雅鲁藏布江中下游河岸植物物种丰富度与海拔(A)、经度(B)、纬度(C)、年均降雨量(D)、年均温(E)和植被盖度(F)之间的关系。最优拟合线都是根据一次曲线广义线性模型建立, 拟合线A、B和F极显著相关(P < 0.001), 拟合线D显著相关(P < 0.05), 拟合线C和E相关不显著。
Fig. 3 Relationships among species richness and altitude (A), longitude (B), latitude (C), average annual precipitation (D), average annual temperature (E) and vegetation cover (F). The fitted line is based on first-order polynomial generalized linear models. Lines A, B, F are highly significant (P < 0.001), line D is significant (P < 0.05), and line C and E are not significant.
4 讨论
4.1 物种组成特征和植被类型
除趋势对应分析(DCA)是基于间接分析物种组成连续体的理论(Hill & Cauch, 1980 ), 通过二维空间上的物种排序, 有效地揭示物种空间分布格局与环境影响因子的关系。本研究DCA物种排序结果表明, 雅鲁藏布江流域河岸植物物种分布格局主要受海拔梯度和经度梯度影响, 由海拔和经度变化引起的降雨量、温度条件变化可能是决定整个流域河岸植物群落物种组成发生变化的主要原因。5个环境因子的相关分析(表4 )也表明, 海拔和经度的变化引起降雨量和温度的显著变化, 从而可能影响物种空间上的分布变化。DCA物种空间排序可明显地反映出从低海拔至高海拔(伴随相对高经度至相对低经度)的物种组成空间排列和变化。从物种组成特征来看, 在上游地区, 随着海拔升高, 像高山嵩草、紫花针茅、青藏苔草和垫状点地梅(Androsace tapete )、藏布红景天等高山植物成分增多, 反映出随着海拔升高物种特有度在增加。有研究表明, 在厄瓜多尔地区, 随着海拔的升高物种特有度也在逐渐增加(Kessler, 2002 )。雅鲁藏布江中游区主要由适应干旱、半干旱气候和偏沙性土壤的灌丛草原植物类型组成, 而下游米林县附近植物组成明显反映出林缘植物组成特征。
CCA排序结果较好地划分了植被的5种不同类型, 沿海拔和经度梯度较清晰地依次划分出高寒草甸或高寒草原、高山草原、两种亚高山灌丛草原植被和林缘灌丛或次生林植被类型。第1排序轴的海拔由低到高的梯度变化正好是经度由高到低的梯度变化过程(图2 ), 两者相耦联, 并互为显著负相关(表4 , 相关系数-0.99), 这与雅鲁藏布江从西向东流向的地形地貌特征相吻合。CCA排序结果表明, 由海拔和经度变化引起的降雨量和温度条件变化(图2 , 表3 )可能控制着整个流域植被类型的形成与分布。张新时(1991)对西藏阿里地区植物群落的分析也表明, 植被类型分布主要取决于热量和湿度梯度。其他相关研究也表明, 与植被类型变化相关联的最重要的环境梯度是海拔、水分和温度(娄安如, 1998 ; 杨小林等, 2010 ; La et al ., 2010 )。
4.2 物种丰富度分布格局
植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 )。本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响。但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低。表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系。一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 )。而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低。因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 )。
本研究中的雅鲁藏布江中游区(行政区域上包括从桑日县至曲水县)是山地灌丛草原植被, 砂生槐是其植物群落中最重要的建群种(赵文智, 1998 )。在浅“U”型海拔梯度分布格局中该区域属于物种最贫乏区段。其可能的原因包括: 一是跟该区域气候条件有关, 中游区气候以半干旱为特点(沈渭寿, 1997 ), 降水少、多风且土壤蒸发量大(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 只比较适合砂生槐、藏沙蒿等耐旱植物生长; 二是由于样方中砂生槐等优势灌木植被盖度太高, 抑制了草本层植物对光照和其他营养资源的利用, 造成该区域样方内物种丰富度十分贫乏(图3A , 海拔3,550-3,590 m范围内), 物种最少的样方内只有6个物种(如样方67、68等), 其他研究也指出优势种对样方内物种多样性可以产生强烈影响(方精云等, 2004 ), 那些植被盖度很高的样方中的物种丰富度较低可能是物种之间对光照竞争的结果(Grace & Pugesek, 1997 )。
与中游区相比, 上游高寒草甸或草原群落尽管处于高海拔、高寒生境中, 但由于其样方内没有明显占优势的灌木植物而使物种丰富度相对较高。下游地区由于水、热等生境条件优越(如海拔2,950 m的米林县境内)表现出物种丰富度最高, 样方内物种最高可达37个(如样方84), 表明水、热是影响物种丰富度的最重要因子。物种丰富度与经度梯度的浅“U”型分布格局(图3B )与上述海拔梯度分布格局特征十分吻合, 其成因机制也应该相同, 因为河流从西到东海拔逐渐降低的过程就是经度逐渐升高的过程。尽管物种丰富度一般随纬度的升高而降低是最普遍的格局(Rohde, 1992 ), 但本研究中此关系不显著, 这主要与雅鲁藏布江流域的纬度梯度跨度不大有关, 整个流域纬度大致处在同一水平上。
物种丰富度与年均降雨量总体上成显著正相关(图3D ), 热带地区的研究也发现这种关系(Gentry, 1982 )。但是在本研究中, 物种丰富度除了在降雨量接近700 mm的最下游区急剧上升(图3D )外, 其余范围内物种丰富度随降雨量增加而升高的趋势并不十分明显, 甚至在某些范围内反而有降低趋势, 如雨量高达680 mm的朗县境内物种丰富度反而十分贫乏, 这种格局的出现可能也与该局地小生境干扰少、优势种盖度大等有紧密关系。反映出局地小生境确实对物种丰富度产生很大影响, 有时甚至可能超过了气候因子的作用。
另外, 本研究中物种丰富度与年均温度关系不显著。一般来讲, 海拔越高, 温度越低, 物种多样性应更低(唐志尧和方精云, 2004 ), 但在本研究中, 即使温度随海拔升高而显著降低(相关系数- 0.87), 也没有发现物种丰富度下降的普遍趋势。这可能也与研究区整个调查样点的纬度梯度小有一定关系, 即纬度引起的温度变化不足以影响物种丰富度的变化。一般来讲, 在更大的环境尺度上温度和水分是影响物种多样性分布格局的重要因素(Whittaker et al ., 2001 ), 但在小尺度上, 如前所述, 物种丰富度更容易受局地生境的影响, 可能在不同尺度影响物种丰富度分布格局的环境因子有较大的差异(Godfray & Lawton, 2001 )。对于长期适应于青藏高原高寒环境的植物来讲, 温度限制作用可能并没有水分那么重要。随着海拔的升高, 物种特有度一般会增加(唐志尧和方精云, 2004 ), 如上游区出现很多耐寒高山特有植物, 如高山嵩草、紫花针茅、垫状点地梅、青藏苔草和藏布红景天等, 这些物种长期适应于高寒环境, 因此其物种丰富度并没有随温度降低而显著下降。
物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系。其中单峰分布格局是最普遍的一种(Grime, 1997 )。但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 )。本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F )。方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关。植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏。如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高。但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低。如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 )。
由于雅鲁藏布江流域东西地理跨度很大, 其河岸植被类型和物种丰富度分布格局在大的尺度上, 除了受环境梯度(主要指海拔和经度)引起的水、热条件变化的综合影响外, 在小尺度上(样方尺度)可能更多地受特定小生境和局地生物作用(如干扰、物种竞争)的影响。因此, 在分析物种丰富度分布格局及其环境解释时, 除了在大尺度上要考虑气候是第一影响因子外, 还应注意在小尺度上其他生态机制的影响, 因为两者有可能同时存在于同一格局机制中(Liu et al ., 2007 )并发挥作用。另外, 本研究还没有完全覆盖到雅鲁藏布江最下游物种最丰富的雅鲁藏布江大拐弯地区, 今后还需补充这一区域的调查数据。
(责任编委: 唐志尧 责任编辑: 周玉荣)
陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义。因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 )。
雅鲁藏布江是世界上海拔最高的大河(何萍等, 2005 )。作为青藏高原上的最重要的河流, 掌握雅鲁藏布江流域植物多样性分布格局对研究青藏高原生物多样性, 确保青藏高原主体作为我国重要生态屏障有着十分重要的科学意义。目前, 有关该区域的植物多样性研究除了涉及局部区域之外, 整个雅鲁藏布江流域河岸的植物物种丰富度分布格局研究还较为缺乏。本研究沿海拔梯度从上游至下游, 对雅鲁藏布江流域河岸谷底植被和维管束植物组成进行了野外样方调查, 揭示了该区域的植物物种丰富度分布格局, 并分析了其与环境因子的关系。本研究结果将丰富这一区域植物多样性的研究, 并可为该流域的植物多样性评估和保护提供科学依据。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Acker SA Gregory SV Lienkaemper GMW Swanson FJ Miller SD 2003 ) Composition, complexity, and tree mortality in riparian forests in the central western Cascades of Oregon.
Forest Ecology and Management , 173 , 293 -308 .
[本文引用: 1]
[2]
Braak CJFT 1986 ) Canonical correspondence analysis: a new eigenvector method for multivariate direct gradient analysis.
Ecology , 67 , 1167 -1179 .
[本文引用: 1]
[3]
Comprehensive Scientific Expedition Team of Chinese Academy of Sciences in Qinghai-Tibet Plateau (中国科学院青藏高原综合科学考察队 ) (1988 ) Tibetan Vegetation (西藏植被 ). Science Press , Beijing . (in Chinese)
[本文引用: 1]
[4]
Connell JH 1978 ) Diversity in tropical rain forests and coral reefs.
Science , 199 , 1302 -1310 .
[本文引用: 1]
[5]
Currie DJ 1991 ) Energy and large-scale patterns of animal- and plant-species richness.
The American Naturalist , 137 , 27 -49 .
[本文引用: 1]
[6]
Fang JY 方精云 ), Kanza KM 神崎护 ), Wang XP 王襄平 ), Yoda K 依田恭二 ), Sun SZ 孙世洲 ), Shimota K 下田胜久 ) (2004 ) Community structure of alpine sparse vegetation and effects of microtopography in Pushila, Everest-Choyu region, Tibet, China.
Biodiversity Science (生物多样性 ), 12 , 190 -199 . (in Chinese with English abstract)
[本文引用: 1]
[7]
Gaston KJ 2000 ) Global patterns in biodiversity.
Nature , 405 , 220 -227 .
[本文引用: 2]
[8]
Gentry A 1982 ) Patterns of neotropical plants species diversity.
Evolutionary Biology , 15 , 1 -84 .
[本文引用: 1]
[9]
Godfray HCJ Lawton JH 2001 ) Scale and species richness numbers.
Trends in Ecology and Evolution , 16 , 400 -404 .
[本文引用: 1]
[10]
Gotelli NJ Colwell RK 2001 ) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness.
Ecology Letters , 4 , 379 -391 .
[本文引用: 1]
[11]
Grace JB Pugesek BH 1997 ) A structural equation model of plant species richness and its application to coastal wetland.
The American Naturalist , 149 , 436 -460 .
[本文引用: 2]
[12]
Grime JP 1997 ) The humped-back model: a response to Oksanen.
Journal of Ecology , 85 , 97 -98 .
[本文引用: 2]
[13]
Grytnes JA 2000 ) Fine-scale vascular plant species richness in different alpine vegetation types: relationships with biomass and cover.
Journal of Vegetation Science , 11 , 87 -92 .
[本文引用: 2]
[14]
Grytnes JA 2003 ) Species-richness patterns of vascular plants along seven altitudinal transects in Norway.
Ecography , 26 , 291 -300 .
[本文引用: 2]
[15]
Guo Q Berry WL 1998 ) Species richness and biomass: dissection of the hump-shaped relationships.
Ecology , 79 , 2555 -2559 .
[本文引用: 1]
[16]
Halbritter AH Alexander JM Edwards PJ Billeter R 2013 ) How comparable are species distributions along elevational and latitudinal climate gradients?
Global Ecology and Biogeography , 22 , 1228 -1237 .
[本文引用: 1]
[17]
He P 何萍 ), Guo K 郭柯 ), Gao JX 高吉喜 ), Shi PJ 史培军 ), Zhang YZ 张永泽 ), Zhuang HX 庄红翔 ) (2005 ) Vegetation types and their geographic distribution in the source area of the Yarlung Zangbo.
Journal of Mountain Science (山地学报 ), 23 , 267 -273 . (in Chinese with English abstract)
[本文引用: 1]
[18]
Hill MO Cauch HG 1980 ) Detrended Correspondence Analysis: an improved ordination technique.
Vegetatio , 42 , 47 -58 .
[本文引用: 2]
[19]
Jiménez-Alfaro B Marcenó C Bueno Á Gavilán R Obeso JR 2014 ) Biogeographic deconstruction of alpine plant communities along altitudinal and topographic gradients.
Journal of Vegetation Science , 25 , 160 -171 .
[本文引用: 1]
[20]
Kesslere M 2002 ) The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels.
Journal of Biogeography , 29 , 1159 -1165 .
[本文引用: 1]
[21]
Kraft NJ Comita LS Chase JM Sanders NJ Swenson NG Crist TO Myers JA 2011 ) Disentangling the drivers of β diversity along latitudinal and elevational gradients.
Science , 333 , 1755 -1758 .
[本文引用: 1]
[22]
Kratochwil A 1999 )Biodiversity in Ecosystems: Principles and Case Studies of Different Complexity Levels. Kluwer Academic Publishers
, Dordrecht.
[本文引用: 1]
[23]
Körner C 2007 ) The use of ‘altitude’ in ecological research.
Trends in Ecology and Evolution , 22 , 569 -574 .
[本文引用: 1]
[24]
Kuglerová L Jansson R Ågren A Laudon H Malm-Renöfält B 2014 ) Groundwater discharge creates hotspots of riparian plant species richness in a boreal forest stream network.
Ecology , 95 , 715 -722 .
[本文引用: 1]
[25]
La Q Grytnes J-A Birks HJB 2010 ) Alpine vegetation and species richness patterns along two altitudinal gradients in the Gyama Valley, south-central Tibet, China.
Plant Ecology and Diversity , 3 , 235 -247 .
[本文引用: 3]
[26]
Lepš J Šmilauer P 2003 ) Multivariate Analysis of Ecological Data Using CANOCO . Cambridge University Press , Lon- don, UK .
[本文引用: 1]
[27]
Li HD 李海东 ), Shen WS 沈渭寿 ), Fang Y 方颖 ), Yan SG 燕守广 ), Zhang H 张慧 ), Zhao W 赵卫 ) (2011 ) Point pattern analysis of several psammophyte population in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet, China.
Chinese Journal of Plant Ecology (植物生态学报 ), 35 , 834 -843 . (in Chinese with English abstract)
[本文引用: 1]
[28]
Liu Y Zhang YP He DM Cao M Zhu H 2007 ) Climatic control of plant species richness along elevation gradients in the Longitudinal Range-Gorge Region.
Chinese Science Bulletin , 52 (Suppl. ), 50 -58 .
[本文引用: 1]
[29]
Lou AR 娄安如 ) (1998 ) Ecological gradient analysis and environmental interpretation of mountain vegetation in the middle stretch of Tianshan Mountian.
Acta Phytoecologica Sinica (植物生态学报 ), 22 , 364 -372 . (in Chinese with English abstract)
[本文引用: 1]
[30]
MacArthur RH 1972 ) Geographical Ecology: Patterns of the Distribution of Species. Harper & Row
, New York.
[本文引用: 1]
[31]
McCullagh P Nelder NJ 1989 ) Generalised Linear Models, 2nd edn
. Chapman & Hall, London.
[本文引用: 1]
[32]
Miyamoto Y Nakano T Hattori M Nara K 2014 ) The mid-domain effect in ectomycorrhizal fungi: range overlap along an elevation gradient on Mount Fuji, Japan.
The ISME Journal, doi: 10.1038/ismej.2014.34
[本文引用: 1]
[33]
Nogués-Bravo D Araújo MB Romdal T Rahbek C 2008 ) Scale effects and human impact on the elevational species richness gradients.
Nature , 453 , 216 -219 .
[本文引用: 1]
[34]
O’Donnell J Fryirs K Leishman MR 2014 ) Digging deep for diversity: riparian seed bank abundance and species richness in relation to burial depth.
Freshwater Biology , 59 , 100 -113 .
[本文引用: 1]
[35]
Qian H Ricklefs RE 2007 ) A latitudinal gradient in large-scale beta diversity for vascular plants in North America.
Ecology Letters , 10 , 737 -744 .
[本文引用: 2]
[36]
Rahbek C 2005 ) The role of spatial scale and the perception of large-scale species-richness patterns.
Ecology Letters , 8 , 224 -239 .
[本文引用: 2]
[37]
Rohde K 1992 ) Latitudinal gradients in species diversity: the search for the primary cause.
Oikos , 65 , 514 -527 .
[本文引用: 2]
[38]
Sabo JL Sponseller R Dixon M Gade K Harms T Heffernan J Welter J 2005 ) Riparian zones increase regional species richness by harboring different, not more, species.
Ecology , 86 , 56 -62 .
[本文引用: 1]
[39]
Shen WS 沈渭寿 )(1997 ) Classification and sorting of the psammophytic vegetation in the middle Yarlung Zangbo River basin, Tibet.
Journal of Desert Research (中国沙漠 ), 17 , 169 -173 . (in Chinese with English abstract)
[本文引用: 1]
[40]
Szava-Kovats RC Ronk A Pärtel M 2013 ) Pattern without bias: local-regional richness relationship revisited.
Ecology , 94 , 1986 -1992 .
[本文引用: 1]
[41]
Tang ZY 唐志尧 ), Fang JY 方精云 ) (2004 ) A review on the elevational patterns of plant species diversity.
Biodiversity Science (生物多样性 ), 12 , 20 -28 . (in Chinese with English abstract)
[本文引用: 3]
[42]
Visser V Clayton WD Simpson DA Freckleton RP Osborne CP 2014 ) Mechanisms driving an unusual latitudinal diversity gradient for grasses.
Global Ecology and Biogeography , 23 , 61 -75 .
[本文引用: 1]
[43]
Waide RB Willig MR Steiner CF Mittelbach G Gough L Dodson SI Parmenter R 1999 ) The relationship between productivity and species richness.
Annual Review of Ecology and Systematics , 30 , 257 -300 .
[本文引用: 1]
[44]
Walter H 1979 ) Vegetation of the Earth and Ecological Systems of the Geo-biosphere. Springer
, New York.
[本文引用: 1]
[45]
Whittaker RJ Willis KJ Field R 2001 ) Scale and species richness: towards a general, hierarchical theory of species diversity.
Journal of Biogeography , 28 , 453 -470 .
[本文引用: 3]
[46]
Willig MR Kaufman DM Stevens RD 2003 ) Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis.
Annual Review of Ecology, Evolution, and Systematics , 34 , 273 -309 .
[本文引用: 1]
[47]
Wu GL 吴甘霖 ), Huang MY 黄敏毅 ), Duan RY 段仁燕 ), Zhao K 赵凯 ) (2006 ) Disturbing effects of tourism on species diversity in Pinus twaiwanensis communities.
Acta Ecologica Sinica (生态学报 ), 16 , 3924 -3929 . (in Chinese with English abstract)
[本文引用: 1]
[48]
Yang XL 杨小林 ), Zhao KT 赵垦田 ), Ma HP 马和平 ), Lu SH 禄树晖 ), Luo J 罗健 ) (2010 ) Ecological studies on vegetation quantity in the semi-arid valley region of Lasa.
Scientia Silvae Sinicae (林业科学 ), 46 , 15 -22 . (in Chinese with English abstract)
[本文引用: 1]
[49]
Yu SL 于顺利 ), Ma KP 马克平 ), Xu CB 徐存宝 ), Jin SF 金淑芳 ), Song XB 宋晓兵 ), Chen LZ 陈灵芝 ) (2004 ) The species diversity characteristics comparison of Quercus mongolica community along environmental gradient factors.
Acta Ecologica Sinica (生态学报 ), 24 , 2933 -2934 . (in Chinese with English abstract)
[本文引用: 2]
[50]
Zhang DC 张大才 ), Sun H 孙航 ) (2009 ) Research advances in altitudinal gradient distribution pattern of plant species richness at a broad spatial scale.
Journal of Southwest Forestry University (西南林学院学报 ), 29 (2 ), 74 -80 . (in Chinese with English abstract)
[本文引用: 1]
[51]
Zhang XS 张新时 ) (1991 ) Indirect gradient analysis, quantitative classification and environmental interpretation of plant communities in Ngari, Xizang (Tibet).
Acta Phytoecologica et Geobotanica Sinica (植物生态学与地植物学报 ), 15 ,101 -113 . (in Chinese with English abstract)
[52]
Zhao WZ 赵文智 ) (1998 ) A preliminary study of the arenaceous adaptability of Sophora moorcroftiana.
Acta Phytoecologica Sinica (植物生态学报 ), 22 , 379 -384 . (in Chinese with English abstract)
[本文引用: 1]
Composition, complexity, and tree mortality in riparian forests in the central western Cascades of Oregon.
1
2003
... 陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义.因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 ). ...
Canonical correspondence analysis: a new eigenvector method for multivariate direct gradient analysis.
1
1986
... 排序的方法能够比较客观地反映植物群落物种分布格局、群落类型及群落与生态环境之间的关系.排序一般分为间接排序和直接排序, 间接排序即在特定的环境轴上探讨物种的变化情况, 分析和推测其可能的环境梯度, 而直接排序用潜在的环境梯度或已测定环境因子数据来解释物种或植被类型变化(Lepš & Šmilauer, 2003 ).因此, 本研究先采用间接排序方法, 即除趋势对应分析方法(Detrended Correspondence Analysis, DCA)(Hill & Cauch, 1980 )对整个样方物种组成进行分析, 揭示所调查植物群落物种组成的空间变化, 并分析影响其格局的可能环境因子.在此基础上, 进一步采用直接排序方法, 即典范对应分析方法(Canonical Correspondence Analysis, CCA), 用本研究直接测得的环境数据, 进一步对物种数据进行分析, 确认植被类型空间分布格局与具体测定的生态环境因子之间是否存在显著相关性, 即海拔、经度、纬度、各海拔样带年均降雨量和年均温度和植被空间分布格局之间关系.CCA排序特别适合用来分析植物群落与具体环境因子之间关系(Braak, 1986 ). ...
1
1988
... 雅鲁藏布江源自于喜马拉雅山北麓的杰马央宗冰川, 全长约3,350 km, 中国境内长达2,052 km(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 由西向东沿喜马拉雅山脉贯穿整个藏南河谷, 在东部绕过喜马拉雅山脉最东端的南迦巴瓦峰转向印度洋.从源头到东部大峡谷, 地质地貌复杂独特, 海拔高差明显, 气候变化多样.上游呈高原宽谷地貌, 谷底海拔约4,500-4,800 m; 中游河谷宽、狭相间, 谷底海拔为3,500-4,400 m; 而下游山高谷深, 水流湍急, 植物组成十分复杂多样.整个雅鲁藏布江流域包括了干旱、半干旱半湿润、湿润3种气候特征(赵鲁青, 2011)① .流域内气温和降水量由西向东随海拔的降低而呈升高或增加趋势.本研究区内雅鲁藏布江下游地区年平均气温可达约11℃以上, 而在上游地区只有约-1℃左右(表1 ).整个流域内植被类型丰富, 上游谷底属于高寒草原和草甸, 而中游广大河谷区是喜温的半干旱草原和落叶阔叶灌丛草原, 下游逐渐向复杂的森林植被类型过渡(中国科学院青藏高原综合科学考察队, 1988 ). ...
Diversity in tropical rain forests and coral reefs.
1
1978
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
Energy and large-scale patterns of animal- and plant-species richness.
1
1991
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Community structure of alpine sparse vegetation and effects of microtopography in Pushila, Everest-Choyu region, Tibet, China.
1
2004
... 本研究中的雅鲁藏布江中游区(行政区域上包括从桑日县至曲水县)是山地灌丛草原植被, 砂生槐是其植物群落中最重要的建群种(赵文智, 1998 ).在浅“U”型海拔梯度分布格局中该区域属于物种最贫乏区段.其可能的原因包括: 一是跟该区域气候条件有关, 中游区气候以半干旱为特点(沈渭寿, 1997 ), 降水少、多风且土壤蒸发量大(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 只比较适合砂生槐、藏沙蒿等耐旱植物生长; 二是由于样方中砂生槐等优势灌木植被盖度太高, 抑制了草本层植物对光照和其他营养资源的利用, 造成该区域样方内物种丰富度十分贫乏(图3A , 海拔3,550-3,590 m范围内), 物种最少的样方内只有6个物种(如样方67、68等), 其他研究也指出优势种对样方内物种多样性可以产生强烈影响(方精云等, 2004 ), 那些植被盖度很高的样方中的物种丰富度较低可能是物种之间对光照竞争的结果(Grace & Pugesek, 1997 ). ...
Global patterns in biodiversity.
2
2000
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Patterns of neotropical plants species diversity.
1
1982
... 物种丰富度与年均降雨量总体上成显著正相关(图3D ), 热带地区的研究也发现这种关系(Gentry, 1982 ).但是在本研究中, 物种丰富度除了在降雨量接近700 mm的最下游区急剧上升(图3D )外, 其余范围内物种丰富度随降雨量增加而升高的趋势并不十分明显, 甚至在某些范围内反而有降低趋势, 如雨量高达680 mm的朗县境内物种丰富度反而十分贫乏, 这种格局的出现可能也与该局地小生境干扰少、优势种盖度大等有紧密关系.反映出局地小生境确实对物种丰富度产生很大影响, 有时甚至可能超过了气候因子的作用. ...
Scale and species richness numbers.
1
2001
... 另外, 本研究中物种丰富度与年均温度关系不显著.一般来讲, 海拔越高, 温度越低, 物种多样性应更低(唐志尧和方精云, 2004 ), 但在本研究中, 即使温度随海拔升高而显著降低(相关系数- 0.87), 也没有发现物种丰富度下降的普遍趋势.这可能也与研究区整个调查样点的纬度梯度小有一定关系, 即纬度引起的温度变化不足以影响物种丰富度的变化.一般来讲, 在更大的环境尺度上温度和水分是影响物种多样性分布格局的重要因素(Whittaker et al ., 2001 ), 但在小尺度上, 如前所述, 物种丰富度更容易受局地生境的影响, 可能在不同尺度影响物种丰富度分布格局的环境因子有较大的差异(Godfray & Lawton, 2001 ).对于长期适应于青藏高原高寒环境的植物来讲, 温度限制作用可能并没有水分那么重要.随着海拔的升高, 物种特有度一般会增加(唐志尧和方精云, 2004 ), 如上游区出现很多耐寒高山特有植物, 如高山嵩草、紫花针茅、垫状点地梅、青藏苔草和藏布红景天等, 这些物种长期适应于高寒环境, 因此其物种丰富度并没有随温度降低而显著下降. ...
Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness.
1
2001
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
A structural equation model of plant species richness and its application to coastal wetland.
2
1997
... 本研究中的雅鲁藏布江中游区(行政区域上包括从桑日县至曲水县)是山地灌丛草原植被, 砂生槐是其植物群落中最重要的建群种(赵文智, 1998 ).在浅“U”型海拔梯度分布格局中该区域属于物种最贫乏区段.其可能的原因包括: 一是跟该区域气候条件有关, 中游区气候以半干旱为特点(沈渭寿, 1997 ), 降水少、多风且土壤蒸发量大(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 只比较适合砂生槐、藏沙蒿等耐旱植物生长; 二是由于样方中砂生槐等优势灌木植被盖度太高, 抑制了草本层植物对光照和其他营养资源的利用, 造成该区域样方内物种丰富度十分贫乏(图3A , 海拔3,550-3,590 m范围内), 物种最少的样方内只有6个物种(如样方67、68等), 其他研究也指出优势种对样方内物种多样性可以产生强烈影响(方精云等, 2004 ), 那些植被盖度很高的样方中的物种丰富度较低可能是物种之间对光照竞争的结果(Grace & Pugesek, 1997 ). ...
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
The humped-back model: a response to Oksanen.
2
1997
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
... ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
Fine-scale vascular plant species richness in different alpine vegetation types: relationships with biomass and cover.
2
2000
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
... ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
Species-richness patterns of vascular plants along seven altitudinal transects in Norway.
2
2003
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Species richness and biomass: dissection of the hump-shaped relationships.
1
1998
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
How comparable are species distributions along elevational and latitudinal climate gradients?
1
2013
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Vegetation types and their geographic distribution in the source area of the Yarlung Zangbo.
1
2005
... 雅鲁藏布江是世界上海拔最高的大河(何萍等, 2005 ).作为青藏高原上的最重要的河流, 掌握雅鲁藏布江流域植物多样性分布格局对研究青藏高原生物多样性, 确保青藏高原主体作为我国重要生态屏障有着十分重要的科学意义.目前, 有关该区域的植物多样性研究除了涉及局部区域之外, 整个雅鲁藏布江流域河岸的植物物种丰富度分布格局研究还较为缺乏.本研究沿海拔梯度从上游至下游, 对雅鲁藏布江流域河岸谷底植被和维管束植物组成进行了野外样方调查, 揭示了该区域的植物物种丰富度分布格局, 并分析了其与环境因子的关系.本研究结果将丰富这一区域植物多样性的研究, 并可为该流域的植物多样性评估和保护提供科学依据. ...
Detrended Correspondence Analysis: an improved ordination technique.
2
1980
... 排序的方法能够比较客观地反映植物群落物种分布格局、群落类型及群落与生态环境之间的关系.排序一般分为间接排序和直接排序, 间接排序即在特定的环境轴上探讨物种的变化情况, 分析和推测其可能的环境梯度, 而直接排序用潜在的环境梯度或已测定环境因子数据来解释物种或植被类型变化(Lepš & Šmilauer, 2003 ).因此, 本研究先采用间接排序方法, 即除趋势对应分析方法(Detrended Correspondence Analysis, DCA)(Hill & Cauch, 1980 )对整个样方物种组成进行分析, 揭示所调查植物群落物种组成的空间变化, 并分析影响其格局的可能环境因子.在此基础上, 进一步采用直接排序方法, 即典范对应分析方法(Canonical Correspondence Analysis, CCA), 用本研究直接测得的环境数据, 进一步对物种数据进行分析, 确认植被类型空间分布格局与具体测定的生态环境因子之间是否存在显著相关性, 即海拔、经度、纬度、各海拔样带年均降雨量和年均温度和植被空间分布格局之间关系.CCA排序特别适合用来分析植物群落与具体环境因子之间关系(Braak, 1986 ). ...
... 除趋势对应分析(DCA)是基于间接分析物种组成连续体的理论(Hill & Cauch, 1980 ), 通过二维空间上的物种排序, 有效地揭示物种空间分布格局与环境影响因子的关系.本研究DCA物种排序结果表明, 雅鲁藏布江流域河岸植物物种分布格局主要受海拔梯度和经度梯度影响, 由海拔和经度变化引起的降雨量、温度条件变化可能是决定整个流域河岸植物群落物种组成发生变化的主要原因.5个环境因子的相关分析(表4 )也表明, 海拔和经度的变化引起降雨量和温度的显著变化, 从而可能影响物种空间上的分布变化.DCA物种空间排序可明显地反映出从低海拔至高海拔(伴随相对高经度至相对低经度)的物种组成空间排列和变化.从物种组成特征来看, 在上游地区, 随着海拔升高, 像高山嵩草、紫花针茅、青藏苔草和垫状点地梅(Androsace tapete )、藏布红景天等高山植物成分增多, 反映出随着海拔升高物种特有度在增加.有研究表明, 在厄瓜多尔地区, 随着海拔的升高物种特有度也在逐渐增加(Kessler, 2002 ).雅鲁藏布江中游区主要由适应干旱、半干旱气候和偏沙性土壤的灌丛草原植物类型组成, 而下游米林县附近植物组成明显反映出林缘植物组成特征. ...
Biogeographic deconstruction of alpine plant communities along altitudinal and topographic gradients.
1
2014
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels.
1
2002
... 除趋势对应分析(DCA)是基于间接分析物种组成连续体的理论(Hill & Cauch, 1980 ), 通过二维空间上的物种排序, 有效地揭示物种空间分布格局与环境影响因子的关系.本研究DCA物种排序结果表明, 雅鲁藏布江流域河岸植物物种分布格局主要受海拔梯度和经度梯度影响, 由海拔和经度变化引起的降雨量、温度条件变化可能是决定整个流域河岸植物群落物种组成发生变化的主要原因.5个环境因子的相关分析(表4 )也表明, 海拔和经度的变化引起降雨量和温度的显著变化, 从而可能影响物种空间上的分布变化.DCA物种空间排序可明显地反映出从低海拔至高海拔(伴随相对高经度至相对低经度)的物种组成空间排列和变化.从物种组成特征来看, 在上游地区, 随着海拔升高, 像高山嵩草、紫花针茅、青藏苔草和垫状点地梅(Androsace tapete )、藏布红景天等高山植物成分增多, 反映出随着海拔升高物种特有度在增加.有研究表明, 在厄瓜多尔地区, 随着海拔的升高物种特有度也在逐渐增加(Kessler, 2002 ).雅鲁藏布江中游区主要由适应干旱、半干旱气候和偏沙性土壤的灌丛草原植物类型组成, 而下游米林县附近植物组成明显反映出林缘植物组成特征. ...
Disentangling the drivers of β diversity along latitudinal and elevational gradients.
1
2011
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Biodiversity in Ecosystems: Principles and Case Studies of Different Complexity Levels. Kluwer Academic Publishers
1
1999
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
The use of ‘altitude’ in ecological research.
1
2007
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Groundwater discharge creates hotspots of riparian plant species richness in a boreal forest stream network.
1
2014
... 陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义.因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 ). ...
Alpine vegetation and species richness patterns along two altitudinal gradients in the Gyama Valley, south-central Tibet, China.
3
2010
... CCA排序结果较好地划分了植被的5种不同类型, 沿海拔和经度梯度较清晰地依次划分出高寒草甸或高寒草原、高山草原、两种亚高山灌丛草原植被和林缘灌丛或次生林植被类型.第1排序轴的海拔由低到高的梯度变化正好是经度由高到低的梯度变化过程(图2 ), 两者相耦联, 并互为显著负相关(表4 , 相关系数-0.99), 这与雅鲁藏布江从西向东流向的地形地貌特征相吻合.CCA排序结果表明, 由海拔和经度变化引起的降雨量和温度条件变化(图2 , 表3 )可能控制着整个流域植被类型的形成与分布.张新时(1991)对西藏阿里地区植物群落的分析也表明, 植被类型分布主要取决于热量和湿度梯度.其他相关研究也表明, 与植被类型变化相关联的最重要的环境梯度是海拔、水分和温度(娄安如, 1998 ; 杨小林等, 2010 ; La et al ., 2010 ). ...
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
1
2003
... 排序的方法能够比较客观地反映植物群落物种分布格局、群落类型及群落与生态环境之间的关系.排序一般分为间接排序和直接排序, 间接排序即在特定的环境轴上探讨物种的变化情况, 分析和推测其可能的环境梯度, 而直接排序用潜在的环境梯度或已测定环境因子数据来解释物种或植被类型变化(Lepš & Šmilauer, 2003 ).因此, 本研究先采用间接排序方法, 即除趋势对应分析方法(Detrended Correspondence Analysis, DCA)(Hill & Cauch, 1980 )对整个样方物种组成进行分析, 揭示所调查植物群落物种组成的空间变化, 并分析影响其格局的可能环境因子.在此基础上, 进一步采用直接排序方法, 即典范对应分析方法(Canonical Correspondence Analysis, CCA), 用本研究直接测得的环境数据, 进一步对物种数据进行分析, 确认植被类型空间分布格局与具体测定的生态环境因子之间是否存在显著相关性, 即海拔、经度、纬度、各海拔样带年均降雨量和年均温度和植被空间分布格局之间关系.CCA排序特别适合用来分析植物群落与具体环境因子之间关系(Braak, 1986 ). ...
Point pattern analysis of several psammophyte population in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet, China.
1
2011
... 陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义.因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 ). ...
Climatic control of plant species richness along elevation gradients in the Longitudinal Range-Gorge Region.
1
2007
... 由于雅鲁藏布江流域东西地理跨度很大, 其河岸植被类型和物种丰富度分布格局在大的尺度上, 除了受环境梯度(主要指海拔和经度)引起的水、热条件变化的综合影响外, 在小尺度上(样方尺度)可能更多地受特定小生境和局地生物作用(如干扰、物种竞争)的影响.因此, 在分析物种丰富度分布格局及其环境解释时, 除了在大尺度上要考虑气候是第一影响因子外, 还应注意在小尺度上其他生态机制的影响, 因为两者有可能同时存在于同一格局机制中(Liu et al ., 2007 )并发挥作用.另外, 本研究还没有完全覆盖到雅鲁藏布江最下游物种最丰富的雅鲁藏布江大拐弯地区, 今后还需补充这一区域的调查数据. ...
Ecological gradient analysis and environmental interpretation of mountain vegetation in the middle stretch of Tianshan Mountian.
1
1998
... CCA排序结果较好地划分了植被的5种不同类型, 沿海拔和经度梯度较清晰地依次划分出高寒草甸或高寒草原、高山草原、两种亚高山灌丛草原植被和林缘灌丛或次生林植被类型.第1排序轴的海拔由低到高的梯度变化正好是经度由高到低的梯度变化过程(图2 ), 两者相耦联, 并互为显著负相关(表4 , 相关系数-0.99), 这与雅鲁藏布江从西向东流向的地形地貌特征相吻合.CCA排序结果表明, 由海拔和经度变化引起的降雨量和温度条件变化(图2 , 表3 )可能控制着整个流域植被类型的形成与分布.张新时(1991)对西藏阿里地区植物群落的分析也表明, 植被类型分布主要取决于热量和湿度梯度.其他相关研究也表明, 与植被类型变化相关联的最重要的环境梯度是海拔、水分和温度(娄安如, 1998 ; 杨小林等, 2010 ; La et al ., 2010 ). ...
Geographical Ecology: Patterns of the Distribution of Species. Harper & Row
1
1972
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Generalised Linear Models, 2nd edn
1
1989
... 为了分析物种丰富度分布格局与环境因子之间关系, 我们采用广义线性回归模型(Generalized Linear Model, GLM)(McCullagh & Nelder, 1989 )进行回归分析, 研究物种丰富度分布格局与海拔、经度、纬度、年均降雨量、年均温度和植被盖度之间的关系. ...
The mid-domain effect in ectomycorrhizal fungi: range overlap along an elevation gradient on Mount Fuji, Japan.
1
2014
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Scale effects and human impact on the elevational species richness gradients.
1
2008
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Digging deep for diversity: riparian seed bank abundance and species richness in relation to burial depth.
1
2014
... 陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义.因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 ). ...
A latitudinal gradient in large-scale beta diversity for vascular plants in North America.
2
2007
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
The role of spatial scale and the perception of large-scale species-richness patterns.
2
2005
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
Latitudinal gradients in species diversity: the search for the primary cause.
2
1992
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... 与中游区相比, 上游高寒草甸或草原群落尽管处于高海拔、高寒生境中, 但由于其样方内没有明显占优势的灌木植物而使物种丰富度相对较高.下游地区由于水、热等生境条件优越(如海拔2,950 m的米林县境内)表现出物种丰富度最高, 样方内物种最高可达37个(如样方84), 表明水、热是影响物种丰富度的最重要因子.物种丰富度与经度梯度的浅“U”型分布格局(图3B )与上述海拔梯度分布格局特征十分吻合, 其成因机制也应该相同, 因为河流从西到东海拔逐渐降低的过程就是经度逐渐升高的过程.尽管物种丰富度一般随纬度的升高而降低是最普遍的格局(Rohde, 1992 ), 但本研究中此关系不显著, 这主要与雅鲁藏布江流域的纬度梯度跨度不大有关, 整个流域纬度大致处在同一水平上. ...
Riparian zones increase regional species richness by harboring different, not more, species.
1
2005
... 陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区(李海东等, 2011 ), 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义.因此, 近年来有关河岸植物多样性与环境之间的关系引起了很多生态学家的兴趣(Acker et al ., 2003 ; Sabo et al ., 2005 ; Kuglerová et al ., 2014 ; O'Donnell et al ., 2014 ). ...
Classification and sorting of the psammophytic vegetation in the middle Yarlung Zangbo River basin, Tibet.
1
1997
... 本研究中的雅鲁藏布江中游区(行政区域上包括从桑日县至曲水县)是山地灌丛草原植被, 砂生槐是其植物群落中最重要的建群种(赵文智, 1998 ).在浅“U”型海拔梯度分布格局中该区域属于物种最贫乏区段.其可能的原因包括: 一是跟该区域气候条件有关, 中游区气候以半干旱为特点(沈渭寿, 1997 ), 降水少、多风且土壤蒸发量大(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 只比较适合砂生槐、藏沙蒿等耐旱植物生长; 二是由于样方中砂生槐等优势灌木植被盖度太高, 抑制了草本层植物对光照和其他营养资源的利用, 造成该区域样方内物种丰富度十分贫乏(图3A , 海拔3,550-3,590 m范围内), 物种最少的样方内只有6个物种(如样方67、68等), 其他研究也指出优势种对样方内物种多样性可以产生强烈影响(方精云等, 2004 ), 那些植被盖度很高的样方中的物种丰富度较低可能是物种之间对光照竞争的结果(Grace & Pugesek, 1997 ). ...
Pattern without bias: local-regional richness relationship revisited.
1
2013
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
A review on the elevational patterns of plant species diversity.
3
2004
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... 另外, 本研究中物种丰富度与年均温度关系不显著.一般来讲, 海拔越高, 温度越低, 物种多样性应更低(唐志尧和方精云, 2004 ), 但在本研究中, 即使温度随海拔升高而显著降低(相关系数- 0.87), 也没有发现物种丰富度下降的普遍趋势.这可能也与研究区整个调查样点的纬度梯度小有一定关系, 即纬度引起的温度变化不足以影响物种丰富度的变化.一般来讲, 在更大的环境尺度上温度和水分是影响物种多样性分布格局的重要因素(Whittaker et al ., 2001 ), 但在小尺度上, 如前所述, 物种丰富度更容易受局地生境的影响, 可能在不同尺度影响物种丰富度分布格局的环境因子有较大的差异(Godfray & Lawton, 2001 ).对于长期适应于青藏高原高寒环境的植物来讲, 温度限制作用可能并没有水分那么重要.随着海拔的升高, 物种特有度一般会增加(唐志尧和方精云, 2004 ), 如上游区出现很多耐寒高山特有植物, 如高山嵩草、紫花针茅、垫状点地梅、青藏苔草和藏布红景天等, 这些物种长期适应于高寒环境, 因此其物种丰富度并没有随温度降低而显著下降. ...
... ).对于长期适应于青藏高原高寒环境的植物来讲, 温度限制作用可能并没有水分那么重要.随着海拔的升高, 物种特有度一般会增加(唐志尧和方精云, 2004 ), 如上游区出现很多耐寒高山特有植物, 如高山嵩草、紫花针茅、垫状点地梅、青藏苔草和藏布红景天等, 这些物种长期适应于高寒环境, 因此其物种丰富度并没有随温度降低而显著下降. ...
Mechanisms driving an unusual latitudinal diversity gradient for grasses.
1
2014
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
The relationship between productivity and species richness.
1
1999
... 物种丰富度与生物生产力关系已在很多生物多样性研究中被报道(Guo & Berry, 1998 ; Grime, 1997 ; Waide, 1999 ), 也发现了很多不同类型的关系.其中单峰分布格局是最普遍的一种(Grime, 1997 ).但是, 在相关研究中直接测定生产力的方法并不多见, 而更多的是用生物量或植被盖度来替代生产力(Grytnes, 2000 ; La et al ., 2010 ), 植被盖度有时比生物量能更好地揭示物种丰富度与生产力的关系(Grytnes, 2000 ).本研究中我们用植被盖度来分析物种丰富度分布格局, 结果显示, 物种丰富度与植被盖度呈显著正相关关系, 当植被盖度接近100%时物种丰富度达到最高(图3F ).方精云等(2004)在西藏珠峰地区山地植被研究中也发现, 随着植被盖度的增加, 转换物种和转换属的丰富度增加, 他们认为这种格局的出现可能与生境质量有关.植被盖度是各种环境因子综合作用于植物的表现, 因此能够反映局地生境质量的好坏.如本研究中的最下游地区由于水热条件优越, 植被盖度高, 物种丰富度也最高.但是, 如果生境内高植被盖度是由某些单一优势物种所决定, 则物种丰富度反而会降低.如上游的高山嵩草和中游的砂生槐等优势种的盖度大约超过80%时, 样方内物种多样性就明显降低, 说明物种丰富度受物种之间竞争作用的影响(Grace & Pugesek, 1997 ). ...
Vegetation of the Earth and Ecological Systems of the Geo-biosphere. Springer
1
1979
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Scale and species richness: towards a general, hierarchical theory of species diversity.
3
2001
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
... 另外, 本研究中物种丰富度与年均温度关系不显著.一般来讲, 海拔越高, 温度越低, 物种多样性应更低(唐志尧和方精云, 2004 ), 但在本研究中, 即使温度随海拔升高而显著降低(相关系数- 0.87), 也没有发现物种丰富度下降的普遍趋势.这可能也与研究区整个调查样点的纬度梯度小有一定关系, 即纬度引起的温度变化不足以影响物种丰富度的变化.一般来讲, 在更大的环境尺度上温度和水分是影响物种多样性分布格局的重要因素(Whittaker et al ., 2001 ), 但在小尺度上, 如前所述, 物种丰富度更容易受局地生境的影响, 可能在不同尺度影响物种丰富度分布格局的环境因子有较大的差异(Godfray & Lawton, 2001 ).对于长期适应于青藏高原高寒环境的植物来讲, 温度限制作用可能并没有水分那么重要.随着海拔的升高, 物种特有度一般会增加(唐志尧和方精云, 2004 ), 如上游区出现很多耐寒高山特有植物, 如高山嵩草、紫花针茅、垫状点地梅、青藏苔草和藏布红景天等, 这些物种长期适应于高寒环境, 因此其物种丰富度并没有随温度降低而显著下降. ...
Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis.
1
2003
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Disturbing effects of tourism on species diversity in Pinus twaiwanensis communities.
1
2006
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
Ecological studies on vegetation quantity in the semi-arid valley region of Lasa.
1
2010
... CCA排序结果较好地划分了植被的5种不同类型, 沿海拔和经度梯度较清晰地依次划分出高寒草甸或高寒草原、高山草原、两种亚高山灌丛草原植被和林缘灌丛或次生林植被类型.第1排序轴的海拔由低到高的梯度变化正好是经度由高到低的梯度变化过程(图2 ), 两者相耦联, 并互为显著负相关(表4 , 相关系数-0.99), 这与雅鲁藏布江从西向东流向的地形地貌特征相吻合.CCA排序结果表明, 由海拔和经度变化引起的降雨量和温度条件变化(图2 , 表3 )可能控制着整个流域植被类型的形成与分布.张新时(1991)对西藏阿里地区植物群落的分析也表明, 植被类型分布主要取决于热量和湿度梯度.其他相关研究也表明, 与植被类型变化相关联的最重要的环境梯度是海拔、水分和温度(娄安如, 1998 ; 杨小林等, 2010 ; La et al ., 2010 ). ...
The species diversity characteristics comparison of Quercus mongolica community along environmental gradient factors.
2
2004
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
... 植物物种丰富度沿海拔梯度分布格局常常因不同研究地域而呈现出不同结果(Rahbek, 2005 ).本研究结果表明, 雅鲁藏布江河岸植物物种丰富度随海拔梯度大致呈浅“U”型分布格局, 相似的格局在拉萨河上游山地植物物种丰富度研究中也被发现(La et al. , 2010 ), 即物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为主要与山地灌木植物盖度有关, 半山腰高密度灌木使物种丰富度受到影响.但本研究的浅“U”型物种海拔梯度分布格局在一些具体区段上并不是十分符合, 如下游(低海拔)朗县一带6个样方的物种丰富度反而很低.表明整个格局的形成除了受海拔梯度引起的水、热条件变化影响外, 还可能跟局地小环境特征有密切关系.一般来讲在小尺度或样方尺度上, 物种丰富度的分布格局更容易受小生境和局地生物因子作用(如光照、生物竞争和干扰史等)的影响(Whittaker et al ., 2001 ; 于顺利等, 2004 ).而上述6个样方位于下游区朗县境内的受大面积保护的灌丛草原围栏内, 人畜干扰较少, 促进了样方内个别优势种的进一步发展(如薄皮木和砂生槐等), 可能抑制了其他小草本植物的生长, 造成物种丰富度的降低.因为适度的干扰或中度干扰有利于物种丰富度的提高(Connell, 1978 ; 吴甘霖等, 2006 ). ...
Research advances in altitudinal gradient distribution pattern of plant species richness at a broad spatial scale.
1
2009
... 物种丰富度的分布格局及其成因机制一直是生态学研究的重要内容(MacArthur, 1972 ; Currie, 1991 ; Gotelli & Colwell, 2001 ; Visser, 2014 ), 特别是一定环境梯度下物种丰富度的分布格局是其研究热点议题之一(Kratochwil, 1999 ; Willig et al ., 2003 ; Körner, 2007 ; Szava-Kovats et al ., 2013 ), 生态学家在全球各种不同环境梯度和空间尺度上进行研究, 发现了各种不同类型的物种丰富度分布格局(Gaston, 2000 ; Grytnes, 2003 ; Qian & Ricklefs, 2007 ; Miyamoto et al ., 2014 ).影响物种多样性分布格局的主要自然环境因素包括海拔、纬度、经度、地形、土壤湿度等(于顺利等, 2004 ; Kraft et al ., 2011 ; Jiménez-Alfaro et al ., 2014 ).由于海拔和纬度梯度包含了水、热和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面(Gaston, 2000 ), 特别是在海拔梯度下, 水、热因子可在相对较小范围内急剧变化(Walter, 1979 ; 唐志尧和方精云, 2004 ).在有关物种海拔梯度分布格局研究中单峰和单调下降型分布格局是最常见的两种类型(Grytnes, 2003 ), 约有一半的研究结果呈现单峰分布格局, 其次是单调下降型分布格局(Rahbek, 2005 ).影响这些不同分布格局的因素往往因研究尺度不同而异(Whittaker et al ., 2001 ; Nogués-Bravo et al ., 2008 ; 张大才等, 2009 ).除了海拔梯度分布格局外, 纬度梯度分布格局也是生物多样性分布的重要特点, 但是气候因子在较小纬度梯度内变化远没有海拔梯度那么剧烈(Halbritter et al ., 2013 ).一般情况下, 物种丰富度随纬度升高而呈降低趋势(Rohde, 1992 ), 但也有峰值出现在中纬度, 甚至无明显梯度变化的现象(Qian & Ricklefs, 2007 ).由于经度梯度引起的气候因子差异没有海拔和纬度梯度那么明显, 所以有关经度梯度下物种丰富度分布格局研究并不多见. ...
Indirect gradient analysis, quantitative classification and environmental interpretation of plant communities in Ngari, Xizang (Tibet).
1991
A preliminary study of the arenaceous adaptability of Sophora moorcroftiana.
1
1998
... 本研究中的雅鲁藏布江中游区(行政区域上包括从桑日县至曲水县)是山地灌丛草原植被, 砂生槐是其植物群落中最重要的建群种(赵文智, 1998 ).在浅“U”型海拔梯度分布格局中该区域属于物种最贫乏区段.其可能的原因包括: 一是跟该区域气候条件有关, 中游区气候以半干旱为特点(沈渭寿, 1997 ), 降水少、多风且土壤蒸发量大(赵鲁青, 2011)(① 赵鲁青 (2011) 雅鲁藏布江中下游区域植被绿期和净初级生产力时空格局及其对气候变化的响应. 硕士论文, 华东师范大学资源与环境科学学院, 上海.), 只比较适合砂生槐、藏沙蒿等耐旱植物生长; 二是由于样方中砂生槐等优势灌木植被盖度太高, 抑制了草本层植物对光照和其他营养资源的利用, 造成该区域样方内物种丰富度十分贫乏(图3A , 海拔3,550-3,590 m范围内), 物种最少的样方内只有6个物种(如样方67、68等), 其他研究也指出优势种对样方内物种多样性可以产生强烈影响(方精云等, 2004 ), 那些植被盖度很高的样方中的物种丰富度较低可能是物种之间对光照竞争的结果(Grace & Pugesek, 1997 ). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}