作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 )。外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 )。预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 )。Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统。
麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 )。麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 )。鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 )。鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 )。最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 )。但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 )。
麦穗鱼和鲫的入侵在世界范围内对生态系统和社会经济都造成了深远的影响。麦穗鱼被认为是世界上最具入侵性的鱼类之一, 它在人类活动的推波助澜下自1960年以来的50年间, 已经入侵到32个国家(Gozlan et al ., 2010 )。麦穗鱼入侵到新的生态系统后, 不仅与当地土著鱼类竞争, 更由于它所携带的传染性病原体还对当地的生态系统造成严重的损害。Gozlan等(2005)的研究揭示麦穗鱼入侵欧洲后, 它携带的病原体导致了当地的濒危鱼类小赤梢鱼(Leucaspius delineatus )死亡率增加, 繁殖遭到强烈抑制。鲫在引进到澳大利亚西部后, 迅速繁殖形成优势种群; 它所携带的病原体, 以及其生活习性改变了当地生态系统的结构与功能(David et al ., 2004 )。我们在西藏雅鲁藏布江的调查也发现, 鲫引入该地区后已经能够自然繁衍并建立种群, 成为当地数量最多、分布最广的外来鱼类(陈锋和陈毅峰, 2010 )。因此, 针对麦穗鱼和鲫这两种具有在世界范围内扩散能力的入侵种, 通过技术手段预测其潜在的分布区, 从而进行早期预警, 是十分必要的。
Maxent是一种根据已知物种分布信息预测该物种潜在分布区域的模型, 广泛应用于生物地理学、生物入侵以及生态位研究等领域(Phillips et al ., 2006 ; Suarez-Seoane et al ., 2008 )。该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法, 即“最大熵原则(the maximum-entropy principle)”, 可以更多地保留物种已有分布数据的有用信息。Giovanelli等(2008 )利用Maxent模型分析了原产于北美的牛蛙(Lithobates catesbeianus )在巴西的入侵范围, 并预测了牛蛙在巴西亚马逊地区和东部的大西洋沿岸的潜在入侵区域。与其他方法相比, Maxent模型所具备的优势使其预测表现始终是最好的(Elith et al ., 2006 )。综合以上情况, 本研究采用Maxent模型对麦穗鱼和鲫在全球的适生区进行了预测, 以期为相关国家和地区对外来鱼类的预警和风险评估提供科学依据。
1 材料与方法
1.1 地理分布数据的收集与整理
麦穗鱼和鲫的分布点数据通过实验室多年野外采样调查和查询FishBase数据库(Froese & Pauly, 2011 , http://www.fishbase.org )两种方法获得。麦穗鱼共有1,503条分布信息, 鲫有376条分布信息。其中麦穗鱼的159个样点和鲫的199个样点来自我们对我国的东北地区、黄河流域、长江流域、云南高原和西藏高原的采样调查; 而麦穗鱼另外的1,344个分布点以及鲫的138个分布点由FishBase下载获得, 此外鲫还采用了我国文献记录的39个样点数据。从FishBase下载的分布点范围除我国以外, 更多地涵盖了世界其他国家和地区。数据使用前首先进行筛选并删除重复分布数据, 在ArcGIS中核对。
1.2 环境数据与地图数据
本研究所采用的环境因子图层为气候数据图层, 来自WorldClim(Hijmans et al ., 2004 , http://www.worldclim.org )。该数据集共包含了24种环境气候数据, 分辨率高达2.5弧分(附录I)。其中, Bio1–Bio19这19种数据由50年(1950–2000年)全球气候数据提取而来, 并特意为预测生物的分布进行了优化, 具有更多的生物学意义, 可以更精确地预测生物的适生区。
本研究的地理底图选用1:10 m Natural Earth I (http://www.naturalearthdata.com )。该地图由北美制图信息学会(North American Cartographic Information Society, NACIS)所支持并绘制而成, 地图信息由卫星土地覆盖数据生成, 不仅包含了地形、水系、三角洲等丰富信息, 而且分辨率高达2.3亿像素, 可以使结果更加清晰精确。国界线参考国家测绘地理信息局提供的《世界地图》(2008版)。
1.3 统计分析
麦穗鱼和鲫的分布点信息在Excel中进行整理后连同环境数据一起导入Maxent。选择80%的样点作为训练数据, 20%的样点用于对结果进行检验。采用AUC检验模型进行准确性判别的原则: 如果数据是随机的, 那么AUC=0.5, 即对角线; 如果AUC曲线在对角线的上方, 说明结果的准确率较高; 如果AUC曲线在对角线下方, 说明与正确结果相反, 需取其反向结果。总之, AUC曲线越远离对角线的结果越可信, 越接近对角线结果则越不可信。用Jackknife检验哪些环境变量对麦穗鱼和鲫的分布起到重要的影响作用, 交叉验证(crossvalidate)重复运算10次, 以确使验证结果更加准确。
最后将Maxent预测结果导入ArcGIS 9.2中, 添加原始样点要素, 通过与地理底图的叠加, 能够更好地观察和分析麦穗鱼和鲫在各个水系的分布, 以及山脉等地形因素对其分布的影响。
2 结果
2.1 麦穗鱼和鲫的适生区
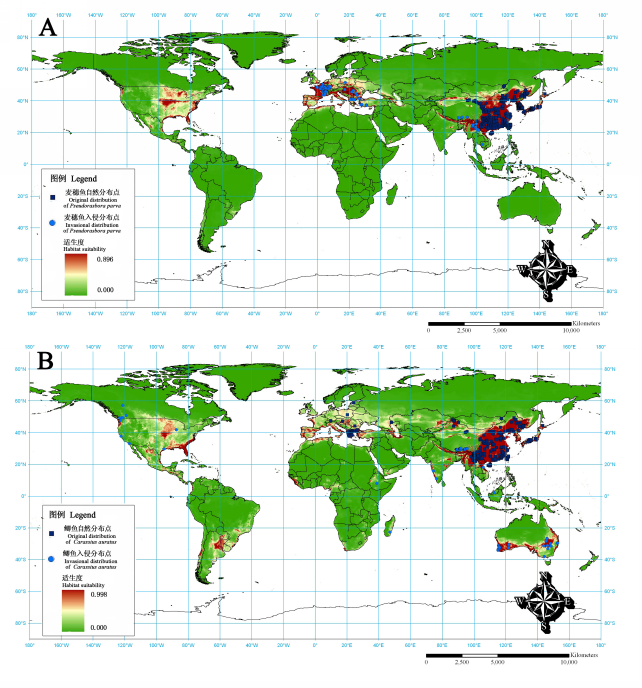
结果表明, 麦穗鱼和鲫在全球除南极洲外的各个大洲都有广泛的适生区(图1 )。由此分析除自然分布区外, 麦穗鱼潜在的入侵区域集中于亚洲、欧洲和北美洲。其中亚洲的潜在入侵区域有自土耳其至里海沿岸, 也门和阿曼的部分地区, 喜马拉雅山脉南麓的巴基斯坦北部、印度北部、尼泊尔和不丹, 我国雅鲁藏布江流域的中下游、怒江上游、塔里木河流域, 以及缅甸、柬埔寨、越南等东南亚国家; 在欧洲, 从其西部的英国、西班牙至东部的希腊、保加利亚均是其潜在的入侵区域, 最北可至55º N附近, 但不包括丹麦、立陶宛及其以北的欧洲国家; 在北美洲, 由于美国中部的密西西比河流域、五大湖、墨西哥湾沿岸和东部的大西洋沿岸有较高的适合度, 因此也是入侵风险性极高的地区, 而美国西北部的西雅图至加拿大温哥华附近区域, 墨西哥、危地马拉和洪都拉斯的部分地区也是其潜在的入侵区域。
图1 麦穗鱼和鲫全球潜在分布区预测结果。A: 麦穗鱼; B: 鲫。
Fig. 1 The results of the potential habitat prediction of Pseudorasbora parva (A) and Carassius auratus (B) in the world
与麦穗鱼相比, 鲫的自然分布区更广, 其潜在的入侵区域主要集中在亚洲、北美洲、南美洲以及大洋洲。在亚洲, 自土耳其至里海沿岸, 也门和阿曼的部分地区, 喜马拉雅山脉南麓的巴基斯坦北部、印度北部、尼泊尔、不丹和孟加拉国, 印度西部沿海和奥里萨邦附近地区, 缅甸、越南、泰国北部、菲律宾、印度尼西亚等东南亚地区; 在北美洲及南美洲, 美国境内的大部分地区, 墨西哥至委内瑞拉, 智利、阿根廷北部、乌拉圭和巴西, 而美国中部的密西西比河流域、墨西哥湾沿岸、佛罗里达州至北卡罗来纳州的大西洋沿岸、西雅图至加拿大温哥华附近的入侵风险性极高; 在大洋洲则包括了澳大利亚的南部和东部、新几内亚、新喀里多尼亚和新西兰的北部。此外, 北非地中海沿岸、塞内加尔和几内亚附近、尼日利亚和喀麦隆、南非、马达加斯加等非洲国家和地区也是鲫的潜在入侵区。
2.2 AUC检验
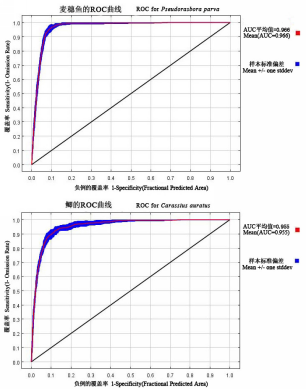
结果表明, 麦穗鱼和鲫的AUC曲线都位于图形的上方(图2 )。其中, 麦穗鱼的AUC=0.966, 而鲫的AUC=0.955, 说明本研究的结果有较高的可信度。
图2 麦穗鱼和鲫的AUC检验结果
Fig. 2 The results of AUC test for Pseudorasbora parva and Carassius auratus
2.3 Jackknife检验
本研究利用Jackknife来检验影响麦穗鱼和鲫自然分布相对重要的环境变量。结果表明, 3月最高温度(Tmax3)、2月最高温度(Tmax2)、11月最高温度(Tmax11)和高程(Alt)是影响麦穗鱼分布最重要的环境因子。而10月最高温度(Tmax10)、3月最高温度(Tmax3)和4月最高温度(Tmax4)是影响鲫分布的重要变量, 高程(Alt)、最冷季节平均温度(Bio19)、7月平均降水量(Prec7)分别在单独使用时具有最高增益, 被省略时对结果影响较大, 因此Tmax10、Tmax3、Tmax4、Alt、Bio19和Prec7是影响鲫分布最重要的环境变量。
3 讨论
本研究的预测结果表明, 麦穗鱼的适生区集中于15º–55º N之间, 而鲫在40º S–60º N。比较麦穗鱼已知的入侵分布与自然分布, 可以发现麦穗鱼从原产于东亚的我国、朝鲜、韩国和日本, 已经入侵到了欧洲的法国、罗马尼亚、希腊和土耳其等国家。我们推测是由于罗马尼亚在20世纪60年代从我国武汉引进草鱼和鲢鱼鱼卵时, 麦穗鱼混在其中偶然带入的, 之后随着其他欧洲国家通过从我国引种以及欧洲国家间相互引种等方式, 在欧洲广泛扩散(Gozlan et al ., 2010 )。麦穗鱼在我国云南的南盘江和金沙江都有自然分布, 但随着家鱼的引进而进一步扩散至洱海、滇池、抚仙湖、泸沽湖、拉市海和丘北普者黑等高原湖泊, 以及澜沧江、李仙江、藤条江和怒江(Chen & Cui, 1993 ; 潘勇等, 2006 ; 陈小勇, 2013 )。进一步比较麦穗鱼已知分布和预测适生区分布, 我们发现除已知分布的地区外, 北美洲的美国和墨西哥, 欧洲的西班牙和葡萄牙, 非洲北部的地中海沿岸麦穗鱼的生境适合度也较高, 预示了这些地区具有极高的入侵风险。
比较鲫已知的入侵分布与自然分布, 发现鲫的自然分布相对广泛, 横跨整个欧亚大陆。但是鲫在美国、加拿大, 非洲的埃塞俄比亚和马达加斯加, 澳大利亚, 印度、越南、马来西亚和我国西藏以及青海也有分布, 可以说其已入侵的区域也同样非常广泛。其原因一方面在于鲫曾以彩鲫和金鱼的形式作为世界最早的观赏鱼类, 以及随后又作为养殖品种在世界各国和地区进行的引种; 另一方面在于鲫对水体环境具有很强的适应能力。比较鲫的已知分布与预测适生区分布, 可以发现南美洲的阿根廷和巴西, 非洲西部的几内亚、喀麦隆等国家鲫的生境适合度较高, 是入侵风险极高的区域。从上述分析可以看出, 尽管麦穗鱼和鲫目前在世界范围内已经有了广泛的分布, 但是它们还将继续扩散至一个更大的范围。
考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 )。可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响。事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a )。此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 )。这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b )。我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现。
比较两种鱼类的Jackknife检验结果, 我们发现虽然影响两者分布的环境因子不尽相同, 但是除了影响鲫分布的7月平均降水量(Prec7)以外, 其余变量都属于与温度和海拔相关的环境因子。而在与温度相关的环境因子中, 只有最冷季节平均温度 (Bio19)不是某月的最高温。海拔对鱼类分布的影响, 一方面是由于山脉、高原等地形的阻隔作用; 另一方面海拔同样是影响温度的因子, 对分布造成间接的影响。温度是限制鱼类分布的重要因子, 鱼类的生长和繁殖都与温度密切相关(殷名称, 1995 )。因此, 影响麦穗鱼和鲫分布的最主要环境因子是温度和海拔, 而我们的研究结果还揭示了这两种鱼类对最高温度更加敏感。
通过本研究, 我们还发现利用Maxent模型来预测物种的潜在分布存在两个方面的不足。首先, 利用气候数据预测物种分布行之有效, 尤其是对物种宏观的大范围分布的预测有着较高的准确性。然而, 受地理环境数据精度和环境数据所涉及的内容的限制, 目前还不能精确到水塘、温泉、水库等小生境, 并且缺少流速、水温和盐度等水文数据, 因此将影响鱼类在小范围内分布的精度和准确性。其次, 该模型并不考虑物种间的相互作用, 这对其预测结果的准确性会有一定的影响。
致谢
感谢北京大学生命科学学院的闻丞老师在模型上提供的帮助。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[40]
Yin MC 殷名称 ) (1995 ) Fish Ecology (鱼类生态学). China Agriculture Press , Beijing . (in Chinese)
[本文引用: 1]
[41]
.Yue PQ 乐佩琦 ) (2000 ) Fauna Sinica · Osteichthyes ·
Cyprini-formes III (中国动物志・硬骨鱼纲・鲤形目(下卷)). Science Press, Beijing. (in Chinese)
[本文引用: 3]
[本文引用: 3]
[42]
Zhang YP 张亚平 ), Jiang YX 蒋有绪 ), Zhang RZ 张润志 ), Sang WG 桑卫国 ), Chen YF 陈毅峰 ), Xue DY 薛大勇 ), Yang JX 杨君兴 ), Peng H 彭华 ), Zhang KQ 张克勤 ) (2009 ) Biological invasion in China: current situation and countermeasures
. Bulletin of Chinese Academy of Sciences (中国科学院院刊 ), 24 , 411 –413 . (in Chinese)
[本文引用: 1]
[1]
Austin MP 2002 ) Spatial prediction of species distribution: an interface between ecological theory and statistical modelling
. Ecological Modelling , 157 , 101 –118 .
[本文引用: 1]
[2]
Bănărescu P 2002 ) The Freshwater Fishes of Europe. Cyprinidae 2, Part Ⅲ: Carassius to Cyprinus. AULA, Wiebelsheim
.
[本文引用: 1]
[3]
Berg LS 1949 ) Freshwater fishes of the USSR and adjacent countries
. Guide to the Fauna of USSR , 27 , 927 –1382 .
[本文引用: 1]
[4]
Britton JR Davies GD Brazier M 2009 ) Eradication of the invasive Pseudorasbora parva results in increased growth and production of native fishes
. Ecology of Freshwater Fish , 18 , 8 –14 .
[本文引用: 2]
[5]
Britton JR Davies GD Brazier M 2010 a) Towards the successful control of the invasive Pseudorasbora parva in the UK
. Biological Invasions , 12 , 125 –131 .
[本文引用: 1]
[6]
Britton JR Davies GD Harrod C 2010 b) Trophic interactions and consequent impacts of the invasive fish Pseudorasbora parva in a native aquatic foodweb: a field investigation in the UK
. Biological Invasions , 12 , 1533 –1542 .
[本文引用: 1]
[7]
Chen F 陈锋 ), Chen YF 陈毅峰 ) (2010 ) Investigation and protection strategies of fishes of Lhasa River
. Acta Hydrobiologica Sinica (水生生物学报 ), 34 , 278 –285 . (in Chinese with English abstract)
[本文引用: 1]
[8]
Chen PX 陈佩薰 ) (1959 ) Fish biological research of Carassius auratus in Liangzi Lake
. Acta Hydrobiologica Sinica (水生生物学集刊 ), (4 ), 411 –419 . (in Chinese)
[本文引用: 1]
[9]
Chen XY 陈小勇 ) (2013 ) Checklist of fishes of Yunnan
. Zoological Research (动物学研究 ), 34 , 281 –343 . (in Chinese with English abstract)
[本文引用: 1]
[10]
Chen YY Cui YB 1993 ) Some aspects of biological diversity in freshwater ecosystems
. Chinese Biodiversity , 1 (Suppl. ), 46 –49 .
[本文引用: 1]
[11]
Chen YY 陈宜瑜 ) (1998 ) Fauna Sinica · Osteichthyes ·Cypriniformes II (中国动物志・硬骨鱼纲・鲤形目 (中卷)). Science Press, Beijing. (in Chinese)
[本文引用: 1]
[本文引用: 1]
[12]
Cheng L 程磊 ), Chang YM 常玉梅 ), Lu CY 鲁翠云 ), Cao DC 曹顶臣 ), Sun XW 孙效文 ) (2012 ) DNA barcoding and species and subspecies classification within genus Carassius
. Zoological Research (动物学研究 ), 33 , 463 –472 . (in Chinese with English abstract)
[本文引用: 1]
[13]
Chu XL 禇新洛 ), Chen YR 陈银瑞 ) (1989 ) Fishes of Yunnan (云南鱼类志). Science Press , Beijing . (in Chinese)
[本文引用: 1]
[14]
David LM Howard SG Mark GM Stephen JB 2004 ) Distribution and impacts of introduced freshwater fishes in Western Australia
. New Zealand Journal of Marine and Freshwater Research , 38 , 511 –523 .
[本文引用: 1]
[15]
Dong CY 董存有 ) (1984 ) An observations of fish biology on Pseudorasbora parva
. Fisheries Science & Technology (水产科技 ), (1 ), 50 –53 . (in Chinese)
[本文引用: 1]
[16]
Elith J Graham CH Anderson R Dudík M Ferrier S Guisan A Hijmans RJ Huettmann F Leathwick JR Lehmann A Li J Lohmann LG Loiselle BA Manion G Moritz C Nakamura M Nakazawa Y Overton JMM Peterson AT Phillips SJ Richardson K Scachetti-Pereira R Schapire RE Soberón J Williams S Wisz MS Zimmermann NE 2006 ) Novel methods improve prediction of species’ distributions from occurrence data
. Ecography , 29 , 129 –151 .
[本文引用: 1]
[17]
Froese R Pauly D 2011 ) FishBase
. . (2013. 04. 10
URL
[本文引用: 1]
[18]
Giovanelli JGR Haddad CFB Alexandrino J 2008 ) Predicting the potential distribution of the alien invasive American bullfrog (Lithobates catesbeianus) in Brazil
. Biological Invasions , 10 , 585 –590 .
[本文引用: 1]
[19]
Gozlan RE Hilaire S Feist SW Martin P Kent ML 2005 ) Biodiversity: disease threat to European fish
. Nature , 435 , 1046 .
[本文引用: 1]
[20]
Gozlan RE Andreou D Asaeda T Beyer K Bouhadad R Burnard D Caiola N Cakic P Djikanovic V Esmaeili HR Falka I Golicher D Harka A Jeney G Kovac V Musil J Nocita A Povz M Poulet N Virbickas T Wolter C Tarkan AS Tricarico E Trichkova T Verreycken H Witkowski A Zhang CG Zweimueller I Britton JR 2010 ) Pan-contin- ental invasion of Pseudorasbora parva: towards a better understanding of freshwater fish invasions
. Fish and Fisheries , 11 , 315 –340 .
[本文引用: 3]
[21]
Hijmans R Cameron S Parra J Jones P Jarvis A 2004 ) The WorldClim interpolated global terrestrial climate surfaces. Version 1.3
. . (2013. 04. 10
URL
[本文引用: 1]
[22]
Humphrey JD Ashburner LD 1993 ) Spread of the bacterial fish pathogen Aeromonas salmonicida after importation of infected goldfish, Carassius auratus, into Australia
. Australian Veterinary Journal , 70 , 453 –454 .
[本文引用: 1]
[23]
Kalous L Bohlen J Rylková K Petrtýl M 2012 ) Hidden diversity within the Prussian carp and designation of a neotype for Carassius gibelio (Teleostei: Cyprinidae)
. Ichthyological Exploration of Freshwaters , 21 , 11 –18 .
[本文引用: 1]
[24]
Kim YU Myung JG Han KH Koh JR 1996 ) The fish fauna of Namdae Stream in Kangreung, Korea
. Journal of the Korean Fisheries Society , 29 , 262 –266 .
[本文引用: 1]
[25]
Kolar CS Lodge DM 2001 ) Progress in invasion biology: predicting invaders
. Trends in Ecology and Evolution , 16 , 199 –204 .
[本文引用: 1]
[26]
Kottelat M Freyhof J 2007 ) Handbook of European Fresh- water Fishes. Publications Kottelat, Cornol, Switzerland
.
[本文引用: 1]
[27]
Lockwood J Hoopes M Marchetti M 2009 ) Invasion Ecology. Wiley-Blackwell, Oxford
.
[本文引用: 1]
[28]
Lusková V Lusk S Halačka K Vetešnik L 2010 ) Carassius auratus gibelio: the most successful invasive fish in waters of the Czech Republic
. Russian Journal of Biological Invasions , 1 , 176 –180 .
[本文引用: 1]
[29]
Mack RN Simberloff D Lonsdale WM Evans H Clout M Bazzaz FA 2000 ) Biotic invasions: causes, epidemiology, global consequences, and control
. Ecological Applications , 10 , 689 –710 .
[本文引用: 1]
[30]
Onikura N Nakajima J 2013 ) Age, growth and habitat use of the topmouth gudgeon, Pseudorasbora parva in irrigation ditches on northwestern Kyushu Island, Japan
. Journal of Applied Ichthyology , 29 , 186 –192 .
[本文引用: 1]
[31]
Pan Y 潘勇 ), Cao WX 曹文宣 ), Xu LP 徐立蒲 ), Yin SR 殷守仁 ) (2006 ) History and approach of invasion from domestic and abroad fishes
. Journal of Dalian Fisheries University (大连水产学院学报 ), 21 , 72 –77 . (in Chinese with English abstract)
[本文引用: 1]
[32]
Phillips SJ Anderson RP Schapire RE 2006 ) Maximum entropy modeling of species geographic distributions
. Ecological Modelling , 190 , 231 –259 .
[本文引用: 1]
[33]
Sang WG 桑卫国 ), Zhu L 朱丽 ), Ma KP 马克平 ) (2006 ) Issues, phenomena and study emphases of alien species invasion in China
. Advances in Earth Science (地球科学进展 ), 21 , 305 –312 . (in Chinese with English abstract)
[本文引用: 1]
[34]
Suarez-Seoane S de la Morena ELG , Prieto MBM Osborne PE de Juana E 2008 ) Maximum entropy niche-based modelling of seasonal changes in little bustard (Tetrax tetrax) distribution
. Ecological Modelling , 219 , 17 –29 .
[本文引用: 1]
[35]
Thuiller W Richardson DM Pysek P Midgley GF Hughes GO Rouget M 2005 ) Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale
. Global Change Biology , 11 , 2234 –2250 .
[本文引用: 1]
[36]
Wu XW 伍献文 ) (1977 ) The Cyprinid Fishes of China , Vol. 2 (中国鲤科鱼类志). Shanghai Science & Technogy Press, Shanghai . (in Chinese)
[本文引用: 1]
[37]
Xie YH 解玉浩 ) (2007 ) Freshwater Fishes in Northeast Region of China (东北地区淡水鱼类). Liaoning Science and Technology Press , Shenyang . (in Chinese)
[本文引用: 1]
[38]
Xu RM 徐汝梅 ), Ye WH 叶万辉 ) (2003 ) Biological Invasions : Theory and Practice (生物入侵: 理论与实践). Science Press, Beijing . (in Chinese)
[本文引用: 1]
[39]
Yang RB 杨瑞斌 ), Bian SJ 边书京 ), Zhou J 周洁 ), Xie CX 谢从新 ) (2004 ) Study on food habits of Pseudorasbora parva in Liangzi Lake
. Journal of Huazhong Agricultural University (华中农业大学学报 ), 23 , 331 –334 . (in Chinese with English abstract)
[本文引用: 1]
1
1995
... 比较两种鱼类的Jackknife检验结果, 我们发现虽然影响两者分布的环境因子不尽相同, 但是除了影响鲫分布的7月平均降水量(Prec7)以外, 其余变量都属于与温度和海拔相关的环境因子.而在与温度相关的环境因子中, 只有最冷季节平均温度 (Bio19)不是某月的最高温.海拔对鱼类分布的影响, 一方面是由于山脉、高原等地形的阻隔作用; 另一方面海拔同样是影响温度的因子, 对分布造成间接的影响.温度是限制鱼类分布的重要因子, 鱼类的生长和繁殖都与温度密切相关(殷名称, 1995 ).因此, 影响麦穗鱼和鲫分布的最主要环境因子是温度和海拔, 而我们的研究结果还揭示了这两种鱼类对最高温度更加敏感. ...
Fauna Sinica · Osteichthyes ·
3
2000
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
... ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
... ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Fauna Sinica · Osteichthyes ·
3
2000
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
... ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
... ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Biological invasion in China: current situation and countermeasures
1
2009
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Spatial prediction of species distribution: an interface between ecological theory and statistical modelling
1
2002
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
The Freshwater Fishes of Europe. Cyprinidae 2, Part Ⅲ: Carassius to Cyprinus. AULA, Wiebelsheim
1
2002
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Freshwater fishes of the USSR and adjacent countries
1
1949
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Eradication of the invasive Pseudorasbora parva results in increased growth and production of native fishes
2
2009
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
... ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
a) Towards the successful control of the invasive Pseudorasbora parva in the UK
1
2010
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
b) Trophic interactions and consequent impacts of the invasive fish Pseudorasbora parva in a native aquatic foodweb: a field investigation in the UK
1
2010
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
Investigation and protection strategies of fishes of Lhasa River
1
2010
... 麦穗鱼和鲫的入侵在世界范围内对生态系统和社会经济都造成了深远的影响.麦穗鱼被认为是世界上最具入侵性的鱼类之一, 它在人类活动的推波助澜下自1960年以来的50年间, 已经入侵到32个国家(Gozlan et al ., 2010 ).麦穗鱼入侵到新的生态系统后, 不仅与当地土著鱼类竞争, 更由于它所携带的传染性病原体还对当地的生态系统造成严重的损害.Gozlan等(2005)的研究揭示麦穗鱼入侵欧洲后, 它携带的病原体导致了当地的濒危鱼类小赤梢鱼(Leucaspius delineatus )死亡率增加, 繁殖遭到强烈抑制.鲫在引进到澳大利亚西部后, 迅速繁殖形成优势种群; 它所携带的病原体, 以及其生活习性改变了当地生态系统的结构与功能(David et al ., 2004 ).我们在西藏雅鲁藏布江的调查也发现, 鲫引入该地区后已经能够自然繁衍并建立种群, 成为当地数量最多、分布最广的外来鱼类(陈锋和陈毅峰, 2010 ).因此, 针对麦穗鱼和鲫这两种具有在世界范围内扩散能力的入侵种, 通过技术手段预测其潜在的分布区, 从而进行早期预警, 是十分必要的. ...
Fish biological research of Carassius auratus in Liangzi Lake
1
1959
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Checklist of fishes of Yunnan
1
2013
... 本研究的预测结果表明, 麦穗鱼的适生区集中于15º–55º N之间, 而鲫在40º S–60º N.比较麦穗鱼已知的入侵分布与自然分布, 可以发现麦穗鱼从原产于东亚的我国、朝鲜、韩国和日本, 已经入侵到了欧洲的法国、罗马尼亚、希腊和土耳其等国家.我们推测是由于罗马尼亚在20世纪60年代从我国武汉引进草鱼和鲢鱼鱼卵时, 麦穗鱼混在其中偶然带入的, 之后随着其他欧洲国家通过从我国引种以及欧洲国家间相互引种等方式, 在欧洲广泛扩散(Gozlan et al ., 2010 ).麦穗鱼在我国云南的南盘江和金沙江都有自然分布, 但随着家鱼的引进而进一步扩散至洱海、滇池、抚仙湖、泸沽湖、拉市海和丘北普者黑等高原湖泊, 以及澜沧江、李仙江、藤条江和怒江(Chen & Cui, 1993 ; 潘勇等, 2006 ; 陈小勇, 2013 ).进一步比较麦穗鱼已知分布和预测适生区分布, 我们发现除已知分布的地区外, 北美洲的美国和墨西哥, 欧洲的西班牙和葡萄牙, 非洲北部的地中海沿岸麦穗鱼的生境适合度也较高, 预示了这些地区具有极高的入侵风险. ...
Some aspects of biological diversity in freshwater ecosystems
1
1993
... 本研究的预测结果表明, 麦穗鱼的适生区集中于15º–55º N之间, 而鲫在40º S–60º N.比较麦穗鱼已知的入侵分布与自然分布, 可以发现麦穗鱼从原产于东亚的我国、朝鲜、韩国和日本, 已经入侵到了欧洲的法国、罗马尼亚、希腊和土耳其等国家.我们推测是由于罗马尼亚在20世纪60年代从我国武汉引进草鱼和鲢鱼鱼卵时, 麦穗鱼混在其中偶然带入的, 之后随着其他欧洲国家通过从我国引种以及欧洲国家间相互引种等方式, 在欧洲广泛扩散(Gozlan et al ., 2010 ).麦穗鱼在我国云南的南盘江和金沙江都有自然分布, 但随着家鱼的引进而进一步扩散至洱海、滇池、抚仙湖、泸沽湖、拉市海和丘北普者黑等高原湖泊, 以及澜沧江、李仙江、藤条江和怒江(Chen & Cui, 1993 ; 潘勇等, 2006 ; 陈小勇, 2013 ).进一步比较麦穗鱼已知分布和预测适生区分布, 我们发现除已知分布的地区外, 北美洲的美国和墨西哥, 欧洲的西班牙和葡萄牙, 非洲北部的地中海沿岸麦穗鱼的生境适合度也较高, 预示了这些地区具有极高的入侵风险. ...
Fauna Sinica · Osteichthyes ·Cypriniformes II (中国动物志・硬骨鱼纲・鲤形目 (中卷)). Science Press, Beijing. (in Chinese)
1
1998
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Fauna Sinica · Osteichthyes ·Cypriniformes II (中国动物志・硬骨鱼纲・鲤形目 (中卷)). Science Press, Beijing. (in Chinese)
1
1998
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
DNA barcoding and species and subspecies classification within genus Carassius
1
2012
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
1
1989
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Distribution and impacts of introduced freshwater fishes in Western Australia
1
2004
... 麦穗鱼和鲫的入侵在世界范围内对生态系统和社会经济都造成了深远的影响.麦穗鱼被认为是世界上最具入侵性的鱼类之一, 它在人类活动的推波助澜下自1960年以来的50年间, 已经入侵到32个国家(Gozlan et al ., 2010 ).麦穗鱼入侵到新的生态系统后, 不仅与当地土著鱼类竞争, 更由于它所携带的传染性病原体还对当地的生态系统造成严重的损害.Gozlan等(2005)的研究揭示麦穗鱼入侵欧洲后, 它携带的病原体导致了当地的濒危鱼类小赤梢鱼(Leucaspius delineatus )死亡率增加, 繁殖遭到强烈抑制.鲫在引进到澳大利亚西部后, 迅速繁殖形成优势种群; 它所携带的病原体, 以及其生活习性改变了当地生态系统的结构与功能(David et al ., 2004 ).我们在西藏雅鲁藏布江的调查也发现, 鲫引入该地区后已经能够自然繁衍并建立种群, 成为当地数量最多、分布最广的外来鱼类(陈锋和陈毅峰, 2010 ).因此, 针对麦穗鱼和鲫这两种具有在世界范围内扩散能力的入侵种, 通过技术手段预测其潜在的分布区, 从而进行早期预警, 是十分必要的. ...
An observations of fish biology on Pseudorasbora parva
1
1984
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Novel methods improve prediction of species’ distributions from occurrence data
1
2006
... Maxent是一种根据已知物种分布信息预测该物种潜在分布区域的模型, 广泛应用于生物地理学、生物入侵以及生态位研究等领域(Phillips et al ., 2006 ; Suarez-Seoane et al ., 2008 ).该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法, 即“最大熵原则(the maximum-entropy principle)”, 可以更多地保留物种已有分布数据的有用信息.Giovanelli等(2008 )利用Maxent模型分析了原产于北美的牛蛙(Lithobates catesbeianus )在巴西的入侵范围, 并预测了牛蛙在巴西亚马逊地区和东部的大西洋沿岸的潜在入侵区域.与其他方法相比, Maxent模型所具备的优势使其预测表现始终是最好的(Elith et al ., 2006 ).综合以上情况, 本研究采用Maxent模型对麦穗鱼和鲫在全球的适生区进行了预测, 以期为相关国家和地区对外来鱼类的预警和风险评估提供科学依据. ...
FishBase
1
2011
... 麦穗鱼和鲫的分布点数据通过实验室多年野外采样调查和查询FishBase数据库(Froese & Pauly, 2011 , http://www.fishbase.org )两种方法获得.麦穗鱼共有1,503条分布信息, 鲫有376条分布信息.其中麦穗鱼的159个样点和鲫的199个样点来自我们对我国的东北地区、黄河流域、长江流域、云南高原和西藏高原的采样调查; 而麦穗鱼另外的1,344个分布点以及鲫的138个分布点由FishBase下载获得, 此外鲫还采用了我国文献记录的39个样点数据.从FishBase下载的分布点范围除我国以外, 更多地涵盖了世界其他国家和地区.数据使用前首先进行筛选并删除重复分布数据, 在ArcGIS中核对. ...
Predicting the potential distribution of the alien invasive American bullfrog (Lithobates catesbeianus) in Brazil
1
2008
... Maxent是一种根据已知物种分布信息预测该物种潜在分布区域的模型, 广泛应用于生物地理学、生物入侵以及生态位研究等领域(Phillips et al ., 2006 ; Suarez-Seoane et al ., 2008 ).该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法, 即“最大熵原则(the maximum-entropy principle)”, 可以更多地保留物种已有分布数据的有用信息.Giovanelli等(2008 )利用Maxent模型分析了原产于北美的牛蛙(Lithobates catesbeianus )在巴西的入侵范围, 并预测了牛蛙在巴西亚马逊地区和东部的大西洋沿岸的潜在入侵区域.与其他方法相比, Maxent模型所具备的优势使其预测表现始终是最好的(Elith et al ., 2006 ).综合以上情况, 本研究采用Maxent模型对麦穗鱼和鲫在全球的适生区进行了预测, 以期为相关国家和地区对外来鱼类的预警和风险评估提供科学依据. ...
Biodiversity: disease threat to European fish
1
2005
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
Pan-contin- ental invasion of Pseudorasbora parva: towards a better understanding of freshwater fish invasions
3
2010
... 麦穗鱼和鲫的入侵在世界范围内对生态系统和社会经济都造成了深远的影响.麦穗鱼被认为是世界上最具入侵性的鱼类之一, 它在人类活动的推波助澜下自1960年以来的50年间, 已经入侵到32个国家(Gozlan et al ., 2010 ).麦穗鱼入侵到新的生态系统后, 不仅与当地土著鱼类竞争, 更由于它所携带的传染性病原体还对当地的生态系统造成严重的损害.Gozlan等(2005)的研究揭示麦穗鱼入侵欧洲后, 它携带的病原体导致了当地的濒危鱼类小赤梢鱼(Leucaspius delineatus )死亡率增加, 繁殖遭到强烈抑制.鲫在引进到澳大利亚西部后, 迅速繁殖形成优势种群; 它所携带的病原体, 以及其生活习性改变了当地生态系统的结构与功能(David et al ., 2004 ).我们在西藏雅鲁藏布江的调查也发现, 鲫引入该地区后已经能够自然繁衍并建立种群, 成为当地数量最多、分布最广的外来鱼类(陈锋和陈毅峰, 2010 ).因此, 针对麦穗鱼和鲫这两种具有在世界范围内扩散能力的入侵种, 通过技术手段预测其潜在的分布区, 从而进行早期预警, 是十分必要的. ...
... 本研究的预测结果表明, 麦穗鱼的适生区集中于15º–55º N之间, 而鲫在40º S–60º N.比较麦穗鱼已知的入侵分布与自然分布, 可以发现麦穗鱼从原产于东亚的我国、朝鲜、韩国和日本, 已经入侵到了欧洲的法国、罗马尼亚、希腊和土耳其等国家.我们推测是由于罗马尼亚在20世纪60年代从我国武汉引进草鱼和鲢鱼鱼卵时, 麦穗鱼混在其中偶然带入的, 之后随着其他欧洲国家通过从我国引种以及欧洲国家间相互引种等方式, 在欧洲广泛扩散(Gozlan et al ., 2010 ).麦穗鱼在我国云南的南盘江和金沙江都有自然分布, 但随着家鱼的引进而进一步扩散至洱海、滇池、抚仙湖、泸沽湖、拉市海和丘北普者黑等高原湖泊, 以及澜沧江、李仙江、藤条江和怒江(Chen & Cui, 1993 ; 潘勇等, 2006 ; 陈小勇, 2013 ).进一步比较麦穗鱼已知分布和预测适生区分布, 我们发现除已知分布的地区外, 北美洲的美国和墨西哥, 欧洲的西班牙和葡萄牙, 非洲北部的地中海沿岸麦穗鱼的生境适合度也较高, 预示了这些地区具有极高的入侵风险. ...
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
The WorldClim interpolated global terrestrial climate surfaces. Version 1.3
1
2004
... 本研究所采用的环境因子图层为气候数据图层, 来自WorldClim(Hijmans et al ., 2004 , http://www.worldclim.org ).该数据集共包含了24种环境气候数据, 分辨率高达2.5弧分(附录I).其中, Bio1–Bio19这19种数据由50年(1950–2000年)全球气候数据提取而来, 并特意为预测生物的分布进行了优化, 具有更多的生物学意义, 可以更精确地预测生物的适生区. ...
Spread of the bacterial fish pathogen Aeromonas salmonicida after importation of infected goldfish, Carassius auratus, into Australia
1
1993
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
Hidden diversity within the Prussian carp and designation of a neotype for Carassius gibelio (Teleostei: Cyprinidae)
1
2012
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
The fish fauna of Namdae Stream in Kangreung, Korea
1
1996
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Progress in invasion biology: predicting invaders
1
2001
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Handbook of European Fresh- water Fishes. Publications Kottelat, Cornol, Switzerland
1
2007
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
Invasion Ecology. Wiley-Blackwell, Oxford
1
2009
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Carassius auratus gibelio: the most successful invasive fish in waters of the Czech Republic
1
2010
... 考虑到这两种鱼类会对当地水生生态系统产生巨大的入侵危害(Gozlan et al ., 2010 ; Lusková et al ., 2010 ).可以预见, 随着这两种鱼类的进一步扩散, 将会对全球水生生态系统产生更加深远的影响.事实上, 麦穗鱼已经是入侵欧洲最严重的外来鱼类(Britton et al ., 2009 ), 而且难以根除, 控制它在新生境的种群数量需要付出巨大的努力和代价 (Britton et al. , 2010a ).此外, 麦穗鱼和鲫在入侵的同时, 还会将其携带的寄生虫、细菌等病原体一同引入入侵地, 感染其他土著鱼类, 并导致这些鱼类严重的种群衰退(Humphrey & Ashburner, 1993 ; Gozlan et al ., 2005 ).这种影响最终会造成土著鱼类种群减少甚至灭绝, 改变原生态系统食物链(Britton et al ., 2009 , 2010b ).我们的研究结果也预示了这些生态问题将会在美国、巴西和阿根廷等国家出现. ...
Biotic invasions: causes, epidemiology, global consequences, and control
1
2000
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Age, growth and habitat use of the topmouth gudgeon, Pseudorasbora parva in irrigation ditches on northwestern Kyushu Island, Japan
1
2013
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
History and approach of invasion from domestic and abroad fishes
1
2006
... 本研究的预测结果表明, 麦穗鱼的适生区集中于15º–55º N之间, 而鲫在40º S–60º N.比较麦穗鱼已知的入侵分布与自然分布, 可以发现麦穗鱼从原产于东亚的我国、朝鲜、韩国和日本, 已经入侵到了欧洲的法国、罗马尼亚、希腊和土耳其等国家.我们推测是由于罗马尼亚在20世纪60年代从我国武汉引进草鱼和鲢鱼鱼卵时, 麦穗鱼混在其中偶然带入的, 之后随着其他欧洲国家通过从我国引种以及欧洲国家间相互引种等方式, 在欧洲广泛扩散(Gozlan et al ., 2010 ).麦穗鱼在我国云南的南盘江和金沙江都有自然分布, 但随着家鱼的引进而进一步扩散至洱海、滇池、抚仙湖、泸沽湖、拉市海和丘北普者黑等高原湖泊, 以及澜沧江、李仙江、藤条江和怒江(Chen & Cui, 1993 ; 潘勇等, 2006 ; 陈小勇, 2013 ).进一步比较麦穗鱼已知分布和预测适生区分布, 我们发现除已知分布的地区外, 北美洲的美国和墨西哥, 欧洲的西班牙和葡萄牙, 非洲北部的地中海沿岸麦穗鱼的生境适合度也较高, 预示了这些地区具有极高的入侵风险. ...
Maximum entropy modeling of species geographic distributions
1
2006
... Maxent是一种根据已知物种分布信息预测该物种潜在分布区域的模型, 广泛应用于生物地理学、生物入侵以及生态位研究等领域(Phillips et al ., 2006 ; Suarez-Seoane et al ., 2008 ).该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法, 即“最大熵原则(the maximum-entropy principle)”, 可以更多地保留物种已有分布数据的有用信息.Giovanelli等(2008 )利用Maxent模型分析了原产于北美的牛蛙(Lithobates catesbeianus )在巴西的入侵范围, 并预测了牛蛙在巴西亚马逊地区和东部的大西洋沿岸的潜在入侵区域.与其他方法相比, Maxent模型所具备的优势使其预测表现始终是最好的(Elith et al ., 2006 ).综合以上情况, 本研究采用Maxent模型对麦穗鱼和鲫在全球的适生区进行了预测, 以期为相关国家和地区对外来鱼类的预警和风险评估提供科学依据. ...
Issues, phenomena and study emphases of alien species invasion in China
1
2006
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Maximum entropy niche-based modelling of seasonal changes in little bustard (Tetrax tetrax) distribution
1
2008
... Maxent是一种根据已知物种分布信息预测该物种潜在分布区域的模型, 广泛应用于生物地理学、生物入侵以及生态位研究等领域(Phillips et al ., 2006 ; Suarez-Seoane et al ., 2008 ).该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法, 即“最大熵原则(the maximum-entropy principle)”, 可以更多地保留物种已有分布数据的有用信息.Giovanelli等(2008 )利用Maxent模型分析了原产于北美的牛蛙(Lithobates catesbeianus )在巴西的入侵范围, 并预测了牛蛙在巴西亚马逊地区和东部的大西洋沿岸的潜在入侵区域.与其他方法相比, Maxent模型所具备的优势使其预测表现始终是最好的(Elith et al ., 2006 ).综合以上情况, 本研究采用Maxent模型对麦穗鱼和鲫在全球的适生区进行了预测, 以期为相关国家和地区对外来鱼类的预警和风险评估提供科学依据. ...
Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale
1
2005
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
1
1977
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
1
2007
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
1
2003
... 作为全球变化的重要组成部分, 生物入侵在世界范围内对生物多样性、环境、生态、社会经济和人类健康都造成了严重的危害, 已引起公众、各国政府和科学家们的广泛关注(Mack et al ., 2000 ; 徐汝梅和叶万辉, 2003 ; 桑卫国等, 2006 ; 张亚平等, 2009 ).外来生物一旦成功入侵, 便难以将其从生态系统中清除出去, 因此早期预防要比后期治理更经济有效(Kolar & Lodge, 2001 ).预测入侵性生物的潜在入侵区域是早期预警的重要手段(Austin, 2002 ; Lockwood et al ., 2009 ).Thuiller等(2005 )利用生态位模型预测了96种原产于南非的植物在世界范围内的潜在入侵地区, 揭示了东非、欧洲、北美洲、南美洲、大洋洲以及我国东部的广大地区都是其适生区, 并且认为可以通过生态位模型预测物种潜在分布的方法来构建外来种的早期预警系统. ...
Study on food habits of Pseudorasbora parva in Liangzi Lake
1
2004
... 麦穗鱼(Pseudorasbora parva )隶属于鲤形目鲤科麦穗鱼属(Pseudorasbora ), 喜好栖息于水浅、流速缓慢水体的中、底层, 尤其是水生植物丛生、富含有机质的池塘、湖泊沿岸带等, 以底栖动物为主要食物(董存有, 1984 ; 杨瑞斌等, 2004 ).麦穗鱼在东亚地区呈自然分布(Kim et al ., 1996 ; 陈宜瑜, 1998 ; Onikura & Nakajima, 2013 ), 也是我国常见的小型鱼类, 自广东省至黑龙江省的广大中东部地区, 以及云南的南盘江流域和金沙江流域都有其自然分布(伍献文, 1982 ; 禇新洛和陈银瑞, 1989 ).鲫(Carassius auratus )隶属于鲤形目鲤科鲫属(Carassius ), 是一种典型的杂食性鱼类, 喜好栖息于静水及缓流水体的底层, 在我国分布广泛且具有很强的适应性, 对水中溶氧和水温都有较高耐受性, 尤其是其产卵场多样(陈佩薰, 1959 ; 乐佩琦, 2000 ).鲫的种下分类尚存争议, 有时作为独立的种对待(Kalous et al ., 2012 ).最近的一些研究表明, 鲫在我国有两个亚种: 鲫的指名亚种(C. a. auratus )和银鲫(C. a. gibelio ) (乐佩琦, 2000 ; 程磊等, 2012 ).但本研究中仍将鲫作为1个种对待, 其自然分布涵盖了东亚的绝大部分地区, 如我国除青藏高原以外的各水系, 以及韩国、朝鲜和日本的各种水体均有分布(乐佩琦, 2000 ; 解玉浩, 2007 ); 在欧洲、西伯利亚和东北亚地区也是自然分布(Berg, 1949 ; Bănărescu, 2002 ; Kottelat & Freyhof, 2007 ). ...
 , 吕植
, 吕植

{kind=link}
{kind=link}
{kind=link}
{kind=link}