The genus Oryza is composed of approximately 24 species. Wild species of Oryza contain a largely untapped resource of agronomically important genes. As an increasing number of genomes of wild rice species have been or will be sequenced, Oryza is becoming a model system for plant comparative, functional and evolutionary genomics studies. Comparative analyses of large genomic regions and whole-genome sequences have revealed molecular mechanisms involved in genome size variation, gene movement, genome evolution of polyploids, transition of euchromatin to heterochromatin and centromere evolution in the genus Oryza. Transposon activity and removal of transposable elements by unequal recombination or illegitimate recombination are two important factors contributing to expansion or contraction of Oryza genomes. Double-strand break repair mediated gene movement, especially non-homologous end joining, is an important source of non-colinear genes. Transition of euchromatin to heterochromatin is accompanied by transposable element amplification, segmental and tandem duplication of genic segments, and acquisition of heterochromatic genes from other genomic locations. Comparative analyses of multiple genomes dramatically improve the precision and sensitivity of evolutionary inference than single-genome analyses can provide. Further investigations on the impact of structural variation, lineage-specific genes and evolution of agriculturally important genes on phenotype diversity and adaptation in the genus Oryza should facilitate molecular breeding and genetic improvement of rice.
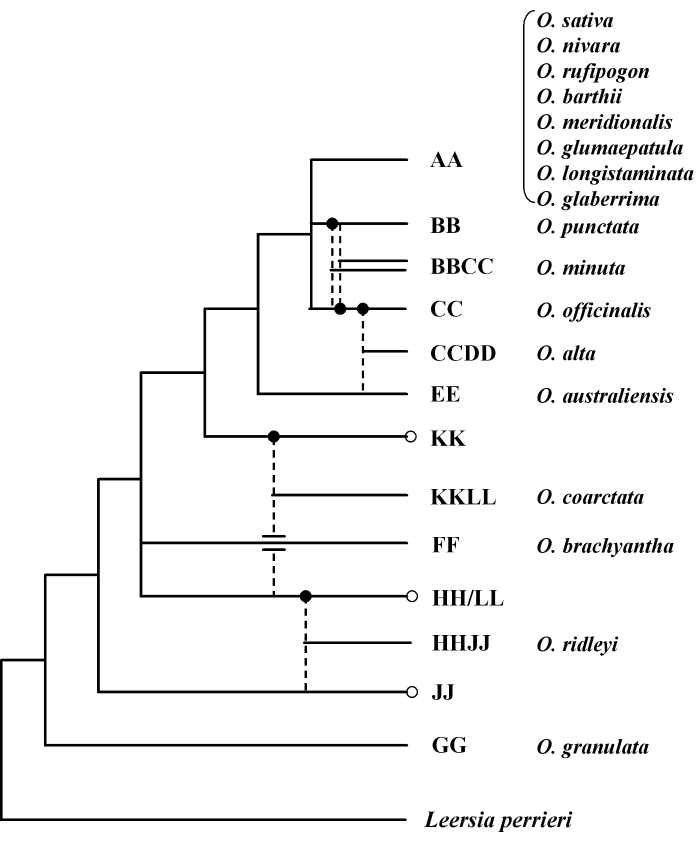
图1 稻属系统进化树(参照Goicoechea et al., 2010)。图中虚线表示四倍体种可能的二倍体父母本。黑色圆点为母本, 空心圆点为不确定的二倍体物种基因组。Leersia perrieri为外类群。17个国际稻属基因组计划(OMAP)中用于基因组研究的代表性物种列在图右侧。
Fig. 1 Phylogenetic tree of Oryza. Dashed lines indicate putative diploid parents involved in the formation of polyploids. Filled in circles indicate maternal parent, and unfilled circles indicate unidentified diploid genome. Leersia perrieri represents the outgroup. Seventeen representative Oryza species that are currently subjected to genomic investigation as part of the International Oryza Map Alignment Project (OMAP) are indicated next to respective phylogenetic branches.
4 稻属基因组多个区段的比较基因组学研究
随着OMAP计划的开展, 稻属基因组进化的研究发展迅速, BAC文库的构建以及指纹图谱的完成使稻属各基因组间大片段比较分析成为可能。通过国际合作, 我们先后选取了水稻Monoculm1 (MOC1)、 Adh1和Heading date 1 (Hd1)3个基因区域, 对稻属代表不同基因组类型的物种进行了序列比较分析, 使我们对于稻属的基因组构成、基因排列、系统发育和分歧时间等方面有了一定的了解 (Ammiraju et al., 2008, 2010; Lu et al., 2009; Sanyal et al., 2010)。3个区段在基因组结构和基因共线性上呈现不同的状态: 在MOC1和Hd1区域, 基因共线性以及基因结构均非常保守, 只是MOC1区域在AA基因组中存在一些特有的基因, 这些基因可能是通过一种从头合成方式形成的新基因; Adh1区域则保守性较差, Adh1基因在O. glaberrima基因组中缺失, 在物种O. rufipogon和O. punctata中突变形成假基因, 同时该区域存在大量串联重复基因。该区段还存在一些基因和片段在不同物种中特异性的获得或者缺失, 特别是在O. australiensis中发生的1个350 kb的倒位和O. granulata中1个219 kb的大片段缺失, 均在一定程度上破坏了该区域的基因共线性特征。
通过对水稻基因组中LTR逆转座子分布进行分析, 发现其密度与重组率和基因密度成反比。高重组区域的LTR趋向于以较小片段和solo LTR形式存在, 说明高重组率加速了LTR的重组删除; 而低重组率区域的LTR则不易被删除, 从而大量累积下来(Tian et al., 2009)。在水稻基因组中, LTR逆转座子的删除一般通过不均等同源重组和非常规重组来完成, 这两种重组方式在维持水稻基因组大小过程中均起到重要作用(Ma et al., 2004)。
7 总结与展望
综上所述, 通过稻属不同基因组区段以及全基因组序列的比较分析, 我们对稻属在基因组大小、基因移动、多倍体进化、着丝粒区域的进化等方面有了比较全面的了解。特别是对常染色质到异染色质的转化区域的研究, 初步揭示了染色质转换的机理, 使人们对植物基因组的进化机制研究更为全面。但是目前的测序和拼接技术成本高、读长短, 对于基因组的从头组装和结构变异分析是巨大的挑战, 这在一定程度上影响了比较基因组学研究的开展。近期发展起来的纳米通道芯片(Nanochannel Arrays)技术(Lam et al., 2012; Hastie et al., 2013)以及三代单分子实时DNA测序(single molecule real time (SMRT™) DNA Sequencing)技术(Flusberg et al., 2010), 将极大地提高复杂物种基因组从头测序和基因组拼接的精确度, 解决基因组结构变异分析必须依赖其他物种间或者物种内部参考序列的难题, 进而促进稻属多物种的从头测序以及水稻多品系的重测序工作。
Chromosome segment substitution lines: a powerful tool for the introgression of valuable genes from Oryza wild species into cultivated rice (O. sativa).
The Oryza bacterial artificial chromosome library resource: construction and analysis of 12 deep-coverage large-insert BAC libraries that represent the 10 genome types of the genus Oryza.
Development of chromosomal segment substitution lines from a backcross recombinant inbred population of interspecific rice cross.
Rice Science, 13, 15-21.
[12]
ChenJ, HuangDR, WangL, LiuGJ, ZhuangJY (2010)
Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa × O. rufipogon.
Mechanisms of different responses of leaf photosynthesis in African rice (Oryza glaberrima Steud.) and rice (Oryza sativa L.) to low leaf water potential.
Japanese Journal of Crop Science, 63, 625-631.
[30]
GeS, SangT, LuBR, HongDY (1999)
Phylogeny of rice genomes with emphasis on origins of allotetraploid species.
Proceedings of the National Academy of Sciences,USA, 96, 14400-14405.
Integration of hybridization-based markers (overgos) into physical maps for comparative and evolutionary explorations in the genus Oryza and in Sorghum.
Doubling genome size without polyploidization: dynamics of retrotransposition-driven genomic expansions in Oryza australiensis, a wild relative of rice.
Orthologous comparisons of the Hd1 region across genera reveal Hd1 gene lability within diploid Oryza species and disruptions to microsynteny in Sorghum.
Introgressions of Oryza grandiglumis chromatin into rice affect plant height and grain length.
1
2002
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
Chromosome segment substitution lines: a powerful tool for the introgression of valuable genes from Oryza wild species into cultivated rice (O. sativa).
2
2010
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
... 区段比较基因组学分析虽然有助于我们对稻属基因组进化的理解, 但是因为各个区段在基因组结构和保守性上呈现不同的状态, 单一区段并不能代表全基因组的进化过程, 因此必须进行基于全基因组序列的稻属比较基因组学分析.O. brachyantha是稻属中与水稻亲缘关系较远的一类野生稻, 具有对多种水稻病原菌和一些非生物胁迫的抗性(Ali et al., 2010); 同时, O. brachyantha具有稻属最小的基因组, 结合其在进化树中的特殊位置, 它可能保持着较为接近稻属祖先基因组的状态.因此通过对O. brachyantha、水稻和其他野生稻的基因组进行比较, 将获取更多稻属在进化过程中发生变异的信息, 从而更为准确地理解稻属基因组的结构和进化过程.Chen等(2013)主要利用二代高通量测序技术完成了261 Mb O. brachyantha全基因组测序, 并与水稻基因组的序列进行了比较, 得到以下结果. ...
Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice, Oryza sativa.
1992
Spatio-temporal patterns of genome evolution in allotetraploid species of the genus Oryza.
1
2010
... 随着OMAP计划的开展, 稻属基因组进化的研究发展迅速, BAC文库的构建以及指纹图谱的完成使稻属各基因组间大片段比较分析成为可能.通过国际合作, 我们先后选取了水稻Monoculm1 (MOC1)、 Adh1和Heading date 1 (Hd1)3个基因区域, 对稻属代表不同基因组类型的物种进行了序列比较分析, 使我们对于稻属的基因组构成、基因排列、系统发育和分歧时间等方面有了一定的了解 (Ammiraju et al., 2008, 2010; Lu et al., 2009; Sanyal et al., 2010).3个区段在基因组结构和基因共线性上呈现不同的状态: 在MOC1和Hd1区域, 基因共线性以及基因结构均非常保守, 只是MOC1区域在AA基因组中存在一些特有的基因, 这些基因可能是通过一种从头合成方式形成的新基因; Adh1区域则保守性较差, Adh1基因在O. glaberrima基因组中缺失, 在物种O. rufipogon和O. punctata中突变形成假基因, 同时该区域存在大量串联重复基因.该区段还存在一些基因和片段在不同物种中特异性的获得或者缺失, 特别是在O. australiensis中发生的1个350 kb的倒位和O. granulata中1个219 kb的大片段缺失, 均在一定程度上破坏了该区域的基因共线性特征. ...
Dynamic evolution of Oryza genomes is revealed by comparative genomic analysis of a genus-wide vertical data set.
1
2008
... 随着OMAP计划的开展, 稻属基因组进化的研究发展迅速, BAC文库的构建以及指纹图谱的完成使稻属各基因组间大片段比较分析成为可能.通过国际合作, 我们先后选取了水稻Monoculm1 (MOC1)、 Adh1和Heading date 1 (Hd1)3个基因区域, 对稻属代表不同基因组类型的物种进行了序列比较分析, 使我们对于稻属的基因组构成、基因排列、系统发育和分歧时间等方面有了一定的了解 (Ammiraju et al., 2008, 2010; Lu et al., 2009; Sanyal et al., 2010).3个区段在基因组结构和基因共线性上呈现不同的状态: 在MOC1和Hd1区域, 基因共线性以及基因结构均非常保守, 只是MOC1区域在AA基因组中存在一些特有的基因, 这些基因可能是通过一种从头合成方式形成的新基因; Adh1区域则保守性较差, Adh1基因在O. glaberrima基因组中缺失, 在物种O. rufipogon和O. punctata中突变形成假基因, 同时该区域存在大量串联重复基因.该区段还存在一些基因和片段在不同物种中特异性的获得或者缺失, 特别是在O. australiensis中发生的1个350 kb的倒位和O. granulata中1个219 kb的大片段缺失, 均在一定程度上破坏了该区域的基因共线性特征. ...
The Oryza bacterial artificial chromosome library resource: construction and analysis of 12 deep-coverage large-insert BAC libraries that represent the 10 genome types of the genus Oryza.
1
2006
... 2005年美国亚利桑那大学启动了“稻属基因组计划”项目(Oryza Map Alignment Project, OMAP) (Wing et al., 2005; Ammiraju et al., 2006).该计划的目的是通过对选择的11个野生稻种及非洲栽培稻O. glaberrima基因组分别构建BAC文库、末端测序和酶切构建指纹图谱, 从而建立一个研究稻属进化、发育、基因组结构、驯化和基因调控网络的系统.OMAP计划的具体目标为: (1)对11个野生稻和1个非洲栽培稻构建BAC文库; (2)对12个BAC文库进行末端测序, 构建酶切指纹图谱; (3)将12个种的物理图谱与国际水稻基因组测序计划(International Rice Genome Sequencing Project, IRGSP) (International Rice Genome Sequencing Project, 2005)发布的日本晴(Oryza sativa L. ssp. japonica cv. Nipponbare)序列对位; (4)对这12个物种的1、3和10号染色体进行精细重构, 并进行比较基因组学分析.在随后的几年里, 科学家完成了18个种的基因组文库构建, 并通过流式细胞仪对各个基因组大小进行了重新计算, 利用野生稻物理图谱中大量的BAC末端序列成功地将物理图谱重叠群(contig)对位到japonica基因组序列上.结果显示, 野生稻与日本晴基因组呈现出高度的共线性(Kim et al., 2008).同时开发了SyMAP软件(Soderlund et al., 2006), 可以在线查询稻属各物种之间以及稻属和玉米、高粱等基因组序列的共线性.这些工作为稻属的比较基因组学研究提供了平台(http://www. omap.org). ...
Evolutionary dynamics of an ancient retrotransposon family provides insights into evolution of genome size in the genus Oryza.
2
2007
... 在已经完成测序的植物基因组中, 占基因组比例最高的转座子一般都是RNA类型的LTR逆转座子(long terminal repeat retrotransposons).在玉米(2,300 Mb)、高粱(730 Mb)和水稻(398 Mb)基因组中, LTR转座子分别约占基因组的75%、54%和23% (International Rice Genome Sequencing Project, 2005; Paterson et al., 2009; Schnable et al., 2009).LTR转座子含量和基因组大小的正相关性暗示该类型转座子在基因组大小变化中起着重要作用.近期对稻属基因组研究表明(Bennetzen & Kellogg, 1997; Ma et al., 2004; Piegu et al., 2006; Ammiraju et al., 2007; Tian et al., 2009), 不同家族的LTR逆转座子的扩增会导致基因组的剧烈膨胀, 这为上述观点提供了直接证据. ...
Breeding rice varieties for resistance against Chilo spp. of stem borers in Asia and Africa.
1990
Development of chromosomal segment substitution lines from a backcross recombinant inbred population of interspecific rice cross.
2006
Identification of quantitative trait loci for resistance to whitebacked planthopper, Sogatella furcifera, from an interspecific cross Oryza sativa × O. rufipogon.
Conservation and purifying selection of transcribed genes located in a rice centromere.
1
2011
... 早期通过荧光原位杂交技术发现水稻着丝粒区域的基因组序列比较复杂, 主要包括CentO串联重复序列以及CRR反转座子(centromere-specific retrotransposon)(Cheng et al., 2002).在12条染色体中只有8号染色体着丝粒区的CentO串联重复序列较少, 并且已经获得了该区域的全长序列(Nagaki et al., 2004; Wu et al., 2004), 因此水稻8号染色体成为研究水稻着丝粒区域基因组结构、进化以及着丝粒功能的模型.研究发现该着丝粒区域不仅含有特异的CentO串联重复序列和CRR反转座子, 还含有少量转录活跃的基因(Nagaki et al., 2004, 2005; Wu et al., 2004).异染色质区由于大量转座子插入引起基因组巨大的结构变异, 一般为重组抑制区.这些区域由于缺乏自然选择, 容易加速有害突变的积累, 进一步造成基因功能的丧失以及染色体的退化 (Haddrill et al., 2007).但是对水稻与O. glaberrima和O. brachyantha在8号染色体着丝粒区域的比较分析却发现了7个保守的基因, 并且这些基因均受到纯化选择(purifying selection), 同时这些基因的序列在不同物种间的差异要明显小于常染色质基因, 说明可能存在某种机制来维持稻属着丝粒区域内基因的保守性以防止功能丧失(Fan et al., 2011). ...
Plant transposable elements: where genetics meets genomics.
1
2002
... 近年来基因组学研究的快速发展使我们认识到转座子在基因组中的重要性.转座子不仅对基因组大小具有重要影响, 而且通过插入和删除突变影响基因表达调控, 甚至可以通过介导新基因形成来影响物种的多样性, 因此已作为功能基因组的研究手段得以广泛应用(Feschotte et al., 2002). ...
Direct detection of DNA methylation during single-molecule, real-time sequencing.
1
2010
... 综上所述, 通过稻属不同基因组区段以及全基因组序列的比较分析, 我们对稻属在基因组大小、基因移动、多倍体进化、着丝粒区域的进化等方面有了比较全面的了解.特别是对常染色质到异染色质的转化区域的研究, 初步揭示了染色质转换的机理, 使人们对植物基因组的进化机制研究更为全面.但是目前的测序和拼接技术成本高、读长短, 对于基因组的从头组装和结构变异分析是巨大的挑战, 这在一定程度上影响了比较基因组学研究的开展.近期发展起来的纳米通道芯片(Nanochannel Arrays)技术(Lam et al., 2012; Hastie et al., 2013)以及三代单分子实时DNA测序(single molecule real time (SMRT™) DNA Sequencing)技术(Flusberg et al., 2010), 将极大地提高复杂物种基因组从头测序和基因组拼接的精确度, 解决基因组结构变异分析必须依赖其他物种间或者物种内部参考序列的难题, 进而促进稻属多物种的从头测序以及水稻多品系的重测序工作. ...
A recombination hotspot delimits a wild-species quantitative trait locus for tomato sugar content to 484 bp within an invertase gene.
Analysis of QTLs for yield-related traits in Yuanjiang common wild rice (Oryza rufipogon Griff.).
2010
Progress on transferring elite genes from non-AA genome wild rice into Oryza sativa through interspecific hybridization.
1
2008
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
Mechanisms of different responses of leaf photosynthesis in African rice (Oryza glaberrima Steud.) and rice (Oryza sativa L.) to low leaf water potential.
1994
Phylogeny of rice genomes with emphasis on origins of allotetraploid species.
2
1999
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
... 清晰明确的系统进化关系对于合理有效地利用稻属野生稻资源是至关重要的, 同时它也是稻属基因组进化研究的基础(Tanksley & McCouch, 1997).稻属各物种之间的亲缘关系一直是植物分类学家所关心的问题.自20世纪60年代以来, 研究人员根据形态特征、杂交育性实验(Harland & De-Wet, 1971)、同工酶分析、RFLP分析(Dally & Second, 1990; Sarkar & Raina, 1992; Wang et al., 1992)等技术将稻属划分成不同的复合群.但是上述分类的准确性欠佳, 尤其是对群内的四倍体物种的分辨能力较差, 其进化关系仍不清楚(Ge et al., 1999). ...
The future of rice genomics: sequencing the collective Oryza genome.
... 稻属系统进化树(参照Goicoechea et al., 2010).图中虚线表示四倍体种可能的二倍体父母本.黑色圆点为母本, 空心圆点为不确定的二倍体物种基因组.Leersia perrieri为外类群.17个国际稻属基因组计划(OMAP)中用于基因组研究的代表性物种列在图右侧. ...
Identification of a rice stripe necrosis virus resistance locus and yield component QTLs using Oryza sativa × O. glaberrima introgression lines.
2010
Reduced efficacy of selection in regions of the Drosophila genome that lack crossing over.
1
2007
... 早期通过荧光原位杂交技术发现水稻着丝粒区域的基因组序列比较复杂, 主要包括CentO串联重复序列以及CRR反转座子(centromere-specific retrotransposon)(Cheng et al., 2002).在12条染色体中只有8号染色体着丝粒区的CentO串联重复序列较少, 并且已经获得了该区域的全长序列(Nagaki et al., 2004; Wu et al., 2004), 因此水稻8号染色体成为研究水稻着丝粒区域基因组结构、进化以及着丝粒功能的模型.研究发现该着丝粒区域不仅含有特异的CentO串联重复序列和CRR反转座子, 还含有少量转录活跃的基因(Nagaki et al., 2004, 2005; Wu et al., 2004).异染色质区由于大量转座子插入引起基因组巨大的结构变异, 一般为重组抑制区.这些区域由于缺乏自然选择, 容易加速有害突变的积累, 进一步造成基因功能的丧失以及染色体的退化 (Haddrill et al., 2007).但是对水稻与O. glaberrima和O. brachyantha在8号染色体着丝粒区域的比较分析却发现了7个保守的基因, 并且这些基因均受到纯化选择(purifying selection), 同时这些基因的序列在不同物种间的差异要明显小于常染色质基因, 说明可能存在某种机制来维持稻属着丝粒区域内基因的保守性以防止功能丧失(Fan et al., 2011). ...
Toward a rational classification of cultivated plants.
2
1971
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
... 清晰明确的系统进化关系对于合理有效地利用稻属野生稻资源是至关重要的, 同时它也是稻属基因组进化研究的基础(Tanksley & McCouch, 1997).稻属各物种之间的亲缘关系一直是植物分类学家所关心的问题.自20世纪60年代以来, 研究人员根据形态特征、杂交育性实验(Harland & De-Wet, 1971)、同工酶分析、RFLP分析(Dally & Second, 1990; Sarkar & Raina, 1992; Wang et al., 1992)等技术将稻属划分成不同的复合群.但是上述分类的准确性欠佳, 尤其是对群内的四倍体物种的分辨能力较差, 其进化关系仍不清楚(Ge et al., 1999). ...
Integration of hybridization-based markers (overgos) into physical maps for comparative and evolutionary explorations in the genus Oryza and in Sorghum.
Rapid genome mapping in nanochannel arrays for highly complete and accurate de novo sequence assembly of the complex Aegilops tauschii genome.
1
2013
... 综上所述, 通过稻属不同基因组区段以及全基因组序列的比较分析, 我们对稻属在基因组大小、基因移动、多倍体进化、着丝粒区域的进化等方面有了比较全面的了解.特别是对常染色质到异染色质的转化区域的研究, 初步揭示了染色质转换的机理, 使人们对植物基因组的进化机制研究更为全面.但是目前的测序和拼接技术成本高、读长短, 对于基因组的从头组装和结构变异分析是巨大的挑战, 这在一定程度上影响了比较基因组学研究的开展.近期发展起来的纳米通道芯片(Nanochannel Arrays)技术(Lam et al., 2012; Hastie et al., 2013)以及三代单分子实时DNA测序(single molecule real time (SMRT™) DNA Sequencing)技术(Flusberg et al., 2010), 将极大地提高复杂物种基因组从头测序和基因组拼接的精确度, 解决基因组结构变异分析必须依赖其他物种间或者物种内部参考序列的难题, 进而促进稻属多物种的从头测序以及水稻多品系的重测序工作. ...
Resistance of wild rices, Oryza spp. to the whorl maggot Hydrellia- Philippina Ferino (Diptera, Ephydridae).
1985
Das Heterochromatin der Moose
1
1928
... 异染色质区域通常指染色较深、结构高度固缩、重组交换和基因表达均受到抑制的区域(Heitz, 1928; Huisinga et al., 2006).异染色质区域一般位于染色体上重复序列富集的区域, 如着丝粒(centromere)、端粒(telomere)及异染色质结(knob)等位置(Martens et al., 2005).重复序列的富集使异染色质区域序列很难获得, 但是随着越来越多的全基因组测序完成, 异染色质区域的比较分析更加深入.目前稻属异染色质区域的研究主要集中在着丝粒区域. ...
Assessing hybrid sterility in Oryza glaberrima × O. sativa hybrid progenies by PCR marker analysis and crossing with wide compatibility varieties.
2003
Identification and characterization of new sources of cytoplasmic male sterility in rice.
1997
The contradictory definitions of heterochromatin: transcription and silencing.
1
2006
... 异染色质区域通常指染色较深、结构高度固缩、重组交换和基因表达均受到抑制的区域(Heitz, 1928; Huisinga et al., 2006).异染色质区域一般位于染色体上重复序列富集的区域, 如着丝粒(centromere)、端粒(telomere)及异染色质结(knob)等位置(Martens et al., 2005).重复序列的富集使异染色质区域序列很难获得, 但是随着越来越多的全基因组测序完成, 异染色质区域的比较分析更加深入.目前稻属异染色质区域的研究主要集中在着丝粒区域. ...
The map-based sequence of the rice genome.
3
2005
... 2005年美国亚利桑那大学启动了“稻属基因组计划”项目(Oryza Map Alignment Project, OMAP) (Wing et al., 2005; Ammiraju et al., 2006).该计划的目的是通过对选择的11个野生稻种及非洲栽培稻O. glaberrima基因组分别构建BAC文库、末端测序和酶切构建指纹图谱, 从而建立一个研究稻属进化、发育、基因组结构、驯化和基因调控网络的系统.OMAP计划的具体目标为: (1)对11个野生稻和1个非洲栽培稻构建BAC文库; (2)对12个BAC文库进行末端测序, 构建酶切指纹图谱; (3)将12个种的物理图谱与国际水稻基因组测序计划(International Rice Genome Sequencing Project, IRGSP) (International Rice Genome Sequencing Project, 2005)发布的日本晴(Oryza sativa L. ssp. japonica cv. Nipponbare)序列对位; (4)对这12个物种的1、3和10号染色体进行精细重构, 并进行比较基因组学分析.在随后的几年里, 科学家完成了18个种的基因组文库构建, 并通过流式细胞仪对各个基因组大小进行了重新计算, 利用野生稻物理图谱中大量的BAC末端序列成功地将物理图谱重叠群(contig)对位到japonica基因组序列上.结果显示, 野生稻与日本晴基因组呈现出高度的共线性(Kim et al., 2008).同时开发了SyMAP软件(Soderlund et al., 2006), 可以在线查询稻属各物种之间以及稻属和玉米、高粱等基因组序列的共线性.这些工作为稻属的比较基因组学研究提供了平台(http://www. omap.org). ...
... 在已经完成测序的植物基因组中, 占基因组比例最高的转座子一般都是RNA类型的LTR逆转座子(long terminal repeat retrotransposons).在玉米(2,300 Mb)、高粱(730 Mb)和水稻(398 Mb)基因组中, LTR转座子分别约占基因组的75%、54%和23% (International Rice Genome Sequencing Project, 2005; Paterson et al., 2009; Schnable et al., 2009).LTR转座子含量和基因组大小的正相关性暗示该类型转座子在基因组大小变化中起着重要作用.近期对稻属基因组研究表明(Bennetzen & Kellogg, 1997; Ma et al., 2004; Piegu et al., 2006; Ammiraju et al., 2007; Tian et al., 2009), 不同家族的LTR逆转座子的扩增会导致基因组的剧烈膨胀, 这为上述观点提供了直接证据. ...
Long-range and targeted ectopic recombination between the two homeologous chromosomes 11 and 12 in Oryza species.
2011
A recent duplication revisited: phylogenetic analysis reveals an ancestral duplication highly-conserved throughout the Oryza genus and beyond.
1
2009
... 多倍体在植物界中普遍存在, 是植物进化的主要动力之一.多倍体形成之后会随即发生染色体重排、基因表达模式改变以及二倍体化等一系列变化, 以缓解基因组加倍对物种的冲击.最初对日本晴进行基因组序列分析时, 国际上多个实验室发现水稻曾经历过一次全基因组加倍, 这次多倍化事件发生在禾本科分化之前, 即约5-7千万年前, 还有一次是约5百万年前发生的位于11、12号染色体短臂间的区段性加倍(Paterson et al., 2004; Tian et al., 2005; Wang et al., 2005; Yu et al., 2005).近期研究显示, 11号和12号染色体短臂加倍事件在高粱和二穗短柄草基因组中也存在(Jacquemin et al., 2009; Paterson et al., 2009), 暗示该区段可能是禾本科二倍体化过程中留下的加倍区段, Jacquemin等(2011)在对日本晴、O. glaberrima和O. brachyantha的加倍区段比较分析发现, 同一物种的加倍区段之间无论是基因还是基因间区都表现出了很高的保守性, 尤其是在O. brachyantha中这个特点更为明显.进化分析表明这种保守性是由于非交互重组(nonreciprocal recombination)即基因转换(gene conversion)造成的, 暗示了基因转换在禾本科基因组进化中起着重要的作用.另外, 在基因组的不同位置, 基因转换发生的频率也并不相同, 一般情况下近端粒区的转换频率高于其他染色体区域(Wang et al., 2009), 说明染色质环境是影响二倍体化过程中重复基因进化的重要因素. ...
Introgression of genes from Oryza officinalis Well ex Watt to cultivated rice, O. sativa L.
1
1990
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
A tiling microarray expression analysis of rice chromosome 4 suggests a chromosome-level regulation of transcription.
1
2005
... 细胞学实验表明, 水稻基因组内除了着丝粒和端粒区域为异染色质结构外, 还分布着大量的异染色质, 尤其是水稻4号和10号染色体, 在减数分裂粗线期着丝粒附近和整条短臂均呈现异染色质状态(Cheng et al., 2001; Jiao et al., 2005).我们对位于4号染色体短臂的H1区域(1.95-4.25 Mb)进行细胞学观察, 结果显示H1区域在稻属3种基因组类型(O. glaberrima [AA], O. punctata [BB]和O. brachyantha [FF])中呈现不同的异染色质状态: 在AA、BB基因组中为异染色质状态, 而在FF基因组中则呈现常染色质状态(Chen et al., 2013).我们对稻属5个基因组(O. japonica, O. glaberrima, O. nivara, O. punctata和O. brachyantha)中H1同源区域进行了比较和进化基因组学分析(实验室未发表数据).序列比较发现, 稻属不同物种中H1同源区域呈现极大变异, 与保持常染色质状态的O. brachyantha基因组区域相比, 水稻该基因组区域扩大了3倍之多, 这种现象主要是由于转座子, 特别是LTR类型的转座子累积引起的.H1区域基因存在特殊的进化方式: 一方面通过高频率的基因重复(包括串联重复以及区段性重复两种方式)不断地扩大现有的基因家族, 而基因重复后功能的冗余进一步介导H1区域大量转座子的积累; 另一方面, 不断通过“复制-粘贴”方式从基因组其他位置上获得外来基因, 而这些外来基因的募集主要以转座子以及DNA双链断裂修复中发生的非法重组所介导. ...
Introgression for grain traits from Oryza minuta into rice, O. sativa.
2004
Origin, dispersal, cultivation and variation of rice.
1
1997
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
Construction, alignment and analysis of twelve framework physical maps that represent the ten genome types of the genus Oryza.
1
2008
... 2005年美国亚利桑那大学启动了“稻属基因组计划”项目(Oryza Map Alignment Project, OMAP) (Wing et al., 2005; Ammiraju et al., 2006).该计划的目的是通过对选择的11个野生稻种及非洲栽培稻O. glaberrima基因组分别构建BAC文库、末端测序和酶切构建指纹图谱, 从而建立一个研究稻属进化、发育、基因组结构、驯化和基因调控网络的系统.OMAP计划的具体目标为: (1)对11个野生稻和1个非洲栽培稻构建BAC文库; (2)对12个BAC文库进行末端测序, 构建酶切指纹图谱; (3)将12个种的物理图谱与国际水稻基因组测序计划(International Rice Genome Sequencing Project, IRGSP) (International Rice Genome Sequencing Project, 2005)发布的日本晴(Oryza sativa L. ssp. japonica cv. Nipponbare)序列对位; (4)对这12个物种的1、3和10号染色体进行精细重构, 并进行比较基因组学分析.在随后的几年里, 科学家完成了18个种的基因组文库构建, 并通过流式细胞仪对各个基因组大小进行了重新计算, 利用野生稻物理图谱中大量的BAC末端序列成功地将物理图谱重叠群(contig)对位到japonica基因组序列上.结果显示, 野生稻与日本晴基因组呈现出高度的共线性(Kim et al., 2008).同时开发了SyMAP软件(Soderlund et al., 2006), 可以在线查询稻属各物种之间以及稻属和玉米、高粱等基因组序列的共线性.这些工作为稻属的比较基因组学研究提供了平台(http://www. omap.org). ...
Resistance to infection of rice tungro viruses and vector resistance in wild species of rice (Oryza spp.).
1993
Screening wild species of rice (Oryza spp.) for resistance to rice tungro disease.
1994
Genome mapping on nanochannel arrays for structural variation analysis and sequence assembly.
1
2012
... 综上所述, 通过稻属不同基因组区段以及全基因组序列的比较分析, 我们对稻属在基因组大小、基因移动、多倍体进化、着丝粒区域的进化等方面有了比较全面的了解.特别是对常染色质到异染色质的转化区域的研究, 初步揭示了染色质转换的机理, 使人们对植物基因组的进化机制研究更为全面.但是目前的测序和拼接技术成本高、读长短, 对于基因组的从头组装和结构变异分析是巨大的挑战, 这在一定程度上影响了比较基因组学研究的开展.近期发展起来的纳米通道芯片(Nanochannel Arrays)技术(Lam et al., 2012; Hastie et al., 2013)以及三代单分子实时DNA测序(single molecule real time (SMRT™) DNA Sequencing)技术(Flusberg et al., 2010), 将极大地提高复杂物种基因组从头测序和基因组拼接的精确度, 解决基因组结构变异分析必须依赖其他物种间或者物种内部参考序列的难题, 进而促进稻属多物种的从头测序以及水稻多品系的重测序工作. ...
Identification and mapping of genes for improving yield from Chinese common wild rice (O. rufipogon Griff.) using advanced backcross QTL analysis.
2002
Mapping quantitative trait loci for heading date and awn length using an advanced backcross line from a cross between Oryza sativa and O. minuta.
2006
Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6.
2002
Taxonomy of the genus Oryza (Poaceae): historical perspective and current status.
... 在已经完成测序的植物基因组中, 占基因组比例最高的转座子一般都是RNA类型的LTR逆转座子(long terminal repeat retrotransposons).在玉米(2,300 Mb)、高粱(730 Mb)和水稻(398 Mb)基因组中, LTR转座子分别约占基因组的75%、54%和23% (International Rice Genome Sequencing Project, 2005; Paterson et al., 2009; Schnable et al., 2009).LTR转座子含量和基因组大小的正相关性暗示该类型转座子在基因组大小变化中起着重要作用.近期对稻属基因组研究表明(Bennetzen & Kellogg, 1997; Ma et al., 2004; Piegu et al., 2006; Ammiraju et al., 2007; Tian et al., 2009), 不同家族的LTR逆转座子的扩增会导致基因组的剧烈膨胀, 这为上述观点提供了直接证据. ...
Doubling genome size without polyploidization: dynamics of retrotransposition-driven genomic expansions in Oryza australiensis, a wild relative of rice.
2
2006
... 在已经完成测序的植物基因组中, 占基因组比例最高的转座子一般都是RNA类型的LTR逆转座子(long terminal repeat retrotransposons).在玉米(2,300 Mb)、高粱(730 Mb)和水稻(398 Mb)基因组中, LTR转座子分别约占基因组的75%、54%和23% (International Rice Genome Sequencing Project, 2005; Paterson et al., 2009; Schnable et al., 2009).LTR转座子含量和基因组大小的正相关性暗示该类型转座子在基因组大小变化中起着重要作用.近期对稻属基因组研究表明(Bennetzen & Kellogg, 1997; Ma et al., 2004; Piegu et al., 2006; Ammiraju et al., 2007; Tian et al., 2009), 不同家族的LTR逆转座子的扩增会导致基因组的剧烈膨胀, 这为上述观点提供了直接证据. ...
Rice sheath blight disease resistance identified in Oryza spp. accessions.
2008
Identification of QTLs for some agronomic traits in rice using an introgression line from Oryza minuta.
2007
a) Identification of new genes for brown planthopper resistance in rice introgressed from O. glaberrima and O. minuta.
1
2010
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
b) Identification of a new gene introgressed from Oryza brachyantha with broad-spectrum resistance to bacterial blight of rice in India.
1
2010
... 自20世纪30年代, 科学家们就开始了稻属各个物种基因组分类工作.到目前为止, 通过形态学、细胞学以及分子标记等研究, 稻属24个种的基因组被划分为10个类型, 包括6个二倍体类型(AA、 BB、 CC、 EE、 FF和GG)以及4个异源四倍体类型(BBCC、 CCDD、 HHJJ和KKLL)(表1) (Ge et al., 1999; Zou et al., 2008; Lu et al., 2009).最初人们认为KKLL基因组类型为HHKK型, 但是在Lu等(2009)的数据比较中发现HHKK中的HH基因组不同于HHJJ中的HH基因组, 因此将原来的HHKK基因组改为KKLL基因组.此外, 育种学家根据各个物种间的生殖隔离以及杂交育性, 将稻属划分成不同的复合群, 包括O. sativa、O. officinalis、O. meyeri- ana和O. ridleyi等(Harland & De-Wet, 1971).处于O. sativa复合群中的稻种均属于AA基因组类型, 它们和栽培稻之间可以通过传统方式进行杂交; 而O. sativa和O. officinalis复合群的物种之间必须依靠胚胎拯救的方法才可以进行杂交(Jena & Khush, 1990; Multani et al., 1994); O. sativa和其他复合群的稻种之间即使使用胚胎拯救的方法, 也极难形成杂种(Khush, 1997).近期染色体片段替换系(chromosome segment substitution lines, CSSLs)和回交重组自交系(backcross inbred lines, BILs)成为鉴定野生稻中优良基因以及将其转化到栽培稻中的有利工具(Ali et al., 2010).目前科学家们已经获得了几十个含有野生稻染色体片段的替换系, 其中有几个非AA基因组物种的替换系, 包括BBCC、CCDD、EE和FF基因组类型(Ahn et al., 2002; Fu et al., 2008; Ram et al., 2010a, b).这些替换系对水稻育种及品种改良具有重要意义. ...
Improving rice for broad-spectrum resistance to blast and salinity tolerance by introgressing genes from O. rufipogon.
2005
Comparative functional genomics of the fission yeasts.
1
2011
... 首先, 我们可以利用稻属其他基因组的序列保守性对水稻基因组进行重新注释.通过稻属不同物种间和物种内部基因组的序列比对, 筛选保守区域, 再根据编码基因和非编码调控序列特征以及cDNA、EST、转录组和表观修饰等数据, 对水稻进行系统的功能注释, 更为精确地鉴定各种功能元件, 提高我们对水稻基因组的认识, 加速水稻功能基因组研究的发展(Roy et al., 2010; Rhind et al., 2011). ...
Identification of functional elements and regulatory circuits by Drosophila modENCODE.
1
2010
... 首先, 我们可以利用稻属其他基因组的序列保守性对水稻基因组进行重新注释.通过稻属不同物种间和物种内部基因组的序列比对, 筛选保守区域, 再根据编码基因和非编码调控序列特征以及cDNA、EST、转录组和表观修饰等数据, 对水稻进行系统的功能注释, 更为精确地鉴定各种功能元件, 提高我们对水稻基因组的认识, 加速水稻功能基因组研究的发展(Roy et al., 2010; Rhind et al., 2011). ...
Analysis of mitochondrial DNAs from Oryza glaberrima and its cytoplasmic substituted line for Oryza sativa associated with cytoplasmic male sterility.
1990
Orthologous comparisons of the Hd1 region across genera reveal Hd1 gene lability within diploid Oryza species and disruptions to microsynteny in Sorghum.
1
2010
... 随着OMAP计划的开展, 稻属基因组进化的研究发展迅速, BAC文库的构建以及指纹图谱的完成使稻属各基因组间大片段比较分析成为可能.通过国际合作, 我们先后选取了水稻Monoculm1 (MOC1)、 Adh1和Heading date 1 (Hd1)3个基因区域, 对稻属代表不同基因组类型的物种进行了序列比较分析, 使我们对于稻属的基因组构成、基因排列、系统发育和分歧时间等方面有了一定的了解 (Ammiraju et al., 2008, 2010; Lu et al., 2009; Sanyal et al., 2010).3个区段在基因组结构和基因共线性上呈现不同的状态: 在MOC1和Hd1区域, 基因共线性以及基因结构均非常保守, 只是MOC1区域在AA基因组中存在一些特有的基因, 这些基因可能是通过一种从头合成方式形成的新基因; Adh1区域则保守性较差, Adh1基因在O. glaberrima基因组中缺失, 在物种O. rufipogon和O. punctata中突变形成假基因, 同时该区域存在大量串联重复基因.该区段还存在一些基因和片段在不同物种中特异性的获得或者缺失, 特别是在O. australiensis中发生的1个350 kb的倒位和O. granulata中1个219 kb的大片段缺失, 均在一定程度上破坏了该区域的基因共线性特征. ...
Assessment of genome relationships in the genus Oryza L. based on seed-protein profile analysis.
1
1992
... 清晰明确的系统进化关系对于合理有效地利用稻属野生稻资源是至关重要的, 同时它也是稻属基因组进化研究的基础(Tanksley & McCouch, 1997).稻属各物种之间的亲缘关系一直是植物分类学家所关心的问题.自20世纪60年代以来, 研究人员根据形态特征、杂交育性实验(Harland & De-Wet, 1971)、同工酶分析、RFLP分析(Dally & Second, 1990; Sarkar & Raina, 1992; Wang et al., 1992)等技术将稻属划分成不同的复合群.但是上述分类的准确性欠佳, 尤其是对群内的四倍体物种的分辨能力较差, 其进化关系仍不清楚(Ge et al., 1999). ...
The B73 maize genome: complexity, diversity, and dynamics.



{kind=link}
{kind=link}