Limin Lu1, 2, Miao Sun1, 2, Jingbo Zhang1, 2, Honglei Li1, 2, Li Lin1, 2, Tuo Yang1, 2, Min Chen1, 2, Zhiduan Chen1, *,
1. State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 1000932. Graduate University of the Chinese Academy of Sciences, Beijing 100049
通讯作者: * 通讯作者 Author for correspondence. E-mail: zhiduan@ibcas.ac.cn* 通讯作者 Author for correspondence. E-mail: zhiduan@ibcas.ac.cn
The term “Tree of Life” was first used by Charles Darwin in 1859 as a metaphor for describing phylogenetic relationships among organisms. Over the past three decades, the recognized tree of life has improved considerably in overall size and reliability due to an increase in diversity of character resources, a dramatic growth in useable data, and the development of tree-reconstruction methods. As a bridge connecting phylogeny, evolution and related disciplines, such as molecular biology, ecology, genomics, bioinformatics and computer science, the tree of life is increasingly widely used. In this paper, we review the history and progress of tree of life studies and focus on its application in the following fields: (1) the reconstruction of phylogenetic trees at different taxonomic hierarchies to understand phylogenetic relationships among taxa; (2) investigation of the origins of taxa and biogeographic patterns based on dating estimation and biogeographic reconstruction; (3) examination of species’ diversification and its causes by integrating dated trees, ecological factors, environmental variation and key innovations; (4) the study of the origin and patterns of biodiversity, predating biodiversity dynamics, and development of conservation strategies. Finally, we evaluate the difficulties from matrix alignment, gene tree incongruence and “rogue taxa” distraction in tree reconstruction due to massive increases of useable data and in the context consider “supertree” building in the future.
Keywords:tree of life
;
phylogeny
;
gene tree incongruence
;
biogeography
;
biodiversity
;
phylogenetic diversity
LiminLu, MiaoSun, JingboZhang, HongleiLi, LiLin, TuoYang, MinChen, ZhiduanChen. Tree of life and its applications[J]. Biodiver Sci, 2014, 22(1): 3-20 https://doi.org/10.3724/SP.J.1003.2014.13170
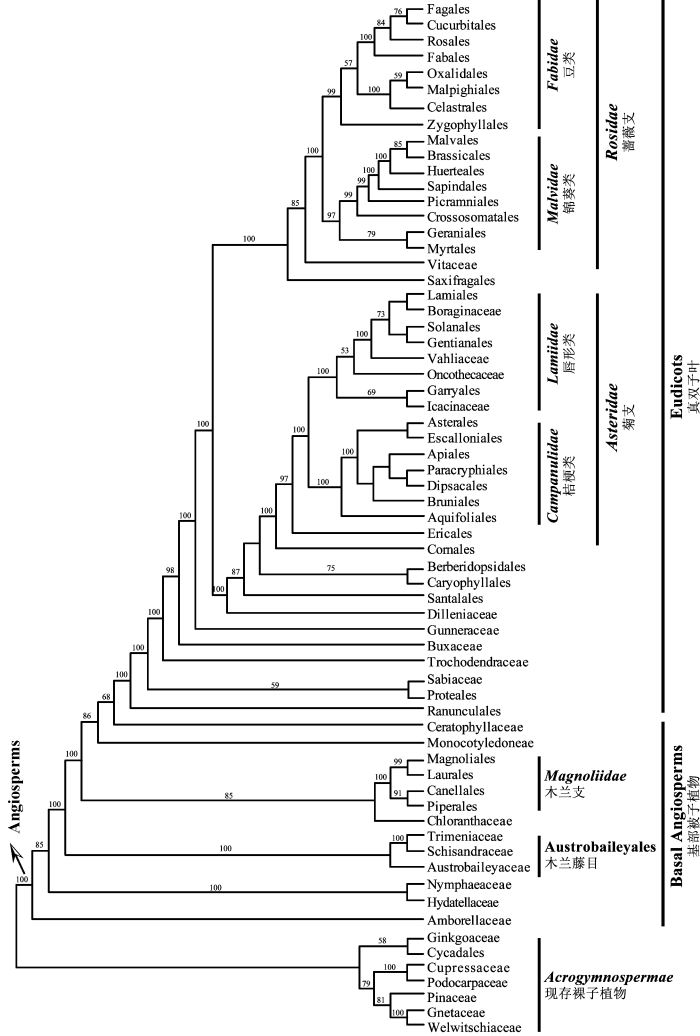
图1 被子植物目级水平的系统发育框架(根据Soltis et al., 2011绘制)。图中的名称与范围与APG III (2009)和Cantino等 (2007)的系统一致; 枝上的数字表示最大似然法BS (bootstrap)支持率。
Fig. 1 Phylogenetic relationships of the angiosperms at the ordinal level (based on Soltis et al., 2011). Names of the families, orders and other major clades follow APG III (2009) and Cantino et al. (2007). Numbers above branches are Maximum Likelihood bootstrap values.
(2)序列拼接(assemblage)。为了提高序列的准确性, 往往需要对所测正反向序列进行拼接和校正, 常用的拼接软件有Contig Express、Geneious (created by Biomatters, available at http://www.geneious.com)、Sequencher (Gene Codes Co., Ann Arbor, Michigan, U.S.A.)等。
Inferences of biogeographical histories within subfamily Hyacinthoideae using S-DIVA and Bayesian binary MCMC analysis implemented in RASP (Reconstruct Ancestral State in Phylogenies).
When are adaptive radiations replicated in areas? Ecological opportunity and unexceptional diversification in West Indian dipsadine snakes (Colubridae: Alsophiini).
Insights into the historical construction of species-rich Mesoamerican seasonally dry tropical forests: the diversification of Bursera (Burseraceae, Sapindales).
New Phytologist, 193, 276-287.
[30]
DegnanJH, RosenbergNA (2009)
Gene tree discordance, phylogenetic inference and the multispecies coalescent.
GeXJ (葛学军) (2013) Community phylogeny and DNA barcoding. In: Frontier in Plant DNA Barcoding (植物DNA条形码前沿探讨) (ed. Shen AM (沈爱民)), pp. 11-21. China Science and Technology Press, Beijing. (in Chinese)
Estimating ancestral distributions of lineages with uncertain sister groups: a statistical approach to Dispersal-Vicariance Analysis and a case using Aesculus L. (Sapindaceae) including fossils.
Journal of Systematics and Evolution, 47, 349-368.
Difference of Relationships Between Spatial Pattern and Environment for Seed Plant Diversity in China (中国种子植物多样性的空间格局-环境关系分异研究). PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing
The contribution of rare species to community phylogenetic diversity across a global network of forest plots.
The American Naturalist, 180, 17-30.
[110]
MiXC (米湘成), PeiNC (裴男才), MaKP (马克平) (2013) On phylogenetic community ecology. In: The New Biology Yearbook 2013 (新生物年鉴2013) (ed. Pu MM (蒲慕明)), pp. 266-289. Science Press, Beijing. (in Chinese)
Nonuniform concerted evolution and chloroplast capture: heterogeneity of observed introgression patterns in three molecular data partition phylogenies of Asian Mitella (Saxifragaceae).
RavenP (1996) Preface. In: The Ecology and Biogeography of Nothofagus Forests (eds Veblen TT, Hill RS, Read J), pp. vii, Yale University Press, New Haven.
Phylogentic placement of two enigmatic genera, Borthwickia and Stixis, based on molecular and pollen data, and the description of a new family of Brassicales, Borthwickiaceae.
WendelJF, DoyleJJ (1998) Phylogenetic incongruence: window into genome history and molecular evolution. In: Molecular Systematics of Plants II: DNA Sequencing (eds Soltis DE, Soltis PS, Doyle JJ), pp. 265-296. Kluwer, Boston.
Integrated fossil and molecular data reveal the biogeographic diversification of the eastern Asian-eastern North American disjunct hickory genus (Carya Nutt.).
Nine exceptinoal radiations plus high turnover explain species diversity in jawed vertebrates.
1
2009
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
Inferences of biogeographical histories within subfamily Hyacinthoideae using S-DIVA and Bayesian binary MCMC analysis implemented in RASP (Reconstruct Ancestral State in Phylogenies).
An ordinal classification for the families of flowering plants.
2
1998
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants APG II.
2
2003
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
3
2009
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... 被子植物目级水平的系统发育框架(根据Soltis et al., 2011绘制).图中的名称与范围与APG III (2009)和Cantino等 (2007)的系统一致; 枝上的数字表示最大似然法BS (bootstrap)支持率. ...
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
Global biogeography and diversification of palms sheds light on the evolution of tropical lineages. I. Historical biogeography.
1
2013
... 随着生物地理学理论和方法的发展, 该领域研究的对象和关注的问题日益广泛, 包括对某一特定类群散布和演化历史的研究, 如探讨葡萄科(Vitaceae)乌蔹莓属(Cayratia)在旧世界间断分布的成因(Lu et al., 2013), 探讨棕榈类植物形成全球热带地区间断分布的过程(Baker & Couvreur, 2013)以及鸟类在全球范围内分布区系的形成历史(Jetz et al., 2012).另外, 也有研究关注不同地理区域间物种相似性及其形成历史, 如讨论东亚和北美地区被子植物的相似性及两区域间物种的迁移历史等(Wen et al., 2010). ...
The origins of modern biodiversity on land.
1
2010
... 具有时间标定的系统发育树(时间树)不仅可以揭示生物的地理分布格局和历史, 还可以关联各种地球环境因子以研究生物的起源和多样化过程(FitzJohn et al., 2009; Benton, 2010; Hoorn et al., 2010; Stadler, 2011).近年来生命科学、地球科学、古气候和古地质学等多学科领域的交叉和渗透, 大大促进了生物多样化研究的发展(Near et al., 2005; Wu et al., 2012; Benton, 2013).多样化分析不仅能够阐明生命演化过程所展现出的不同式样, 而且可以揭示参与塑造这一过程的诸多因素及生物对不同地史时期地质变迁和气候变化的响应(Donoghue, 2008; Gehrke & Linder, 2011). ...
Origins of biodiversity.
1
2013
... 具有时间标定的系统发育树(时间树)不仅可以揭示生物的地理分布格局和历史, 还可以关联各种地球环境因子以研究生物的起源和多样化过程(FitzJohn et al., 2009; Benton, 2010; Hoorn et al., 2010; Stadler, 2011).近年来生命科学、地球科学、古气候和古地质学等多学科领域的交叉和渗透, 大大促进了生物多样化研究的发展(Near et al., 2005; Wu et al., 2012; Benton, 2013).多样化分析不仅能够阐明生命演化过程所展现出的不同式样, 而且可以揭示参与塑造这一过程的诸多因素及生物对不同地史时期地质变迁和气候变化的响应(Donoghue, 2008; Gehrke & Linder, 2011). ...
Reconstructing web evolution and spider diversification in the molecular era.
1
2009
... 生物类群所栖息的生态环境的变化是物种的快速多样化所必需的外因条件, 然而外界环境的剧变并不能直接导致物种的快速多样化, 类群本身还必须通过进化创新来响应环境的变化(Renaud et al., 2005; Blackledge et al., 2009; Burbrink et al., 2012; Puzey et al., 2012).只有阐明了关键创新和生态机会之间的相互联系, 才能真正理解物种多样性的进化历史或动态变化规律.生命之树是连接生物自身与外界环境的桥梁.在探讨协同演化的问题时需要比较两棵树(如宿主和寄主, 植物和传粉者)的分化时间, 才能确定两者之间是否真的存在着协同演化关系(Cruaud et al., 2012).基于生命之树的形态性状重建发现, 毛茛科(Ranunculaceae)耧斗菜属(Aquilegia)植物蜜腺距的长度和弯曲式样与传粉者密切相关, 随着传粉者从蜂类到蜂鸟、天蛾的转变, 其距的长度也呈现伸长的趋势; 此外, 花被的颜色和花的生长方向也随着传粉者的偏好而不同.正是由于发展了创新性状来适应不同的传粉者和生境, 耧斗菜属植物在北美才发生了快速多样化(Hodges & Derieg, 2009; Kramer, 2009). ...
The origins and diversification of C4 grasses and savanna-adapted ungulates.
1
2009
... 一个分类群物种多样性高可能是由物种的快速多样化(Haffer, 1969)、较低的灭绝速率(Stebbins, 1974)或古老支系多样性的长期积累造成的(Couvreur et al., 2011).目前有关物种快速多样化的研究多数归因于生态机会(ecological opportunity), 如气候变化、地质变化、栖息地和生境的转变及外界生物因素等(Bouchenak-Khelladi et al., 2009; Yoder et al., 2010; Hou et al., 2011; Schweizer et al., 2011).在生命演化过程中, 全球气候的变化常常导致新的生态系统的形成, 从而造成部分物种的绝灭和新物种的产生.如白垩纪晚期, 小行星撞击地球引起的全球变冷和光照不足等关键环境因素的改变致使陆地生态系统发生了巨大变化, 最终导致了著名的K-Pg Boundary生物大灭绝事件(Schulte et al., 2010).许多动、植物类群在K-Pg Boundary之后占据了空白生态位, 发生了辐射进化, 从而形成了现今的多样化格局.O’Lear等(2013)认为白垩纪晚期恐龙的绝灭、地球环境的变化及食谱的变化促进了有胎盘类哺乳动物在K-Pg Boundary之后发生快速辐射.Töpel等(2012)的研究认为古新世以来, 北美地区的气候变化促进了蔷薇科(Rosaceae)植物的快速多样化.近年来也有利用某一植被类型指示成分的多样化来探讨该植被类型的起源与演化的研究.如: De-Nova等(2012)对裂榄属(Bursera)的研究发现, 中美洲季节性干旱森林的演化与中新世气候的干旱密切相关; Wang等(2012)通过研究热带雨林指示植物防己科(Menispermaceae)的起源和多样化, 推测现代热带雨林于约6,500万年前在3个热带区域几乎同时出现. ...
When are adaptive radiations replicated in areas? Ecological opportunity and unexceptional diversification in West Indian dipsadine snakes (Colubridae: Alsophiini).
1
2012
... 生物类群所栖息的生态环境的变化是物种的快速多样化所必需的外因条件, 然而外界环境的剧变并不能直接导致物种的快速多样化, 类群本身还必须通过进化创新来响应环境的变化(Renaud et al., 2005; Blackledge et al., 2009; Burbrink et al., 2012; Puzey et al., 2012).只有阐明了关键创新和生态机会之间的相互联系, 才能真正理解物种多样性的进化历史或动态变化规律.生命之树是连接生物自身与外界环境的桥梁.在探讨协同演化的问题时需要比较两棵树(如宿主和寄主, 植物和传粉者)的分化时间, 才能确定两者之间是否真的存在着协同演化关系(Cruaud et al., 2012).基于生命之树的形态性状重建发现, 毛茛科(Ranunculaceae)耧斗菜属(Aquilegia)植物蜜腺距的长度和弯曲式样与传粉者密切相关, 随着传粉者从蜂类到蜂鸟、天蛾的转变, 其距的长度也呈现伸长的趋势; 此外, 花被的颜色和花的生长方向也随着传粉者的偏好而不同.正是由于发展了创新性状来适应不同的传粉者和生境, 耧斗菜属植物在北美才发生了快速多样化(Hodges & Derieg, 2009; Kramer, 2009). ...
Genome-scale phylogenetics: inferring the plant tree of life from 18,896 gene trees.
1
2011
... 随着测序技术的进步和越来越多的分子片段应用于系统发育重建, 随之产生的基因组间、基因间的建树冲突成为系统发育研究中日益突出的问题(邹新慧和葛颂, 2008).为此, 生物学家们提出许多理论来解释这些冲突, 如杂交/渐渗、基因水平转移和谱系筛选等, 并创建了许多新的方法和模型以重建较为真实、可靠的系统发育关系(Liu, 2008; Burleigh et al., 2011; Song et al., 2012).当然这并不说明物种树比基因树更重要, 因为在研究多基因家族的进化时, 基因树更有助于了解成员基因的进化历史和基因重复过程(Nei & Kumar, 2000). ...
Phylogenetic diversity metrics for ecological communities: integrating species richness, abundance and evolutionary history.
2
2010
... 目前, 已经有不少学者依据系统发育多样性指数对部分生物多样性热点地区展开了研究, 并提出一系列相关的分析方法(Redding & Mooers, 2006; Cadotte et al., 2010).例如, 南非好望角有明显的东西部植被分区界限, 在西部, 由于全年降雨(包括冬季), 其植被覆盖度是东部的两倍之多, 在物种丰富度上极具优势.Forest等(2007)利用rbcL序列重建了好望角735属本土特有物种的生命之树, 发现物种数量丰富和类群多样的区域在系统发育多样性上不一定占优势.因此, 在制定生物多样性保护策略时不仅要依据物种丰富度, 还要考虑生物在基因和功能等方面特有的进化属性.另外, 在系统发育多样性指数(Faith, 1992)和进化特异性指数(Isaac et al., 2007)的基础上, 系统发育特有性指数(Rosauer et al., 2009)和地理空间权重进化特异性指数(Cadotte et al., 2010)分别被提出, 完善了生物多样性的评估指标. ...
... )和地理空间权重进化特异性指数(Cadotte et al., 2010)分别被提出, 完善了生物多样性的评估指标. ...
The merging of community ecology and phylogenetic biology.
On Barcode of Life and Tree of Life. Plant Diversity and Resources (植物分类与资源学报
1
2013
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
Oligocene CO2 decline promoted C4 photosynthesis in grass.
1
2008
... 由于测序技术的快速发展, 尤其是二代测序技术(如Roche 454, Illumina Solexa, ABI SOLiD)的日益成熟, 大规模分子数据的获取已经不再是进行系统发育分析的难题, 因此系统学领域对大尺度系统发育树的构建也逐渐关注(Bininda-Emonds, 2004; Cracraft & Donoghue, 2004; Hodkinson & Parnell, 2006).基于大量物种取样所进行的系统发育研究已经解决了许多进化上的问题, 例如类群的起源、快速辐射以及多样化过程等(Bininda-Emonds et al., 2007; Christin et al., 2008; Magallón & Castillo, 2009; Springer et al., 2012).目前, 构建超大数据系统发育树主要有两类方法: 超树方法和超矩阵方法(Sanderson et al., 1998; De Queiroz & Gatesy, 2007).超树方法利用一系列基于不同基因、具有部分共同物种的基因树, 得到最终的物种树; 而超矩阵方法直接将各类数据整合为“supermatrix”而构建物种树, 目前得到了更广泛的应用(Marjoram & Tavaré, 2006). ...
Chloroplast gene sequences and the study of plant evolution.
... 南水青冈属(Nothofagus)一直被视为理解南半球冈瓦纳大陆板块分裂过程的关键类群(Raven, 1996).根据化石记录, 该属植物在冈瓦纳大陆分裂前就已经出现并广泛分布于南半球(Cook & Crisp, 2005).随着分子系统学的发展, 许多研究利用叶绿体、线粒体及核基因序列重建了南水青冈属的系统关系(Martin & Dowd, 1993; Manos & Steele, 1997; Jordan & Hill, 1999; Li et al., 2004), 为该属的生物地理学研究提供了新的线索.分子钟结果显示, 新西兰和澳大利亚的南水青冈物种分化大约发生于渐新世, 晚于澳大利亚和南美洲物种的分化时间, 这与新西兰约8,000万年前与冈瓦纳大陆分离, 而澳大利亚和南美洲约3,500万年前才与冈瓦纳大陆分离的地质事件不符, 说明长距离传播在该属的物种形成过程中发挥了更为重要的作用(Kodandar- amaiah, 2010; Crisp et al., 2011).由此可见, 南水青冈的生物地理学历史是极为复杂的过程, 隔离在演化早期发挥了一定作用, 而后伴随的多次长距离传播和灭绝事件是导致该属现存类群分布格局的重要因素(Manos, 1997; Knapp et al., 2005; Kodanda- ramaiah, 2010; Crisp et al., 2011). ...
... ; Crisp et al., 2011). ...
The Evolution and Classification of Flowering Plants, 2nd edn. New York Botanical Garden
1
1988
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
An extreme case of plant-insect codiversification: figs and fig-pollinating wasps.
1
2012
... 生物类群所栖息的生态环境的变化是物种的快速多样化所必需的外因条件, 然而外界环境的剧变并不能直接导致物种的快速多样化, 类群本身还必须通过进化创新来响应环境的变化(Renaud et al., 2005; Blackledge et al., 2009; Burbrink et al., 2012; Puzey et al., 2012).只有阐明了关键创新和生态机会之间的相互联系, 才能真正理解物种多样性的进化历史或动态变化规律.生命之树是连接生物自身与外界环境的桥梁.在探讨协同演化的问题时需要比较两棵树(如宿主和寄主, 植物和传粉者)的分化时间, 才能确定两者之间是否真的存在着协同演化关系(Cruaud et al., 2012).基于生命之树的形态性状重建发现, 毛茛科(Ranunculaceae)耧斗菜属(Aquilegia)植物蜜腺距的长度和弯曲式样与传粉者密切相关, 随着传粉者从蜂类到蜂鸟、天蛾的转变, 其距的长度也呈现伸长的趋势; 此外, 花被的颜色和花的生长方向也随着传粉者的偏好而不同.正是由于发展了创新性状来适应不同的传粉者和生境, 耧斗菜属植物在北美才发生了快速多样化(Hodges & Derieg, 2009; Kramer, 2009). ...
An updated angiosperm classification.
1
1989
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray
1859
Insights into the historical construction of species-rich Mesoamerican seasonally dry tropical forests: the diversification of Bursera (Burseraceae, Sapindales).
2012
Gene tree discordance, phylogenetic inference and the multispecies coalescent.
4
2009
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
MUSCLE: multiple sequence alignment with high accuracy and high throughput.
1
2004
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
Conservation evaluation and phylogenetic diversity.
Next-generation museomics disentangles one of the largest primate radiations.
1
2013
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
Speciation in Amazonian forest birds.
1
1969
... 一个分类群物种多样性高可能是由物种的快速多样化(Haffer, 1969)、较低的灭绝速率(Stebbins, 1974)或古老支系多样性的长期积累造成的(Couvreur et al., 2011).目前有关物种快速多样化的研究多数归因于生态机会(ecological opportunity), 如气候变化、地质变化、栖息地和生境的转变及外界生物因素等(Bouchenak-Khelladi et al., 2009; Yoder et al., 2010; Hou et al., 2011; Schweizer et al., 2011).在生命演化过程中, 全球气候的变化常常导致新的生态系统的形成, 从而造成部分物种的绝灭和新物种的产生.如白垩纪晚期, 小行星撞击地球引起的全球变冷和光照不足等关键环境因素的改变致使陆地生态系统发生了巨大变化, 最终导致了著名的K-Pg Boundary生物大灭绝事件(Schulte et al., 2010).许多动、植物类群在K-Pg Boundary之后占据了空白生态位, 发生了辐射进化, 从而形成了现今的多样化格局.O’Lear等(2013)认为白垩纪晚期恐龙的绝灭、地球环境的变化及食谱的变化促进了有胎盘类哺乳动物在K-Pg Boundary之后发生快速辐射.Töpel等(2012)的研究认为古新世以来, 北美地区的气候变化促进了蔷薇科(Rosaceae)植物的快速多样化.近年来也有利用某一植被类型指示成分的多样化来探讨该植被类型的起源与演化的研究.如: De-Nova等(2012)对裂榄属(Bursera)的研究发现, 中美洲季节性干旱森林的演化与中新世气候的干旱密切相关; Wang等(2012)通过研究热带雨林指示植物防己科(Menispermaceae)的起源和多样化, 推测现代热带雨林于约6,500万年前在3个热带区域几乎同时出现. ...
BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.
1
1999
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
Comparison of the accuracies of several phylogenetic methods using protein and DNA sequences.
Estimating ancestral distributions of lineages with uncertain sister groups: a statistical approach to Dispersal-Vicariance Analysis and a case using Aesculus L. (Sapindaceae) including fossils.
... 由于测序技术的快速发展, 尤其是二代测序技术(如Roche 454, Illumina Solexa, ABI SOLiD)的日益成熟, 大规模分子数据的获取已经不再是进行系统发育分析的难题, 因此系统学领域对大尺度系统发育树的构建也逐渐关注(Bininda-Emonds, 2004; Cracraft & Donoghue, 2004; Hodkinson & Parnell, 2006).基于大量物种取样所进行的系统发育研究已经解决了许多进化上的问题, 例如类群的起源、快速辐射以及多样化过程等(Bininda-Emonds et al., 2007; Christin et al., 2008; Magallón & Castillo, 2009; Springer et al., 2012).目前, 构建超大数据系统发育树主要有两类方法: 超树方法和超矩阵方法(Sanderson et al., 1998; De Queiroz & Gatesy, 2007).超树方法利用一系列基于不同基因、具有部分共同物种的基因树, 得到最终的物种树; 而超矩阵方法直接将各类数据整合为“supermatrix”而构建物种树, 目前得到了更广泛的应用(Marjoram & Tavaré, 2006). ...
Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity.
1
2010
... 具有时间标定的系统发育树(时间树)不仅可以揭示生物的地理分布格局和历史, 还可以关联各种地球环境因子以研究生物的起源和多样化过程(FitzJohn et al., 2009; Benton, 2010; Hoorn et al., 2010; Stadler, 2011).近年来生命科学、地球科学、古气候和古地质学等多学科领域的交叉和渗透, 大大促进了生物多样化研究的发展(Near et al., 2005; Wu et al., 2012; Benton, 2013).多样化分析不仅能够阐明生命演化过程所展现出的不同式样, 而且可以揭示参与塑造这一过程的诸多因素及生物对不同地史时期地质变迁和气候变化的响应(Donoghue, 2008; Gehrke & Linder, 2011). ...
Eocene habitat shift from saline to freshwater promoted Tethyan amphipod diversification.
1
2011
... 一个分类群物种多样性高可能是由物种的快速多样化(Haffer, 1969)、较低的灭绝速率(Stebbins, 1974)或古老支系多样性的长期积累造成的(Couvreur et al., 2011).目前有关物种快速多样化的研究多数归因于生态机会(ecological opportunity), 如气候变化、地质变化、栖息地和生境的转变及外界生物因素等(Bouchenak-Khelladi et al., 2009; Yoder et al., 2010; Hou et al., 2011; Schweizer et al., 2011).在生命演化过程中, 全球气候的变化常常导致新的生态系统的形成, 从而造成部分物种的绝灭和新物种的产生.如白垩纪晚期, 小行星撞击地球引起的全球变冷和光照不足等关键环境因素的改变致使陆地生态系统发生了巨大变化, 最终导致了著名的K-Pg Boundary生物大灭绝事件(Schulte et al., 2010).许多动、植物类群在K-Pg Boundary之后占据了空白生态位, 发生了辐射进化, 从而形成了现今的多样化格局.O’Lear等(2013)认为白垩纪晚期恐龙的绝灭、地球环境的变化及食谱的变化促进了有胎盘类哺乳动物在K-Pg Boundary之后发生快速辐射.Töpel等(2012)的研究认为古新世以来, 北美地区的气候变化促进了蔷薇科(Rosaceae)植物的快速多样化.近年来也有利用某一植被类型指示成分的多样化来探讨该植被类型的起源与演化的研究.如: De-Nova等(2012)对裂榄属(Bursera)的研究发现, 中美洲季节性干旱森林的演化与中新世气候的干旱密切相关; Wang等(2012)通过研究热带雨林指示植物防己科(Menispermaceae)的起源和多样化, 推测现代热带雨林于约6,500万年前在3个热带区域几乎同时出现. ...
Identifying hotspots of endemic woody seed plant diversity in China.
1
2012
... 生物多样性是生物与环境形成的生态复合体以及与此相关的各种生态过程的总和, 重要的有遗传多样性、物种多样性和生态系统多样性三个层次(Hawksworth, 1996).近年来, 中国在生物多样性研究领域取得了重要进展(马克平, 2013), 如对于中国特有木本种子植物多样性的研究(如Huang et al., 2012), 基于代谢理论(Metabolic Theory of Ecology)对物种多样性分布规律的解释(如王志恒等, 2009; Wang et al., 2009), 以及对中国山地生物多样性分布格局的研究(Tang et al., 2006).然而, 生物多样性在不同区域丰富度的变化不仅受到该区域的生态环境和历史地理因素(包括隔离、保存、迁移和绝灭)的深刻影响, 而且与生物类群的系统发育和进化过程密切相关.对于物种不均匀分化程度较强的地区, 在解释气候生态因子与生物多样性之间的关联度时, 就要充分考虑到进化过程的影响(Francis & Currie, 2003).如中国东喜马拉雅和横断山地区在1,000万年内迅速抬升, 导致物种的快速分化, 对生物多样性格局产生了巨大影响. ...
Mammals on the EDGE: conservation priorities based on threat and phylogeny.
1
2007
... 目前, 已经有不少学者依据系统发育多样性指数对部分生物多样性热点地区展开了研究, 并提出一系列相关的分析方法(Redding & Mooers, 2006; Cadotte et al., 2010).例如, 南非好望角有明显的东西部植被分区界限, 在西部, 由于全年降雨(包括冬季), 其植被覆盖度是东部的两倍之多, 在物种丰富度上极具优势.Forest等(2007)利用rbcL序列重建了好望角735属本土特有物种的生命之树, 发现物种数量丰富和类群多样的区域在系统发育多样性上不一定占优势.因此, 在制定生物多样性保护策略时不仅要依据物种丰富度, 还要考虑生物在基因和功能等方面特有的进化属性.另外, 在系统发育多样性指数(Faith, 1992)和进化特异性指数(Isaac et al., 2007)的基础上, 系统发育特有性指数(Rosauer et al., 2009)和地理空间权重进化特异性指数(Cadotte et al., 2010)分别被提出, 完善了生物多样性的评估指标. ...
The global diversity of birds in space and time.
1
2012
... 随着生物地理学理论和方法的发展, 该领域研究的对象和关注的问题日益广泛, 包括对某一特定类群散布和演化历史的研究, 如探讨葡萄科(Vitaceae)乌蔹莓属(Cayratia)在旧世界间断分布的成因(Lu et al., 2013), 探讨棕榈类植物形成全球热带地区间断分布的过程(Baker & Couvreur, 2013)以及鸟类在全球范围内分布区系的形成历史(Jetz et al., 2012).另外, 也有研究关注不同地理区域间物种相似性及其形成历史, 如讨论东亚和北美地区被子植物的相似性及两区域间物种的迁移历史等(Wen et al., 2010). ...
Resolving an ancient, rapid radiation in Saxifragales.
1
2008
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
The phylogenetic affinities of Nothofagus (Nothofagaceae) leaf fossils based on combined molecular and morphological data.
1
1999
... 南水青冈属(Nothofagus)一直被视为理解南半球冈瓦纳大陆板块分裂过程的关键类群(Raven, 1996).根据化石记录, 该属植物在冈瓦纳大陆分裂前就已经出现并广泛分布于南半球(Cook & Crisp, 2005).随着分子系统学的发展, 许多研究利用叶绿体、线粒体及核基因序列重建了南水青冈属的系统关系(Martin & Dowd, 1993; Manos & Steele, 1997; Jordan & Hill, 1999; Li et al., 2004), 为该属的生物地理学研究提供了新的线索.分子钟结果显示, 新西兰和澳大利亚的南水青冈物种分化大约发生于渐新世, 晚于澳大利亚和南美洲物种的分化时间, 这与新西兰约8,000万年前与冈瓦纳大陆分离, 而澳大利亚和南美洲约3,500万年前才与冈瓦纳大陆分离的地质事件不符, 说明长距离传播在该属的物种形成过程中发挥了更为重要的作用(Kodandar- amaiah, 2010; Crisp et al., 2011).由此可见, 南水青冈的生物地理学历史是极为复杂的过程, 隔离在演化早期发挥了一定作用, 而后伴随的多次长距离传播和灭绝事件是导致该属现存类群分布格局的重要因素(Manos, 1997; Knapp et al., 2005; Kodanda- ramaiah, 2010; Crisp et al., 2011). ...
MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform.
1
2002
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
Problems due to missing data in phylogenetic analyses including fossils: a critical review.
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
PhyloBayes 3: a Bayesian software package for phylogenetic reconstruction and molecular dating.
Estimating a binary character’s effect on speciation and extinction.
1
2007
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
Absolute diversification rates in angiosperm clades.
1
2001
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
Angiosperm diversification through time.
1
2009
... 由于测序技术的快速发展, 尤其是二代测序技术(如Roche 454, Illumina Solexa, ABI SOLiD)的日益成熟, 大规模分子数据的获取已经不再是进行系统发育分析的难题, 因此系统学领域对大尺度系统发育树的构建也逐渐关注(Bininda-Emonds, 2004; Cracraft & Donoghue, 2004; Hodkinson & Parnell, 2006).基于大量物种取样所进行的系统发育研究已经解决了许多进化上的问题, 例如类群的起源、快速辐射以及多样化过程等(Bininda-Emonds et al., 2007; Christin et al., 2008; Magallón & Castillo, 2009; Springer et al., 2012).目前, 构建超大数据系统发育树主要有两类方法: 超树方法和超矩阵方法(Sanderson et al., 1998; De Queiroz & Gatesy, 2007).超树方法利用一系列基于不同基因、具有部分共同物种的基因树, 得到最终的物种树; 而超矩阵方法直接将各类数据整合为“supermatrix”而构建物种树, 目前得到了更广泛的应用(Marjoram & Tavaré, 2006). ...
Systematics of Nothofagus (Nothofagaceae) based on rDNA spacer sequences (ITS): taxonomic congruence with morphology and plastid sequences.
1997
Phylogenetic analyses of “higher” Hamamelididae based on plastid sequence data.
2
1997
... 南水青冈属(Nothofagus)一直被视为理解南半球冈瓦纳大陆板块分裂过程的关键类群(Raven, 1996).根据化石记录, 该属植物在冈瓦纳大陆分裂前就已经出现并广泛分布于南半球(Cook & Crisp, 2005).随着分子系统学的发展, 许多研究利用叶绿体、线粒体及核基因序列重建了南水青冈属的系统关系(Martin & Dowd, 1993; Manos & Steele, 1997; Jordan & Hill, 1999; Li et al., 2004), 为该属的生物地理学研究提供了新的线索.分子钟结果显示, 新西兰和澳大利亚的南水青冈物种分化大约发生于渐新世, 晚于澳大利亚和南美洲物种的分化时间, 这与新西兰约8,000万年前与冈瓦纳大陆分离, 而澳大利亚和南美洲约3,500万年前才与冈瓦纳大陆分离的地质事件不符, 说明长距离传播在该属的物种形成过程中发挥了更为重要的作用(Kodandar- amaiah, 2010; Crisp et al., 2011).由此可见, 南水青冈的生物地理学历史是极为复杂的过程, 隔离在演化早期发挥了一定作用, 而后伴随的多次长距离传播和灭绝事件是导致该属现存类群分布格局的重要因素(Manos, 1997; Knapp et al., 2005; Kodanda- ramaiah, 2010; Crisp et al., 2011). ...
... ).由此可见, 南水青冈的生物地理学历史是极为复杂的过程, 隔离在演化早期发挥了一定作用, 而后伴随的多次长距离传播和灭绝事件是导致该属现存类群分布格局的重要因素(Manos, 1997; Knapp et al., 2005; Kodanda- ramaiah, 2010; Crisp et al., 2011). ...
Difference of Relationships Between Spatial Pattern and Environment for Seed Plant Diversity in China (中国种子植物多样性的空间格局-环境关系分异研究). PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing
Reconciling molecular phylogenies with the fossil record.
1
2011
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
Assessing concordance of fossil calibration points in molecular clock studies: an example using turtles.
1
2005
... 具有时间标定的系统发育树(时间树)不仅可以揭示生物的地理分布格局和历史, 还可以关联各种地球环境因子以研究生物的起源和多样化过程(FitzJohn et al., 2009; Benton, 2010; Hoorn et al., 2010; Stadler, 2011).近年来生命科学、地球科学、古气候和古地质学等多学科领域的交叉和渗透, 大大促进了生物多样化研究的发展(Near et al., 2005; Wu et al., 2012; Benton, 2013).多样化分析不仅能够阐明生命演化过程所展现出的不同式样, 而且可以揭示参与塑造这一过程的诸多因素及生物对不同地史时期地质变迁和气候变化的响应(Donoghue, 2008; Gehrke & Linder, 2011). ...
The reconstructed evolutionary process.
1
1994
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
1
2000
... 随着测序技术的进步和越来越多的分子片段应用于系统发育重建, 随之产生的基因组间、基因间的建树冲突成为系统发育研究中日益突出的问题(邹新慧和葛颂, 2008).为此, 生物学家们提出许多理论来解释这些冲突, 如杂交/渐渗、基因水平转移和谱系筛选等, 并创建了许多新的方法和模型以重建较为真实、可靠的系统发育关系(Liu, 2008; Burleigh et al., 2011; Song et al., 2012).当然这并不说明物种树比基因树更重要, 因为在研究多基因家族的进化时, 基因树更有助于了解成员基因的进化历史和基因重复过程(Nei & Kumar, 2000). ...
MrModeltest. Version 2. Program distributed by the author. Evolutionary Biology Center, Uppsala University
Nonuniform concerted evolution and chloroplast capture: heterogeneity of observed introgression patterns in three molecular data partition phylogenies of Asian Mitella (Saxifragaceae).
Evolution of spur-length diversity in Aquilegia petals is achieved solely through cell-shape anisotropy.
1
2012
... 生物类群所栖息的生态环境的变化是物种的快速多样化所必需的外因条件, 然而外界环境的剧变并不能直接导致物种的快速多样化, 类群本身还必须通过进化创新来响应环境的变化(Renaud et al., 2005; Blackledge et al., 2009; Burbrink et al., 2012; Puzey et al., 2012).只有阐明了关键创新和生态机会之间的相互联系, 才能真正理解物种多样性的进化历史或动态变化规律.生命之树是连接生物自身与外界环境的桥梁.在探讨协同演化的问题时需要比较两棵树(如宿主和寄主, 植物和传粉者)的分化时间, 才能确定两者之间是否真的存在着协同演化关系(Cruaud et al., 2012).基于生命之树的形态性状重建发现, 毛茛科(Ranunculaceae)耧斗菜属(Aquilegia)植物蜜腺距的长度和弯曲式样与传粉者密切相关, 随着传粉者从蜂类到蜂鸟、天蛾的转变, 其距的长度也呈现伸长的趋势; 此外, 花被的颜色和花的生长方向也随着传粉者的偏好而不同.正是由于发展了创新性状来适应不同的传粉者和生境, 耧斗菜属植物在北美才发生了快速多样化(Hodges & Derieg, 2009; Kramer, 2009). ...
Angiosperm phylogeny inferred from sequences of four mitochondrial genes.
1
2010
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
a) Likelihood methods for detecting temporal shifts in diversification rates.
1
2006
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
b) LASER: a maximum likelihood toolkit for detecting temporal shifts in diversification rates from molecular phylogenies.
1
2006
... 目前, 生物多样化分析主要基于Birth-death模型.该模型假定物种在固定时间间隔内以恒定的概率发生分化与灭绝(Nee, 1994), 通过基于似然值的模型分析来判断物种的多样化速率是否发生改变(Rabosky, 2006a).相关分析软件主要包括R语言平台下的APE (Paradis, 2004)和LASER (Rabosky, 2006b)软件包等.在多样化研究中, 取样缺失是一个普遍存在的问题.目前有许多方法致力于研究不同程度取样缺失情况下如何正确评估生物多样化(Magallón & Sanderson, 2001).其中R语言平台下的MEDUSA软件包在解决取样缺失和优化分析结果方面均有良好表现, 成为当下的主流分析方法(Alfaro et al., 2009).目前更多的研究倾向于探寻影响生物多样化的内因与外因, 相应的模型和分析手段也在迅速发展.在R语言环境下的diversitree软件包能综合分析性状、地理分布等因素与生物多样化速率的关系, 有助于进一步阐明生物多样化的规律(Maddison et al., 2007; Goldberg et al., 2011). ...
Se-Al: Sequence Alignment Editor. Available at
1
2002
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... Phylogenetic relationships of the angiosperms at the ordinal level (based on Soltis et al., 2011). Names of the families, orders and other major clades follow APG III (2009) and Cantino et al. (2007). Numbers above branches are Maximum Likelihood bootstrap values. ...
Angiosperm phylogeny: a framework for studies of genome evolution.
3
2013
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
Angiosperm Phylogeny Website. Version 9, June 2008. Available at
1
2001
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
The Study on Molecular Phylogeny of Rosids (蔷薇分支的分子系统学研究). PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing
1
2012
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
Phylogentic placement of two enigmatic genera, Borthwickia and Stixis, based on molecular and pollen data, and the description of a new family of Brassicales, Borthwickiaceae.
1
2012
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
Paup*: Phylogenetic Anlysis Using Parsimony (*and Other Methods), Version 4.0b10. Sinauer Associates
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.
The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools.
1
1997
... (3)序列比对(alignment).为了保证序列的同源性和所得系统发育关系的可靠性, 需要对原始序列进行比对和校正.自动比对序列的软件包括Clustal (Thompson et al., 1997; Larkin et al., 2007)、MAFFT (Katoh et al., 2002)、MUSCLE (Edgar, 2004)等; 手工校对序列的软件有BioEdit (Hall, 1999)、Se-Al (Rambaut, 2002)、Geneious (created by Biomatters, available at http://www.geneious.com)等. ...
Sparse supermatrices for phylogenetic inference: taxonomy alignment, rogue taxa, and the phylogeny of living turtles.
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
Phy- logeny and classification of Ranunculales: Evidence from four molecular loci and morphological data.
2009
Menispermaceae and the diversification of tropical rainforests near the Cretaceous-Paleogene boundary.
2012
Metabolic theory of ecology: an explanation for species richness patterns based on the metabolic processes of organisms.
1
2009
... 生物多样性是生物与环境形成的生态复合体以及与此相关的各种生态过程的总和, 重要的有遗传多样性、物种多样性和生态系统多样性三个层次(Hawksworth, 1996).近年来, 中国在生物多样性研究领域取得了重要进展(马克平, 2013), 如对于中国特有木本种子植物多样性的研究(如Huang et al., 2012), 基于代谢理论(Metabolic Theory of Ecology)对物种多样性分布规律的解释(如王志恒等, 2009; Wang et al., 2009), 以及对中国山地生物多样性分布格局的研究(Tang et al., 2006).然而, 生物多样性在不同区域丰富度的变化不仅受到该区域的生态环境和历史地理因素(包括隔离、保存、迁移和绝灭)的深刻影响, 而且与生物类群的系统发育和进化过程密切相关.对于物种不均匀分化程度较强的地区, 在解释气候生态因子与生物多样性之间的关联度时, 就要充分考虑到进化过程的影响(Francis & Currie, 2003).如中国东喜马拉雅和横断山地区在1,000万年内迅速抬升, 导致物种的快速分化, 对生物多样性格局产生了巨大影响. ...
Temperature dependence, spatial scale, and tree species diversity in eastern Asia and North America.
3
2009
... 研究生物类群之间的亲缘关系一直以来都是生命之树最直接和最根本的用途(陈之端和李德铢, 2013).生命之树的发展历史就是人们利用不同手段研究生物类群间系统关系的过程.随着测序技术和生物信息技术的进步, 通过广泛地利用多源生物信息, 包括形态、化石、分子等数据, 建立所有有机体的生命之树已成为系统发育研究的重要方面(Morlon et al., 2011).例如, 自20世纪80年代以来, 根据形态性状, 完善的被子植物分类系统相继被提出(如Cronquist, 1988; Dahlgren, 1989; Takhtajan, 1997).到了20世纪90年代, 人们开始利用分子数据, 特别是DNA序列, 来探讨被子植物各大类群之间的关系.随着植物分子系统学的快速发展, 系统学家针对被子植物大尺度的系统发育分析提出了APG系统, 并不断对其进行修订(APG, 1998; APG II, 2003; APG III, 2009); 同时也有大量针对目、科、属级水平的专门研究(Stevens, 2001; Wang et al., 2009; Su et al., 2012).这些不同尺度的研究为重建生命之树积累了丰富的分子数据和理论借鉴.目前被子植物系统在目、科级水平上的框架基本稳定(Soltis et al., 2011; 图1), 仅有部分类群的系统位置或内部关系不确定、支持率不高, 或根据不同基因组数据得出的拓扑结构不一致, 如第伦桃科(Dilleniaceae)、金虎尾目(Malpighiales)等(Soltis & Soltis, 2013). ...
... 生物多样性是生物与环境形成的生态复合体以及与此相关的各种生态过程的总和, 重要的有遗传多样性、物种多样性和生态系统多样性三个层次(Hawksworth, 1996).近年来, 中国在生物多样性研究领域取得了重要进展(马克平, 2013), 如对于中国特有木本种子植物多样性的研究(如Huang et al., 2012), 基于代谢理论(Metabolic Theory of Ecology)对物种多样性分布规律的解释(如王志恒等, 2009; Wang et al., 2009), 以及对中国山地生物多样性分布格局的研究(Tang et al., 2006).然而, 生物多样性在不同区域丰富度的变化不仅受到该区域的生态环境和历史地理因素(包括隔离、保存、迁移和绝灭)的深刻影响, 而且与生物类群的系统发育和进化过程密切相关.对于物种不均匀分化程度较强的地区, 在解释气候生态因子与生物多样性之间的关联度时, 就要充分考虑到进化过程的影响(Francis & Currie, 2003).如中国东喜马拉雅和横断山地区在1,000万年内迅速抬升, 导致物种的快速分化, 对生物多样性格局产生了巨大影响. ...
... 近十年来, 基于生命之树框架的系统发育多样性(phylogenetic diversity, PD)研究备受关注(Mace et al., 2003; Santamaria & Mendez, 2012; Winter et al., 2013).系统发育多样性也称作进化多样性(evolutionary diversity), 是基于系统发育关系(或物种的进化历史)衡量物种间距离, 对某一特定关联类群进化的独特性或基因和功能的多样性进行评估的重要指标(Hartmann & Andre, 2013; Winter et al., 2013).对于有限的保护资源来说, 系统发育多样性保护策略比盲目地汇集或单纯保护特有物种的数量更有效(Faith, 1992).系统发育多样性不仅能提供许多物种多样性无法反映的重要信息, 还可以准确评估和预测生态系统的稳定性, 对于未来生物多样性中心的确定具有重要指导意义(Vane-Wright et al., 1991; Faith, 1992).根据物种的分类阶元, 可以建立科水平(APG, 1998; APG II, 2003; APGIII, 2009)、属水平(Wang et al., 2009; 苏俊霞, 2012)、种水平(DNA条形码)及居群水平的生命之树(任保青和陈之端, 2010).不同层次生命之树的构建将为全面评估与预测生物多样性提供更好的平台. ...
Phylocom: software for the analysis of phylogenetic community structure and trait evolution.
2008
Phylogenies and community ecology.
2002
Phylomatic: tree assembly for applied phylogenetics.
Integrated fossil and molecular data reveal the biogeographic diversification of the eastern Asian-eastern North American disjunct hickory genus (Carya Nutt.).
1
2013
... 目前化石标定应用最为广泛(Sauquet, 2013; Zhang et al., 2013).除了利用化石对相关节点进行最小时间限定外, 还需对生命之树进行一个最大时间的限定, 从而保证整个生命之树的所有节点在一个较为合理的时间尺度上评估. ...
Highly conserved low-copy nuclear genes as effective markers for phylogenetic analyses in angiosperms.
1
2012
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
Spatial patterns of wood traits in China are controlled by phylogeny and the environment.
2011
Mitochondrial matR sequences help to resolve deep phylogenetic relationships in rosids.
1
2007
... 随着各类生物系统发育大框架的基本确立, 人们开始较多地关注生命之树中系统关系支持率不高或存在冲突的疑难类群, 如被子植物中的虎耳草目(Fishbein et al., 2001; Jian et al., 2008)、蔷薇类的COM分支(Zhu et al., 2007; Qiu et al., 2010; Zhang et al., 2012; Soltis & Soltis, 2013), 动物类群中的蜥蜴(Fu, 2000)、长尾猴(Guschanski et al., 2013)等.目前已有不少探讨系统发育关系冲突原因及其解决方法的研究和综述(Maddison, 1997; Wendel & Doyle, 1998; Delsuc et al., 2005; Degnan & Rosenberg, 2009).Seelanan等(1997)根据冲突的显著与否, 将造成系统发育关系冲突的原因分为软冲突(soft incongruence)和硬冲突(hard incongruence).软冲突主要包括人为因素和序列因素, 通常是由各种分析方法或实验设计(如取样不足)的缺陷所引起的数据异质性、系统发育信息“噪音”或非同源相似(homoplasy)而得出错误的基因树; 硬冲突一般出现在来自不同基因组的基因之间, 或者非连锁的核基因之间, 常常来自生物过程本身, 如杂交/渐渗、谱系筛选、基因水平转移、基因重复/丢失、基因重组等(Rokas & Carroll, 2006; Galtier & Daubin, 2008; Degnan & Rosenberg, 2009; Simpson, 2012).导致系统发育关系冲突的原因与对应的解决方案见表1. ...
Conflicting gene trees and phylogenomics.
1
2008
... 随着测序技术的进步和越来越多的分子片段应用于系统发育重建, 随之产生的基因组间、基因间的建树冲突成为系统发育研究中日益突出的问题(邹新慧和葛颂, 2008).为此, 生物学家们提出许多理论来解释这些冲突, 如杂交/渐渗、基因水平转移和谱系筛选等, 并创建了许多新的方法和模型以重建较为真实、可靠的系统发育关系(Liu, 2008; Burleigh et al., 2011; Song et al., 2012).当然这并不说明物种树比基因树更重要, 因为在研究多基因家族的进化时, 基因树更有助于了解成员基因的进化历史和基因重复过程(Nei & Kumar, 2000). ...



{kind=link}
{kind=link}