Advances and challenges in resolving the angiosperm phylogeny
Liping Zeng, Ning Zhang, Hong Ma*,
State Key Laboratory of Genetic Engineering and Institute of Plant Biology, Center for Evolutionary Biology, School of Life Sciences, Fudan University, Shanghai 200433
Angiosperm phylogenetics investigates the evolutionary history and relationships of angiosperms based on the construction of phylogenetic trees. Since the 1990s, nucleotide or amino acid sequences have been widely used for this and angiosperm phylogenetic analysis has advanced from using single or a combination of a few organellar genes to whole plastid genome sequences, resulting in the widely accepted modern molecular systematics of angiosperms. The current framework of the angiosperm phylogeny includes highly supported basal angiosperm relationships, five major clades (eudicots, monocots, magnoliids, Chloranthales, and Ceratophyllales), orders grouped within these clades, and core groups in the monocots or eudicots. However, organellar genes have some limitations; these involve uniparental inheritance in most instances and a relatively low percentage of phylogenetic informative sites. Thus, they are unable to resolve some relationships even when whole plastid genome sequences are used. Therefore, the utility of biparentally inherited nuclear genes with more information about evolutionary history, has gradually received more attention. Nevertheless, there are still some plant groups that are difficult to place in the angiosperm phylogeny, such as those involving the relative positions of the five major groups as well as those of several orders of eudicots. In this review, we discuss the applications, advantages and disadvantages of marker genes, the deep relationships that have been resolved in angiosperm phylogeny, groups with uncertain positions, and the challenges that remain in resolving an accurate phylogeny for angiosperms.
Fig. 1 The relationships of major lineages of angiosperms. Except for three basal orders, all angiosperms formed a monophyletic group composed of five major lineages: Chloranthaceae, magnoliids, monocots, Ceratophyllaceae and eudicots. However, their relationships are not clear. Among monocots, Acorales is the basal order and commelinids (shown as triangle in this figure) with four orders are the core groups. The relationships among four magnoliid orders have been clarified. In eudicots, Ranunculales is sister to all other eudicots and Gunnerales is sister to all other core eudicots, which formed a monophyletic group known as Pentapetalae. Rosids and asterids are core groups of Pentapetalae, but there are many uncertain relationships in Pentapetalae. In rosids, it is still uncertain whether the sister group of the COM clade is the nitrogen-fixing clade or malvids.
2.1 被子植物的基部类群
目前被子植物系统发育框架已经基本形成, 已知的现存物种隶属于哪个类群已经基本清楚, 另外无油樟目、睡莲目和木兰藤目依次被认为是现存被子植物的最基部类群, 也被称为“ANITA”(即Amborellaceae, Nymphaeaceae, Illiciaceae, Trimeniaceae和Austrobaileyaceae的缩写)(Qiu et al., 1999; Bremer et al., 2009)。大多数研究表明, 无油樟目是现存被子植物中最先分化出来的类群, 此目仅含1个物种(Amborella trichopoda), 为单科单属种, 现存的自然群落只在新喀里多尼亚发现(New Caledonia)。但有少数研究结果也把无油樟目和睡莲目形成姊妹群放在被子植物最基部(Barkman et al., 2000; Soltis & Soltis, 2004; Qiu et al., 2005)。从形态学上来看, 上述基部被子植物的种子都具2枚子叶。从系统发育树上看, 单子叶植物作为单系群嵌在传统意义上的双子叶植物内部, 因此, 传统意义上的双子叶植物就不再是单系群。这些结果也说明被子植物祖先的子叶数目可能是2枚, 而后在单子叶植物起源前后丢失成为1枚, 其他被子植物类群则保留了祖先中子叶的数目。
除了ANITA基部类群外, 约占被子植物99.95%的其他类群聚为单系, 此类群包括数量不等的5个分支, 即真双子叶植物、单子叶植物(monocots)、木兰类(magnoliids)、金鱼藻目(Ceratophyllales)和金粟兰目(Chloranthales)(Cantino et al., 2007)。真双子叶植物为五个分支中物种多样性最丰富的类群, 包含了约40个目300余科, 约占整个被子植物的75%, 该类群的共同形态特征为花粉粒具有三孔沟(tricolpate pollen), 因此也被称为“三孔花粉组”(Doyle & Hotton, 1991)。真双子叶植物起源古老, 在中国发现的早白垩纪化石中就存在该类群植物(Drinnan et al., 1991); Sun et al., 2011), 该类群的主要物种在约八千多万年前的晚白垩纪桑托期(late Santonian)就已经形成(Herendeen et al., 1999)。
2.2 真双子叶植物的系统发育关系
真双子叶植物中, 毛茛目(Ranunculales)位于最基部, 是其他所有真双子叶植物的姊妹群; 清风藤科(Sabiaceae)、山龙眼目(Proteales)、黄杨目(Buxales)和昆栏树目(Trochodendrales)是真双子叶植物的其他基部类群; 其余所有真双子叶物种形成单系, 被命名为核心真双子叶(core eudicots)植物(Magallón et al., 1999)。在核心真双子叶植物中, 洋二仙草目(Gunnerales)是其他类群的姊妹群; 其他类群的花多是五基数, 因此也被称为Pentapetalae, 主要由蔷薇类(rosids)和菊类植物(asterids)两大类群组成, 另外还包含系统位置尚未完全确定的几个类群, 即虎耳草目(Saxifragales)、葡萄目(Vitales)、檀香目(Santalales)、五桠果科(Dilleniaceae)和石竹目(Caryophyllales)(Judd & Olmstead, 2004; Worberg et al., 2007; Moore et al., 2010)。
在蔷薇类植物中, 主要类群为固氮分支(nitrogen-fixing clade), 包括葫芦目(Cucurbitales)、壳斗目(Fagales)、蔷薇目(Rosales)和豆目(Fabales), 是支持率很高的单系群; 另外两个高支持率的分支是COM分支和锦葵类(malvids)分支, 前者包括卫矛目(Celastrales)、酢浆草目(Oxalidales)和金虎尾目(Malpighiales), 后者包括锦葵目(Malvales)、包含模式植物拟南芥的十字花目(Brassicales)、燧体木目(Crossosomatales)、无患子目(Sapindales)、十齿花目(Huerteales)和美洲苦木目(Picramniales)(Bremer et al., 2009; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012)(图2a)。在菊类植物中, 大部分物种组成一个被称为真菊的单系, 而山茱萸目(Cornales)和杜鹃花目(Ericales)则是真菊的姊妹群(Bremer et al., 2009; Zhang et al., 2012)(图2b)。APG III又将真菊植物分为真菊I类(也被称为唇形类lamiids)和真菊II类(也被称为桔梗类campanulids); 其中真菊I类的核心类群是唇形目(Lamiales)、茄目(Solanales)、龙胆目(Gentianles)、紫草科(Boraginaceae), 而真菊II类的核心类群是盔瓣花目(Paracryphiales)、川续断目(Dipsacales)、伞形目(Apiales)、鳞叶树目(Bruniales)、菊目(Asterales)和南鼠刺目(Escalloniales)。
Fig. 2 The relationships within rosids, asterids, monocots and magnoliids, respectively. (a) Rosids contain three clades: malvids, COM and the nitrogen-fixing clade, but whether the sister group of the COM clade is the nitrogen-fixing clade or malvids remains unclear. (b) Asterids are composed of euasterid I, euasterid II and two basal orders: Ericales and Cornales. The relationships among euasterids I are not clear. Garryales and Aquifoliales were respectively regarded as the basal group of euasterids I and II; however, new evidence based on nuclear genes suggested that these two together formed a sister group of other euasterids I. (c) Commelinids is the core group of monocots, and Acorales is the basal order. The relationships among commelinids are not clear. (d) In magnollids, Magnoliales and Laurales are sister groups, and Canellales and Piperales are sister groups.
2.3 单子叶植物的系统发育关系
单子叶植物是五大分支中的第二大类群, 约占整个被子植物的22%, 为人类提供了主要的粮食作物(谷物与其他淀粉类作物等)(Bremer, 2000; Stevens, 2001)。该类植物绝大多数为草本, 种子仅有1枚子叶, 平行叶脉, 花基数为3。单子叶植物包含11个目和1个科(Bremer et al., 2009), 其中兰科植物约占单子叶物种的34%, 禾本科植物约占17%。几乎所有的分子证据都表明菖蒲目(Acorales)是单子叶植物的最基部分支, 泽泻目(Alismatales)紧随 其后; 另外, 单子叶植物还包含天门冬目(Aspara-gales)、薯蓣目(Dioscoreales)、百合目(Liliales)、露兜树目(Pandanales)、无叶莲目(Petrosaviales)和鸭跖草类植物(commelinids)。其中鸭跖草类植物是单子叶植物的核心类群, 包括棕榈目(Arecales; 木本)、鸭跖草目(Commelinales)、禾本目(Poales)、姜目(Zingiberales)和多须草科(Dasypogonaceae) (Davis et al., 2004; Graham et al., 2006)(图2c)。单子叶植物是被子植物各大类群中系统发育研究得比较清楚的类群之一, 除了兰科和禾本科外, 几乎其他单子叶植物所有属都曾进行了系统发育研究, 大部分研究基于叶绿体rbcL基因, 这为研究单子叶植物各分支的起源、扩张、重要形态性状的演化与适应等奠定了基础(Soltis et al., 2005)。
测序技术的发展使核基因组和转录组测序变得高效与快速, 测序成本也大大降低, 这为获得多个物种核基因的数据提供了坚实的技术基础, 将使得联合大量基因乃至全基因组数据研究被子植物系统发育关系成为重要趋势, 也将极大地促进核基因系统发育基因组学的研究。不同于利用PCR方法扩增少量基因, 利用基因组或转录组测序能快速地获得大量的基因序列, 可以寻找系统发育信息量和进化速率等适用于研究不同分类阶元亲缘关系的直系同源基因, 应用于系统发育分析, 常用的寻找直系同源基因的分析软件有InParanoid(O'Brien et al., 2005), OrthoMCL(Li et al., 2003)和HaMStR (Ebersberger et al., 2009)等。动物中应用系统发育基因组学解决系统发育问题已经有了很多成功的先例。例如Smith等(2012)获取了15种软体动物的转录组序列, 联合1,185个基因重建了软体动物系统发育树; Kocot等(2011)测定了20种软体动物的转录组序列并联合基因组序列, 利用HaMStR鉴定出308个共有的直系同源基因后, 得到了高支持率的软体动物主要分支间的亲缘关系树; Johnson等(2013)利用基因组数据重建了膜翅目中社会性昆虫蚂蚁、蜜蜂和黄蜂的系统发育树, 发现蚂蚁与蜜蜂总科(包含spheciform wasps和bees)是姊妹群。系统发育基因组学方法也逐步被应用于研究被子植物各类群的亲缘关系, 如Wen等(2013)测定了葡萄科15个物种的转录组序列, 联合229个单拷贝的直系同源基因序列, 确定了葡萄科5个分支间的亲缘关系。
目前, 系统发育基因组学的策略主要是超级矩阵(supermatrix)法和超级树(supertree)法。超级矩阵法是将所有比对后的单基因序列拼接(concatenate)成一个完整大矩阵后建树, 在叶绿体基因中已经被成功应用(Jansen et al., 2007; Moore et al., 2007, 2010), 但在核基因中的应用尚不多。华大基因研究院(BGI)正在开展测定1,000种植物转录组的计划(1KP Project)(http://www.onekp.com/), 被子植物系统发育研究中一直难以解决的重要问题都有望利用核基因系统发育基因组学得到解决, 例如被子植物5个主要分支之间的关系, 真双子叶植物内部位置不确定类群的关系等。如前所述, 超级矩阵法无法避免系统误差, 很可能会得到统计上支持率很高的拓扑结构, 实际却不能反映物种之间真实的亲缘关系, 所以也需要谨慎选择基因。
超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等。Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009)。Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者。
Potential phylogenetic utility of the low-copy nuclear gene pistillata in dicotyledonous plants: comparison to nrDNA ITS and trnL intron in Sphaerocardamum and other Brassicaceae.
A phylogeny of the monocots, as inferred from rbcL and atpA sequence variation, and a comparison of methods for calculating jackknife and bootstrap values.
DoyleJA, HottonCL (1991) Diversification of early angiosperm pollen in a cladistic context. In: Pollen and Spores: Pattern of Diversification (eds Blackmore S, Barnes SH), pp. 169-195. Clarendon Press, Oxford , England.
Identification of shared single copy nuclear genes in Arabidopsis, Populus, Vitis and Oryza and their phylogenetic utility across various taxonomic levels.
BMC Evolutionary Biology, 10, 61.
[42]
EatonDA, ReeRH (2013)
Inferring phylogeny and introgression using RADseq Data: an example from flowering plants (Pedicularis: Orobanchaceae).
GIGANTEA: a circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains.
Complete plastid genome sequences of three rosids (Castanea, Prunus, Theobroma): evidence for at least two independent transfers of rpl22 to the nucleus.
Molecular cloning and characterization of GPA1, a G protein alpha subunit gene from Arabidopsis thaliana. Proceedings of the National Academy of Sciences,
Phylogenetic analysis of the plastid inverted repeat for 244 species: insights into deeper-level angiosperm relationships from a long, slowly evolving sequence region.
International Journal of Plant Sciences, 172, 541-558.
Reconciling gene and genome duplication events: using multiple nuclear gene families to infer the phylogeny of the aquatic plant family Pontederiaceae.
Complete plastid genome sequencing of trochodendraceae reveals a significant expansion of the inverted repeat and suggests a paleogene divergence between the two extant species.
PLoS ONE, 8, e60429.
[155]
SwamyB (1953)
The morphology and relationships of the Chloranthaceae.
Combining bioinformatics and phylogenetics to identify large sets of single-copy orthologous genes (COSII) for comparative, evolutionary and systematic studies: a test case in the euasterid plant clade.
Genetics, 174, 1407-1420.
[173]
WuZQ, GeS (2012)
The phylogeny of the BEP clade in grasses revisited: evidence from the whole-genome sequences of chloroplasts.
Molecular Phylogenetics and Evolution, 62, 573-578.
An empirical demonstration of using pentatricopeptide repeat (PPR) genes as plant phylogenetic tools: phylogeny of Verbenaceae and the Verbena complex.
Phylogeny of the New World diploid cottons (Gossypium L., Malvaceae) based on sequences of three low-copy nuclear genes.
2005
Selecting single-copy nuclear genes for plant phylogenetics: a preliminary analysis for the Senecioneae (Asteraceae).
2008
2
2007
... 第三, 相比于高等分类阶元, 被子植物低等分类阶元如目、科和属水平未解决的问题则更多.例如菊科(Asteraceae或Compositae)是真双子叶植物中最大的科, 约1,600属, 占开花植物属的10%(Funk et al., 2005; Anderberg et al., 2007).按照头状花序中小花的构造以及植物有无乳汁等特征, 菊科分为舌状花亚科(Liguliflorae)和管状花亚科(Tubuliforae), 但也有研究将菊科分为12个亚科, 其中前4个较大的亚科为Asteroideae(有16,000多种, 如青蒿、向日葵), Cichoroideae(3,200种, 如莴苣), Carduoideae (2,500种, 如大蓟)和Mutisioideae(750种)(Panero & Funk, 2008).另外还有其他一些属的位置无法确定(Anderberg et al., 2007), 各亚科之间以及亚科内也有很多关系尚未确定.十字花科(Brassicaceae)中有44个族(tribe)307属约3,600种, 被分为3个主要的系(lineage), 但是还有约38个属和71个种无法确定属于哪个系或哪个族; 3个系间的亲缘关系及系内部各族间和各属间的问题也仍然很多(Bailey et al., 2006; Couvreur et al., 2010; Franzke et al., 2011). ...
... ).另外还有其他一些属的位置无法确定(Anderberg et al., 2007), 各亚科之间以及亚科内也有很多关系尚未确定.十字花科(Brassicaceae)中有44个族(tribe)307属约3,600种, 被分为3个主要的系(lineage), 但是还有约38个属和71个种无法确定属于哪个系或哪个族; 3个系间的亲缘关系及系内部各族间和各属间的问题也仍然很多(Bailey et al., 2006; Couvreur et al., 2010; Franzke et al., 2011). ...
Phylogeny and divergence of basal angiosperms inferred from APETALA-3 and PISTILLATA-like MADS-box genes.
1
2004
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
An ordinal classification for the families of flowering plants.
1
1998
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG II.
1
2003
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
Potential phylogenetic utility of the low-copy nuclear gene pistillata in dicotyledonous plants: comparison to nrDNA ITS and trnL intron in Sphaerocardamum and other Brassicaceae.
1
1999
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Toward a global phylogeny of the Brassicaceae.
1
2006
... 第三, 相比于高等分类阶元, 被子植物低等分类阶元如目、科和属水平未解决的问题则更多.例如菊科(Asteraceae或Compositae)是真双子叶植物中最大的科, 约1,600属, 占开花植物属的10%(Funk et al., 2005; Anderberg et al., 2007).按照头状花序中小花的构造以及植物有无乳汁等特征, 菊科分为舌状花亚科(Liguliflorae)和管状花亚科(Tubuliforae), 但也有研究将菊科分为12个亚科, 其中前4个较大的亚科为Asteroideae(有16,000多种, 如青蒿、向日葵), Cichoroideae(3,200种, 如莴苣), Carduoideae (2,500种, 如大蓟)和Mutisioideae(750种)(Panero & Funk, 2008).另外还有其他一些属的位置无法确定(Anderberg et al., 2007), 各亚科之间以及亚科内也有很多关系尚未确定.十字花科(Brassicaceae)中有44个族(tribe)307属约3,600种, 被分为3个主要的系(lineage), 但是还有约38个属和71个种无法确定属于哪个系或哪个族; 3个系间的亲缘关系及系内部各族间和各属间的问题也仍然很多(Bailey et al., 2006; Couvreur et al., 2010; Franzke et al., 2011). ...
The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny.
Early Cretaceous lineages of monocot flowering plants.
1
2000
... 单子叶植物是五大分支中的第二大类群, 约占整个被子植物的22%, 为人类提供了主要的粮食作物(谷物与其他淀粉类作物等)(Bremer, 2000; Stevens, 2001).该类植物绝大多数为草本, 种子仅有1枚子叶, 平行叶脉, 花基数为3.单子叶植物包含11个目和1个科(Bremer et al., 2009), 其中兰科植物约占单子叶物种的34%, 禾本科植物约占17%.几乎所有的分子证据都表明菖蒲目(Acorales)是单子叶植物的最基部分支, 泽泻目(Alismatales)紧随 其后; 另外, 单子叶植物还包含天门冬目(Aspara-gales)、薯蓣目(Dioscoreales)、百合目(Liliales)、露兜树目(Pandanales)、无叶莲目(Petrosaviales)和鸭跖草类植物(commelinids).其中鸭跖草类植物是单子叶植物的核心类群, 包括棕榈目(Arecales; 木本)、鸭跖草目(Commelinales)、禾本目(Poales)、姜目(Zingiberales)和多须草科(Dasypogonaceae) (Davis et al., 2004; Graham et al., 2006)(图2c).单子叶植物是被子植物各大类群中系统发育研究得比较清楚的类群之一, 除了兰科和禾本科外, 几乎其他单子叶植物所有属都曾进行了系统发育研究, 大部分研究基于叶绿体rbcL基因, 这为研究单子叶植物各分支的起源、扩张、重要形态性状的演化与适应等奠定了基础(Soltis et al., 2005). ...
Inferring phylogenies with incomplete data sets: a 5-gene, 567-taxon analysis of angiosperms.
3
2009
... 叶绿体是植物特有的细胞器, 其环状基因组DNA被两个反向重复序列(inverted repeat sequence, IR)分为大单拷贝区(large single copy, LSC)和小单拷贝区(small single copy, SSC).虽然不同物种中叶绿体DNA(cpDNA)拷贝数目有差异, 但基因组成和排列相似, 基因数目几乎相同, 不同物种中的叶绿体基因是直系同源基因(ortholog), 因此利用叶绿体基因构建系统发育树时很少像使用核基因那样受旁系同源基因(paralog)的干扰; 同时, 叶绿体基因序列相对比较保守, 便于扩增和克隆, 因此在被子植物系统发育研究中得到广泛应用(Olmstead & Palmer, 1994).例如Chase等(1993)使用叶绿体基因rbcL分析了代表种子植物各类群的499个物种, 全面探讨了被子植物主要类群的系统发育关系, 是植物系统发育研究的典范.在叶绿体基因中, rbcL, atpB, ndhF和matK是比较常用的被子植物系统发育标记基因(Kim & Jansen, 1995; Soltis et al., 2000; Hilu et al., 2003; Qiu et al., 2005; Burleigh et al., 2009), 其中atpB和rbcL长度接近(氨基酸编码区序列长度大约为1,500 bp), 其核苷酸替换速率(substi- tution site/unit time)约为0.068-0.108, 序列易于比对(alignment); ndhF的序列更长(> 2,000 bp), 进化速率也相对较快; matK进化速率在叶绿体基因中最快(≥1,500 bp, ~0.141), 更适用于研究相对近缘物种之间的系统关系, 但因其插入/缺失(indel)比较多, 较其他叶绿体基因更难比对.线粒体基因如mtSSU, mtLSU, atpA, cox1, matR, atp1, nad5和rps3等也被经常使用(Qiu et al., 1999; Meng et al., 2002; Zanis et al., 2002), 其中atp1, rps3, nad5和matR等的进化速率比较慢, 核酸替换速率约为0.029-0.044, 对于研究远缘物种之间的亲缘关系很有帮助(Qiu et al., 2010). ...
... 目前被子植物系统发育研究主要分为两大类型: 第一类是物种数目很多但基因数目较少(Burleigh et al., 2009; Qiu et al., 2010; Soltis et al., 2011).当选取的基因序列较短或包含的系统发育信息少时, 并不能解决所有关注的问题, 或者很可能产生随机误差.降低随机误差的方法是选择系统发育信息充足的直系同源基因, 加大基因数据量(Wortley et al., 2005; Jian et al., 2008), 但是当物种数目太多时, 每增加一个基因片段信息都会增加工作量和计算时间.第二类研究是物种数量较少但基因数目很多, 即系统发育基因组学(Delsuc et al., 2005).被子植物的叶绿体系统发育基因组学在近几年取得了很大的进展, 例如Jansen等(2007)用81个叶绿体基因构建包含64个物种的系统树, Moore等(2007)用61个叶绿体基因(约42,000 bp)构建包含45个物种的系统树. ...
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Towards a phylogenetic nomenclature of Tracheophyta.
1
2007
... 除了ANITA基部类群外, 约占被子植物99.95%的其他类群聚为单系, 此类群包括数量不等的5个分支, 即真双子叶植物、单子叶植物(monocots)、木兰类(magnoliids)、金鱼藻目(Ceratophyllales)和金粟兰目(Chloranthales)(Cantino et al., 2007).真双子叶植物为五个分支中物种多样性最丰富的类群, 包含了约40个目300余科, 约占整个被子植物的75%, 该类群的共同形态特征为花粉粒具有三孔沟(tricolpate pollen), 因此也被称为“三孔花粉组”(Doyle & Hotton, 1991).真双子叶植物起源古老, 在中国发现的早白垩纪化石中就存在该类群植物(Drinnan et al., 1991); Sun et al., 2011), 该类群的主要物种在约八千多万年前的晚白垩纪桑托期(late Santonian)就已经形成(Herendeen et al., 1999). ...
Specialized structures in the leaf epidermis of basal angiosperms: morphology, distribution, and homology.
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Impact of missing data, gene choice, and taxon sampling on phylogenetic reconstruction: the Caryophyllales (angiosperms).
1
2012
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
The Evolution and Classification of Flowering Plants. Houghton Mifflin
Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences.
2
2002
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
Development of phylogenetic markers from single-copy nuclear genes for multi locus, species level analyses in the mint family (Lamiaceae).
1
2012
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
1
1903
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Darwin's abominable mystery: insights from a supertree of the angiosperms.
2
2004
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
Host-to-parasite gene transfer in flowering plants: phylogenetic evidence from Malpighiales.
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
A phylogeny of the monocots, as inferred from rbcL and atpA sequence variation, and a comparison of methods for calculating jackknife and bootstrap values.
1
2004
... 单子叶植物是五大分支中的第二大类群, 约占整个被子植物的22%, 为人类提供了主要的粮食作物(谷物与其他淀粉类作物等)(Bremer, 2000; Stevens, 2001).该类植物绝大多数为草本, 种子仅有1枚子叶, 平行叶脉, 花基数为3.单子叶植物包含11个目和1个科(Bremer et al., 2009), 其中兰科植物约占单子叶物种的34%, 禾本科植物约占17%.几乎所有的分子证据都表明菖蒲目(Acorales)是单子叶植物的最基部分支, 泽泻目(Alismatales)紧随 其后; 另外, 单子叶植物还包含天门冬目(Aspara-gales)、薯蓣目(Dioscoreales)、百合目(Liliales)、露兜树目(Pandanales)、无叶莲目(Petrosaviales)和鸭跖草类植物(commelinids).其中鸭跖草类植物是单子叶植物的核心类群, 包括棕榈目(Arecales; 木本)、鸭跖草目(Commelinales)、禾本目(Poales)、姜目(Zingiberales)和多须草科(Dasypogonaceae) (Davis et al., 2004; Graham et al., 2006)(图2c).单子叶植物是被子植物各大类群中系统发育研究得比较清楚的类群之一, 除了兰科和禾本科外, 几乎其他单子叶植物所有属都曾进行了系统发育研究, 大部分研究基于叶绿体rbcL基因, 这为研究单子叶植物各分支的起源、扩张、重要形态性状的演化与适应等奠定了基础(Soltis et al., 2005). ...
Chloroplast DNA deletions associated with wheat plants regenerated from pollen: possible basis for maternal inheritance of chloroplasts.
1
1984
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
Phylogenomics and the reconstruction of the tree of life.
2
2005
... 然而, 基于不同单基因构建的被子植物系统发育树之间往往存在分歧, 而且很多分支的支持率并不高, 这主要是由于单基因的序列较短和信息位点太少, 造成了随机误差(stochastic error)(Rokas et al., 2003; Delsuc et al., 2005; Jeffroy et al., 2006).例如Savolainen等(2000)使用atpB序列构建被子植物系统发育树时认为无油樟(Amborella trichocarpa)和睡莲目(Nymphaeales)是被子植物最基部的类群, 而Chase等(1993)基于rbcL序列的结果则显示金鱼藻(Ceratophyllum)位于被子植物最基部.随着数据的积累, 植物系统学家们联合几个单基因的序列来重建被子植物系统发育关系, 发现联合基因的系统树分支间的关系更明确, 各分支的支持率也得到了显著提高(Qiu et al., 1999; Soltis et al., 1999, 2002; Fishbein et al., 2001). ...
... 目前被子植物系统发育研究主要分为两大类型: 第一类是物种数目很多但基因数目较少(Burleigh et al., 2009; Qiu et al., 2010; Soltis et al., 2011).当选取的基因序列较短或包含的系统发育信息少时, 并不能解决所有关注的问题, 或者很可能产生随机误差.降低随机误差的方法是选择系统发育信息充足的直系同源基因, 加大基因数据量(Wortley et al., 2005; Jian et al., 2008), 但是当物种数目太多时, 每增加一个基因片段信息都会增加工作量和计算时间.第二类研究是物种数量较少但基因数目很多, 即系统发育基因组学(Delsuc et al., 2005).被子植物的叶绿体系统发育基因组学在近几年取得了很大的进展, 例如Jansen等(2007)用81个叶绿体基因构建包含64个物种的系统树, Moore等(2007)用61个叶绿体基因(约42,000 bp)构建包含45个物种的系统树. ...
Usefulness of RNA polymerase II coding sequences for estimation of green plant phylogeny.
1
1998
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
The occurrence of fruits with affinities to Ceratophyllaceae in lower and mid-Cretaceous sediments.
1
1989
... 有些来自化石方面的资料却似乎仍表明金鱼藻目可能是比较原始的类群.例如在早期化石中发现的角状果(horned fruit)与金鱼藻的果实很像(Dilcher, 1989); 另外被子植物最早的化石辽宁古果(Archaefructus liaoningensis)的花很简单, 这与现存的基部被子植物睡莲目的独蕊草科(Hydatellaeae)、金粟兰和金鱼藻的花很类似, 因此有研究者推测可能金鱼藻目与金粟兰目的关系比较近, 且位置都偏基部(Eklund et al., 2004; Friis et al., 2007; Endress & Doyle, 2009).但是, Moore等(2007)基于叶绿体基因组的结果以及APG III, 认为金鱼藻目是真双子叶植物的姊妹群, 然后是单子叶植物, 而木兰类与金粟兰目形成的姊妹群是五大分支最基部的类群. ...
1
1991
... 除了ANITA基部类群外, 约占被子植物99.95%的其他类群聚为单系, 此类群包括数量不等的5个分支, 即真双子叶植物、单子叶植物(monocots)、木兰类(magnoliids)、金鱼藻目(Ceratophyllales)和金粟兰目(Chloranthales)(Cantino et al., 2007).真双子叶植物为五个分支中物种多样性最丰富的类群, 包含了约40个目300余科, 约占整个被子植物的75%, 该类群的共同形态特征为花粉粒具有三孔沟(tricolpate pollen), 因此也被称为“三孔花粉组”(Doyle & Hotton, 1991).真双子叶植物起源古老, 在中国发现的早白垩纪化石中就存在该类群植物(Drinnan et al., 1991); Sun et al., 2011), 该类群的主要物种在约八千多万年前的晚白垩纪桑托期(late Santonian)就已经形成(Herendeen et al., 1999). ...
Angiosperm flowers and tricolpate pollen of buxaceous affinity from the Potomac Group (mid-Cretaceous) of eastern North America.
2
1991
... 除了ANITA基部类群外, 约占被子植物99.95%的其他类群聚为单系, 此类群包括数量不等的5个分支, 即真双子叶植物、单子叶植物(monocots)、木兰类(magnoliids)、金鱼藻目(Ceratophyllales)和金粟兰目(Chloranthales)(Cantino et al., 2007).真双子叶植物为五个分支中物种多样性最丰富的类群, 包含了约40个目300余科, 约占整个被子植物的75%, 该类群的共同形态特征为花粉粒具有三孔沟(tricolpate pollen), 因此也被称为“三孔花粉组”(Doyle & Hotton, 1991).真双子叶植物起源古老, 在中国发现的早白垩纪化石中就存在该类群植物(Drinnan et al., 1991); Sun et al., 2011), 该类群的主要物种在约八千多万年前的晚白垩纪桑托期(late Santonian)就已经形成(Herendeen et al., 1999). ...
Identification of shared single copy nuclear genes in Arabidopsis, Populus, Vitis and Oryza and their phylogenetic utility across various taxonomic levels.
2010
Inferring phylogeny and introgression using RADseq Data: an example from flowering plants (Pedicularis: Orobanchaceae).
... 有些来自化石方面的资料却似乎仍表明金鱼藻目可能是比较原始的类群.例如在早期化石中发现的角状果(horned fruit)与金鱼藻的果实很像(Dilcher, 1989); 另外被子植物最早的化石辽宁古果(Archaefructus liaoningensis)的花很简单, 这与现存的基部被子植物睡莲目的独蕊草科(Hydatellaeae)、金粟兰和金鱼藻的花很类似, 因此有研究者推测可能金鱼藻目与金粟兰目的关系比较近, 且位置都偏基部(Eklund et al., 2004; Friis et al., 2007; Endress & Doyle, 2009).但是, Moore等(2007)基于叶绿体基因组的结果以及APG III, 认为金鱼藻目是真双子叶植物的姊妹群, 然后是单子叶植物, 而木兰类与金粟兰目形成的姊妹群是五大分支最基部的类群. ...
The granule-bound starch synthase (GBSSI) gene in the Rosaceae: multiple loci and phylogenetic utility.
1
2000
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
“Inordinate fondness” explained: why are there so many beetles?
1
1998
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Maternal inheritance of chloroplast genome and paternal inheritance of mitochondrial genome in bananas (Musa acuminata).
1
1994
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
Multigene phylogeny of the green lineage reveals the origin and diversification of land plants.
4
2010
... 在蔷薇类植物中, 主要类群为固氮分支(nitrogen-fixing clade), 包括葫芦目(Cucurbitales)、壳斗目(Fagales)、蔷薇目(Rosales)和豆目(Fabales), 是支持率很高的单系群; 另外两个高支持率的分支是COM分支和锦葵类(malvids)分支, 前者包括卫矛目(Celastrales)、酢浆草目(Oxalidales)和金虎尾目(Malpighiales), 后者包括锦葵目(Malvales)、包含模式植物拟南芥的十字花目(Brassicales)、燧体木目(Crossosomatales)、无患子目(Sapindales)、十齿花目(Huerteales)和美洲苦木目(Picramniales)(Bremer et al., 2009; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012)(图2a).在菊类植物中, 大部分物种组成一个被称为真菊的单系, 而山茱萸目(Cornales)和杜鹃花目(Ericales)则是真菊的姊妹群(Bremer et al., 2009; Zhang et al., 2012)(图2b).APG III又将真菊植物分为真菊I类(也被称为唇形类lamiids)和真菊II类(也被称为桔梗类campanulids); 其中真菊I类的核心类群是唇形目(Lamiales)、茄目(Solanales)、龙胆目(Gentianles)、紫草科(Boraginaceae), 而真菊II类的核心类群是盔瓣花目(Paracryphiales)、川续断目(Dipsacales)、伞形目(Apiales)、鳞叶树目(Bruniales)、菊目(Asterales)和南鼠刺目(Escalloniales). ...
... 其次, 除了被子植物5个主要分支的亲缘关系还不能确定外, 各大类群内部主要分支的关系也没有完全厘清(图2).真双子叶植物中, 除了最基部的毛茛目外, 其他基部类群中的清风藤科与山龙眼目、黄杨目与昆栏树目是否互为姊妹群, 尚需进一步的研究(Moore et al., 2010; Zhang et al., 2011).核心真双子叶植物中除了蔷薇类和菊类植物两大分支外, 其他重要分支, 即虎耳草目、檀香目、葡萄目、五桠果科和石竹目的系统发育位置也还没有最终确定(图1).蔷薇类内部的很多关系也有争论(Finet et al., 2010; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... ; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Phylogeny of Saxifragales (angiosperms, eudicots): analysis of a rapid, ancient radiation.
3
2001
... 然而, 基于不同单基因构建的被子植物系统发育树之间往往存在分歧, 而且很多分支的支持率并不高, 这主要是由于单基因的序列较短和信息位点太少, 造成了随机误差(stochastic error)(Rokas et al., 2003; Delsuc et al., 2005; Jeffroy et al., 2006).例如Savolainen等(2000)使用atpB序列构建被子植物系统发育树时认为无油樟(Amborella trichocarpa)和睡莲目(Nymphaeales)是被子植物最基部的类群, 而Chase等(1993)基于rbcL序列的结果则显示金鱼藻(Ceratophyllum)位于被子植物最基部.随着数据的积累, 植物系统学家们联合几个单基因的序列来重建被子植物系统发育关系, 发现联合基因的系统树分支间的关系更明确, 各分支的支持率也得到了显著提高(Qiu et al., 1999; Soltis et al., 1999, 2002; Fishbein et al., 2001). ...
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
Evolutionary dynamics of Waxy and the origin of hexaploid Spartina species (Poaceae).
1
2007
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
GIGANTEA: a circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains.
1
1999
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Cabbage family affairs: the evolutionary history of Brassicaceae.
1
2011
... 第三, 相比于高等分类阶元, 被子植物低等分类阶元如目、科和属水平未解决的问题则更多.例如菊科(Asteraceae或Compositae)是真双子叶植物中最大的科, 约1,600属, 占开花植物属的10%(Funk et al., 2005; Anderberg et al., 2007).按照头状花序中小花的构造以及植物有无乳汁等特征, 菊科分为舌状花亚科(Liguliflorae)和管状花亚科(Tubuliforae), 但也有研究将菊科分为12个亚科, 其中前4个较大的亚科为Asteroideae(有16,000多种, 如青蒿、向日葵), Cichoroideae(3,200种, 如莴苣), Carduoideae (2,500种, 如大蓟)和Mutisioideae(750种)(Panero & Funk, 2008).另外还有其他一些属的位置无法确定(Anderberg et al., 2007), 各亚科之间以及亚科内也有很多关系尚未确定.十字花科(Brassicaceae)中有44个族(tribe)307属约3,600种, 被分为3个主要的系(lineage), 但是还有约38个属和71个种无法确定属于哪个系或哪个族; 3个系间的亲缘关系及系内部各族间和各属间的问题也仍然很多(Bailey et al., 2006; Couvreur et al., 2010; Franzke et al., 2011). ...
The meaning of Darwin’s “abominable mystery”.
1
2009
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Floral evidence for Cretaceous chloranthoid angiosperms.
... 从Hamby和Zimmer(1992)首次用18S和26S rDNA序列片段构建被子植物系统发育树, 到1998年被子植物系统发育组基于rbcL, atpB和18S rDNA的序列首次提出APG分类标准并不断修改更新至APG III(Bremer et al., 2009), 以及最近运用编码蛋白的核基因序列研究各分类阶元被子植物的系统发育关系, 人们对被子植物各类群间的进化关系有了全新的认识(Hamby & Zimmer, 1992; Jansen et al., 2007; Moore et al., 2007; Bremer et al., 2009; Zhang et al., 2012)(图1). ...
A preliminary conspectus of the Allon flora from the late Cretaceous (late Santonian) of central Georgia, USA.
1
1999
... 除了ANITA基部类群外, 约占被子植物99.95%的其他类群聚为单系, 此类群包括数量不等的5个分支, 即真双子叶植物、单子叶植物(monocots)、木兰类(magnoliids)、金鱼藻目(Ceratophyllales)和金粟兰目(Chloranthales)(Cantino et al., 2007).真双子叶植物为五个分支中物种多样性最丰富的类群, 包含了约40个目300余科, 约占整个被子植物的75%, 该类群的共同形态特征为花粉粒具有三孔沟(tricolpate pollen), 因此也被称为“三孔花粉组”(Doyle & Hotton, 1991).真双子叶植物起源古老, 在中国发现的早白垩纪化石中就存在该类群植物(Drinnan et al., 1991); Sun et al., 2011), 该类群的主要物种在约八千多万年前的晚白垩纪桑托期(late Santonian)就已经形成(Herendeen et al., 1999). ...
Is sparse taxon sampling a problem for phylogenetic inference?
1
2003
... 系统发育分析中随着数据量的增加虽然能尽可能降低随机误差带来的影响, 但是却可能引入系统误差; 这些误差在某些情况下还可能被放大, 从而导致产生统计学上支持率很高但却不能反映真实物种关系的进化树(Phillips et al., 2004; Jeffroy et al., 2006).因此, 相对于使用较少基因序列构建系统发育树, 用系统发育基因组学来研究物种间的亲缘关系时更需要谨慎(Soltis et al., 2004).被子植物系统发育基因组学研究中比较典型的反例即由系统误差带来的长枝吸引假象(Philippe et al., 2011), 其典型原因就是取样不当或取样量不够.被子植物基部类群的确定就出现过类似的问题.虽然很多研究表明Amborella或Amborella和Nymphaeales形成的姊妹群是被子植物的最基部类群, 但是Goremykin等(2003)测定Amborella trichopoda和Nymphaea alba的叶绿体全基因组序列后, 结合公共数据库中12个叶绿体基因组序列, 提出单子叶植物(禾本科为代表物种)为被子植物的最基部类群.然而Soltis和Stefanovic等发现在增加关键类群, 尤其是基部类群如Nymphaea、Austrobaileya、木兰类或基部单子叶植物后, Amborella或Amborella + Nymphaeales仍然是被子植物的最基部类群(Soltis & Soltis, 2004; Stefanović et al., 2004), 这一结果在增加取样量后的叶绿体系统发育基因组学研究结果中也得到了验证(Jansen et al., 2007; Moore et al., 2007).Goremykin等(2003, 2013)的结果很大程度上是由于取样不当或不够导致的, 禾本科的进化速率很快, 因此当仅以长枝的禾本科植物作为单子叶植物的代表, 以及选用买麻藤属(Gnetum)植物作为外类群时, 就会造成单子叶被吸引到基部的假象.除了选取合适的进化模型和分析方法之外, 密集采样以减少长枝, 目前被认为是降低长枝吸引影响的有效方法(Graybeal, 1998; Hillis et al., 2003).但是目前大部分实验室还是较难对物种密集测序, 而且当数据量增多时, 系统发育“噪音”也会增多, 从而可能放大系统误差.无论使用叶绿体基因组还是核基因组, 系统误差的影响都不容忽视. ...
The matK gene: sequence variation and application in plant systematics.
1
1997
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
Angiosperm phylogeny based on matK sequence information.
1
2003
... 叶绿体是植物特有的细胞器, 其环状基因组DNA被两个反向重复序列(inverted repeat sequence, IR)分为大单拷贝区(large single copy, LSC)和小单拷贝区(small single copy, SSC).虽然不同物种中叶绿体DNA(cpDNA)拷贝数目有差异, 但基因组成和排列相似, 基因数目几乎相同, 不同物种中的叶绿体基因是直系同源基因(ortholog), 因此利用叶绿体基因构建系统发育树时很少像使用核基因那样受旁系同源基因(paralog)的干扰; 同时, 叶绿体基因序列相对比较保守, 便于扩增和克隆, 因此在被子植物系统发育研究中得到广泛应用(Olmstead & Palmer, 1994).例如Chase等(1993)使用叶绿体基因rbcL分析了代表种子植物各类群的499个物种, 全面探讨了被子植物主要类群的系统发育关系, 是植物系统发育研究的典范.在叶绿体基因中, rbcL, atpB, ndhF和matK是比较常用的被子植物系统发育标记基因(Kim & Jansen, 1995; Soltis et al., 2000; Hilu et al., 2003; Qiu et al., 2005; Burleigh et al., 2009), 其中atpB和rbcL长度接近(氨基酸编码区序列长度大约为1,500 bp), 其核苷酸替换速率(substi- tution site/unit time)约为0.068-0.108, 序列易于比对(alignment); ndhF的序列更长(> 2,000 bp), 进化速率也相对较快; matK进化速率在叶绿体基因中最快(≥1,500 bp, ~0.141), 更适用于研究相对近缘物种之间的系统关系, 但因其插入/缺失(indel)比较多, 较其他叶绿体基因更难比对.线粒体基因如mtSSU, mtLSU, atpA, cox1, matR, atp1, nad5和rps3等也被经常使用(Qiu et al., 1999; Meng et al., 2002; Zanis et al., 2002), 其中atp1, rps3, nad5和matR等的进化速率比较慢, 核酸替换速率约为0.029-0.044, 对于研究远缘物种之间的亲缘关系很有帮助(Qiu et al., 2010). ...
Phylogenetic analysis of the acetyl-CoA carboxylase and 3-phosphoglycerate kinase loci in wheat and other grasses.
2
2002
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
... (plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns.
5
2007
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
... 从Hamby和Zimmer(1992)首次用18S和26S rDNA序列片段构建被子植物系统发育树, 到1998年被子植物系统发育组基于rbcL, atpB和18S rDNA的序列首次提出APG分类标准并不断修改更新至APG III(Bremer et al., 2009), 以及最近运用编码蛋白的核基因序列研究各分类阶元被子植物的系统发育关系, 人们对被子植物各类群间的进化关系有了全新的认识(Hamby & Zimmer, 1992; Jansen et al., 2007; Moore et al., 2007; Bremer et al., 2009; Zhang et al., 2012)(图1). ...
... 关于这5个类群的亲缘关系, 不同研究者根据不同基因或不同数目的物种得到的结果不尽相同, 目前已发表的拓扑结构约有15种, 但支持率都不高(Soltis et al., 1999; Jansen et al., 2007; Moore et al., 2011; Zhang et al., 2012).其中, 有研究分别将单子叶植物、单子叶植物与金鱼藻、金鱼藻和金粟兰、木兰类、金粟兰、木兰类与金粟兰作为这5个分支中的最基部类群.例如, Qiu等(1999)联合2个叶绿体基因、2个线粒体基因和18S rDNA的序列构建了包含105个种的系统树, 认为真双子叶植物与木兰类植物是姊妹群, 金粟兰目是两者的姊妹群, 而单子叶植物与金鱼藻目形成的姊妹群位于最基部.有一些源于核基因、线粒体和叶绿体基因的证据也曾认为金鱼藻目与单子叶植物是姊妹群(Soltis et al., 1997; Savolainen et al., 2000), 但金鱼藻目是比较特化的类群, 而单子叶植物多为草本, 进化速率快, 两者在进化树上都表现为长枝, 因此有研究者认为二者聚为姊妹群很可能是因为长枝吸引(long-branch attraction, LBA)(Soltis et al., 2005). ...
... 系统发育分析中随着数据量的增加虽然能尽可能降低随机误差带来的影响, 但是却可能引入系统误差; 这些误差在某些情况下还可能被放大, 从而导致产生统计学上支持率很高但却不能反映真实物种关系的进化树(Phillips et al., 2004; Jeffroy et al., 2006).因此, 相对于使用较少基因序列构建系统发育树, 用系统发育基因组学来研究物种间的亲缘关系时更需要谨慎(Soltis et al., 2004).被子植物系统发育基因组学研究中比较典型的反例即由系统误差带来的长枝吸引假象(Philippe et al., 2011), 其典型原因就是取样不当或取样量不够.被子植物基部类群的确定就出现过类似的问题.虽然很多研究表明Amborella或Amborella和Nymphaeales形成的姊妹群是被子植物的最基部类群, 但是Goremykin等(2003)测定Amborella trichopoda和Nymphaea alba的叶绿体全基因组序列后, 结合公共数据库中12个叶绿体基因组序列, 提出单子叶植物(禾本科为代表物种)为被子植物的最基部类群.然而Soltis和Stefanovic等发现在增加关键类群, 尤其是基部类群如Nymphaea、Austrobaileya、木兰类或基部单子叶植物后, Amborella或Amborella + Nymphaeales仍然是被子植物的最基部类群(Soltis & Soltis, 2004; Stefanović et al., 2004), 这一结果在增加取样量后的叶绿体系统发育基因组学研究结果中也得到了验证(Jansen et al., 2007; Moore et al., 2007).Goremykin等(2003, 2013)的结果很大程度上是由于取样不当或不够导致的, 禾本科的进化速率很快, 因此当仅以长枝的禾本科植物作为单子叶植物的代表, 以及选用买麻藤属(Gnetum)植物作为外类群时, 就会造成单子叶被吸引到基部的假象.除了选取合适的进化模型和分析方法之外, 密集采样以减少长枝, 目前被认为是降低长枝吸引影响的有效方法(Graybeal, 1998; Hillis et al., 2003).但是目前大部分实验室还是较难对物种密集测序, 而且当数据量增多时, 系统发育“噪音”也会增多, 从而可能放大系统误差.无论使用叶绿体基因组还是核基因组, 系统误差的影响都不容忽视. ...
... 目前, 系统发育基因组学的策略主要是超级矩阵(supermatrix)法和超级树(supertree)法.超级矩阵法是将所有比对后的单基因序列拼接(concatenate)成一个完整大矩阵后建树, 在叶绿体基因中已经被成功应用(Jansen et al., 2007; Moore et al., 2007, 2010), 但在核基因中的应用尚不多.华大基因研究院(BGI)正在开展测定1,000种植物转录组的计划(1KP Project)(http://www.onekp.com/), 被子植物系统发育研究中一直难以解决的重要问题都有望利用核基因系统发育基因组学得到解决, 例如被子植物5个主要分支之间的关系, 真双子叶植物内部位置不确定类群的关系等.如前所述, 超级矩阵法无法避免系统误差, 很可能会得到统计上支持率很高的拓扑结构, 实际却不能反映物种之间真实的亲缘关系, 所以也需要谨慎选择基因. ...
Complete plastid genome sequences of three rosids (Castanea, Prunus, Theobroma): evidence for at least two independent transfers of rpl22 to the nucleus.
1
2011
... 测序技术的成熟与计算能力的提高促使越来越多的植物叶绿体基因组被测序, 叶绿体系统发育基因组学(phylogenomics)也逐步成为被子植物系统发育研究的趋势之一(Moore et al., 2010; Jansen et al., 2011; Weng et al., 2012).例如, Jansen等(2007)利用64个物种的叶绿体基因组的81个基因重建了被子植物大类群之间的亲缘关系; Moore等(2010)根据86个物种的83个叶绿体基因重建了真双子叶(eudicots)植物内部大类群之间的亲缘关系.叶绿体基因组也开始被应用于被子植物科以下分类阶元的进化研究.例如Zhang等(2011)成功运用叶绿体基因组序列研究了竹亚科(Bambusoideae)在禾本科(Poaceae)中的系统位置, 提出了竹亚科与早熟禾亚科(Pooideae)为姊妹群的假说; Wu和Ge(2012)利用22个禾本科植物的叶绿体基因组成功解决了禾本科三大类群即竹亚科、早熟禾亚科和稻亚科(Ehrhartoidea)之间的亲缘关系; Xi等(2012)对约占热带雨林物种数40%的金虎尾目(Malpighiales)进行了系统发育重建, 并鉴定出了新的12个分支; Sun等(2013)推断基部真双子叶植物昆栏树科(Trochon- dendraceae)在30-44百万年前分化形成昆栏树属(Trochodendron)和水青树属(Tetracentron). ...
Phylogenomics: the beginning of incongruence?
4
2006
... 然而, 基于不同单基因构建的被子植物系统发育树之间往往存在分歧, 而且很多分支的支持率并不高, 这主要是由于单基因的序列较短和信息位点太少, 造成了随机误差(stochastic error)(Rokas et al., 2003; Delsuc et al., 2005; Jeffroy et al., 2006).例如Savolainen等(2000)使用atpB序列构建被子植物系统发育树时认为无油樟(Amborella trichocarpa)和睡莲目(Nymphaeales)是被子植物最基部的类群, 而Chase等(1993)基于rbcL序列的结果则显示金鱼藻(Ceratophyllum)位于被子植物最基部.随着数据的积累, 植物系统学家们联合几个单基因的序列来重建被子植物系统发育关系, 发现联合基因的系统树分支间的关系更明确, 各分支的支持率也得到了显著提高(Qiu et al., 1999; Soltis et al., 1999, 2002; Fishbein et al., 2001). ...
... 系统发育研究中, 可能导致结果不准确的原因主要分为如下三类(Jeffroy et al., 2006; Philippe et al., 2011): (1)使用的分子标记不是真正的直系同源基因, 即hidden paralogs.被子植物中全基因组加倍和其他基因重复事件多次发生, 基因重复后不同拷贝的丢失事件使得直系同源基因的鉴定变得复杂(Jiao et al., 2011).(2)用于系统发育分析的基因长度较短或者信息含量较低时, “噪音”信息例如回复突变(reverse mutation)和平行突变(parallel mutation, 也被称为homoplasy)等, 可能会掩盖真实的系统发育信息, 从而可能引入随机误差.(3)不同物种中核苷酸成分或碱基替代模式的偏差(compositional signal)、类群之间进化速率的差异(rate signal)以及特定碱基位点替换速率的差异(heterotachous signal)等因素造成的系统误差(systematic error)(Lopez et al., 2002; Collins et al., 2005).由于世代长短、居群大小和核苷酸替换速率差异等因素, 不同被子植物类群的进化速率是不同的, 当类群间进化速率差异很大时, 进化快的类群(在进化树上表现为长枝)可能因为多重突变在某些碱基位点随机获得了相同的碱基, 从而使亲缘关系较远的物种在序列的部分位置上反而相似, 当这些非同源相似过多而足以掩盖真实的同源信息时, 就会错误地将进化快的类群聚在一起(即长枝吸引)(Goremykin et al., 2003).另外, 虽然目前常用的序列替代模型已经考虑了基因内部不同碱基位点因选择压力不同而有相异的进化速率, 但是几乎所有的模型都没有考虑选择压力会随时间发生变化, 即假设特定碱基位点的替代速率随着时间推移在类群之间保持恒定, 这与实际的分子进化过程并不一定相符(Yang, 1994; Gu et al., 1995; Lopez et al., 2002). ...
... 系统发育分析中随着数据量的增加虽然能尽可能降低随机误差带来的影响, 但是却可能引入系统误差; 这些误差在某些情况下还可能被放大, 从而导致产生统计学上支持率很高但却不能反映真实物种关系的进化树(Phillips et al., 2004; Jeffroy et al., 2006).因此, 相对于使用较少基因序列构建系统发育树, 用系统发育基因组学来研究物种间的亲缘关系时更需要谨慎(Soltis et al., 2004).被子植物系统发育基因组学研究中比较典型的反例即由系统误差带来的长枝吸引假象(Philippe et al., 2011), 其典型原因就是取样不当或取样量不够.被子植物基部类群的确定就出现过类似的问题.虽然很多研究表明Amborella或Amborella和Nymphaeales形成的姊妹群是被子植物的最基部类群, 但是Goremykin等(2003)测定Amborella trichopoda和Nymphaea alba的叶绿体全基因组序列后, 结合公共数据库中12个叶绿体基因组序列, 提出单子叶植物(禾本科为代表物种)为被子植物的最基部类群.然而Soltis和Stefanovic等发现在增加关键类群, 尤其是基部类群如Nymphaea、Austrobaileya、木兰类或基部单子叶植物后, Amborella或Amborella + Nymphaeales仍然是被子植物的最基部类群(Soltis & Soltis, 2004; Stefanović et al., 2004), 这一结果在增加取样量后的叶绿体系统发育基因组学研究结果中也得到了验证(Jansen et al., 2007; Moore et al., 2007).Goremykin等(2003, 2013)的结果很大程度上是由于取样不当或不够导致的, 禾本科的进化速率很快, 因此当仅以长枝的禾本科植物作为单子叶植物的代表, 以及选用买麻藤属(Gnetum)植物作为外类群时, 就会造成单子叶被吸引到基部的假象.除了选取合适的进化模型和分析方法之外, 密集采样以减少长枝, 目前被认为是降低长枝吸引影响的有效方法(Graybeal, 1998; Hillis et al., 2003).但是目前大部分实验室还是较难对物种密集测序, 而且当数据量增多时, 系统发育“噪音”也会增多, 从而可能放大系统误差.无论使用叶绿体基因组还是核基因组, 系统误差的影响都不容忽视. ...
... 综上所述, 通过仔细筛选直系同源基因(Zhang et al., 2012), 选用适当的分子数据, 例如研究起源较早的类群之间的亲缘关系时使用氨基酸序列以减少密码子简并性带来的“噪音”, 或者使用系统发育信号强的基因降低干扰(Goremykin et al., 2009; Townsend et al., 2012; Salichos & Rokas, 2013), 使用核苷酸数据研究亲缘关系较远的类群时去除变化速率快的第三位密码子或位点(Jeffroy et al., 2006; Goremykin et al., 2010), 适当增加物种或选取进化较慢的物种(Graybeal, 1998), 开发新的符合基因序列进化过程的替换模型, 寻找其他序列特征例如插入缺失等策略来尽量降低各种误差的影响显得非常重要. ...
Resolving an ancient, rapid radiation in Saxifragales.
2
2008
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
... 目前被子植物系统发育研究主要分为两大类型: 第一类是物种数目很多但基因数目较少(Burleigh et al., 2009; Qiu et al., 2010; Soltis et al., 2011).当选取的基因序列较短或包含的系统发育信息少时, 并不能解决所有关注的问题, 或者很可能产生随机误差.降低随机误差的方法是选择系统发育信息充足的直系同源基因, 加大基因数据量(Wortley et al., 2005; Jian et al., 2008), 但是当物种数目太多时, 每增加一个基因片段信息都会增加工作量和计算时间.第二类研究是物种数量较少但基因数目很多, 即系统发育基因组学(Delsuc et al., 2005).被子植物的叶绿体系统发育基因组学在近几年取得了很大的进展, 例如Jansen等(2007)用81个叶绿体基因构建包含64个物种的系统树, Moore等(2007)用61个叶绿体基因(约42,000 bp)构建包含45个物种的系统树. ...
Ancestral polyploidy in seed plants and angiosperms.
2
2011
... 虽然核基因在被子植物系统发育研究中的重要性不容忽视, 但是编码蛋白质的核基因或核基因组的使用远远滞后于叶绿体基因的运用.一方面, 由于核基因组拷贝低, 导致使用传统的PCR扩增和测序得到序列信息相对困难, 有时不得不依赖于克隆测序, 工作量增大, 限制了核基因在被子植物系统发育研究中的应用.另一方面, 由于被子植物在进化过程中经历多次全基因组加倍(whole genome duplication)(Jiao et al., 2011), 近期的多倍化事件更加频繁, 据保守估计被子植物中约有50%的物种是由于多倍化形成的(Soltis et al., 2009); 其他机制导致的基因加倍事件也频繁发生(Ng & Yanofsky, 2001; Lin et al., 2006; Xu et al., 2009), 导致不同类群中存在丢失不同拷贝的现象(Xu et al., 2009).这些因素使得同源核基因是直系同源还是旁系同源的关系难以确定.尽管如此, 研究者们通过利用大量的测序资源与基因序列数据, 开始在不同亲缘关系的类群间寻找可能是直系同源关系的单拷贝基因, 并取得了一定进展.例如, Fulton等(2002)通过比较番茄(Solanum lycopersicum)的EST序列和拟南芥(Arabidopsis thaliana)的基因组序列, 鉴定了两者共有的1,025个保守的低拷贝候选直系同源基因; Wu等(2006)比较番茄、土豆(Solanum tuberosum)、辣椒(Capsicum annuum)和咖啡(Coffea arabica)的EST序列与拟南芥的基因组序列, 认为真菊类植物(euasterids)与拟南芥共有2,869个单拷贝候选直系同源基因; Duarte等(2010)鉴定了拟南芥、杨树(Pop- ulus trichocarpa)、葡萄(Vitis vinifera)和水稻(Oryza sativa)共有的959个单拷贝候选直系同源基因, 并发现其中大部分基因在小立碗藓(Physcomitrella patens)和卷柏(Selaginella moellendorffii)中也能找到; Zhang等(2012)通过比较拟南芥、杨树、桃(Prunus persica)、葡萄、猴面花(Mimulus guttatus)、水稻、高粱(Sorghum bicolor)和小立碗藓的序列鉴定了1,083个高度保守的低拷贝核基因.这些工作为低拷贝核基因的利用奠定了基础. ...
... 系统发育研究中, 可能导致结果不准确的原因主要分为如下三类(Jeffroy et al., 2006; Philippe et al., 2011): (1)使用的分子标记不是真正的直系同源基因, 即hidden paralogs.被子植物中全基因组加倍和其他基因重复事件多次发生, 基因重复后不同拷贝的丢失事件使得直系同源基因的鉴定变得复杂(Jiao et al., 2011).(2)用于系统发育分析的基因长度较短或者信息含量较低时, “噪音”信息例如回复突变(reverse mutation)和平行突变(parallel mutation, 也被称为homoplasy)等, 可能会掩盖真实的系统发育信息, 从而可能引入随机误差.(3)不同物种中核苷酸成分或碱基替代模式的偏差(compositional signal)、类群之间进化速率的差异(rate signal)以及特定碱基位点替换速率的差异(heterotachous signal)等因素造成的系统误差(systematic error)(Lopez et al., 2002; Collins et al., 2005).由于世代长短、居群大小和核苷酸替换速率差异等因素, 不同被子植物类群的进化速率是不同的, 当类群间进化速率差异很大时, 进化快的类群(在进化树上表现为长枝)可能因为多重突变在某些碱基位点随机获得了相同的碱基, 从而使亲缘关系较远的物种在序列的部分位置上反而相似, 当这些非同源相似过多而足以掩盖真实的同源信息时, 就会错误地将进化快的类群聚在一起(即长枝吸引)(Goremykin et al., 2003).另外, 虽然目前常用的序列替代模型已经考虑了基因内部不同碱基位点因选择压力不同而有相异的进化速率, 但是几乎所有的模型都没有考虑选择压力会随时间发生变化, 即假设特定碱基位点的替代速率随着时间推移在类群之间保持恒定, 这与实际的分子进化过程并不一定相符(Yang, 1994; Gu et al., 1995; Lopez et al., 2002). ...
Phylogenomics resolves evolutionary relationships among ants, bees, and wasps.
2013
A survey of tricolpate (eudicot) phylogenetic relationships.
2
2004
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
... 真双子叶植物中, 毛茛目(Ranunculales)位于最基部, 是其他所有真双子叶植物的姊妹群; 清风藤科(Sabiaceae)、山龙眼目(Proteales)、黄杨目(Buxales)和昆栏树目(Trochodendrales)是真双子叶植物的其他基部类群; 其余所有真双子叶物种形成单系, 被命名为核心真双子叶(core eudicots)植物(Magallón et al., 1999).在核心真双子叶植物中, 洋二仙草目(Gunnerales)是其他类群的姊妹群; 其他类群的花多是五基数, 因此也被称为Pentapetalae, 主要由蔷薇类(rosids)和菊类植物(asterids)两大类群组成, 另外还包含系统位置尚未完全确定的几个类群, 即虎耳草目(Saxifragales)、葡萄目(Vitales)、檀香目(Santalales)、五桠果科(Dilleniaceae)和石竹目(Caryophyllales)(Judd & Olmstead, 2004; Worberg et al., 2007; Moore et al., 2010). ...
ndhF sequence evolution and the major clades in the sunflower family.
1
1995
... 叶绿体是植物特有的细胞器, 其环状基因组DNA被两个反向重复序列(inverted repeat sequence, IR)分为大单拷贝区(large single copy, LSC)和小单拷贝区(small single copy, SSC).虽然不同物种中叶绿体DNA(cpDNA)拷贝数目有差异, 但基因组成和排列相似, 基因数目几乎相同, 不同物种中的叶绿体基因是直系同源基因(ortholog), 因此利用叶绿体基因构建系统发育树时很少像使用核基因那样受旁系同源基因(paralog)的干扰; 同时, 叶绿体基因序列相对比较保守, 便于扩增和克隆, 因此在被子植物系统发育研究中得到广泛应用(Olmstead & Palmer, 1994).例如Chase等(1993)使用叶绿体基因rbcL分析了代表种子植物各类群的499个物种, 全面探讨了被子植物主要类群的系统发育关系, 是植物系统发育研究的典范.在叶绿体基因中, rbcL, atpB, ndhF和matK是比较常用的被子植物系统发育标记基因(Kim & Jansen, 1995; Soltis et al., 2000; Hilu et al., 2003; Qiu et al., 2005; Burleigh et al., 2009), 其中atpB和rbcL长度接近(氨基酸编码区序列长度大约为1,500 bp), 其核苷酸替换速率(substi- tution site/unit time)约为0.068-0.108, 序列易于比对(alignment); ndhF的序列更长(> 2,000 bp), 进化速率也相对较快; matK进化速率在叶绿体基因中最快(≥1,500 bp, ~0.141), 更适用于研究相对近缘物种之间的系统关系, 但因其插入/缺失(indel)比较多, 较其他叶绿体基因更难比对.线粒体基因如mtSSU, mtLSU, atpA, cox1, matR, atp1, nad5和rps3等也被经常使用(Qiu et al., 1999; Meng et al., 2002; Zanis et al., 2002), 其中atp1, rps3, nad5和matR等的进化速率比较慢, 核酸替换速率约为0.029-0.044, 对于研究远缘物种之间的亲缘关系很有帮助(Qiu et al., 2010). ...
Allopolyploid speciation in Persicaria (Polygonaceae): insights from a low-copy nuclear region.
2008
On the genealogy of large populations.
1
1982
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Origins of the coalescent: 1974-1982.
1
2000
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Phylogenomics reveals deep molluscan relationships.
2011
Comparative morphology of leaf epidermis in the Chloranthaceae.
Development of novel low-copy nuclear markers for Hieraciinae (Asteraceae) and their perspective for other tribes.
1
2012
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
STEM: species tree estimation using maximum likelihood for gene trees under coalescence.
1
2009
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
A functional phylogenomic view of the seed plants.
1
2011
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporated into the core barcode for seed plants.
Origins and evolution of the recA/RAD51 gene family: evidence for ancient gene duplication and endosymbiotic gene transfer.
1
2006
... 虽然核基因在被子植物系统发育研究中的重要性不容忽视, 但是编码蛋白质的核基因或核基因组的使用远远滞后于叶绿体基因的运用.一方面, 由于核基因组拷贝低, 导致使用传统的PCR扩增和测序得到序列信息相对困难, 有时不得不依赖于克隆测序, 工作量增大, 限制了核基因在被子植物系统发育研究中的应用.另一方面, 由于被子植物在进化过程中经历多次全基因组加倍(whole genome duplication)(Jiao et al., 2011), 近期的多倍化事件更加频繁, 据保守估计被子植物中约有50%的物种是由于多倍化形成的(Soltis et al., 2009); 其他机制导致的基因加倍事件也频繁发生(Ng & Yanofsky, 2001; Lin et al., 2006; Xu et al., 2009), 导致不同类群中存在丢失不同拷贝的现象(Xu et al., 2009).这些因素使得同源核基因是直系同源还是旁系同源的关系难以确定.尽管如此, 研究者们通过利用大量的测序资源与基因序列数据, 开始在不同亲缘关系的类群间寻找可能是直系同源关系的单拷贝基因, 并取得了一定进展.例如, Fulton等(2002)通过比较番茄(Solanum lycopersicum)的EST序列和拟南芥(Arabidopsis thaliana)的基因组序列, 鉴定了两者共有的1,025个保守的低拷贝候选直系同源基因; Wu等(2006)比较番茄、土豆(Solanum tuberosum)、辣椒(Capsicum annuum)和咖啡(Coffea arabica)的EST序列与拟南芥的基因组序列, 认为真菊类植物(euasterids)与拟南芥共有2,869个单拷贝候选直系同源基因; Duarte等(2010)鉴定了拟南芥、杨树(Pop- ulus trichocarpa)、葡萄(Vitis vinifera)和水稻(Oryza sativa)共有的959个单拷贝候选直系同源基因, 并发现其中大部分基因在小立碗藓(Physcomitrella patens)和卷柏(Selaginella moellendorffii)中也能找到; Zhang等(2012)通过比较拟南芥、杨树、桃(Prunus persica)、葡萄、猴面花(Mimulus guttatus)、水稻、高粱(Sorghum bicolor)和小立碗藓的序列鉴定了1,083个高度保守的低拷贝核基因.这些工作为低拷贝核基因的利用奠定了基础. ...
BEST: Bayesian estimation of species trees under the coalescent model.
1
2008
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Estimating species phylogenies using coalescence times among sequences.
2
2009
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
... )、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Maximum tree: a consistent estimator of the species tree.
1
2010
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
Molecular evolution and phylogenetic utility of the petD group II intron: a case study in basal angiosperms.
1
2005
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
The plant mitochondrial genome: homologous recombination as a mechanism for generating heterogeneity.
Heterotachy, an important process of protein evolution.
2
2002
... 系统发育研究中, 可能导致结果不准确的原因主要分为如下三类(Jeffroy et al., 2006; Philippe et al., 2011): (1)使用的分子标记不是真正的直系同源基因, 即hidden paralogs.被子植物中全基因组加倍和其他基因重复事件多次发生, 基因重复后不同拷贝的丢失事件使得直系同源基因的鉴定变得复杂(Jiao et al., 2011).(2)用于系统发育分析的基因长度较短或者信息含量较低时, “噪音”信息例如回复突变(reverse mutation)和平行突变(parallel mutation, 也被称为homoplasy)等, 可能会掩盖真实的系统发育信息, 从而可能引入随机误差.(3)不同物种中核苷酸成分或碱基替代模式的偏差(compositional signal)、类群之间进化速率的差异(rate signal)以及特定碱基位点替换速率的差异(heterotachous signal)等因素造成的系统误差(systematic error)(Lopez et al., 2002; Collins et al., 2005).由于世代长短、居群大小和核苷酸替换速率差异等因素, 不同被子植物类群的进化速率是不同的, 当类群间进化速率差异很大时, 进化快的类群(在进化树上表现为长枝)可能因为多重突变在某些碱基位点随机获得了相同的碱基, 从而使亲缘关系较远的物种在序列的部分位置上反而相似, 当这些非同源相似过多而足以掩盖真实的同源信息时, 就会错误地将进化快的类群聚在一起(即长枝吸引)(Goremykin et al., 2003).另外, 虽然目前常用的序列替代模型已经考虑了基因内部不同碱基位点因选择压力不同而有相异的进化速率, 但是几乎所有的模型都没有考虑选择压力会随时间发生变化, 即假设特定碱基位点的替代速率随着时间推移在类群之间保持恒定, 这与实际的分子进化过程并不一定相符(Yang, 1994; Gu et al., 1995; Lopez et al., 2002). ...
... ; Lopez et al., 2002). ...
Angiosperm diversification through time.
1
2009
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Phylogenetic pattern, diversity, and diversification of eudicots.
1
1999
... 真双子叶植物中, 毛茛目(Ranunculales)位于最基部, 是其他所有真双子叶植物的姊妹群; 清风藤科(Sabiaceae)、山龙眼目(Proteales)、黄杨目(Buxales)和昆栏树目(Trochodendrales)是真双子叶植物的其他基部类群; 其余所有真双子叶物种形成单系, 被命名为核心真双子叶(core eudicots)植物(Magallón et al., 1999).在核心真双子叶植物中, 洋二仙草目(Gunnerales)是其他类群的姊妹群; 其他类群的花多是五基数, 因此也被称为Pentapetalae, 主要由蔷薇类(rosids)和菊类植物(asterids)两大类群组成, 另外还包含系统位置尚未完全确定的几个类群, 即虎耳草目(Saxifragales)、葡萄目(Vitales)、檀香目(Santalales)、五桠果科(Dilleniaceae)和石竹目(Caryophyllales)(Judd & Olmstead, 2004; Worberg et al., 2007; Moore et al., 2010). ...
Molecular cloning and characterization of GPA1, a G protein alpha subunit gene from Arabidopsis thaliana. Proceedings of the National Academy of Sciences,
1
1990
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Granule-bound starch synthase: structure, function, and phylogenetic utility.
2
1998
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
... Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
The root of angiosperm phylogeny inferred from duplicate phytochrome genes.
1
1999
... 然而, 目前仍只有少数保守的核基因被用于研究被子植物各大类群间的亲缘关系.例如Mathews和Donoghue(1999)使用PHYA(Phytochrome A)和PHYC(Phytochrome C)推断Amborella是被子植物的最基部类群, 是其他所有被子植物的姊妹分支(Mathews & Donoghue, 1999).最近, Zhang等(2012)从分布在各大类群的73种被子植物中获得了5个十分保守的与染色体或核酸代谢相关的核基因序列, 即SMC1(structure maintenance of chromosomes family protein), SMC2(structure maintenance of chromosomes family protein), MCM5(minichromosome maintenance family protein), MSH1(DNA mismatch repair MUTS related protein)和MLH1(DNA mismatch repair protein), 发现它们的序列分化速度比叶绿体基因快, 可以提供更多的系统发育信息; 结合已完成全基因组测序的被子植物共91种(代表46个目), 他们利用这些物种中5个核基因的信息重建了被子植物系统发育关系.他们的研究结果高度支持无油樟目(Amborellales)、睡莲目和木兰藤目(Austrobaileyales)为最基部的被子植物类群, 并且在多个层面与叶绿体基因组的研究结果总体保持一致, 即被子植物几大分支分别为单系类群, 如单子叶、真双子叶、木兰类、真双子叶中的蔷薇类和菊类, 以及各个目的划分等(图1; 细节请见第2节), 同时在许多之前有争议的分类位置上也得到了有高支持率的新结果. ...
Phylogeny of Saururaceae based on mitochondrial matR gene sequence data.
1
2002
... 叶绿体是植物特有的细胞器, 其环状基因组DNA被两个反向重复序列(inverted repeat sequence, IR)分为大单拷贝区(large single copy, LSC)和小单拷贝区(small single copy, SSC).虽然不同物种中叶绿体DNA(cpDNA)拷贝数目有差异, 但基因组成和排列相似, 基因数目几乎相同, 不同物种中的叶绿体基因是直系同源基因(ortholog), 因此利用叶绿体基因构建系统发育树时很少像使用核基因那样受旁系同源基因(paralog)的干扰; 同时, 叶绿体基因序列相对比较保守, 便于扩增和克隆, 因此在被子植物系统发育研究中得到广泛应用(Olmstead & Palmer, 1994).例如Chase等(1993)使用叶绿体基因rbcL分析了代表种子植物各类群的499个物种, 全面探讨了被子植物主要类群的系统发育关系, 是植物系统发育研究的典范.在叶绿体基因中, rbcL, atpB, ndhF和matK是比较常用的被子植物系统发育标记基因(Kim & Jansen, 1995; Soltis et al., 2000; Hilu et al., 2003; Qiu et al., 2005; Burleigh et al., 2009), 其中atpB和rbcL长度接近(氨基酸编码区序列长度大约为1,500 bp), 其核苷酸替换速率(substi- tution site/unit time)约为0.068-0.108, 序列易于比对(alignment); ndhF的序列更长(> 2,000 bp), 进化速率也相对较快; matK进化速率在叶绿体基因中最快(≥1,500 bp, ~0.141), 更适用于研究相对近缘物种之间的系统关系, 但因其插入/缺失(indel)比较多, 较其他叶绿体基因更难比对.线粒体基因如mtSSU, mtLSU, atpA, cox1, matR, atp1, nad5和rps3等也被经常使用(Qiu et al., 1999; Meng et al., 2002; Zanis et al., 2002), 其中atp1, rps3, nad5和matR等的进化速率比较慢, 核酸替换速率约为0.029-0.044, 对于研究远缘物种之间的亲缘关系很有帮助(Qiu et al., 2010). ...
Phylogeny and biogeography of the genus Ainsliaea (Asteraceae) in the Sino-Japanese region based on nuclear rDNA and plastid DNA sequence data.
1
2008
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms.
5
2007
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
... 从Hamby和Zimmer(1992)首次用18S和26S rDNA序列片段构建被子植物系统发育树, 到1998年被子植物系统发育组基于rbcL, atpB和18S rDNA的序列首次提出APG分类标准并不断修改更新至APG III(Bremer et al., 2009), 以及最近运用编码蛋白的核基因序列研究各分类阶元被子植物的系统发育关系, 人们对被子植物各类群间的进化关系有了全新的认识(Hamby & Zimmer, 1992; Jansen et al., 2007; Moore et al., 2007; Bremer et al., 2009; Zhang et al., 2012)(图1). ...
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
... 系统发育分析中随着数据量的增加虽然能尽可能降低随机误差带来的影响, 但是却可能引入系统误差; 这些误差在某些情况下还可能被放大, 从而导致产生统计学上支持率很高但却不能反映真实物种关系的进化树(Phillips et al., 2004; Jeffroy et al., 2006).因此, 相对于使用较少基因序列构建系统发育树, 用系统发育基因组学来研究物种间的亲缘关系时更需要谨慎(Soltis et al., 2004).被子植物系统发育基因组学研究中比较典型的反例即由系统误差带来的长枝吸引假象(Philippe et al., 2011), 其典型原因就是取样不当或取样量不够.被子植物基部类群的确定就出现过类似的问题.虽然很多研究表明Amborella或Amborella和Nymphaeales形成的姊妹群是被子植物的最基部类群, 但是Goremykin等(2003)测定Amborella trichopoda和Nymphaea alba的叶绿体全基因组序列后, 结合公共数据库中12个叶绿体基因组序列, 提出单子叶植物(禾本科为代表物种)为被子植物的最基部类群.然而Soltis和Stefanovic等发现在增加关键类群, 尤其是基部类群如Nymphaea、Austrobaileya、木兰类或基部单子叶植物后, Amborella或Amborella + Nymphaeales仍然是被子植物的最基部类群(Soltis & Soltis, 2004; Stefanović et al., 2004), 这一结果在增加取样量后的叶绿体系统发育基因组学研究结果中也得到了验证(Jansen et al., 2007; Moore et al., 2007).Goremykin等(2003, 2013)的结果很大程度上是由于取样不当或不够导致的, 禾本科的进化速率很快, 因此当仅以长枝的禾本科植物作为单子叶植物的代表, 以及选用买麻藤属(Gnetum)植物作为外类群时, 就会造成单子叶被吸引到基部的假象.除了选取合适的进化模型和分析方法之外, 密集采样以减少长枝, 目前被认为是降低长枝吸引影响的有效方法(Graybeal, 1998; Hillis et al., 2003).但是目前大部分实验室还是较难对物种密集测序, 而且当数据量增多时, 系统发育“噪音”也会增多, 从而可能放大系统误差.无论使用叶绿体基因组还是核基因组, 系统误差的影响都不容忽视. ...
... 目前, 系统发育基因组学的策略主要是超级矩阵(supermatrix)法和超级树(supertree)法.超级矩阵法是将所有比对后的单基因序列拼接(concatenate)成一个完整大矩阵后建树, 在叶绿体基因中已经被成功应用(Jansen et al., 2007; Moore et al., 2007, 2010), 但在核基因中的应用尚不多.华大基因研究院(BGI)正在开展测定1,000种植物转录组的计划(1KP Project)(http://www.onekp.com/), 被子植物系统发育研究中一直难以解决的重要问题都有望利用核基因系统发育基因组学得到解决, 例如被子植物5个主要分支之间的关系, 真双子叶植物内部位置不确定类群的关系等.如前所述, 超级矩阵法无法避免系统误差, 很可能会得到统计上支持率很高的拓扑结构, 实际却不能反映物种之间真实的亲缘关系, 所以也需要谨慎选择基因. ...
Phylogenetic analysis of 83 plastid genes further resolves the early diversification of eudicots.
7
2010
... 由于受研究手段的限制, 早期分类学家主要依赖于形态学、细胞学和生物化学等方面的证据进行分类, 但是形态性状有很强的环境可塑性, 以至于趋同进化(convergent evolution)和平行进化(parallel evolution)多次发生, 在一定程度上影响了它们的适用范围; 而且不同研究者判断形态性状相似性的标准有差异, 因此存在很多分类关系有争议的物种或类群.分子生物学和计算科学的快速发展, 使得利用核苷酸或氨基酸等分子性状阐明被子植物间的亲缘关系成为可能.经过二十余年的发展, 分子系统学研究也从最初的基于单个基因(Hamby & Zimmer, 1992; Chase et al., 1993; Hilu & Liang, 1997)向联合多个基因(Qiu et al., 1999; Soltis et al., 1999), 乃至整个细胞器基因组的方向发展(Jansen et al., 2007; Moore et al., 2007, 2010).世界各地的植物系统学家通力合作, 构建了现代的被子植物系统发育框架(Judd & Olmstead, 2004; Soltis et al., 2005, 2011; Soltis & Soltis, 2013), 例如提出并不断改善了被子植物APG(1998, 2003, 2009)分类系统, 并已更新至APG III(APG, 1998, 2003; Bremer et al., 2009), 使人们对被子植物主要类群间的亲缘关系以及演化过程有了全新的认识. ...
... 测序技术的成熟与计算能力的提高促使越来越多的植物叶绿体基因组被测序, 叶绿体系统发育基因组学(phylogenomics)也逐步成为被子植物系统发育研究的趋势之一(Moore et al., 2010; Jansen et al., 2011; Weng et al., 2012).例如, Jansen等(2007)利用64个物种的叶绿体基因组的81个基因重建了被子植物大类群之间的亲缘关系; Moore等(2010)根据86个物种的83个叶绿体基因重建了真双子叶(eudicots)植物内部大类群之间的亲缘关系.叶绿体基因组也开始被应用于被子植物科以下分类阶元的进化研究.例如Zhang等(2011)成功运用叶绿体基因组序列研究了竹亚科(Bambusoideae)在禾本科(Poaceae)中的系统位置, 提出了竹亚科与早熟禾亚科(Pooideae)为姊妹群的假说; Wu和Ge(2012)利用22个禾本科植物的叶绿体基因组成功解决了禾本科三大类群即竹亚科、早熟禾亚科和稻亚科(Ehrhartoidea)之间的亲缘关系; Xi等(2012)对约占热带雨林物种数40%的金虎尾目(Malpighiales)进行了系统发育重建, 并鉴定出了新的12个分支; Sun等(2013)推断基部真双子叶植物昆栏树科(Trochon- dendraceae)在30-44百万年前分化形成昆栏树属(Trochodendron)和水青树属(Tetracentron). ...
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
... 真双子叶植物中, 毛茛目(Ranunculales)位于最基部, 是其他所有真双子叶植物的姊妹群; 清风藤科(Sabiaceae)、山龙眼目(Proteales)、黄杨目(Buxales)和昆栏树目(Trochodendrales)是真双子叶植物的其他基部类群; 其余所有真双子叶物种形成单系, 被命名为核心真双子叶(core eudicots)植物(Magallón et al., 1999).在核心真双子叶植物中, 洋二仙草目(Gunnerales)是其他类群的姊妹群; 其他类群的花多是五基数, 因此也被称为Pentapetalae, 主要由蔷薇类(rosids)和菊类植物(asterids)两大类群组成, 另外还包含系统位置尚未完全确定的几个类群, 即虎耳草目(Saxifragales)、葡萄目(Vitales)、檀香目(Santalales)、五桠果科(Dilleniaceae)和石竹目(Caryophyllales)(Judd & Olmstead, 2004; Worberg et al., 2007; Moore et al., 2010). ...
... 其次, 除了被子植物5个主要分支的亲缘关系还不能确定外, 各大类群内部主要分支的关系也没有完全厘清(图2).真双子叶植物中, 除了最基部的毛茛目外, 其他基部类群中的清风藤科与山龙眼目、黄杨目与昆栏树目是否互为姊妹群, 尚需进一步的研究(Moore et al., 2010; Zhang et al., 2011).核心真双子叶植物中除了蔷薇类和菊类植物两大分支外, 其他重要分支, 即虎耳草目、檀香目、葡萄目、五桠果科和石竹目的系统发育位置也还没有最终确定(图1).蔷薇类内部的很多关系也有争论(Finet et al., 2010; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... ; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... 目前, 系统发育基因组学的策略主要是超级矩阵(supermatrix)法和超级树(supertree)法.超级矩阵法是将所有比对后的单基因序列拼接(concatenate)成一个完整大矩阵后建树, 在叶绿体基因中已经被成功应用(Jansen et al., 2007; Moore et al., 2007, 2010), 但在核基因中的应用尚不多.华大基因研究院(BGI)正在开展测定1,000种植物转录组的计划(1KP Project)(http://www.onekp.com/), 被子植物系统发育研究中一直难以解决的重要问题都有望利用核基因系统发育基因组学得到解决, 例如被子植物5个主要分支之间的关系, 真双子叶植物内部位置不确定类群的关系等.如前所述, 超级矩阵法无法避免系统误差, 很可能会得到统计上支持率很高的拓扑结构, 实际却不能反映物种之间真实的亲缘关系, 所以也需要谨慎选择基因. ...
Phylogenetic analysis of the plastid inverted repeat for 244 species: insights into deeper-level angiosperm relationships from a long, slowly evolving sequence region.
1
2011
... 关于这5个类群的亲缘关系, 不同研究者根据不同基因或不同数目的物种得到的结果不尽相同, 目前已发表的拓扑结构约有15种, 但支持率都不高(Soltis et al., 1999; Jansen et al., 2007; Moore et al., 2011; Zhang et al., 2012).其中, 有研究分别将单子叶植物、单子叶植物与金鱼藻、金鱼藻和金粟兰、木兰类、金粟兰、木兰类与金粟兰作为这5个分支中的最基部类群.例如, Qiu等(1999)联合2个叶绿体基因、2个线粒体基因和18S rDNA的序列构建了包含105个种的系统树, 认为真双子叶植物与木兰类植物是姊妹群, 金粟兰目是两者的姊妹群, 而单子叶植物与金鱼藻目形成的姊妹群位于最基部.有一些源于核基因、线粒体和叶绿体基因的证据也曾认为金鱼藻目与单子叶植物是姊妹群(Soltis et al., 1997; Savolainen et al., 2000), 但金鱼藻目是比较特化的类群, 而单子叶植物多为草本, 进化速率快, 两者在进化树上都表现为长枝, 因此有研究者认为二者聚为姊妹群很可能是因为长枝吸引(long-branch attraction, LBA)(Soltis et al., 2005). ...
Phylogeny of the ants: diversification in the age of angiosperms.
1
2006
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
The continuing search: low-copy nuclear sequences for lower-level plant molecular phylogenetic studies.
1
2004
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Reconciling gene and genome duplication events: using multiple nuclear gene families to infer the phylogeny of the aquatic plant family Pontederiaceae.
1
2011
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
Function and evolution of the plant MADS-box gene family.
1
2001
... 虽然核基因在被子植物系统发育研究中的重要性不容忽视, 但是编码蛋白质的核基因或核基因组的使用远远滞后于叶绿体基因的运用.一方面, 由于核基因组拷贝低, 导致使用传统的PCR扩增和测序得到序列信息相对困难, 有时不得不依赖于克隆测序, 工作量增大, 限制了核基因在被子植物系统发育研究中的应用.另一方面, 由于被子植物在进化过程中经历多次全基因组加倍(whole genome duplication)(Jiao et al., 2011), 近期的多倍化事件更加频繁, 据保守估计被子植物中约有50%的物种是由于多倍化形成的(Soltis et al., 2009); 其他机制导致的基因加倍事件也频繁发生(Ng & Yanofsky, 2001; Lin et al., 2006; Xu et al., 2009), 导致不同类群中存在丢失不同拷贝的现象(Xu et al., 2009).这些因素使得同源核基因是直系同源还是旁系同源的关系难以确定.尽管如此, 研究者们通过利用大量的测序资源与基因序列数据, 开始在不同亲缘关系的类群间寻找可能是直系同源关系的单拷贝基因, 并取得了一定进展.例如, Fulton等(2002)通过比较番茄(Solanum lycopersicum)的EST序列和拟南芥(Arabidopsis thaliana)的基因组序列, 鉴定了两者共有的1,025个保守的低拷贝候选直系同源基因; Wu等(2006)比较番茄、土豆(Solanum tuberosum)、辣椒(Capsicum annuum)和咖啡(Coffea arabica)的EST序列与拟南芥的基因组序列, 认为真菊类植物(euasterids)与拟南芥共有2,869个单拷贝候选直系同源基因; Duarte等(2010)鉴定了拟南芥、杨树(Pop- ulus trichocarpa)、葡萄(Vitis vinifera)和水稻(Oryza sativa)共有的959个单拷贝候选直系同源基因, 并发现其中大部分基因在小立碗藓(Physcomitrella patens)和卷柏(Selaginella moellendorffii)中也能找到; Zhang等(2012)通过比较拟南芥、杨树、桃(Prunus persica)、葡萄、猴面花(Mimulus guttatus)、水稻、高粱(Sorghum bicolor)和小立碗藓的序列鉴定了1,083个高度保守的低拷贝核基因.这些工作为低拷贝核基因的利用奠定了基础. ...
Molecular phylogeny and habitat diversification of the genus Farfugium (Asteraceae) based on nuclear rDNA and plastid DNA.
1
2010
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
Inparanoid: a comprehensive database of eukaryotic orthologs.
Nuclear DNA-based markers for plant evolutionary biology.
1
1997
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Archaefructaceae, a new basal angiosperm family.
1
2002
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
A eudicot from the Early Cretaceous of China.
2011
Complete plastid genome sequencing of trochodendraceae reveals a significant expansion of the inverted repeat and suggests a paleogene divergence between the two extant species.
2013
The morphology and relationships of the Chloranthaceae.
Phylogeny and biogeography of the rice tribe (Oryzeae): evidence from combined analysis of 20 chloroplast fragments.
1
2010
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
Phylogenetic utility of the Glycerol-3-Phosphate acyltransferase gene: evolution and implications in Paeonia (Paeoniaceae).
1
2001
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
Classification and geography of the flowering plants.
Phylogeny, rate variation, and genome size evolution of Pelargonium (Geraniaceae).
1
2012
... 测序技术的成熟与计算能力的提高促使越来越多的植物叶绿体基因组被测序, 叶绿体系统发育基因组学(phylogenomics)也逐步成为被子植物系统发育研究的趋势之一(Moore et al., 2010; Jansen et al., 2011; Weng et al., 2012).例如, Jansen等(2007)利用64个物种的叶绿体基因组的81个基因重建了被子植物大类群之间的亲缘关系; Moore等(2010)根据86个物种的83个叶绿体基因重建了真双子叶(eudicots)植物内部大类群之间的亲缘关系.叶绿体基因组也开始被应用于被子植物科以下分类阶元的进化研究.例如Zhang等(2011)成功运用叶绿体基因组序列研究了竹亚科(Bambusoideae)在禾本科(Poaceae)中的系统位置, 提出了竹亚科与早熟禾亚科(Pooideae)为姊妹群的假说; Wu和Ge(2012)利用22个禾本科植物的叶绿体基因组成功解决了禾本科三大类群即竹亚科、早熟禾亚科和稻亚科(Ehrhartoidea)之间的亲缘关系; Xi等(2012)对约占热带雨林物种数40%的金虎尾目(Malpighiales)进行了系统发育重建, 并鉴定出了新的12个分支; Sun等(2013)推断基部真双子叶植物昆栏树科(Trochon- dendraceae)在30-44百万年前分化形成昆栏树属(Trochodendron)和水青树属(Tetracentron). ...
Timing the radiations of leaf beetles: Hispines on gingers from latest Cretaceous to recent.
1
2000
... 被子植物也称有花植物, 约起源于一亿八千万年前的早侏罗纪(early Jurassic)甚至可能更早(Sun et al., 2002; Crane et al., 2004; Smith et al., 2010), 并在短时间内快速分化形成种类繁多、形态各异的类群(Davies et al., 2004; Magallón & Castillo, 2009).早期被子植物的过渡型化石很少被发现, 因此, 被子植物在演化过程中呈现出的快速起源及其在较短地质时期内物种多样化的现象被达尔文称为“令人烦恼之谜”(abominable mystery)(Darwin & Seward, 1903; Friedman, 2009).已知的被子植物约有35万种, 占整个植物界的一半, 在现今陆地生态系统中占绝对优势, 为人类提供氧气、食物、木材、纤维、医药和能源等重要生存物资(Tilman et al., 2002), 也为其他生物类群如昆虫(Farrell, 1998; Wilf et al., 2000; Moreau et al., 2006)、两栖类(Roelants et al., 2007)、哺乳动物(Bininda-Emonds et al., 2007; Roelants et al., 2007)和蕨类植物(Schneider et al., 2004)的扩张或再次繁荣带来新的契机. ...
Horizontal gene transfer from flowering plants to Gnetum.
1
2003
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...
Phylogeny of basal eudicots: insights from non-coding and rapidly evolving DNA.
1
2007
... 真双子叶植物中, 毛茛目(Ranunculales)位于最基部, 是其他所有真双子叶植物的姊妹群; 清风藤科(Sabiaceae)、山龙眼目(Proteales)、黄杨目(Buxales)和昆栏树目(Trochodendrales)是真双子叶植物的其他基部类群; 其余所有真双子叶物种形成单系, 被命名为核心真双子叶(core eudicots)植物(Magallón et al., 1999).在核心真双子叶植物中, 洋二仙草目(Gunnerales)是其他类群的姊妹群; 其他类群的花多是五基数, 因此也被称为Pentapetalae, 主要由蔷薇类(rosids)和菊类植物(asterids)两大类群组成, 另外还包含系统位置尚未完全确定的几个类群, 即虎耳草目(Saxifragales)、葡萄目(Vitales)、檀香目(Santalales)、五桠果科(Dilleniaceae)和石竹目(Caryophyllales)(Judd & Olmstead, 2004; Worberg et al., 2007; Moore et al., 2010). ...
How much data are needed to resolve a difficult phylogeny? Case study in Lamiales.
1
2005
... 目前被子植物系统发育研究主要分为两大类型: 第一类是物种数目很多但基因数目较少(Burleigh et al., 2009; Qiu et al., 2010; Soltis et al., 2011).当选取的基因序列较短或包含的系统发育信息少时, 并不能解决所有关注的问题, 或者很可能产生随机误差.降低随机误差的方法是选择系统发育信息充足的直系同源基因, 加大基因数据量(Wortley et al., 2005; Jian et al., 2008), 但是当物种数目太多时, 每增加一个基因片段信息都会增加工作量和计算时间.第二类研究是物种数量较少但基因数目很多, 即系统发育基因组学(Delsuc et al., 2005).被子植物的叶绿体系统发育基因组学在近几年取得了很大的进展, 例如Jansen等(2007)用81个叶绿体基因构建包含64个物种的系统树, Moore等(2007)用61个叶绿体基因(约42,000 bp)构建包含45个物种的系统树. ...
Combining bioinformatics and phylogenetics to identify large sets of single-copy orthologous genes (COSII) for comparative, evolutionary and systematic studies: a test case in the euasterid plant clade.
2006
The phylogeny of the BEP clade in grasses revisited: evidence from the whole-genome sequences of chloroplasts.
2012
Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation Malpighiales.
2
2012
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
... 被子植物在起源和漫长演化过程的不同时期中经历了若干次不同程度的扩张, 有些分支在较短地质年代内爆发式形成大类群, 导致了被子植物各个种群大小不一, 而且产生了丰富的物种多样性 (Davies et al., 2004).研究表明, 除了5个分支形成时经历了快速物种扩张之外(Moore et al., 2007), 现存真双子叶的主要类群也源自至少1次物种扩张事件, 蔷薇类(Wang et al., 2009)、金虎尾目(Xi et al., 2012)、虎耳草目(Fishbein et al., 2001; Jian et al., 2008)和石竹目(Cuénoud et al., 2002)等在演化过程中也都经历了物种扩张.被子植物这些古老的物种扩张事件使各类群在形态和分子序列上的共有祖征较少, 系统发育信号较少, 从而导致物种亲缘关系难以解决(Rokas & Carroll, 2006), 在进化树上表现为“梳齿”(polytomies)结构. ...
Evolution of F-box genes in plants: different modes of sequence divergence and their relationships with functional diversification.
2
2009
... 虽然核基因在被子植物系统发育研究中的重要性不容忽视, 但是编码蛋白质的核基因或核基因组的使用远远滞后于叶绿体基因的运用.一方面, 由于核基因组拷贝低, 导致使用传统的PCR扩增和测序得到序列信息相对困难, 有时不得不依赖于克隆测序, 工作量增大, 限制了核基因在被子植物系统发育研究中的应用.另一方面, 由于被子植物在进化过程中经历多次全基因组加倍(whole genome duplication)(Jiao et al., 2011), 近期的多倍化事件更加频繁, 据保守估计被子植物中约有50%的物种是由于多倍化形成的(Soltis et al., 2009); 其他机制导致的基因加倍事件也频繁发生(Ng & Yanofsky, 2001; Lin et al., 2006; Xu et al., 2009), 导致不同类群中存在丢失不同拷贝的现象(Xu et al., 2009).这些因素使得同源核基因是直系同源还是旁系同源的关系难以确定.尽管如此, 研究者们通过利用大量的测序资源与基因序列数据, 开始在不同亲缘关系的类群间寻找可能是直系同源关系的单拷贝基因, 并取得了一定进展.例如, Fulton等(2002)通过比较番茄(Solanum lycopersicum)的EST序列和拟南芥(Arabidopsis thaliana)的基因组序列, 鉴定了两者共有的1,025个保守的低拷贝候选直系同源基因; Wu等(2006)比较番茄、土豆(Solanum tuberosum)、辣椒(Capsicum annuum)和咖啡(Coffea arabica)的EST序列与拟南芥的基因组序列, 认为真菊类植物(euasterids)与拟南芥共有2,869个单拷贝候选直系同源基因; Duarte等(2010)鉴定了拟南芥、杨树(Pop- ulus trichocarpa)、葡萄(Vitis vinifera)和水稻(Oryza sativa)共有的959个单拷贝候选直系同源基因, 并发现其中大部分基因在小立碗藓(Physcomitrella patens)和卷柏(Selaginella moellendorffii)中也能找到; Zhang等(2012)通过比较拟南芥、杨树、桃(Prunus persica)、葡萄、猴面花(Mimulus guttatus)、水稻、高粱(Sorghum bicolor)和小立碗藓的序列鉴定了1,083个高度保守的低拷贝核基因.这些工作为低拷贝核基因的利用奠定了基础. ...
Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: approximate methods.
1
1994
... 系统发育研究中, 可能导致结果不准确的原因主要分为如下三类(Jeffroy et al., 2006; Philippe et al., 2011): (1)使用的分子标记不是真正的直系同源基因, 即hidden paralogs.被子植物中全基因组加倍和其他基因重复事件多次发生, 基因重复后不同拷贝的丢失事件使得直系同源基因的鉴定变得复杂(Jiao et al., 2011).(2)用于系统发育分析的基因长度较短或者信息含量较低时, “噪音”信息例如回复突变(reverse mutation)和平行突变(parallel mutation, 也被称为homoplasy)等, 可能会掩盖真实的系统发育信息, 从而可能引入随机误差.(3)不同物种中核苷酸成分或碱基替代模式的偏差(compositional signal)、类群之间进化速率的差异(rate signal)以及特定碱基位点替换速率的差异(heterotachous signal)等因素造成的系统误差(systematic error)(Lopez et al., 2002; Collins et al., 2005).由于世代长短、居群大小和核苷酸替换速率差异等因素, 不同被子植物类群的进化速率是不同的, 当类群间进化速率差异很大时, 进化快的类群(在进化树上表现为长枝)可能因为多重突变在某些碱基位点随机获得了相同的碱基, 从而使亲缘关系较远的物种在序列的部分位置上反而相似, 当这些非同源相似过多而足以掩盖真实的同源信息时, 就会错误地将进化快的类群聚在一起(即长枝吸引)(Goremykin et al., 2003).另外, 虽然目前常用的序列替代模型已经考虑了基因内部不同碱基位点因选择压力不同而有相异的进化速率, 但是几乎所有的模型都没有考虑选择压力会随时间发生变化, 即假设特定碱基位点的替代速率随着时间推移在类群之间保持恒定, 这与实际的分子进化过程并不一定相符(Yang, 1994; Gu et al., 1995; Lopez et al., 2002). ...
The pentatricopeptide repeat (PPR) gene family, a tremendous resource for plant phylogenetic studies.
1
2009
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
An empirical demonstration of using pentatricopeptide repeat (PPR) genes as plant phylogenetic tools: phylogeny of Verbenaceae and the Verbena complex.
1
2010
... 在低等分类阶元系统发育重建过程中, 越来越多的研究者开始使用核基因或其内含子区域(Strand et al., 1997; Sang, 2002; Mort & Crawford, 2004).Ness等(2011)利用5个核基因重建了水生植物雨久花科(Pontederiaceae)的系统发育关系, 解决了叶绿体基因研究结果中凤眼莲属(Eichhornia)是并系(paraphyly)的问题, 并推断出凤眼莲(E. meyeri)位于该科的基部.Kim等(2008)利用LEAFY的第二个内含子阐明了蓼科(Polygonaceae)春蓼属(Persicaria)内复杂的杂交和多倍化事件.Álvarez等(2008)在菊科千里光族(Senecioneae)中筛选出了9个候选的直系同源基因, 并通过实验获得基因序列, 验证了CesA(cellulose synthase)、CHS (chalcone synthase)、DHS(DAHP synthase)和QG8140这4个核基因的可用性.Small和Wendel(2000)、Alvarez等(2005)利用核基因Adh(alcohol dehydrogenase)、A1341和CesA1b研究了棉属(Gossypium)内的物种关系.其他众多核基因也被用于研究不同类群的亲缘关系, 包括B类MADS-box基因家族中的AP3(APETALA3)和PI(PISTILLATA)(Bailey & Doyle, 1999; Aoki et al., 2004), Adc(arginine decarboxylase)(Galloway et al., 1998), RPB2(RNA polymerase II)(Denton et al., 1998), Waxy(Mason-Gamer et al., 1998; Fortune et al., 2007), GAP3DH(glyceraldehyde-3-phosphate dehydrogenase)(Gouy & Li, 1989), ACCase(acetyl-CoA carboxylase)(Huang et al., 2002), PGK(plastid 3-phosphoglycerate kinase)(Huang et al., 2002), petD(Löhne & Borsch, 2005), GBSS1(granule-bound starch synthase)(Mason-Gamer et al., 1998; Evans et al., 2000), Gpat(glycerol-3-phosphate acyltransferase)(Tank & Sang, 2001), ncpGS(Emshwiller & Doyle, 1999), GIGANTEA(Fowler et al., 1999), GPA1(G protein α subunit)(Ma et al., 1990), GAI (GA Insensitive)(Wen et al., 2007), AGB1(G proteinβsubunit)(Weiss et al., 1994)和PPR(pentatricopeptide repeat)(Yuan et al., 2009, 2010)等.另外, 为了推动核基因在被子植物系统发育研究中的应用, 研究者们在豆科、菊科和唇形科等科中开展了寻找适用的核基因, 并设计可用于研究相应科的系统发育通用引物的工作(Choi et al., 2006; Chapman et al., 2007; Curto et al., 2012; Krak et al., 2012). ...
The root of the angiosperms revisited.
1
2002
... 叶绿体是植物特有的细胞器, 其环状基因组DNA被两个反向重复序列(inverted repeat sequence, IR)分为大单拷贝区(large single copy, LSC)和小单拷贝区(small single copy, SSC).虽然不同物种中叶绿体DNA(cpDNA)拷贝数目有差异, 但基因组成和排列相似, 基因数目几乎相同, 不同物种中的叶绿体基因是直系同源基因(ortholog), 因此利用叶绿体基因构建系统发育树时很少像使用核基因那样受旁系同源基因(paralog)的干扰; 同时, 叶绿体基因序列相对比较保守, 便于扩增和克隆, 因此在被子植物系统发育研究中得到广泛应用(Olmstead & Palmer, 1994).例如Chase等(1993)使用叶绿体基因rbcL分析了代表种子植物各类群的499个物种, 全面探讨了被子植物主要类群的系统发育关系, 是植物系统发育研究的典范.在叶绿体基因中, rbcL, atpB, ndhF和matK是比较常用的被子植物系统发育标记基因(Kim & Jansen, 1995; Soltis et al., 2000; Hilu et al., 2003; Qiu et al., 2005; Burleigh et al., 2009), 其中atpB和rbcL长度接近(氨基酸编码区序列长度大约为1,500 bp), 其核苷酸替换速率(substi- tution site/unit time)约为0.068-0.108, 序列易于比对(alignment); ndhF的序列更长(> 2,000 bp), 进化速率也相对较快; matK进化速率在叶绿体基因中最快(≥1,500 bp, ~0.141), 更适用于研究相对近缘物种之间的系统关系, 但因其插入/缺失(indel)比较多, 较其他叶绿体基因更难比对.线粒体基因如mtSSU, mtLSU, atpA, cox1, matR, atp1, nad5和rps3等也被经常使用(Qiu et al., 1999; Meng et al., 2002; Zanis et al., 2002), 其中atp1, rps3, nad5和matR等的进化速率比较慢, 核酸替换速率约为0.029-0.044, 对于研究远缘物种之间的亲缘关系很有帮助(Qiu et al., 2010). ...
Highly conserved low-copy nuclear genes as effective markers for phylogenetic analyses in angiosperms.
8
2012
... 从Hamby和Zimmer(1992)首次用18S和26S rDNA序列片段构建被子植物系统发育树, 到1998年被子植物系统发育组基于rbcL, atpB和18S rDNA的序列首次提出APG分类标准并不断修改更新至APG III(Bremer et al., 2009), 以及最近运用编码蛋白的核基因序列研究各分类阶元被子植物的系统发育关系, 人们对被子植物各类群间的进化关系有了全新的认识(Hamby & Zimmer, 1992; Jansen et al., 2007; Moore et al., 2007; Bremer et al., 2009; Zhang et al., 2012)(图1). ...
... 在蔷薇类植物中, 主要类群为固氮分支(nitrogen-fixing clade), 包括葫芦目(Cucurbitales)、壳斗目(Fagales)、蔷薇目(Rosales)和豆目(Fabales), 是支持率很高的单系群; 另外两个高支持率的分支是COM分支和锦葵类(malvids)分支, 前者包括卫矛目(Celastrales)、酢浆草目(Oxalidales)和金虎尾目(Malpighiales), 后者包括锦葵目(Malvales)、包含模式植物拟南芥的十字花目(Brassicales)、燧体木目(Crossosomatales)、无患子目(Sapindales)、十齿花目(Huerteales)和美洲苦木目(Picramniales)(Bremer et al., 2009; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012)(图2a).在菊类植物中, 大部分物种组成一个被称为真菊的单系, 而山茱萸目(Cornales)和杜鹃花目(Ericales)则是真菊的姊妹群(Bremer et al., 2009; Zhang et al., 2012)(图2b).APG III又将真菊植物分为真菊I类(也被称为唇形类lamiids)和真菊II类(也被称为桔梗类campanulids); 其中真菊I类的核心类群是唇形目(Lamiales)、茄目(Solanales)、龙胆目(Gentianles)、紫草科(Boraginaceae), 而真菊II类的核心类群是盔瓣花目(Paracryphiales)、川续断目(Dipsacales)、伞形目(Apiales)、鳞叶树目(Bruniales)、菊目(Asterales)和南鼠刺目(Escalloniales). ...
... ; Zhang et al., 2012)(图2b).APG III又将真菊植物分为真菊I类(也被称为唇形类lamiids)和真菊II类(也被称为桔梗类campanulids); 其中真菊I类的核心类群是唇形目(Lamiales)、茄目(Solanales)、龙胆目(Gentianles)、紫草科(Boraginaceae), 而真菊II类的核心类群是盔瓣花目(Paracryphiales)、川续断目(Dipsacales)、伞形目(Apiales)、鳞叶树目(Bruniales)、菊目(Asterales)和南鼠刺目(Escalloniales). ...
... 关于这5个类群的亲缘关系, 不同研究者根据不同基因或不同数目的物种得到的结果不尽相同, 目前已发表的拓扑结构约有15种, 但支持率都不高(Soltis et al., 1999; Jansen et al., 2007; Moore et al., 2011; Zhang et al., 2012).其中, 有研究分别将单子叶植物、单子叶植物与金鱼藻、金鱼藻和金粟兰、木兰类、金粟兰、木兰类与金粟兰作为这5个分支中的最基部类群.例如, Qiu等(1999)联合2个叶绿体基因、2个线粒体基因和18S rDNA的序列构建了包含105个种的系统树, 认为真双子叶植物与木兰类植物是姊妹群, 金粟兰目是两者的姊妹群, 而单子叶植物与金鱼藻目形成的姊妹群位于最基部.有一些源于核基因、线粒体和叶绿体基因的证据也曾认为金鱼藻目与单子叶植物是姊妹群(Soltis et al., 1997; Savolainen et al., 2000), 但金鱼藻目是比较特化的类群, 而单子叶植物多为草本, 进化速率快, 两者在进化树上都表现为长枝, 因此有研究者认为二者聚为姊妹群很可能是因为长枝吸引(long-branch attraction, LBA)(Soltis et al., 2005). ...
... 其次, 除了被子植物5个主要分支的亲缘关系还不能确定外, 各大类群内部主要分支的关系也没有完全厘清(图2).真双子叶植物中, 除了最基部的毛茛目外, 其他基部类群中的清风藤科与山龙眼目、黄杨目与昆栏树目是否互为姊妹群, 尚需进一步的研究(Moore et al., 2010; Zhang et al., 2011).核心真双子叶植物中除了蔷薇类和菊类植物两大分支外, 其他重要分支, 即虎耳草目、檀香目、葡萄目、五桠果科和石竹目的系统发育位置也还没有最终确定(图1).蔷薇类内部的很多关系也有争论(Finet et al., 2010; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
... 综上所述, 通过仔细筛选直系同源基因(Zhang et al., 2012), 选用适当的分子数据, 例如研究起源较早的类群之间的亲缘关系时使用氨基酸序列以减少密码子简并性带来的“噪音”, 或者使用系统发育信号强的基因降低干扰(Goremykin et al., 2009; Townsend et al., 2012; Salichos & Rokas, 2013), 使用核苷酸数据研究亲缘关系较远的类群时去除变化速率快的第三位密码子或位点(Jeffroy et al., 2006; Goremykin et al., 2010), 适当增加物种或选取进化较慢的物种(Graybeal, 1998), 开发新的符合基因序列进化过程的替换模型, 寻找其他序列特征例如插入缺失等策略来尽量降低各种误差的影响显得非常重要. ...
... 超级树法即按照溯祖理论(coalescent model)(Kingman, 1982, 2000)基于大量单基因树推断物种树, 常用的分析软件有BEST(Bayesian Estimation of Species Trees)(Liu, 2008)、GLASS(Global LAteSt Split)(Liu et al., 2010)、STAR(Species Tree Estimation using Average Ranks of Coalescences)(Liu et al., 2009)、STEAC(Species Tree Estimation using Average Coalescence Times)(Liu et al., 2009)和STEM(Species Tree Estimation Using Maximum Likelihood for Gene Trees under Coalescence) (Kubatko et al., 2009)等.Burleigh等(2009)联合18,896个核基因树重建了136种植物的系统发育树, 拓扑结构框架与前人基本一致, 但是该结果并不支持基于叶绿体基因划分的真蔷薇I类和II类, 这与其他核基因的结果一致(Finet et al., 2010; Zhang et al., 2012); 并发现经历过基因重复和丢失事件的核基因可以通过超级树法构建系统发育树, 从而推动了核基因在系统发育研究中的应用(Burleigh et al., 2009).Shaw等(2013)开发了在线分析工具STRAW (Species TRee Analysis Web server), 大大方便了使用者. ...
High-throughput sequencing of six bamboo chloroplast genomes: phylogenetic implications for temperate woody bamboos (Poaceae: Bambusoideae).
2
2011
... 另外, 除了上述五大类群之外, 被子植物较低等分类阶元的研究也获得了重要进展, 例如目水平的真双子叶的金虎尾目(Malpighiales)(Xi et al., 2012)、虎耳草目(Saxifragales)(Fishbein et al., 2001)和石竹目(Caryophyllales)(Cuénoud et al., 2002; Crawley & Hilu, 2012)等, 科水平的单子叶植物的禾本科(Tang et al., 2010; Zhang et al., 2011)和雨久花科(Ness et al., 2011)等; 属水平的真双子叶植物菊科(Asteraceae)的大吴风草属(Farfugium)(Nomura et al., 2010)和兔儿风属(Ainsliaea)(Mitsui et al., 2008)等. ...
... 其次, 除了被子植物5个主要分支的亲缘关系还不能确定外, 各大类群内部主要分支的关系也没有完全厘清(图2).真双子叶植物中, 除了最基部的毛茛目外, 其他基部类群中的清风藤科与山龙眼目、黄杨目与昆栏树目是否互为姊妹群, 尚需进一步的研究(Moore et al., 2010; Zhang et al., 2011).核心真双子叶植物中除了蔷薇类和菊类植物两大分支外, 其他重要分支, 即虎耳草目、檀香目、葡萄目、五桠果科和石竹目的系统发育位置也还没有最终确定(图1).蔷薇类内部的很多关系也有争论(Finet et al., 2010; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
Mitochondrial matR sequences help to resolve deep phylogenetic relationships in rosids.
1
2007
... 其次, 除了被子植物5个主要分支的亲缘关系还不能确定外, 各大类群内部主要分支的关系也没有完全厘清(图2).真双子叶植物中, 除了最基部的毛茛目外, 其他基部类群中的清风藤科与山龙眼目、黄杨目与昆栏树目是否互为姊妹群, 尚需进一步的研究(Moore et al., 2010; Zhang et al., 2011).核心真双子叶植物中除了蔷薇类和菊类植物两大分支外, 其他重要分支, 即虎耳草目、檀香目、葡萄目、五桠果科和石竹目的系统发育位置也还没有最终确定(图1).蔷薇类内部的很多关系也有争论(Finet et al., 2010; Moore et al., 2010)(图2a), 例如COM分支曾被认为与固氮分支形成姊妹群, 这两支共同被称为真蔷薇I类, 但是最近利用5个核基因得到的具有高度支持的结果却表明, COM分支中的卫矛目和金虎尾目与锦葵类植物(原来认为属于真蔷薇II类)形成姊妹群, 这与线粒体基因的研究结果一致(Zhu et al., 2007; Finet et al., 2010; Qiu et al., 2010; Zhang et al., 2012).菊类植物中(图2b), 叶绿体证据大都支持山茱萸目和杜鹃花目依次为其他菊类植物的姊妹群, 但近来基于核基因的结果表明两者互为姊妹群, 并且支持率高达100%, 而两者合并的一支则与其他真菊类植物为姊妹群; 绞木目(Garryales)和冬青目(Aquifoliales)曾被认为分别是真菊I和II类的最基部类群, 但是近来核基因的结果则认为二者互为姊妹群, 并位于原来真菊I类的基部(Zhang et al., 2012).单子叶植物中, 除了已知菖蒲目和泽泻目依次位于基部外, 鸭跖草分支内部4个目1个科的关系尚不明晰; APG III认为天门冬目与鸭跖草分支关系最近, 而后依次是百合目、薯蓣目+露兜树目以及无叶莲目, 但是Zhang等(2012)利用5个核基因得到的结果却认为薯蓣目可能与鸭跖草分支靠近(图2c). ...
Reprint of: using nuclear gene data for plant phylogenetics: progress and prospects.
1
2013
... 虽然细胞器基因, 尤其是叶绿体基因组在研究被子植物不同分类阶元的物种亲缘关系中取得了极为重要的进展, 但是近年来对于仅使用细胞器基因研究被子植物间的亲缘关系开始产生争议.在大多数被子植物类群中, 细胞器基因是单亲遗传的, 叶绿体为双亲遗传的物种仅占约14%(Corriveau & Coleman, 1988).单亲遗传的细胞器基因只能反映母系或父系的进化历史(Day & Ellis, 1984; Fauré et al., 1994), 不能完全揭示被子植物的进化历史; 在寄生型的被子植物中, 叶绿体基因可能会部分丢失(Palmer, 1990), 线粒体基因也存在基因水平转移现象(Bergthorsson et al., 2003; Won & Renner, 2003; Davis & Wurdack, 2004); 而且, 细胞器基因系统发育信息有限, 即使应用了整个叶绿体基因组数据, 被子植物某些关键类群的系统位置仍然无法确定(Moore et al., 2010), 因此基于双亲遗传且蕴含丰富系统发育信息的核基因的系统发育研究结果显得越来越重要(Sang, 2002; Lee et al., 2011; Zimmer & Wen, 2013).另外, 核基因的使用也有助于发现和分析被子植物进化过程中的杂交(hybridization)、多倍化(polyploidization)和渐渗(introgression)等现象, 并且便于将被子植物系统发育的研究结果整合到整个生物界从而构建出生命之树(tree of life). ...



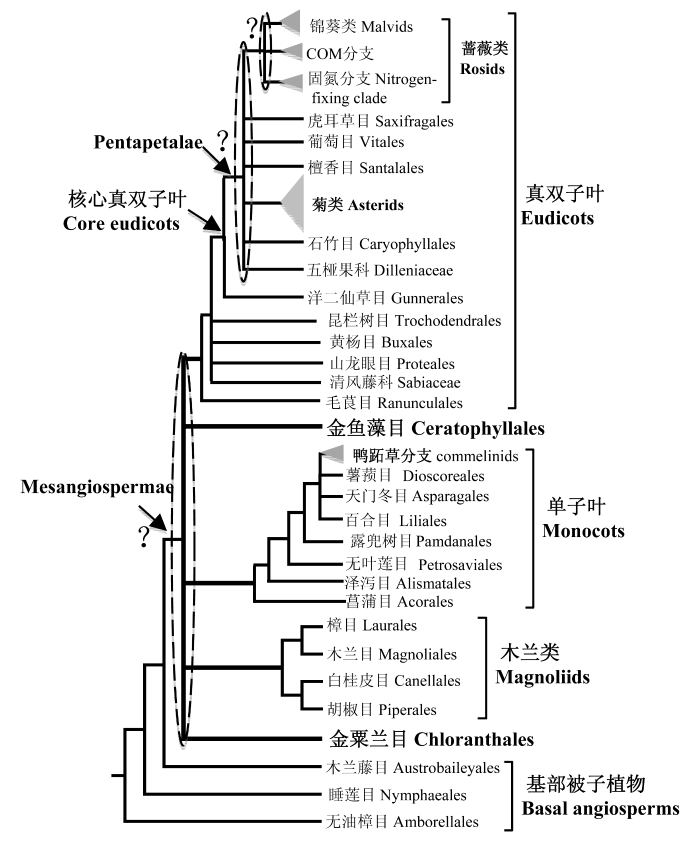
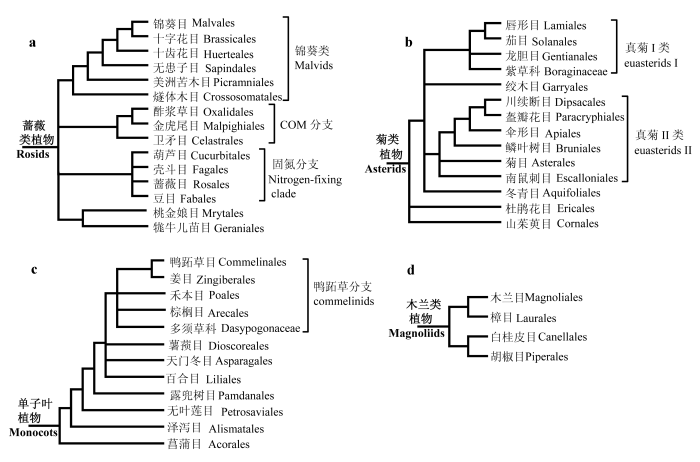
{kind=link}
{kind=link}
{kind=link}
{kind=link}