0 引言
湖泊生态系统中细菌数量众多且种类丰富, 是湖泊生态系统中食物链和食物网的重要组成部分, 驱动着湖泊生态系统中绝大多数生物活性元素的形态转化和地球化学循环。细菌作为有机化合物的主要分解和矿化者, 通过将有机污染物(如碳、氮、磷、硫等有机营养物质)分解成无机化学成分, 影响和调节着水体的环境质量, 实现水质的净化。因此全面了解湖泊生态系统中细菌群落多样性、分布特征及其在生态系统中的功能和作用, 对于未来更好地管理和维护湖泊生态环境具有深远的意义。
在自然界中, 只有不到1%的微生物可以纯培养(Amann et al ., 1995 ), 这使得在纯培养技术时代, 人们缺乏对自然界中绝大部分微生物的认识。20世纪80年代末, 微生物分子生态学技术和方法的出现, 使得人们对水生细菌群落多样性和组成有了比较深入的认识。各种分子指纹图谱(denaturing gradient gel electrophoresis, DGGE; terminal-restriction fragment-length polymorphism, T-RFLP; amplified ribosomal DNA restriction analysis, ARDRA)、荧光杂交技术(fluorescent in situ hybridization, FISH; reverse line blot hybridization, RLB)及分子克隆文库技术被应用到对海洋及湖泊水体微生物的研究之中, 增加了人们对这一微生物群体多样性及动态规律的认识。近年来方兴未艾的宏基因组学、蛋白质组学、高通量测序技术及基因芯片技术的发展, 使得人们有机会对湖泊中的细菌群落结构和功能进行更加深入而全面的研究。但迄今为止, 人们对于细菌在湖泊生态系统中如何分布、分布的机制, 及如何在生态系统中发挥作用等问题并没有清晰而明确的认识。本文拟对近年来湖泊生态系统细菌群落分布特征、形成机制、生态功能等方面的研究进行简要的梳理和评述。
1 细菌“种”的定义和多样性评价
1.1 对细菌“种”的定义
对细菌“种”的科学且合理的定义, 影响着微生物学者对微生物多样性时空分布特征的认识。但由于细菌本身的特点, 例如频繁的基因水平转移, 使得细菌的基因组存在着高度的动态变化, 导致对细菌“种”的定义存在着巨大困难。目前, 微生物学家对如何定义细菌“种”仍存在争议, 例如: 是否需要同时考虑细菌“种”在表型和基因型上的相似性, 是否需要同时考虑细菌“种”在基因、系统发育史、进化以及生态特征上的相似性(Gevers et al ., 2005 ; Logue et al ., 2008 )。目前, 对细菌“种”的常用定义为: 基因组具有一致性的一组分离菌株, 若在相同条件下对这些菌株进行检测, 它们在多项指标上具有高度的相似性(Rosselló-Mora & Amann, 2001 )。
在实际运用过程中, 细菌“种”常被简单定义为DNA-DNA杂交相似性大于70%的一组菌株(或者变性温度间的差异小于5%, 即ΔTm < 5%), 或16S rRNA基因序列的相似性大于97%的一组菌株。但这种简单的定义实际上是比较武断的, 并没有在真正意义上考虑到细菌的进化过程。一些研究者认为仅考虑16S rRNA基因序列的相似性来定义细菌“种”, 会使对不同生境中细菌多样性的认识存在偏差。Cohan(2002)提出用更小的单元“生态型”来定义细菌“种”, 并将“生态型”定义为: 占有相同或者极为相似生态位的一组菌株, 这组菌株中若出现某一个自适应变异菌株, 该菌株将通过竞争作用导致同一生态型中其他菌株的灭绝, 但不能导致其他生态型中菌株的灭绝。为了更好地认识细菌的“生态型”, 需要研究影响种内基因结合和分化的生物过程及其机理(Curtis & Sloan, 2004 )。Gevers等(2005)建议细菌“种”的界定应同时包括种内和种间表型的一致性和差异性, 且细菌“种”应该同时具备以下特征: (1)表型的一致性(生物化学指标); (2)基因型的一致性(DNA指纹图谱信息); (3)系统发育史的一致性(rRNA基因序列数据); (4)生态特征上的相似性。
1.2 评价细菌多样性的常用指标
尽管微生物研究者已认识到仅用16S rRNA基因序列的相似性来定义细菌的分类单元存在着很大弊端, 但由于研究条件和手段的限制, 目前关于微生物多样性的大部分研究, 仍然依赖于16S rRNA基因序列的相似性来解释微生物的多样性。目前, 有两种解读方法被广泛使用: 一是计算未知序列与参考序列库中序列的相似性, 将未知序列分成不同的分类单元, 例如: 种系型(phylotypes; Huse et al ., 2008 ); 二是通过比较序列与群落中其他未知序列间的相似性, 将序列分成不同的操作性分类单元(OTUs)(Schloss, 2009 ; Schloss & Westcott, 2011 )。目前第二代测序技术(高通量测序)广泛使用, 越来越多先前难以检测到的稀有细菌种类被检测出来。如何将难以培养的稀有细菌准确分类是研究者们将要面临的严峻挑战。
湖泊水体中细菌数量众多, 约105 -108 个/mL, 且种类丰富, 将每一种细菌从湖泊中都采集到几乎不可能, 因此研究者们只能采集有限的样本量, 并通过外推的方法, 估算出湖泊细菌的物种丰度和多样性。目前常用于描述物种多样性的参数有3个, 分别是: α多样性, β多样性和γ多样性(Whittaker, 1960 )。
通过计算基于OTUs或种系型(种类和相对丰度)的细菌α多样性和β多样性, 可以描述细菌的多样性及其与环境因子之间的相互关系。其中用于表征α多样性的方法有两大类: (1)借助物种丰度来估算多样性, 主要包括: 参数类型(Curtis指数)、非参数类型(Chao1、ACE、Jackknife1指数和Jackknife2指数)、曲线外推法(Michaelis-Menton指数)和稀释曲线法(Coleman指数); (2)通过丰度和均匀度来估算细菌的多样性, 主要包括: Fisher’s α指数、Shannon指数和Simpson指数。用于表征细菌β多样性的主要方法有: (1)相似性(相异性)指数, 用于度量群落物种组成在时空尺度上的变化, 主要的指数包括: Sørensen指数、Jaccard指数、Simpson指数和Bray-Curtis指数; (2)系统发育β多样性(phylogenetic β-diversity), 用于度量不同区域物种库共有进化历史长短, 主要指数有PD dissimilarity指数、UniFrac指数和系统发育Sørensen指数; (3)相似性随时间和空间距离衰减的斜率。斜率的绝对值越大, 代表β多样性越高; (4)排序空间的梯度长度, 例如除趋势对应分析(detrended correspondence analysis, DCA)中的梯度长度和逐步约束对应分析(constrained correspondence analysis, CCA)中的总方差都可以作为β多样性的度量值。
尽管基于目前的微生物分子检测技术和方法, 还不能检测出环境中细菌真实的多样性。但在尽可能地增加样本量的基础上(使得样本中细菌的多样性尽可能地反映环境中真实的细菌多样性), 上述多样性指数仍可用于比较不同样本间细菌相对多样性的大小。
2 湖泊水体典型的细菌组成类群
20世纪80年代末, 基于纯培养的研究表明, 土壤、海洋和淡水等生态系统中的细菌群落结构和组成相似(Rheinheimer, 1980 ), 但分子生态学技术的出现与不断发展已逐渐打破了这种认识。Glöckner等(2000)、Zwart等(2002)和Logares等(2009)运用现代分子生态学方法比较了不同生境中的细菌多样性, 发现淡水生境的细菌组成与土壤及海洋生态系统中的细菌组成有着明显差异。Zwart等(2002)从不同淡水环境(湖泊和河流)得到的689个序列中至少有34个16S rRNA序列为典型本土淡水细菌类群, 而且它们的分布非常广泛。这些典型的淡水细菌大都隶属于α-、β-、γ-变形菌亚门(Proteobacteria)、蓝细菌门(Cyanobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)和疣微菌门(Verrucom- icrobia)。Newton等(2011)在Zwart等(2002, 2003)研究的基础上, 结合淡水生态系统中细菌多样性的最新研究成果, 发现湖泊生态系统变温层中共有21个典型的淡水细菌门类(ribosomal database project, RDP, 相似性为75%)。除Zwart等(2002)已报道的5个优势门类, 以放线菌和β-变形菌数量最为丰富; 其他的16个细菌门类仅约占所收集到的全部序列的2.6%, 包括: 酸杆菌门(Acidobacteria), 绿菌门(Chlorobi), 绿弯菌门(Chloroflexi), 纤维杆菌门(Fibrobacteres), 厚壁菌门(Firmicutes), 梭杆菌门(Fusobacteria), 芽单胞菌门(Gemmatimonadetes), 黏胶球形菌门(Lentisphaerae), 硝化螺菌门(Nitrospira), 浮霉菌门(Planctomycetes), 螺旋体门(Spirochaetes), BRC1, OD1, OP10, SR1和TM7。
在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 )。
3 细菌多样性的维持机制
在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍。 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用。细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 )。另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 )。基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中。该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”。最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 )。一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 )。同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 )。例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等)。那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战。
与Beijerinck和Baas-Becking所提出的细菌全球性分布的观点相对应的是meta-群落假说(Leibold et al ., 2004 ; Brendan Logue & Lindström, 2008 )。Hanski 和Gilpin(1991)对meta-群落的定义是: 因扩散作用而联系在一起的一系列本地群落的集合。与之前的群落理论只强调群落受当地当时的环境条件作用的观点不同的是, meta-群落假说更强调不同时空尺度(本地、区域)上群落的结构和动态过程。且meta-群落假说认为不同本地群落在时空尺度上的动态变化, 不仅可以改变本地群落结构, 而且可通过反馈作用改变区域尺度上生物群落的组成和结构(Holyoak et al ., 2005 )。Meta-群落假说由4个简单的观点组成, 它们都在一定程度上解释了细菌群落的生物地理学分布及动态规律(图1 )。
图1 meta-群落假说的4种观点对细菌群落的相似性与环境和地理距离之间的相互关系的解释(改自Logue et al ., 2008 )
Fig. 1 The interpretation of the relationship between bacterial community similarity and environmental gradients and geographic distances under four perspectives of meta- community hypothesis (revised from Logue et al ., 2008 )
(1)物种筛选观点(species-sorting perspective, 图1A ), 该观点假设环境由不同的斑块组成, 且斑块与斑块之间存在着环境异质性。不同的斑块中物种组成存在差异, 且这种差异与不同斑块间物种的交流没有关系, 因为物种的扩散作用是全球性的。物种筛选观点与Beijerinck和Baas-Becking的观点相同, 均强调细菌具有全球性的分布特征, 但本土环境特征是调节细菌组成与分布的主要因素。物种筛选观点也可以理解为: 在地球上, 任何两个环境特征完全相同的斑块其细菌组成也相同。
(2)质量效应观点(mass effects perspective, 图1B ), 该观点在支持环境异质性对细菌群落分布有选择作用的同时, 更强调细菌的迁入和迁出对本土细菌群落动态的影响。细菌因在斑块与斑块之间群落组成上不对称而发生交流, 影响着细菌的迁入和迁出。这种“源—库”间交流的动态影响着本土细菌群落结构和空间分布特征。质量效应观点认为细菌群落的动态变化受环境和空间距离的共同作用。例如, Lindström等(2006)研究发现, 湖泊水体中本土细菌的多样性受连续的入湖溪流中细菌组成的显著影响。
(3)斑块-动态观点(patch-dynamics perspective, 图1C ), 该观点认为相同的斑块是存在的, 且每一个斑块均有能力包含一定的细菌群落。这些斑块可能是已经被占领的, 也可能是未被占领的。斑块中细菌种类的多样性受细菌扩散作用的影响, 扩散作用通过向斑块输送源细菌, 从而补偿在斑块中因随机性或确定性而消失了的细菌。斑块-动态观点认为斑块中细菌群落结构不受环境因子的影响(Chase et al ., 2005 ), 而是否受斑块间地理距离的影响还无定论。斑块-动态观点与Baas-Becking的观点形成鲜明的对比, 认为群落组成的空间变化主要受历史原因的影响。
(4)中性理论(neutral perspective, 图1D ), 该理论假定所有细菌种类具有相似的竞争能力、扩散能力和适应性(Hubbell, 2001 )。不同类群细菌间的相互作用由细菌的随机扩散作用组成, 因此, 物种的减少(灭绝和迁出)和增加(迁入和物种的形成)的随机概率决定了物种的多样性。中性理论认为细菌群落组成动态变化受地理距离远近的驱动, 而不受环境条件的影响。
从meta-群落假说以及Beijerinck和Baas- Becking的观点(Loreau et al ., 2003 ; Leibold et al ., 2004 ; Martiny et al ., 2006 )出发, 可以推测出淡水细菌在全球、区域和本土3个梯度下的物种分布特征和群落多样性, 更可能是由“随机分布”和“环境决定”共同作用的结果。与Forbes(1887)提出的“湖泊正如一个微宇宙”的观点不同, 现在研究者们更多将湖泊看作一个流动系统, 认为它是更大尺度空间中的一个组成单元。湖泊不是一个与外界环境隔离的生态系统, 它与外界环境之间存在着频繁的物质、能量及基因的交流。对于水力停留时间很长的湖泊, 其细菌群落组成主要受湖泊内部环境因子的影响, 即适用物种筛选(species-sorting perspective)和Baas- Becking的观点(Brendan Logue & Lindström, 2008 )。但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 )。
4 环境因子对湖泊水体细菌多样性的影响
如果以湖岸线为界限, 将湖泊看作一个整体, 我们可以把影响湖泊细菌多样性的环境因子分为湖泊生态系统的内部环境因子和区域尺度上的外部环境因子。区域尺度上的非生物因素, 例如温度和主要离子浓度, 在不同湖泊间表现出年际变化同步性(Magnuson et al ., 1990 ), 而湖泊特有的内部环境因子, 例如食物网中生物间相互作用及物种的随机动态, 却往往抑制并减弱了区域尺度上的外界因子对细菌多样性的作用(Magnuson et al ., 2006 ; Kent et al ., 2007 )。
4.1 区域尺度上的外部环境因子
湖泊在区域尺度上的外来补水改变着湖泊营养盐、理化因子状态的同时, 也改变了湖泊内部食物网的组成, 并不断将区域尺度上的细菌群落输送至湖泊内部, 影响着湖泊系统内部细菌群落的组成和多样性。Lindström等(2006)研究了瑞典的12个不同水力停留时间(从1天到10年)的湖泊, 发现区域尺度上的外部环境因子(例如: 细菌的扩散速率)对细菌群落组成起决定作用。当水力停留时间小于200天时, 区域尺度上细菌的输入显著改变了湖泊水体细菌群落的组成和多样性。
4.2 湖泊特有的内部环境因子
影响细菌多样性的湖泊内部环境因子主要包括: 湖泊的形态特征(例如: 湖泊的大小和深度)、物理化学特征(例如: 温度、pH、盐度、无机营养盐等)、有机质(DOM)的浓度和类型以及食物网的组成和物种间的相互作用(例如: 鞭毛虫的捕食作用和病毒的溶菌作用)。
4.2.1 湖泊的大小和深度
物种的多样性与生境大小的关系一直是生态学关注的热点之一(Rosenzweig, 1995 ; Lawton, 1999 )。MacArthur和Wilson(1967)针对生物多样性与生境大小之间的关系提出“生物地理学的岛屿效应”, 它主要包括两个方面: (1)岛屿上物种数目会随着岛屿面积的增加而增加; (2)岛屿上物种数目会随着岛屿与大陆间的距离增加而下降。如果把陆地看作“海洋”, 湖泊就是陆地上的“岛屿”。那么, 湖泊中的细菌是否也如大型生物一样, 呈现出“岛屿效应”? Reche等(2005)研究位于西班牙内华达山脉上的11个高原湖泊中的细菌群落结构和多样性, 发现细菌物种数和湖泊的面积呈显著的正相关关系。但湖泊的偏僻程度并不影响细菌物种数目, 这在一定程度上表明细菌分布并不存在“地理隔离”。
细菌群落组成和多样性随湖泊深度的变化与环境异质性密切相关, 例如: 光照、溶解氧和温度随着水深增加而下降; 水体初级生产力、营养盐浓度等也沿着水深而发生相应的变化。Hollibaugh等(2001)借助于DGGE指纹图谱, 发现细菌群落的组成沿着湖泊水深呈现显著性的差异。Humayoun等(2003)通过克隆文库法, 发现湖泊中好氧层和厌氧层间细菌多样性呈现显著差异, 多样性在表层水体中较低, 且细菌主要为高G+C含量的革兰氏阳性细菌, 放线菌(Actinobacteria ); 在湖泊底层较高, 且细菌的组成以低G+C含量的革兰氏阴性细菌为主(如: 芽孢杆菌(Bacillus )和梭菌属(Clostridium ))。
4.2.2 湖泊的理化特征
温度是反映不同纬度、海拔及季节梯度上湖泊环境差异的主要因子。在全球变暖背景下, 由增温所引起的一系列湖泊生态效应也是当前和未来科学界关注的焦点之一。越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一(Simon & Wünsch, 1998 ; Adams et al ., 2010 ; Dziallas & Grossart, 2011 )。Adams等(2010)通过24个湖泊和溪流样品的野外研究和室内培养试验, 发现在温度因子的驱动下细菌群落的组成发生了显著的改变。不同的细菌种类有着不同的生态最适温度, 在最适温度下, 细菌表现出最大的适合度。因此温度可能对水体细菌群落组成具有选择作用。Wu和Hahn (2006b)对湖泊水体模式细菌Polynucleobacte 的研究进一步表明温度对水体细菌组成在属的水平上的选择作用, 温度差异促使了细菌在同一属内的分化, Polynucleobacte 属细菌在不同温度的湖泊间表现出明显的生态多样化。温度还可以作为湖泊水体细菌在季节水平上的动态变化最好的预测因子。Wu和Hahn(2006a)通过荧光原位杂交的方法研究了3年内月亮湖中模式浮游细菌Polynucleobacter 亚群(PnecB)的丰度变化, 发现PnecB细菌丰度在研究的3年内表现出相似的季节动态特征。因细菌的生物过程(包括繁殖速率、扩散作用、物种间的相互作用、基因的突变、适应进化及物种的形成)受到温度的调控(Pomeroy & Wiebe, 2001 ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加。他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”。细菌个体的大小分布特征同样受温度的影响。Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低。
pH是决定湖泊细菌群落结构的另一关键因子, 野外原位试验研究表明, 湖泊水体细菌群落结构和多样性随着水体pH变化而发生显著变化(如: Lindström et al ., 2005 ; Yannarell & Triplett, 2005 )。pH可以通过影响不同种类细菌的生长状况(包括: 绝灭、繁衍、种的形成等; Langenheder et al ., 2006 ), 直接影响多样性; 也可通过影响湖泊生态系统中的其他环境因子(如: 有机物质的分子结构; Yannarell & Triplett, 2005 )来间接影响水体细菌群落的结构和多样性。
盐度是内陆水体另一重要环境因子, 但目前相关研究较少。Wu等(2006)通过研究位于青藏高原青海-西藏段上16个高山湖泊(盐度从0.02%的淡水到22.3%的超盐水), 发现在淡水系统和超盐水系统中只有很少一部分细菌组成发生重叠。与大型生物不同的是, 湖泊水体细菌多样性随着水体盐度增加并未呈现减少趋势。细菌在高盐度水体中的高度分化是其多样性显著增加的重要机制(Wang et al ., 2011 )。
4.2.3 营养盐水平
按照营养盐的化学组成形式, 我们可以将水体营养盐分成无机营养盐和有机营养盐。它们既可通过“上行效应”直接影响水体细菌群落的组成和分布(Crump et al ., 2003 ; Pérez & Sommaruga, 2006 ; Bouvy et al ., 2011 ); 也可通过影响水体细菌的捕食者(异样鞭毛虫和纤毛虫)的组成和丰度(Nakano et al ., 1998 ), 间接影响水体细菌群落结构和多样性。
4.2.4 湖泊食物网物种间的相互作用
湖泊生态系统, 特别是浅水湖泊生态系统, 一般按初级生产者的组成分成两种类型: 一种是当营养负荷较低时, 以大型沉水植物为主要初级生产者的清水湖泊; 另一种是当营养负荷较高时, 以浮游植物(藻类)为主要初级生产者的浊水湖泊。湖泊从清水草型向浊水藻型转变时(湖泊富营养化), 生物多样性下降(Hanson & Butler, 1994 ; Riemann & Winding, 2001 ; Villaescusa et al ., 2010 )。Hanson和Butler(1994)认为湖泊从清水状态转变为浊水状态后, 虽然没有改变湖泊生态系统中的经典食物链的传递(藻类→浮游动物→鱼类), 但是却导致组成微生物食物网的所有生物的生物量显著增加(Mathes & Arndt, 1994 )。
研究表明, 大型沉水植物的生物量和组成是影响太湖中水体细菌群落结构和多样性的关键因子, 同时也是调控太湖水体细菌群落组成季节动态的主要驱动因子(Wu et al ., 2007 ; Zeng et al ., 2012 )。当湖泊水体发生富营养化, 蓝藻爆发有助于具有溶菌作用的病毒和噬细胞菌属(Cytophaga )细菌的生长, 同样显著影响着水体细菌群落结构和多样性(Rashidan & Bird, 2001 )。
原生动物与细菌间的捕食和被捕食关系构成了湖泊微生物网的基础。在捕食细菌的原生动物中, 异养鞭毛虫(HNFs)被认为是影响细菌群落结构和组成的最主要的因子之一(Šimek & Chrzanowski, 1992 ; Young, 2006 )。研究表明HNFs对细菌的捕食作用同样显著改变了细菌群落的形态分布特征, 在高强度的捕食压力下, 中等大小的细菌更容易被捕食, 使得细菌群落形态呈极小或丝状分布(Hahn & Höfle, 2001 )。除大小特征外, Wu等(2004)认为极小或丝状细菌难以被异养鞭毛虫消化也是导致其不被捕食的重要原因。尽管研究者们认识到水体病毒对细菌群落结构的形成存在重要影响(Weinbauer et al ., 2006 ), 但目前, 对病毒如何引起细菌的死亡及影响细菌组成间的竞争的研究仍非常之少。
5 湖泊水体细菌的功能
5.1 湖泊水体细菌的生产力
从生产力的角度来看, 水生生态系统浮游细菌的生产力也常常被人们提及。采用亮氨酸、14 C标记、测定分裂细菌比例的方式可以测算湖泊浮游细菌的生产力。浮游细菌生产力(数量级约为几十mg·C·m-2 ·h-1 )有着明显的季节变化, 夏季高于冬季; 在一天之中也有变化, 夜间和凌晨的生产力最大。在年际范围内, 浮游细菌的生产力与浮游植物的初级生产力有着很强的正相关, 约占浮游植物生产力的2-45%, 这种相关性主要依赖于温度因子的作用(Di Siervi et al ., 1995 )。
浮游细菌的生产力也可以细分为初级生产力和次级生产力。初级生产力是由自养细菌来完成的, 自养细菌中普遍分布的、发挥主要功能的一类是蓝细菌, 在一些研究中它们占据自养浮游细菌生产力的95%。而自养细菌占整个浮游细菌生产力的比例也同样有着季节的变化, 在7月的午后达到最大, 约为55.6%, 而在1月只有2.3%(Affronti & Marshall, 1994 )。浮游细菌的次级生产力主要表现在有机物质(OM)的降解利用上, 由异养型浮游细菌来完成, 它们是湖泊浮游细菌的主要组成部分, 主要扮演着分解者的角色。对浮游细菌分解过程的研究表明, 细菌可以产生胞外酶来分解颗粒性有机物(POM)和溶解性有机物(DOM); 对于比较复杂的腐植酸, 可能需要多种细菌的多种代谢途径来共同完成。颗粒类有机物和复杂有机物会被分解成有机物“微羽(plume)”, 这些“微羽”又能够通过信号物质吸引浮游细菌, 并进一步被分解成小分子有机物。对于有机物“微羽”的分解在水柱细菌生产力中占有很大的比例(Grossart, 2010 )。
另外值得注意的是, 水体细菌可以按生活状态分为自由浮动和聚集体(>5 μm)两种形式, 研究表明聚集体形式的细菌在生产力和能量系数上要高于自由浮动的细菌(Shchur et al ., 2002 )。
5.2 湖泊水体细菌参与的生物地球化学过程
从元素利用和循环的角度来看, 同其他生境一样, 湖泊水体中细菌在C、N、P、S等元素的生物地球化学循环过程中也发挥着关键作用。
5.2.1 氮循环
湖泊水体中参与N循环的细菌主要包括固氮细菌、硝化细菌、反硝化细菌和厌氧氨氧化细菌。
(1)湖泊中的固氮细菌发现的主要是蓝细菌的某些种类, 如念珠藻、水华束丝藻等。这些种类的蓝藻在夏季表层水体无机氮被消耗匮乏时有着很强的生存优势(Torrey & Lee, 1975 )。不过也有学者认为蓝细菌的固氮作用并不显著, 因其可直接利用来自于沉积物中的铵离子和硝酸根离子生存, 并不完全依赖于固氮作用(Ferber et al ., 2004 )。另外一些梭菌、硫螺菌属的细菌在5-10 m深的水柱可以通过异养代谢来进行固氮作用(Keirn & Brezonik, 1971 )。
(2)湖泊中的硝化细菌主要存在于湖泊氧化性水体和表层沉积物之中。但是在湖泊水体分层时期(一般在夏季), 变温层以下的水体由于获得了来自于表层水的氧气, 及来自于湖泊沉积物的还原性氮, 也能观察到明显的硝化作用, 产生硝酸盐的能力大概在0.1-23.2 mmol NO3 - -N/m-3 (Vincent & Downes, 1981 )。硝化螺旋菌、硝化单胞菌是分布较为广泛的水体硝化细菌, 富集试验发现不同来源的硝化细菌对铵离子的亲和力有所不同: 来自于深水层的硝化细菌只能利用中低浓度的氨离子(<5 μM), 而来自于沉积物的硝化细菌能够利用浓度高达12.5 μM的铵离子(Whitby et al ., 2001 )。
(3)反硝化细菌大部分存在于沉积物中, 只有在深水湖泊的底层或者局部厌氧的环境中才存在着反硝化作用。硝化-反硝化的联合产氮作用一直被认为是湖泊氮损失的主要途径。
(4)在湖泊水体中还可能存在着厌氧氨氧化细菌, 这类细菌可以在厌氧条件下利用铵离子和亚硝酸根产生氮气。有研究表明在深水湖泊中等温层以下的水体中厌氧氨氧化(最大504 nmol N·L-1 ·d-1 )和反硝化(最大74 nmol N·L-1 ·d-1 )对氮气产出有着非常大的贡献(Hamersley et al ., 2009 )。
5.2.2 硫循环
湖泊水体中与硫循环有关的细菌主要包括硫氧化细菌和硫酸盐还原细菌。其中紫硫细菌是一类化能自养不产氧细菌, 它们以硫化物为电子受体, 在一些盐碱性湖泊中是主要的初级生产者, 在分层湖泊的化学跃变层以下光线可达的水体中, 在小范围内能够积累很大的生物量, 并在湖泊中形成肉眼可见的紫色水层(Rogozin et al ., 2012 )。在化学跃变层的厌氧性水体以及蓝藻暴发后形成的厌氧环境下, 都能够检测到较高丰度的硫酸盐还原细菌(每毫升水体104 -106 个细胞)。湖泊水体细菌进行的硫氧化还原过程中常常会利用重金属作为中间电子供体或受体, 从而也被认为在重金属甲基化等过程中发挥重要作用(Teske et al ., 1998 ; Chirwa & Wang, 2000 ; Sorokin et al ., 2011 )。
5.2.3 碳、磷循环
碳和磷是构成细菌的最基本元素。在湖泊生态系统中几乎所有细菌均参与碳、磷元素的生物地球化学过程。但是目前相关研究匮乏。在湖泊水体中, 与C循环有关的一类细菌为甲基营养细菌, 它是一类以单甲基分子为营养的自养细菌, 并最终将甲基分子氧化为CO2 。甲基营养细菌在淡水水体中广泛分布, 在一些湖泊中能够达到每升水0.6-5.5×109 个细胞, 约占浮游细菌总数的10-46%(Ross et al ., 1997 )。另一类与C循环有关的是产甲烷微生物, 主要存在于湖泊沉积物等厌氧环境下, 且以产甲烷古菌为主, 本文不再详述。在湖泊生态系统中多种形态的水体浮游细菌都可以在胞内或胞外分泌磷酸酶, 进而利用各种复杂形态的有机磷, 形成不同类型的适应状态和生态位分化(Spijkerman & Coesel, 1998 )。
5.3 湖泊水体细菌功能研究的相关技术
将细菌群落结构和特定生态功能联系起来, 一直是众多微生物生态学家关心的热点。BIOLOGTM Plates、稳定性同位素、高级FISH技术、微阵列杂交、功能基因的扩增测序和定量研究、蛋白质组测序技术, 都大大促进了人们对湖泊水体细菌各种生态功能的认识。尽管有种种研究表明不同的物质循环过程会导致非常迥异的细菌群落结构(Grossart, 2010 ), 但还是难以把某一类细菌和某一特定生态功能直接联系起来。这是因为即使同一“种”的细菌, 在不同的环境下也常常会显示出截然不同的生理生态特性(Jaspers & Overmann, 2004 )。这些基本功能, 在不同门类的细菌中应该是普遍存在的, 只是典型种类的细菌已经被人们分离纯培养并为人们所熟知。分子生物学、显微成像、连续自动采样等技术的进一步发展, 将使得人们可以在更小的时空范围内比较亲缘关系上接近或遥远的细菌之间蛋白质组的差异, 追踪细菌的形态和生理特征方面的变化(Logue et al ., 2008 ), 从而可以为人们认识湖泊水体细菌的相关功能提供更有利的支持。
6 展望
尽管在过去十几年里, 分子生物学等相关技术的不断发展, 对微生物生态学的发展起了巨大的推动作用, 但关于细菌在湖泊生态系统中的分布、形成机制, 及其生态功能等仍没有清晰而明确的答案。湖泊细菌生态学仍是一门年轻的科学, 现有的湖泊细菌群落多样性的研究是十分有限的, 这限制着人们对湖泊微生物群落的进一步认识。未来的研究者们面临着更多严峻的挑战: (1)如何更加科学地界定细菌“种”的概念, 以兼顾细菌表型、基因型、系统发育史及生态特征的一致性。提高分离纯培养技术, 获得更多纯培养菌株, 依然是未来更好地研究细菌“种”的前提; (2)在区域尺度上, 如何运用景观生态学的相关概念, 例如“meta-群落假说”来研究细菌群落组成在不同斑块间的扩散作用; (3)在微观尺度上, 借助于新技术的发展, 在更小的时空范围内研究细菌群落多样性及功能特征; (4)在研究细菌群落动态及相关影响因子的基础上, 提出或验证湖泊细菌群落多样性的生态理论及假说, 完善微生物生态学相关理论框架。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Adams HE Crump BC Kling GW 2010 ) Temperature controls on aquatic bacterial production and community dynamics in arctic lakes and streams.
Environmental Microbiology , 12 , 1319 -1333 .
[本文引用: 2]
[2]
Affronti LF Marshall HG 1994 ) Using frequency of dividing cells in estimating autotrophic picoplankton growth and productivity in the Chesapeake Bay.
Hydrobiologia , 284 , 193 -203 .
[本文引用: 1]
[3]
Amann RI Ludwig W Schleifer KH 1995 ) Phylogenetic identification and in situ detection of individual microbial cells without cultivation.
Microbiological Reviews , 59 , 143 -169 .
[本文引用: 1]
[4]
Baas-Becking LGM 1934 ) Geobiologie of Inleiding tot de Milieukunde . WP Van Stockum & Zoon, The Hague, The Netherlands. (in Dutch)
[5]
Beijerinck MW 1913 ) De Infusies en de Ontdekking der Backteriën. Jaarboek van de Koninklijke Aakademie voor Wetenschappen. Müller. Amsterdam, The Netherlands. (in Dutch)
[6]
Bouvy M Bettarel Y Bouvier C Domaizon I Jacquet S Floc’h L Montanié H Mostajir B Sime-Ngando T Torréton JP Vidussi F Bouvier T 2011 ) Trophic interactions between viruses, bacteria and nanoflagellates under various nutrient conditions and simulated climate change.
Environmental Microbiology , 13 , 1842 -1857 .
[本文引用: 1]
[7]
Brendan Logue J Lindström ES 2008 ) Biogeography of bacterioplankton in inland waters.
Freshwater Reviews , 1 , 99 -114 .
[本文引用: 3]
[8]
Chase JM Amarasekare P Cottenie K Gonzalez A Holt RD Holyoak M Hoopes MF Leibold MA Loreau M Mouquet NM Shurin JB Tilman D 2005 ) Competing theories for competitive metacommunities.
In: Metacommunities: Spatial Dynamics and Ecological Communities (eds Holyoak M, Leibold MA, Holt RD), pp. 335 -354 . The University of Chicago Press , Chicago and London .
[本文引用: 1]
[9]
Chirwa EN Wang YT 2000 ) Simultaneous chromium (VI) reduction and phenol degradation in an anaerobic consortium of bacteria.
Water Research , 34 , 2376 -2384 .
[本文引用: 1]
[10]
Cohan FM 2002 ) What are bacterial species?
Annual Reviews in Microbiology , 56 , 457 -487 .
[11]
Crump BC Kling GW Bahr M Hobbie JE 2003 ) Bacterioplankton community shifts in an arctic lake correlate with seasonal changes in organic matter source.
Applied and Environmental Microbiology , 69 , 2253 -2268 .
[本文引用: 1]
[12]
Curtis TP Sloan WT 2004 ) Prokaryotic diversity and its limits: microbial community structure in nature and implications for microbial ecology.
Current Opinion in Microbiology , 7 , 221 -226 .
[本文引用: 1]
[13]
Curtis TP Sloan WT Scannell JW 2002 ) Estimating prokaryotic diversity and its limits
. Proceedings of the National Academy of Sciences, USA , 99 , 10494 -10499 .
[本文引用: 1]
[14]
Daufresne M Lengfellner K Sommer U 2009 ) Global warming benefits the small in aquatic ecosystems
. Proceedings of the National Academy of Sciences, USA , 106 , 12788 -12793 .
[15]
Di Siervi M Mariazzi AA Donadelli JL 1995 ) Bacterioplankton and phytoplankton production in a large patagonian reservoir (República Argentina).
Hydrobiologia , 297 , 123 -129 .
[本文引用: 1]
[16]
Dziallas C Grossart HP 2011 ) Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp.
Environmental Microbiology , 13 , 1632 -1641 .
[本文引用: 1]
[17]
Eiler A Bertilsson S 2004 ) Composition of freshwater bacterial communities associated with cyanobacterial blooms in four Swedish lakes.
Environmental Microbiology , 6 , 1228 -1243 .
[本文引用: 1]
[18]
Fenchel TOM Finlay BJ 2004 ) The ubiquity of small species: patterns of local and global diversity.
BioScience , 54 , 777 -784 .
[本文引用: 1]
[19]
Ferber LR Levine SN Lini A Livingston GP 2004 ) Do cyanobacteria dominate in eutrophic lakes because they fix atmospheric nitrogen?
Freshwater Biology , 49 , 690 -708 .
[本文引用: 1]
[20]
Forbes SA 1887 ) The lake as a microcosm
. Bulletin Science Association of Peoria, Illinois , 1887 , 77 -87 .
[21]
Fuhrman JA Steele JA Hewson I Schwalbach MS Brown MV Green JL Brown JH 2008 ) A latitudinal diversity gradient in planktonic marine bacteria
. Proceedings of the National Academy of Sciences, USA , 105 , 7774 -7778 .
[22]
Gevers D Cohan FM Lawrence JG Spratt BG Coenye T Feil EJ Stackebrandt E de Peer YV Vandamme P Thompson FL Swings J 2005 ) Re-evaluating prokaryotic species.
Nature Reviews Microbiology , 3 , 733 -739 .
[本文引用: 1]
[23]
Glöckner FO Zaichikov E Belkova N Denissova L Pernthaler J Pernthaler A Amann R 2000 ) Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of Actinobacteria.
Applied and Environmental Microbiology , 66 , 5053 -5065 .
[24]
Grossart HP 2010 ) Ecological consequences of bacterioplankton lifestyles: changes in concepts are needed.
Environmental Microbiology Reports , 2 , 706 -714 .
[本文引用: 2]
[25]
Hahn MW 2003 ) Isolation of strains belonging to the cosmopolitan Polynucleobacter necessarius cluster from freshwater habitats located in three climatic zones.
Applied and Environmental Microbiology , 69 , 5248 -5254 .
[本文引用: 1]
[26]
Hahn MW Höfle MG 2001 ) Grazing of protozoa and its effect on populations of aquatic bacteria.
FEMS Microbiology Ecology , 35 , 113 -121 .
[本文引用: 1]
[27]
Hahn MW Lang E Tarao M Brandt U 2011 ) Polynucleobacter rarus sp. nov., a free-living planktonic bacterium isolated from an acidic lake.
International Journal of Systematic and Evolutionary Microbiology , 61 , 781 -787 .
[本文引用: 1]
[28]
Hamersley MR Woebken D Boehrer B Schultze M Lavik G Kuypers MMM 2009 ) Water column anammox and denitrification in a temperate permanently stratified lake (Lake Rassnitzer, Germany).
Systematic and Applied Microbiology , 32 , 571 -582 .
[本文引用: 1]
[29]
Hanski I Gilpin M 1991 ) Metapopulation dynamics: brief history and conceptual domain.
Biological Journal of the Linnean Society , 42 , 3 -16 .
[30]
Hanson MA Butler MG 1994 ) Responses of plankton, turbidity, and macrophytes to biomanipulation in a shallow prairie lake.
Canadian Journal of Fisheries and Aquatic Sciences , 51 , 1180 -1188 .
[本文引用: 1]
[31]
Hollibaugh JT Wong PS Bano N Pak SK Prager EM Orrego C 2001 ) Stratification of microbial assemblages in Mono Lake, California, and response to a mixing event.
Hydrobiologia , 466 , 45 -60 .
[32]
Holyoak M Leibold MA Mouquet NM Holt RD Hoopes MF 2005 ) Metacommunities: a framework for large-scale community ecology.
In: Metacommunities: Spatial Dynamics and Ecological Communities (eds Holyoak M, Leibold MA, Holt RD), pp. 1 -31 . The University of Chicago Press , Chicago and London .
[本文引用: 1]
[33]
Hubbell SP 2001 ) The Unified Neutral Theory of Biodiversity and Biogeography . Princeton University Press , Princeton, NJ .
[本文引用: 1]
[34]
Humayoun SB Bano N Hollibaugh JT 2003 ) Depth distribution of microbial diversity in Mono Lake, a meromictic soda lake in California.
Applied and Environmental Microbiology , 69 , 1030 -1042 .
[35]
Huse SM Dethlefsen L Huber JA Welch DM Relman DA Sogin ML 2008 ) Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing.
PLoS Genetics , 4 , e1000255 .
[本文引用: 1]
[36]
Jaspers E Overmann J 2004 ) Ecological significance of microdiversity: identical 16S rRNA gene sequences can be found in bacteria with highly divergent genomes and ecophysiologies.
Applied and Environmental Microbiology , 70 , 4831 -4839 .
[本文引用: 1]
[37]
Jezberová J Jezbera J Brandt U Lindström ES Langenheder S Hahn MW 2010 ) Ubiquity of Polynucleobacter necessarius ssp. asymbioticus in lentic freshwater habitats of a heterogenous 2000 km2 area.
Environmental Microbiology , 12 , 658 -669 .
[本文引用: 1]
[38]
Jurasinski G Retzer V Beierkuhnlein C 2009 ) Inventory, differentiation, and proportional diversity: a consistent terminology for quantifying species diversity.
Oecologia , 159 , 15 -26 .
[39]
Jürgens K Güde H 1994 ) The potential importance of grazing-resistant bacteria in planktonic systems.
Marine Ecology Progress Series , 112 , 169 -188 .
[本文引用: 1]
[40]
Keirn MA Brezonik PL 1971 ) Nitrogen fixation by bacteria in Lake Mize, Florida, and in some lacustrine sediments.
Limnology and Oceanography , 16 , 720 -731 .
[本文引用: 1]
[41]
Kent AD Yannarell AC Rusak JA Triplett EW McMahon KD 2007 ) Synchrony in aquatic microbial community dynamics.
The ISME Journal , 1 , 38 -47 .
[本文引用: 1]
[42]
Langenheder S Lindström ES Tranvik LJ 2006 ) Structure and function of bacterial communities emerging from different sources under identical conditions.
Applied and Environmental Microbiology , 72 , 212 -220 .
[本文引用: 1]
[43]
Lawton JH 1999 ) Are there general laws in ecology?
Oikos , 84 , 177 -192 .
[本文引用: 1]
[44]
Leibold MA Holyoak M Mouquet N Amarasekare P Chase JM Hoopes MF Holt RD Shurin JB Law R Tilman D Loreau M Gonzalez A 2004 ) The metacommunity concept: a framework for multi-scale community ecology.
Ecology Letters , 7 , 601 -613 .
[本文引用: 2]
[45]
Lindström ES Forslund M Algesten G Bergström AK 2006 ) External control of bacterial community structure in lakes.
Limnology and Oceanography , 51 , 339 -342 .
[46]
Lindström ES Kamst-Van Agterveld MP Zwart G 2005 ) Distribution of typical freshwater bacterial groups is associated with pH, temperature, and lake water retention time.
Applied and Environmental Microbiology , 71 , 8201 -8206 .
[本文引用: 1]
[47]
Logares R Bråte J Bertilsson S Clasen JL Shalchian-Tabrizi K Rengefors K 2009 ) Infrequent marine-freshwater transitions in the microbial world.
Trends in Microbiology , 17 , 414 -422 .
[本文引用: 2]
[48]
Logue JB Bürgmann H Robinson CT 2008 ) Progress in the ecological genetics and biodiversity of freshwater bacteria.
BioScience , 58 , 103 -113 .
[本文引用: 4]
[49]
Loreau M Mouquet N Holt RD 2003 ) Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology.
Ecology Letters , 6 , 673 -679 .
[本文引用: 1]
[50]
MacArthur RH Wilson EO 1967 ) The Theory of Island Biogeography . Princeton University Press , Princeton, New Jersey .
[51]
Magnuson JJ Benson BJ Kratz TK 1990 ) Temporal coherence in the limnology of a suite of lakes in Wisconsin, USA.
Freshwater Biology , 23 , 145 -159 .
[本文引用: 1]
[52]
Magnuson JJ Kratz TK Benson BJ 2006 ) Long-term Dynamics of Lakes in the Landscape: term Ecological Research North Temperate Lakes . Oxford University Press , USA .
[本文引用: 1]
[53]
Martiny JBH Bohannan BJ Brown JH Colwell RK Fuhrman JA Green JL Horner-Devine MC Kane M Krumins JA Kuske CR Morin PJ Naeem S Øvreås L Reysenbach AL Smith VH Staley JT 2006 ) Microbial biogeography: putting microorganisms on the map.
Nature Reviews Microbiology , 4 , 102 -112 .
[本文引用: 2]
[54]
Mathes J Arndt H 1994 ) Biomass and composition of protozooplankton in relation to lake trophy in north German lakes.
Aquatic Microbial Ecology , 8 , 357 -375 .
[本文引用: 1]
[55]
Nakano SI Ishii N Manage PM Kawabata ZI 1998 ) Trophic roles of heterotrophic nanoflagellates and ciliates among planktonic organisms in a hypereutrophic pond.
Aquatic Microbial Ecology , 16 , 153 -161 .
[本文引用: 1]
[56]
Newton RJ Jones SE Eiler A McMahon KD Bertilsson S 2011 ) A guide to the natural history of freshwater lake bacteria.
Microbiology and Molecular Biology Reviews , 75 , 14 -49 .
[57]
Pérez MT Sommaruga R 2006 ) Differential effect of algal- and soil-derived dissolved organic matter on alpine lake bacterial community composition and activity.
Limnology and Oceanography , 51 , 2527 -2537 .
[本文引用: 1]
[58]
Pomeroy LR Wiebe WJ 2001 ) Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria.
Aquatic Microbial Ecology , 23 , 187 -204 .
[本文引用: 1]
[59]
Rashidan KK Bird DF 2001 ) Role of predatory bacteria in the termination of a cyanobacterial bloom.
Microbial Ecology , 41 , 97 -105 .
[本文引用: 1]
[60]
Reche I Pulido-Villena E Morales-Baquero R Casamayor EO 2005 ) Does ecosystem size determine aquatic bacterial richness?
Ecology , 86 , 1715 -1722 .
[61]
Rheinheimer G 1980 ) Aquatic Microbiology , 2nd edn. Wiley, New York.
[本文引用: 1]
[62]
Riemann L Winding A 2001 ) Community dynamics of free-living and particle-associated bacterial assemblages during a freshwater phytoplankton bloom.
Microbial Ecology , 42 , 274 -285 .
[本文引用: 1]
[63]
Rogozin DY Zykov VV Degermendzhi AG 2012 ) Ecology of purple sulfur bacteria in the highly stratified meromictic Lake Shunet (Siberia, Khakassia) in 2002-2009.
Microbiology , 81 , 727 -735 .
[本文引用: 1]
[64]
Rosenzweig ML 1995 ) Species Diversity in Space and Time . Cambridge University Press , Cambridge, UK .
[本文引用: 1]
[65]
Ross JL Boon PI Ford P Hart BT 1997 ) Detection and quantification with 16S rRNA probes of planktonic methylotrophic bacteria in a floodplain lake.
Microbial Ecology , 34 , 97 -108 .
[本文引用: 1]
[66]
Rosselló-Mora R Amann R 2001 ) The species concept for prokaryotes.
FEMS Microbiology Reviews , 25 , 39 -67 .
[本文引用: 1]
[67]
Schloss PD 2009 ) A high-throughput DNA sequence aligner for microbial ecology studies.
PLoS ONE , 4 , e8230 .
[本文引用: 1]
[68]
Schloss PD Westcott SL 2011 ) Assessing and improving methods used in operational taxonomic unit-based approaches for 16S rRNA gene sequence analysis.
Applied and Environmental Microbiology , 77 , 3219 -3226 .
[本文引用: 1]
[69]
Shchur LA Aponasenko AD Lopatin VN Makarskaya GV 2002 ) Functional characteristics of bacterioplankton with reference to its aggregation in water bodies of different types.
Biology Bulletin of the Russian Academy of Sciences , 29 , 431 -436 .
[本文引用: 1]
[70]
Šimek K Chrzanowski TH 1992 ) Direct and indirect evidence of size-selective grazing on pelagic bacteria by freshwater nanoflagellates.
Applied and Environmental Microbiology , 58 , 3715 -3720 .
[本文引用: 1]
[71]
Simon M Wünsch C 1998 ) Temperature control of bacterioplankton growth in a temperate large lake.
Aquatic Microbial Ecology , 16 , 119 -130 .
[本文引用: 1]
[72]
Sorokin DY Kuenen JG Muyzer G 2011 ) The microbial sulfur cycle at extremely haloalkaline conditions of soda lakes.
Frontiers in Microbiology , 2 , 44 .
[本文引用: 1]
[73]
Spijkerman E Coesel PFM 1998 ) Alkaline phosphatase activity in two planktonic desmid species and the possible role of an extracellular envelope.
Freshwater Biology , 39 , 503 -513 .
[本文引用: 1]
[74]
Teske A Ramsing NB Habicht K Fukui M Küver J Jørgensen BB Cohen Y 1998 ) Sulfate-reducing bacteria and their activities in cyanobacterial mats of Solar Lake (Sinai, Egypt).
Applied and Environmental Microbiology , 64 , 2943 -2951 .
[本文引用: 1]
[75]
Torrey MS Lee GF 1975 ) Nitrogen fixation in Lake Mendota, Madison, Wisconsin.
Limnology and Oceanography , 21 , 365 -378 .
[本文引用: 1]
[76]
Torsvik V Øvreås L Thingstad TF 2002 ) Prokaryotic diversity: magnitude, dynamics, and controlling factors.
Science , 296 , 1064 -1066 .
[本文引用: 1]
[77]
Villaescusa JA Casamayor EO Rochera C Velázquez D Chicote Á Quesada A Camacho A 2010 ) A close link between bacterial community composition and environmental heterogeneity in maritime Antarctic lakes.
International Microbiology , 13 , 67 -77 .
[本文引用: 1]
[78]
Vincent WF Downes MT 1981 ) Nitrate accumulation in aerobic hypolimnia: relative importance of benthic and planktonic nitrifiers in an oligotrophic lake.
Applied and Environmental Microbiology , 42 , 565 -573 .
[本文引用: 1]
[79]
Wang J Yang D Zhang Y Shen J van der Gast C Hahn MW Wu Q 2011 ) Do patterns of bacterial diversity along salinity gradients differ from those observed for macroorganisms?
PLoS ONE , 6 , e27597 .
[本文引用: 1]
[80]
Warnecke F Sommaruga R Sekar R Hofer JS Pernthaler J 2005 ) Abundances, identity, and growth state of Actinobacteria in mountain lakes of different UV transparency.
Applied and Environmental Microbiology , 71 , 5551 -5559 .
[本文引用: 1]
[81]
Weinbauer MG Hornák K Jezbera J Nedoma J Dolan JR Šimek K 2006 ) Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity.
Environmental Microbiology , 9 , 777 -788 .
[本文引用: 1]
[82]
Whitby CB Saunders JR Pickup RW McCarthy AJ 2001 ) A comparison of ammonia-oxidiser populations in eutrophic and oligotrophic basins of a large freshwater lake.
Antonie van Leeuwenhoek , 79 , 179 -188 .
[本文引用: 1]
[83]
Whittaker RH 1960 ) Vegetation of the Siskiyou mountains, Oregon and California.
Ecological Monographs , 30 , 279 -338 .
[本文引用: 1]
[84]
Whittaker RH 1972 ) Evolution and measurement of species diversity.
Taxon , 21 , 213 -251 .
[85]
Wu QL Boenigk J Hahn MW 2004 ) Successful predation of filamentous bacteria by a nanoflagellate challenges current models of flagellate bacterivory.
Applied and Environmental Microbiology , 70 , 332 -339 .
[86]
Wu QL Hahn MW 2006 a) High predictability of the seasonal dynamics of a species-like Polynucleobacter population in a freshwater lake.
Environmental Microbiology , 8 , 1660 -1666 .
[本文引用: 1]
[87]
Wu QL Hahn MW 2006 b) Differences in structure and dynamics of Polynucleobacter communities in a temperate and a subtropical lake, revealed at three phylogenetic levels.
FEMS Microbiology Ecology , 57 , 67 -79 .
[本文引用: 1]
[88]
Wu QL Zwart G Schauer M Kamst-van Agterveld MP Hahn MW 2006 ) Bacterioplankton community composition along a salinity gradient of sixteen high-mountain lakes located on the Tibetan Plateau, China.
Applied and Environmental Microbiology , 72 , 5478 -5485 .
[89]
Wu QL Zwart G Wu J Kamst-van Agterveld MP Liu S Hahn MW 2007 ) Submersed macrophytes play a key role in structuring bacterioplankton community composition in the large, shallow, subtropical Taihu Lake, China.
Environmental Microbiology , 9 , 2765 -2774 .
[本文引用: 1]
[90]
Yannarell AC Triplett EW 2005 ) Geographic and environmental sources of variation in lake bacterial community composition.
Applied and Environmental Microbiology , 71 , 227 -239 .
[本文引用: 2]
[91]
Young KD 2006 ) The selective value of bacterial shape.
Microbiology and Molecular Biology Reviews , 70 , 660 -703 .
[本文引用: 1]
[92]
Zeng J Bian YQ Xing P Wu QL 2012 ) Macrophyte species drive the variation of bacterioplankton community composition in a shallow freshwater lake.
Applied and Environmental Microbiology , 78 , 177 -184 .
[本文引用: 1]
[93]
Zwart G Crump BC Kamst-van Agterveld MP Hagen F Han SK 2002 ) Typical freshwater bacteria: an analysis of available 16S rRNA gene sequences from plankton of lakes and rivers.
Aquatic Microbial Ecology , 28 , 141 -155 .
[94]
Zwart G Van Hannen EJ Kamst-van Agterveld MP Van der Gucht K Lindström ES Van Wichelen J Lauridisen T Han SK Declerck S 2003 ) Rapid screening for freshwater bacterial groups by using reverse line blot hybridization.
Applied and Environmental Microbiology , 69 , 5875 -5883 .
[本文引用: 2]
Temperature controls on aquatic bacterial production and community dynamics in arctic lakes and streams.
2
2010
... 温度是反映不同纬度、海拔及季节梯度上湖泊环境差异的主要因子.在全球变暖背景下, 由增温所引起的一系列湖泊生态效应也是当前和未来科学界关注的焦点之一.越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一(Simon & Wünsch, 1998 ; Adams et al ., 2010 ; Dziallas & Grossart, 2011 ).Adams等(2010)通过24个湖泊和溪流样品的野外研究和室内培养试验, 发现在温度因子的驱动下细菌群落的组成发生了显著的改变.不同的细菌种类有着不同的生态最适温度, 在最适温度下, 细菌表现出最大的适合度.因此温度可能对水体细菌群落组成具有选择作用.Wu和Hahn (2006b)对湖泊水体模式细菌Polynucleobacte 的研究进一步表明温度对水体细菌组成在属的水平上的选择作用, 温度差异促使了细菌在同一属内的分化, Polynucleobacte 属细菌在不同温度的湖泊间表现出明显的生态多样化.温度还可以作为湖泊水体细菌在季节水平上的动态变化最好的预测因子.Wu和Hahn(2006a)通过荧光原位杂交的方法研究了3年内月亮湖中模式浮游细菌Polynucleobacter 亚群(PnecB)的丰度变化, 发现PnecB细菌丰度在研究的3年内表现出相似的季节动态特征.因细菌的生物过程(包括繁殖速率、扩散作用、物种间的相互作用、基因的突变、适应进化及物种的形成)受到温度的调控(Pomeroy & Wiebe, 2001 ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加.他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”.细菌个体的大小分布特征同样受温度的影响.Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低. ...
... ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加.他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”.细菌个体的大小分布特征同样受温度的影响.Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低. ...
Using frequency of dividing cells in estimating autotrophic picoplankton growth and productivity in the Chesapeake Bay.
1
1994
... 浮游细菌的生产力也可以细分为初级生产力和次级生产力.初级生产力是由自养细菌来完成的, 自养细菌中普遍分布的、发挥主要功能的一类是蓝细菌, 在一些研究中它们占据自养浮游细菌生产力的95%.而自养细菌占整个浮游细菌生产力的比例也同样有着季节的变化, 在7月的午后达到最大, 约为55.6%, 而在1月只有2.3%(Affronti & Marshall, 1994 ).浮游细菌的次级生产力主要表现在有机物质(OM)的降解利用上, 由异养型浮游细菌来完成, 它们是湖泊浮游细菌的主要组成部分, 主要扮演着分解者的角色.对浮游细菌分解过程的研究表明, 细菌可以产生胞外酶来分解颗粒性有机物(POM)和溶解性有机物(DOM); 对于比较复杂的腐植酸, 可能需要多种细菌的多种代谢途径来共同完成.颗粒类有机物和复杂有机物会被分解成有机物“微羽(plume)”, 这些“微羽”又能够通过信号物质吸引浮游细菌, 并进一步被分解成小分子有机物.对于有机物“微羽”的分解在水柱细菌生产力中占有很大的比例(Grossart, 2010 ). ...
Phylogenetic identification and in situ detection of individual microbial cells without cultivation.
1
1995
... 在自然界中, 只有不到1%的微生物可以纯培养(Amann et al ., 1995 ), 这使得在纯培养技术时代, 人们缺乏对自然界中绝大部分微生物的认识.20世纪80年代末, 微生物分子生态学技术和方法的出现, 使得人们对水生细菌群落多样性和组成有了比较深入的认识.各种分子指纹图谱(denaturing gradient gel electrophoresis, DGGE; terminal-restriction fragment-length polymorphism, T-RFLP; amplified ribosomal DNA restriction analysis, ARDRA)、荧光杂交技术(fluorescent in situ hybridization, FISH; reverse line blot hybridization, RLB)及分子克隆文库技术被应用到对海洋及湖泊水体微生物的研究之中, 增加了人们对这一微生物群体多样性及动态规律的认识.近年来方兴未艾的宏基因组学、蛋白质组学、高通量测序技术及基因芯片技术的发展, 使得人们有机会对湖泊中的细菌群落结构和功能进行更加深入而全面的研究.但迄今为止, 人们对于细菌在湖泊生态系统中如何分布、分布的机制, 及如何在生态系统中发挥作用等问题并没有清晰而明确的认识.本文拟对近年来湖泊生态系统细菌群落分布特征、形成机制、生态功能等方面的研究进行简要的梳理和评述. ...
Trophic interactions between viruses, bacteria and nanoflagellates under various nutrient conditions and simulated climate change.
1
2011
... 按照营养盐的化学组成形式, 我们可以将水体营养盐分成无机营养盐和有机营养盐.它们既可通过“上行效应”直接影响水体细菌群落的组成和分布(Crump et al ., 2003 ; Pérez & Sommaruga, 2006 ; Bouvy et al ., 2011 ); 也可通过影响水体细菌的捕食者(异样鞭毛虫和纤毛虫)的组成和丰度(Nakano et al ., 1998 ), 间接影响水体细菌群落结构和多样性. ...
Biogeography of bacterioplankton in inland waters.
3
2008
... 与Beijerinck和Baas-Becking所提出的细菌全球性分布的观点相对应的是meta-群落假说(Leibold et al ., 2004 ; Brendan Logue & Lindström, 2008 ).Hanski 和Gilpin(1991)对meta-群落的定义是: 因扩散作用而联系在一起的一系列本地群落的集合.与之前的群落理论只强调群落受当地当时的环境条件作用的观点不同的是, meta-群落假说更强调不同时空尺度(本地、区域)上群落的结构和动态过程.且meta-群落假说认为不同本地群落在时空尺度上的动态变化, 不仅可以改变本地群落结构, 而且可通过反馈作用改变区域尺度上生物群落的组成和结构(Holyoak et al ., 2005 ).Meta-群落假说由4个简单的观点组成, 它们都在一定程度上解释了细菌群落的生物地理学分布及动态规律(图1 ). ...
... 从meta-群落假说以及Beijerinck和Baas- Becking的观点(Loreau et al ., 2003 ; Leibold et al ., 2004 ; Martiny et al ., 2006 )出发, 可以推测出淡水细菌在全球、区域和本土3个梯度下的物种分布特征和群落多样性, 更可能是由“随机分布”和“环境决定”共同作用的结果.与Forbes(1887)提出的“湖泊正如一个微宇宙”的观点不同, 现在研究者们更多将湖泊看作一个流动系统, 认为它是更大尺度空间中的一个组成单元.湖泊不是一个与外界环境隔离的生态系统, 它与外界环境之间存在着频繁的物质、能量及基因的交流.对于水力停留时间很长的湖泊, 其细菌群落组成主要受湖泊内部环境因子的影响, 即适用物种筛选(species-sorting perspective)和Baas- Becking的观点(Brendan Logue & Lindström, 2008 ).但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 ). ...
... ).但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 ). ...
Competing theories for competitive metacommunities.
1
2005
... (3)斑块-动态观点(patch-dynamics perspective, 图1C ), 该观点认为相同的斑块是存在的, 且每一个斑块均有能力包含一定的细菌群落.这些斑块可能是已经被占领的, 也可能是未被占领的.斑块中细菌种类的多样性受细菌扩散作用的影响, 扩散作用通过向斑块输送源细菌, 从而补偿在斑块中因随机性或确定性而消失了的细菌.斑块-动态观点认为斑块中细菌群落结构不受环境因子的影响(Chase et al ., 2005 ), 而是否受斑块间地理距离的影响还无定论.斑块-动态观点与Baas-Becking的观点形成鲜明的对比, 认为群落组成的空间变化主要受历史原因的影响. ...
Simultaneous chromium (VI) reduction and phenol degradation in an anaerobic consortium of bacteria.
1
2000
... 湖泊水体中与硫循环有关的细菌主要包括硫氧化细菌和硫酸盐还原细菌.其中紫硫细菌是一类化能自养不产氧细菌, 它们以硫化物为电子受体, 在一些盐碱性湖泊中是主要的初级生产者, 在分层湖泊的化学跃变层以下光线可达的水体中, 在小范围内能够积累很大的生物量, 并在湖泊中形成肉眼可见的紫色水层(Rogozin et al ., 2012 ).在化学跃变层的厌氧性水体以及蓝藻暴发后形成的厌氧环境下, 都能够检测到较高丰度的硫酸盐还原细菌(每毫升水体104 -106 个细胞).湖泊水体细菌进行的硫氧化还原过程中常常会利用重金属作为中间电子供体或受体, 从而也被认为在重金属甲基化等过程中发挥重要作用(Teske et al ., 1998 ; Chirwa & Wang, 2000 ; Sorokin et al ., 2011 ). ...
What are bacterial species?
2002
Bacterioplankton community shifts in an arctic lake correlate with seasonal changes in organic matter source.
1
2003
... 按照营养盐的化学组成形式, 我们可以将水体营养盐分成无机营养盐和有机营养盐.它们既可通过“上行效应”直接影响水体细菌群落的组成和分布(Crump et al ., 2003 ; Pérez & Sommaruga, 2006 ; Bouvy et al ., 2011 ); 也可通过影响水体细菌的捕食者(异样鞭毛虫和纤毛虫)的组成和丰度(Nakano et al ., 1998 ), 间接影响水体细菌群落结构和多样性. ...
Prokaryotic diversity and its limits: microbial community structure in nature and implications for microbial ecology.
1
2004
... 在实际运用过程中, 细菌“种”常被简单定义为DNA-DNA杂交相似性大于70%的一组菌株(或者变性温度间的差异小于5%, 即ΔTm < 5%), 或16S rRNA基因序列的相似性大于97%的一组菌株.但这种简单的定义实际上是比较武断的, 并没有在真正意义上考虑到细菌的进化过程.一些研究者认为仅考虑16S rRNA基因序列的相似性来定义细菌“种”, 会使对不同生境中细菌多样性的认识存在偏差.Cohan(2002)提出用更小的单元“生态型”来定义细菌“种”, 并将“生态型”定义为: 占有相同或者极为相似生态位的一组菌株, 这组菌株中若出现某一个自适应变异菌株, 该菌株将通过竞争作用导致同一生态型中其他菌株的灭绝, 但不能导致其他生态型中菌株的灭绝.为了更好地认识细菌的“生态型”, 需要研究影响种内基因结合和分化的生物过程及其机理(Curtis & Sloan, 2004 ).Gevers等(2005)建议细菌“种”的界定应同时包括种内和种间表型的一致性和差异性, 且细菌“种”应该同时具备以下特征: (1)表型的一致性(生物化学指标); (2)基因型的一致性(DNA指纹图谱信息); (3)系统发育史的一致性(rRNA基因序列数据); (4)生态特征上的相似性. ...
Estimating prokaryotic diversity and its limits
1
2002
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
Global warming benefits the small in aquatic ecosystems
2009
Bacterioplankton and phytoplankton production in a large patagonian reservoir (República Argentina).
1
1995
... 从生产力的角度来看, 水生生态系统浮游细菌的生产力也常常被人们提及.采用亮氨酸、14 C标记、测定分裂细菌比例的方式可以测算湖泊浮游细菌的生产力.浮游细菌生产力(数量级约为几十mg·C·m-2 ·h-1 )有着明显的季节变化, 夏季高于冬季; 在一天之中也有变化, 夜间和凌晨的生产力最大.在年际范围内, 浮游细菌的生产力与浮游植物的初级生产力有着很强的正相关, 约占浮游植物生产力的2-45%, 这种相关性主要依赖于温度因子的作用(Di Siervi et al ., 1995 ). ...
Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp.
1
2011
... 温度是反映不同纬度、海拔及季节梯度上湖泊环境差异的主要因子.在全球变暖背景下, 由增温所引起的一系列湖泊生态效应也是当前和未来科学界关注的焦点之一.越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一(Simon & Wünsch, 1998 ; Adams et al ., 2010 ; Dziallas & Grossart, 2011 ).Adams等(2010)通过24个湖泊和溪流样品的野外研究和室内培养试验, 发现在温度因子的驱动下细菌群落的组成发生了显著的改变.不同的细菌种类有着不同的生态最适温度, 在最适温度下, 细菌表现出最大的适合度.因此温度可能对水体细菌群落组成具有选择作用.Wu和Hahn (2006b)对湖泊水体模式细菌Polynucleobacte 的研究进一步表明温度对水体细菌组成在属的水平上的选择作用, 温度差异促使了细菌在同一属内的分化, Polynucleobacte 属细菌在不同温度的湖泊间表现出明显的生态多样化.温度还可以作为湖泊水体细菌在季节水平上的动态变化最好的预测因子.Wu和Hahn(2006a)通过荧光原位杂交的方法研究了3年内月亮湖中模式浮游细菌Polynucleobacter 亚群(PnecB)的丰度变化, 发现PnecB细菌丰度在研究的3年内表现出相似的季节动态特征.因细菌的生物过程(包括繁殖速率、扩散作用、物种间的相互作用、基因的突变、适应进化及物种的形成)受到温度的调控(Pomeroy & Wiebe, 2001 ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加.他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”.细菌个体的大小分布特征同样受温度的影响.Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低. ...
Composition of freshwater bacterial communities associated with cyanobacterial blooms in four Swedish lakes.
1
2004
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
The ubiquity of small species: patterns of local and global diversity.
1
2004
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
Do cyanobacteria dominate in eutrophic lakes because they fix atmospheric nitrogen?
1
2004
... (1)湖泊中的固氮细菌发现的主要是蓝细菌的某些种类, 如念珠藻、水华束丝藻等.这些种类的蓝藻在夏季表层水体无机氮被消耗匮乏时有着很强的生存优势(Torrey & Lee, 1975 ).不过也有学者认为蓝细菌的固氮作用并不显著, 因其可直接利用来自于沉积物中的铵离子和硝酸根离子生存, 并不完全依赖于固氮作用(Ferber et al ., 2004 ).另外一些梭菌、硫螺菌属的细菌在5-10 m深的水柱可以通过异养代谢来进行固氮作用(Keirn & Brezonik, 1971 ). ...
The lake as a microcosm
1887
A latitudinal diversity gradient in planktonic marine bacteria
2008
Re-evaluating prokaryotic species.
1
2005
... 对细菌“种”的科学且合理的定义, 影响着微生物学者对微生物多样性时空分布特征的认识.但由于细菌本身的特点, 例如频繁的基因水平转移, 使得细菌的基因组存在着高度的动态变化, 导致对细菌“种”的定义存在着巨大困难.目前, 微生物学家对如何定义细菌“种”仍存在争议, 例如: 是否需要同时考虑细菌“种”在表型和基因型上的相似性, 是否需要同时考虑细菌“种”在基因、系统发育史、进化以及生态特征上的相似性(Gevers et al ., 2005 ; Logue et al ., 2008 ).目前, 对细菌“种”的常用定义为: 基因组具有一致性的一组分离菌株, 若在相同条件下对这些菌株进行检测, 它们在多项指标上具有高度的相似性(Rosselló-Mora & Amann, 2001 ). ...
Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of Actinobacteria.
2000
Ecological consequences of bacterioplankton lifestyles: changes in concepts are needed.
2
2010
... 浮游细菌的生产力也可以细分为初级生产力和次级生产力.初级生产力是由自养细菌来完成的, 自养细菌中普遍分布的、发挥主要功能的一类是蓝细菌, 在一些研究中它们占据自养浮游细菌生产力的95%.而自养细菌占整个浮游细菌生产力的比例也同样有着季节的变化, 在7月的午后达到最大, 约为55.6%, 而在1月只有2.3%(Affronti & Marshall, 1994 ).浮游细菌的次级生产力主要表现在有机物质(OM)的降解利用上, 由异养型浮游细菌来完成, 它们是湖泊浮游细菌的主要组成部分, 主要扮演着分解者的角色.对浮游细菌分解过程的研究表明, 细菌可以产生胞外酶来分解颗粒性有机物(POM)和溶解性有机物(DOM); 对于比较复杂的腐植酸, 可能需要多种细菌的多种代谢途径来共同完成.颗粒类有机物和复杂有机物会被分解成有机物“微羽(plume)”, 这些“微羽”又能够通过信号物质吸引浮游细菌, 并进一步被分解成小分子有机物.对于有机物“微羽”的分解在水柱细菌生产力中占有很大的比例(Grossart, 2010 ). ...
... 将细菌群落结构和特定生态功能联系起来, 一直是众多微生物生态学家关心的热点.BIOLOGTM Plates、稳定性同位素、高级FISH技术、微阵列杂交、功能基因的扩增测序和定量研究、蛋白质组测序技术, 都大大促进了人们对湖泊水体细菌各种生态功能的认识.尽管有种种研究表明不同的物质循环过程会导致非常迥异的细菌群落结构(Grossart, 2010 ), 但还是难以把某一类细菌和某一特定生态功能直接联系起来.这是因为即使同一“种”的细菌, 在不同的环境下也常常会显示出截然不同的生理生态特性(Jaspers & Overmann, 2004 ).这些基本功能, 在不同门类的细菌中应该是普遍存在的, 只是典型种类的细菌已经被人们分离纯培养并为人们所熟知.分子生物学、显微成像、连续自动采样等技术的进一步发展, 将使得人们可以在更小的时空范围内比较亲缘关系上接近或遥远的细菌之间蛋白质组的差异, 追踪细菌的形态和生理特征方面的变化(Logue et al ., 2008 ), 从而可以为人们认识湖泊水体细菌的相关功能提供更有利的支持. ...
Isolation of strains belonging to the cosmopolitan Polynucleobacter necessarius cluster from freshwater habitats located in three climatic zones.
1
2003
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
Grazing of protozoa and its effect on populations of aquatic bacteria.
1
2001
... 原生动物与细菌间的捕食和被捕食关系构成了湖泊微生物网的基础.在捕食细菌的原生动物中, 异养鞭毛虫(HNFs)被认为是影响细菌群落结构和组成的最主要的因子之一(Šimek & Chrzanowski, 1992 ; Young, 2006 ).研究表明HNFs对细菌的捕食作用同样显著改变了细菌群落的形态分布特征, 在高强度的捕食压力下, 中等大小的细菌更容易被捕食, 使得细菌群落形态呈极小或丝状分布(Hahn & Höfle, 2001 ).除大小特征外, Wu等(2004)认为极小或丝状细菌难以被异养鞭毛虫消化也是导致其不被捕食的重要原因.尽管研究者们认识到水体病毒对细菌群落结构的形成存在重要影响(Weinbauer et al ., 2006 ), 但目前, 对病毒如何引起细菌的死亡及影响细菌组成间的竞争的研究仍非常之少. ...
Polynucleobacter rarus sp. nov., a free-living planktonic bacterium isolated from an acidic lake.
1
2011
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
Water column anammox and denitrification in a temperate permanently stratified lake (Lake Rassnitzer, Germany).
1
2009
... (4)在湖泊水体中还可能存在着厌氧氨氧化细菌, 这类细菌可以在厌氧条件下利用铵离子和亚硝酸根产生氮气.有研究表明在深水湖泊中等温层以下的水体中厌氧氨氧化(最大504 nmol N·L-1 ·d-1 )和反硝化(最大74 nmol N·L-1 ·d-1 )对氮气产出有着非常大的贡献(Hamersley et al ., 2009 ). ...
Metapopulation dynamics: brief history and conceptual domain.
1991
Responses of plankton, turbidity, and macrophytes to biomanipulation in a shallow prairie lake.
1
1994
... 湖泊生态系统, 特别是浅水湖泊生态系统, 一般按初级生产者的组成分成两种类型: 一种是当营养负荷较低时, 以大型沉水植物为主要初级生产者的清水湖泊; 另一种是当营养负荷较高时, 以浮游植物(藻类)为主要初级生产者的浊水湖泊.湖泊从清水草型向浊水藻型转变时(湖泊富营养化), 生物多样性下降(Hanson & Butler, 1994 ; Riemann & Winding, 2001 ; Villaescusa et al ., 2010 ).Hanson和Butler(1994)认为湖泊从清水状态转变为浊水状态后, 虽然没有改变湖泊生态系统中的经典食物链的传递(藻类→浮游动物→鱼类), 但是却导致组成微生物食物网的所有生物的生物量显著增加(Mathes & Arndt, 1994 ). ...
Stratification of microbial assemblages in Mono Lake, California, and response to a mixing event.
2001
Metacommunities: a framework for large-scale community ecology.
1
2005
... 与Beijerinck和Baas-Becking所提出的细菌全球性分布的观点相对应的是meta-群落假说(Leibold et al ., 2004 ; Brendan Logue & Lindström, 2008 ).Hanski 和Gilpin(1991)对meta-群落的定义是: 因扩散作用而联系在一起的一系列本地群落的集合.与之前的群落理论只强调群落受当地当时的环境条件作用的观点不同的是, meta-群落假说更强调不同时空尺度(本地、区域)上群落的结构和动态过程.且meta-群落假说认为不同本地群落在时空尺度上的动态变化, 不仅可以改变本地群落结构, 而且可通过反馈作用改变区域尺度上生物群落的组成和结构(Holyoak et al ., 2005 ).Meta-群落假说由4个简单的观点组成, 它们都在一定程度上解释了细菌群落的生物地理学分布及动态规律(图1 ). ...
1
2001
... (4)中性理论(neutral perspective, 图1D ), 该理论假定所有细菌种类具有相似的竞争能力、扩散能力和适应性(Hubbell, 2001 ).不同类群细菌间的相互作用由细菌的随机扩散作用组成, 因此, 物种的减少(灭绝和迁出)和增加(迁入和物种的形成)的随机概率决定了物种的多样性.中性理论认为细菌群落组成动态变化受地理距离远近的驱动, 而不受环境条件的影响. ...
Depth distribution of microbial diversity in Mono Lake, a meromictic soda lake in California.
2003
Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing.
1
2008
... 尽管微生物研究者已认识到仅用16S rRNA基因序列的相似性来定义细菌的分类单元存在着很大弊端, 但由于研究条件和手段的限制, 目前关于微生物多样性的大部分研究, 仍然依赖于16S rRNA基因序列的相似性来解释微生物的多样性.目前, 有两种解读方法被广泛使用: 一是计算未知序列与参考序列库中序列的相似性, 将未知序列分成不同的分类单元, 例如: 种系型(phylotypes; Huse et al ., 2008 ); 二是通过比较序列与群落中其他未知序列间的相似性, 将序列分成不同的操作性分类单元(OTUs)(Schloss, 2009 ; Schloss & Westcott, 2011 ).目前第二代测序技术(高通量测序)广泛使用, 越来越多先前难以检测到的稀有细菌种类被检测出来.如何将难以培养的稀有细菌准确分类是研究者们将要面临的严峻挑战. ...
Ecological significance of microdiversity: identical 16S rRNA gene sequences can be found in bacteria with highly divergent genomes and ecophysiologies.
1
2004
... 将细菌群落结构和特定生态功能联系起来, 一直是众多微生物生态学家关心的热点.BIOLOGTM Plates、稳定性同位素、高级FISH技术、微阵列杂交、功能基因的扩增测序和定量研究、蛋白质组测序技术, 都大大促进了人们对湖泊水体细菌各种生态功能的认识.尽管有种种研究表明不同的物质循环过程会导致非常迥异的细菌群落结构(Grossart, 2010 ), 但还是难以把某一类细菌和某一特定生态功能直接联系起来.这是因为即使同一“种”的细菌, 在不同的环境下也常常会显示出截然不同的生理生态特性(Jaspers & Overmann, 2004 ).这些基本功能, 在不同门类的细菌中应该是普遍存在的, 只是典型种类的细菌已经被人们分离纯培养并为人们所熟知.分子生物学、显微成像、连续自动采样等技术的进一步发展, 将使得人们可以在更小的时空范围内比较亲缘关系上接近或遥远的细菌之间蛋白质组的差异, 追踪细菌的形态和生理特征方面的变化(Logue et al ., 2008 ), 从而可以为人们认识湖泊水体细菌的相关功能提供更有利的支持. ...
Ubiquity of Polynucleobacter necessarius ssp. asymbioticus in lentic freshwater habitats of a heterogenous 2000 km2 area.
1
2010
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
Inventory, differentiation, and proportional diversity: a consistent terminology for quantifying species diversity.
2009
The potential importance of grazing-resistant bacteria in planktonic systems.
1
1994
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
Nitrogen fixation by bacteria in Lake Mize, Florida, and in some lacustrine sediments.
1
1971
... (1)湖泊中的固氮细菌发现的主要是蓝细菌的某些种类, 如念珠藻、水华束丝藻等.这些种类的蓝藻在夏季表层水体无机氮被消耗匮乏时有着很强的生存优势(Torrey & Lee, 1975 ).不过也有学者认为蓝细菌的固氮作用并不显著, 因其可直接利用来自于沉积物中的铵离子和硝酸根离子生存, 并不完全依赖于固氮作用(Ferber et al ., 2004 ).另外一些梭菌、硫螺菌属的细菌在5-10 m深的水柱可以通过异养代谢来进行固氮作用(Keirn & Brezonik, 1971 ). ...
Synchrony in aquatic microbial community dynamics.
1
2007
... 如果以湖岸线为界限, 将湖泊看作一个整体, 我们可以把影响湖泊细菌多样性的环境因子分为湖泊生态系统的内部环境因子和区域尺度上的外部环境因子.区域尺度上的非生物因素, 例如温度和主要离子浓度, 在不同湖泊间表现出年际变化同步性(Magnuson et al ., 1990 ), 而湖泊特有的内部环境因子, 例如食物网中生物间相互作用及物种的随机动态, 却往往抑制并减弱了区域尺度上的外界因子对细菌多样性的作用(Magnuson et al ., 2006 ; Kent et al ., 2007 ). ...
Structure and function of bacterial communities emerging from different sources under identical conditions.
1
2006
... pH是决定湖泊细菌群落结构的另一关键因子, 野外原位试验研究表明, 湖泊水体细菌群落结构和多样性随着水体pH变化而发生显著变化(如: Lindström et al ., 2005 ; Yannarell & Triplett, 2005 ).pH可以通过影响不同种类细菌的生长状况(包括: 绝灭、繁衍、种的形成等; Langenheder et al ., 2006 ), 直接影响多样性; 也可通过影响湖泊生态系统中的其他环境因子(如: 有机物质的分子结构; Yannarell & Triplett, 2005 )来间接影响水体细菌群落的结构和多样性. ...
Are there general laws in ecology?
1
1999
... 物种的多样性与生境大小的关系一直是生态学关注的热点之一(Rosenzweig, 1995 ; Lawton, 1999 ).MacArthur和Wilson(1967)针对生物多样性与生境大小之间的关系提出“生物地理学的岛屿效应”, 它主要包括两个方面: (1)岛屿上物种数目会随着岛屿面积的增加而增加; (2)岛屿上物种数目会随着岛屿与大陆间的距离增加而下降.如果把陆地看作“海洋”, 湖泊就是陆地上的“岛屿”.那么, 湖泊中的细菌是否也如大型生物一样, 呈现出“岛屿效应”? Reche等(2005)研究位于西班牙内华达山脉上的11个高原湖泊中的细菌群落结构和多样性, 发现细菌物种数和湖泊的面积呈显著的正相关关系.但湖泊的偏僻程度并不影响细菌物种数目, 这在一定程度上表明细菌分布并不存在“地理隔离”. ...
The metacommunity concept: a framework for multi-scale community ecology.
2
2004
... 与Beijerinck和Baas-Becking所提出的细菌全球性分布的观点相对应的是meta-群落假说(Leibold et al ., 2004 ; Brendan Logue & Lindström, 2008 ).Hanski 和Gilpin(1991)对meta-群落的定义是: 因扩散作用而联系在一起的一系列本地群落的集合.与之前的群落理论只强调群落受当地当时的环境条件作用的观点不同的是, meta-群落假说更强调不同时空尺度(本地、区域)上群落的结构和动态过程.且meta-群落假说认为不同本地群落在时空尺度上的动态变化, 不仅可以改变本地群落结构, 而且可通过反馈作用改变区域尺度上生物群落的组成和结构(Holyoak et al ., 2005 ).Meta-群落假说由4个简单的观点组成, 它们都在一定程度上解释了细菌群落的生物地理学分布及动态规律(图1 ). ...
... 从meta-群落假说以及Beijerinck和Baas- Becking的观点(Loreau et al ., 2003 ; Leibold et al ., 2004 ; Martiny et al ., 2006 )出发, 可以推测出淡水细菌在全球、区域和本土3个梯度下的物种分布特征和群落多样性, 更可能是由“随机分布”和“环境决定”共同作用的结果.与Forbes(1887)提出的“湖泊正如一个微宇宙”的观点不同, 现在研究者们更多将湖泊看作一个流动系统, 认为它是更大尺度空间中的一个组成单元.湖泊不是一个与外界环境隔离的生态系统, 它与外界环境之间存在着频繁的物质、能量及基因的交流.对于水力停留时间很长的湖泊, 其细菌群落组成主要受湖泊内部环境因子的影响, 即适用物种筛选(species-sorting perspective)和Baas- Becking的观点(Brendan Logue & Lindström, 2008 ).但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 ). ...
External control of bacterial community structure in lakes.
2006
Distribution of typical freshwater bacterial groups is associated with pH, temperature, and lake water retention time.
1
2005
... pH是决定湖泊细菌群落结构的另一关键因子, 野外原位试验研究表明, 湖泊水体细菌群落结构和多样性随着水体pH变化而发生显著变化(如: Lindström et al ., 2005 ; Yannarell & Triplett, 2005 ).pH可以通过影响不同种类细菌的生长状况(包括: 绝灭、繁衍、种的形成等; Langenheder et al ., 2006 ), 直接影响多样性; 也可通过影响湖泊生态系统中的其他环境因子(如: 有机物质的分子结构; Yannarell & Triplett, 2005 )来间接影响水体细菌群落的结构和多样性. ...
Infrequent marine-freshwater transitions in the microbial world.
2
2009
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
Progress in the ecological genetics and biodiversity of freshwater bacteria.
4
2008
... 对细菌“种”的科学且合理的定义, 影响着微生物学者对微生物多样性时空分布特征的认识.但由于细菌本身的特点, 例如频繁的基因水平转移, 使得细菌的基因组存在着高度的动态变化, 导致对细菌“种”的定义存在着巨大困难.目前, 微生物学家对如何定义细菌“种”仍存在争议, 例如: 是否需要同时考虑细菌“种”在表型和基因型上的相似性, 是否需要同时考虑细菌“种”在基因、系统发育史、进化以及生态特征上的相似性(Gevers et al ., 2005 ; Logue et al ., 2008 ).目前, 对细菌“种”的常用定义为: 基因组具有一致性的一组分离菌株, 若在相同条件下对这些菌株进行检测, 它们在多项指标上具有高度的相似性(Rosselló-Mora & Amann, 2001 ). ...
... meta-群落假说的4种观点对细菌群落的相似性与环境和地理距离之间的相互关系的解释(改自Logue et al ., 2008 ) ...
... The interpretation of the relationship between bacterial community similarity and environmental gradients and geographic distances under four perspectives of meta- community hypothesis (revised from Logue et al ., 2008 ) ...
... 将细菌群落结构和特定生态功能联系起来, 一直是众多微生物生态学家关心的热点.BIOLOGTM Plates、稳定性同位素、高级FISH技术、微阵列杂交、功能基因的扩增测序和定量研究、蛋白质组测序技术, 都大大促进了人们对湖泊水体细菌各种生态功能的认识.尽管有种种研究表明不同的物质循环过程会导致非常迥异的细菌群落结构(Grossart, 2010 ), 但还是难以把某一类细菌和某一特定生态功能直接联系起来.这是因为即使同一“种”的细菌, 在不同的环境下也常常会显示出截然不同的生理生态特性(Jaspers & Overmann, 2004 ).这些基本功能, 在不同门类的细菌中应该是普遍存在的, 只是典型种类的细菌已经被人们分离纯培养并为人们所熟知.分子生物学、显微成像、连续自动采样等技术的进一步发展, 将使得人们可以在更小的时空范围内比较亲缘关系上接近或遥远的细菌之间蛋白质组的差异, 追踪细菌的形态和生理特征方面的变化(Logue et al ., 2008 ), 从而可以为人们认识湖泊水体细菌的相关功能提供更有利的支持. ...
Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology.
1
2003
... 从meta-群落假说以及Beijerinck和Baas- Becking的观点(Loreau et al ., 2003 ; Leibold et al ., 2004 ; Martiny et al ., 2006 )出发, 可以推测出淡水细菌在全球、区域和本土3个梯度下的物种分布特征和群落多样性, 更可能是由“随机分布”和“环境决定”共同作用的结果.与Forbes(1887)提出的“湖泊正如一个微宇宙”的观点不同, 现在研究者们更多将湖泊看作一个流动系统, 认为它是更大尺度空间中的一个组成单元.湖泊不是一个与外界环境隔离的生态系统, 它与外界环境之间存在着频繁的物质、能量及基因的交流.对于水力停留时间很长的湖泊, 其细菌群落组成主要受湖泊内部环境因子的影响, 即适用物种筛选(species-sorting perspective)和Baas- Becking的观点(Brendan Logue & Lindström, 2008 ).但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 ). ...
Temporal coherence in the limnology of a suite of lakes in Wisconsin, USA.
1
1990
... 如果以湖岸线为界限, 将湖泊看作一个整体, 我们可以把影响湖泊细菌多样性的环境因子分为湖泊生态系统的内部环境因子和区域尺度上的外部环境因子.区域尺度上的非生物因素, 例如温度和主要离子浓度, 在不同湖泊间表现出年际变化同步性(Magnuson et al ., 1990 ), 而湖泊特有的内部环境因子, 例如食物网中生物间相互作用及物种的随机动态, 却往往抑制并减弱了区域尺度上的外界因子对细菌多样性的作用(Magnuson et al ., 2006 ; Kent et al ., 2007 ). ...
1
2006
... 如果以湖岸线为界限, 将湖泊看作一个整体, 我们可以把影响湖泊细菌多样性的环境因子分为湖泊生态系统的内部环境因子和区域尺度上的外部环境因子.区域尺度上的非生物因素, 例如温度和主要离子浓度, 在不同湖泊间表现出年际变化同步性(Magnuson et al ., 1990 ), 而湖泊特有的内部环境因子, 例如食物网中生物间相互作用及物种的随机动态, 却往往抑制并减弱了区域尺度上的外界因子对细菌多样性的作用(Magnuson et al ., 2006 ; Kent et al ., 2007 ). ...
Microbial biogeography: putting microorganisms on the map.
2
2006
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
... 从meta-群落假说以及Beijerinck和Baas- Becking的观点(Loreau et al ., 2003 ; Leibold et al ., 2004 ; Martiny et al ., 2006 )出发, 可以推测出淡水细菌在全球、区域和本土3个梯度下的物种分布特征和群落多样性, 更可能是由“随机分布”和“环境决定”共同作用的结果.与Forbes(1887)提出的“湖泊正如一个微宇宙”的观点不同, 现在研究者们更多将湖泊看作一个流动系统, 认为它是更大尺度空间中的一个组成单元.湖泊不是一个与外界环境隔离的生态系统, 它与外界环境之间存在着频繁的物质、能量及基因的交流.对于水力停留时间很长的湖泊, 其细菌群落组成主要受湖泊内部环境因子的影响, 即适用物种筛选(species-sorting perspective)和Baas- Becking的观点(Brendan Logue & Lindström, 2008 ).但对于水力停留时间较短的湖泊, 由于与外界环境之间存在着频繁的物质、能量及基因的交流, 其细菌群落结构和多样性受湖泊内部环境特征和细菌在不同生境(或斑块)间随机分布作用的影响, 即更适用meta-群落假说(Brendan Logue & Lindström, 2008 ). ...
Biomass and composition of protozooplankton in relation to lake trophy in north German lakes.
1
1994
... 湖泊生态系统, 特别是浅水湖泊生态系统, 一般按初级生产者的组成分成两种类型: 一种是当营养负荷较低时, 以大型沉水植物为主要初级生产者的清水湖泊; 另一种是当营养负荷较高时, 以浮游植物(藻类)为主要初级生产者的浊水湖泊.湖泊从清水草型向浊水藻型转变时(湖泊富营养化), 生物多样性下降(Hanson & Butler, 1994 ; Riemann & Winding, 2001 ; Villaescusa et al ., 2010 ).Hanson和Butler(1994)认为湖泊从清水状态转变为浊水状态后, 虽然没有改变湖泊生态系统中的经典食物链的传递(藻类→浮游动物→鱼类), 但是却导致组成微生物食物网的所有生物的生物量显著增加(Mathes & Arndt, 1994 ). ...
Trophic roles of heterotrophic nanoflagellates and ciliates among planktonic organisms in a hypereutrophic pond.
1
1998
... 按照营养盐的化学组成形式, 我们可以将水体营养盐分成无机营养盐和有机营养盐.它们既可通过“上行效应”直接影响水体细菌群落的组成和分布(Crump et al ., 2003 ; Pérez & Sommaruga, 2006 ; Bouvy et al ., 2011 ); 也可通过影响水体细菌的捕食者(异样鞭毛虫和纤毛虫)的组成和丰度(Nakano et al ., 1998 ), 间接影响水体细菌群落结构和多样性. ...
A guide to the natural history of freshwater lake bacteria.
2011
Differential effect of algal- and soil-derived dissolved organic matter on alpine lake bacterial community composition and activity.
1
2006
... 按照营养盐的化学组成形式, 我们可以将水体营养盐分成无机营养盐和有机营养盐.它们既可通过“上行效应”直接影响水体细菌群落的组成和分布(Crump et al ., 2003 ; Pérez & Sommaruga, 2006 ; Bouvy et al ., 2011 ); 也可通过影响水体细菌的捕食者(异样鞭毛虫和纤毛虫)的组成和丰度(Nakano et al ., 1998 ), 间接影响水体细菌群落结构和多样性. ...
Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria.
1
2001
... 温度是反映不同纬度、海拔及季节梯度上湖泊环境差异的主要因子.在全球变暖背景下, 由增温所引起的一系列湖泊生态效应也是当前和未来科学界关注的焦点之一.越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一(Simon & Wünsch, 1998 ; Adams et al ., 2010 ; Dziallas & Grossart, 2011 ).Adams等(2010)通过24个湖泊和溪流样品的野外研究和室内培养试验, 发现在温度因子的驱动下细菌群落的组成发生了显著的改变.不同的细菌种类有着不同的生态最适温度, 在最适温度下, 细菌表现出最大的适合度.因此温度可能对水体细菌群落组成具有选择作用.Wu和Hahn (2006b)对湖泊水体模式细菌Polynucleobacte 的研究进一步表明温度对水体细菌组成在属的水平上的选择作用, 温度差异促使了细菌在同一属内的分化, Polynucleobacte 属细菌在不同温度的湖泊间表现出明显的生态多样化.温度还可以作为湖泊水体细菌在季节水平上的动态变化最好的预测因子.Wu和Hahn(2006a)通过荧光原位杂交的方法研究了3年内月亮湖中模式浮游细菌Polynucleobacter 亚群(PnecB)的丰度变化, 发现PnecB细菌丰度在研究的3年内表现出相似的季节动态特征.因细菌的生物过程(包括繁殖速率、扩散作用、物种间的相互作用、基因的突变、适应进化及物种的形成)受到温度的调控(Pomeroy & Wiebe, 2001 ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加.他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”.细菌个体的大小分布特征同样受温度的影响.Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低. ...
Role of predatory bacteria in the termination of a cyanobacterial bloom.
1
2001
... 研究表明, 大型沉水植物的生物量和组成是影响太湖中水体细菌群落结构和多样性的关键因子, 同时也是调控太湖水体细菌群落组成季节动态的主要驱动因子(Wu et al ., 2007 ; Zeng et al ., 2012 ).当湖泊水体发生富营养化, 蓝藻爆发有助于具有溶菌作用的病毒和噬细胞菌属(Cytophaga )细菌的生长, 同样显著影响着水体细菌群落结构和多样性(Rashidan & Bird, 2001 ). ...
Does ecosystem size determine aquatic bacterial richness?
2005
1
1980
... 20世纪80年代末, 基于纯培养的研究表明, 土壤、海洋和淡水等生态系统中的细菌群落结构和组成相似(Rheinheimer, 1980 ), 但分子生态学技术的出现与不断发展已逐渐打破了这种认识.Glöckner等(2000)、Zwart等(2002)和Logares等(2009)运用现代分子生态学方法比较了不同生境中的细菌多样性, 发现淡水生境的细菌组成与土壤及海洋生态系统中的细菌组成有着明显差异.Zwart等(2002)从不同淡水环境(湖泊和河流)得到的689个序列中至少有34个16S rRNA序列为典型本土淡水细菌类群, 而且它们的分布非常广泛.这些典型的淡水细菌大都隶属于α-、β-、γ-变形菌亚门(Proteobacteria)、蓝细菌门(Cyanobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)和疣微菌门(Verrucom- icrobia).Newton等(2011)在Zwart等(2002, 2003)研究的基础上, 结合淡水生态系统中细菌多样性的最新研究成果, 发现湖泊生态系统变温层中共有21个典型的淡水细菌门类(ribosomal database project, RDP, 相似性为75%).除Zwart等(2002)已报道的5个优势门类, 以放线菌和β-变形菌数量最为丰富; 其他的16个细菌门类仅约占所收集到的全部序列的2.6%, 包括: 酸杆菌门(Acidobacteria), 绿菌门(Chlorobi), 绿弯菌门(Chloroflexi), 纤维杆菌门(Fibrobacteres), 厚壁菌门(Firmicutes), 梭杆菌门(Fusobacteria), 芽单胞菌门(Gemmatimonadetes), 黏胶球形菌门(Lentisphaerae), 硝化螺菌门(Nitrospira), 浮霉菌门(Planctomycetes), 螺旋体门(Spirochaetes), BRC1, OD1, OP10, SR1和TM7. ...
Community dynamics of free-living and particle-associated bacterial assemblages during a freshwater phytoplankton bloom.
1
2001
... 湖泊生态系统, 特别是浅水湖泊生态系统, 一般按初级生产者的组成分成两种类型: 一种是当营养负荷较低时, 以大型沉水植物为主要初级生产者的清水湖泊; 另一种是当营养负荷较高时, 以浮游植物(藻类)为主要初级生产者的浊水湖泊.湖泊从清水草型向浊水藻型转变时(湖泊富营养化), 生物多样性下降(Hanson & Butler, 1994 ; Riemann & Winding, 2001 ; Villaescusa et al ., 2010 ).Hanson和Butler(1994)认为湖泊从清水状态转变为浊水状态后, 虽然没有改变湖泊生态系统中的经典食物链的传递(藻类→浮游动物→鱼类), 但是却导致组成微生物食物网的所有生物的生物量显著增加(Mathes & Arndt, 1994 ). ...
Ecology of purple sulfur bacteria in the highly stratified meromictic Lake Shunet (Siberia, Khakassia) in 2002-2009.
1
2012
... 湖泊水体中与硫循环有关的细菌主要包括硫氧化细菌和硫酸盐还原细菌.其中紫硫细菌是一类化能自养不产氧细菌, 它们以硫化物为电子受体, 在一些盐碱性湖泊中是主要的初级生产者, 在分层湖泊的化学跃变层以下光线可达的水体中, 在小范围内能够积累很大的生物量, 并在湖泊中形成肉眼可见的紫色水层(Rogozin et al ., 2012 ).在化学跃变层的厌氧性水体以及蓝藻暴发后形成的厌氧环境下, 都能够检测到较高丰度的硫酸盐还原细菌(每毫升水体104 -106 个细胞).湖泊水体细菌进行的硫氧化还原过程中常常会利用重金属作为中间电子供体或受体, 从而也被认为在重金属甲基化等过程中发挥重要作用(Teske et al ., 1998 ; Chirwa & Wang, 2000 ; Sorokin et al ., 2011 ). ...
1
1995
... 物种的多样性与生境大小的关系一直是生态学关注的热点之一(Rosenzweig, 1995 ; Lawton, 1999 ).MacArthur和Wilson(1967)针对生物多样性与生境大小之间的关系提出“生物地理学的岛屿效应”, 它主要包括两个方面: (1)岛屿上物种数目会随着岛屿面积的增加而增加; (2)岛屿上物种数目会随着岛屿与大陆间的距离增加而下降.如果把陆地看作“海洋”, 湖泊就是陆地上的“岛屿”.那么, 湖泊中的细菌是否也如大型生物一样, 呈现出“岛屿效应”? Reche等(2005)研究位于西班牙内华达山脉上的11个高原湖泊中的细菌群落结构和多样性, 发现细菌物种数和湖泊的面积呈显著的正相关关系.但湖泊的偏僻程度并不影响细菌物种数目, 这在一定程度上表明细菌分布并不存在“地理隔离”. ...
Detection and quantification with 16S rRNA probes of planktonic methylotrophic bacteria in a floodplain lake.
1
1997
... 碳和磷是构成细菌的最基本元素.在湖泊生态系统中几乎所有细菌均参与碳、磷元素的生物地球化学过程.但是目前相关研究匮乏.在湖泊水体中, 与C循环有关的一类细菌为甲基营养细菌, 它是一类以单甲基分子为营养的自养细菌, 并最终将甲基分子氧化为CO2 .甲基营养细菌在淡水水体中广泛分布, 在一些湖泊中能够达到每升水0.6-5.5×109 个细胞, 约占浮游细菌总数的10-46%(Ross et al ., 1997 ).另一类与C循环有关的是产甲烷微生物, 主要存在于湖泊沉积物等厌氧环境下, 且以产甲烷古菌为主, 本文不再详述.在湖泊生态系统中多种形态的水体浮游细菌都可以在胞内或胞外分泌磷酸酶, 进而利用各种复杂形态的有机磷, 形成不同类型的适应状态和生态位分化(Spijkerman & Coesel, 1998 ). ...
The species concept for prokaryotes.
1
2001
... 对细菌“种”的科学且合理的定义, 影响着微生物学者对微生物多样性时空分布特征的认识.但由于细菌本身的特点, 例如频繁的基因水平转移, 使得细菌的基因组存在着高度的动态变化, 导致对细菌“种”的定义存在着巨大困难.目前, 微生物学家对如何定义细菌“种”仍存在争议, 例如: 是否需要同时考虑细菌“种”在表型和基因型上的相似性, 是否需要同时考虑细菌“种”在基因、系统发育史、进化以及生态特征上的相似性(Gevers et al ., 2005 ; Logue et al ., 2008 ).目前, 对细菌“种”的常用定义为: 基因组具有一致性的一组分离菌株, 若在相同条件下对这些菌株进行检测, 它们在多项指标上具有高度的相似性(Rosselló-Mora & Amann, 2001 ). ...
A high-throughput DNA sequence aligner for microbial ecology studies.
1
2009
... 尽管微生物研究者已认识到仅用16S rRNA基因序列的相似性来定义细菌的分类单元存在着很大弊端, 但由于研究条件和手段的限制, 目前关于微生物多样性的大部分研究, 仍然依赖于16S rRNA基因序列的相似性来解释微生物的多样性.目前, 有两种解读方法被广泛使用: 一是计算未知序列与参考序列库中序列的相似性, 将未知序列分成不同的分类单元, 例如: 种系型(phylotypes; Huse et al ., 2008 ); 二是通过比较序列与群落中其他未知序列间的相似性, 将序列分成不同的操作性分类单元(OTUs)(Schloss, 2009 ; Schloss & Westcott, 2011 ).目前第二代测序技术(高通量测序)广泛使用, 越来越多先前难以检测到的稀有细菌种类被检测出来.如何将难以培养的稀有细菌准确分类是研究者们将要面临的严峻挑战. ...
Assessing and improving methods used in operational taxonomic unit-based approaches for 16S rRNA gene sequence analysis.
1
2011
... 尽管微生物研究者已认识到仅用16S rRNA基因序列的相似性来定义细菌的分类单元存在着很大弊端, 但由于研究条件和手段的限制, 目前关于微生物多样性的大部分研究, 仍然依赖于16S rRNA基因序列的相似性来解释微生物的多样性.目前, 有两种解读方法被广泛使用: 一是计算未知序列与参考序列库中序列的相似性, 将未知序列分成不同的分类单元, 例如: 种系型(phylotypes; Huse et al ., 2008 ); 二是通过比较序列与群落中其他未知序列间的相似性, 将序列分成不同的操作性分类单元(OTUs)(Schloss, 2009 ; Schloss & Westcott, 2011 ).目前第二代测序技术(高通量测序)广泛使用, 越来越多先前难以检测到的稀有细菌种类被检测出来.如何将难以培养的稀有细菌准确分类是研究者们将要面临的严峻挑战. ...
Functional characteristics of bacterioplankton with reference to its aggregation in water bodies of different types.
1
2002
... 另外值得注意的是, 水体细菌可以按生活状态分为自由浮动和聚集体(>5 μm)两种形式, 研究表明聚集体形式的细菌在生产力和能量系数上要高于自由浮动的细菌(Shchur et al ., 2002 ). ...
Direct and indirect evidence of size-selective grazing on pelagic bacteria by freshwater nanoflagellates.
1
1992
... 原生动物与细菌间的捕食和被捕食关系构成了湖泊微生物网的基础.在捕食细菌的原生动物中, 异养鞭毛虫(HNFs)被认为是影响细菌群落结构和组成的最主要的因子之一(Šimek & Chrzanowski, 1992 ; Young, 2006 ).研究表明HNFs对细菌的捕食作用同样显著改变了细菌群落的形态分布特征, 在高强度的捕食压力下, 中等大小的细菌更容易被捕食, 使得细菌群落形态呈极小或丝状分布(Hahn & Höfle, 2001 ).除大小特征外, Wu等(2004)认为极小或丝状细菌难以被异养鞭毛虫消化也是导致其不被捕食的重要原因.尽管研究者们认识到水体病毒对细菌群落结构的形成存在重要影响(Weinbauer et al ., 2006 ), 但目前, 对病毒如何引起细菌的死亡及影响细菌组成间的竞争的研究仍非常之少. ...
Temperature control of bacterioplankton growth in a temperate large lake.
1
1998
... 温度是反映不同纬度、海拔及季节梯度上湖泊环境差异的主要因子.在全球变暖背景下, 由增温所引起的一系列湖泊生态效应也是当前和未来科学界关注的焦点之一.越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一(Simon & Wünsch, 1998 ; Adams et al ., 2010 ; Dziallas & Grossart, 2011 ).Adams等(2010)通过24个湖泊和溪流样品的野外研究和室内培养试验, 发现在温度因子的驱动下细菌群落的组成发生了显著的改变.不同的细菌种类有着不同的生态最适温度, 在最适温度下, 细菌表现出最大的适合度.因此温度可能对水体细菌群落组成具有选择作用.Wu和Hahn (2006b)对湖泊水体模式细菌Polynucleobacte 的研究进一步表明温度对水体细菌组成在属的水平上的选择作用, 温度差异促使了细菌在同一属内的分化, Polynucleobacte 属细菌在不同温度的湖泊间表现出明显的生态多样化.温度还可以作为湖泊水体细菌在季节水平上的动态变化最好的预测因子.Wu和Hahn(2006a)通过荧光原位杂交的方法研究了3年内月亮湖中模式浮游细菌Polynucleobacter 亚群(PnecB)的丰度变化, 发现PnecB细菌丰度在研究的3年内表现出相似的季节动态特征.因细菌的生物过程(包括繁殖速率、扩散作用、物种间的相互作用、基因的突变、适应进化及物种的形成)受到温度的调控(Pomeroy & Wiebe, 2001 ; Adams et al ., 2010 ), 且温度升高细菌的生物过程加剧, 因此 Fuhrman等(2008)认为细菌的多样性受温度的影响, 温度升高细菌多样性增加.他把这一现象称为红色皇后假设, 即“红色皇后(Red Queen)为保持其在环境中的原位, 在环境变化越快时跑得越快”.细菌个体的大小分布特征同样受温度的影响.Daufresne等(2009)在模拟研究全球变化(温度分别增加2, 4和6°C)对水体生物影响的实验中发现水体浮游细菌群落的平均细胞大小随着温度增加而降低. ...
The microbial sulfur cycle at extremely haloalkaline conditions of soda lakes.
1
2011
... 湖泊水体中与硫循环有关的细菌主要包括硫氧化细菌和硫酸盐还原细菌.其中紫硫细菌是一类化能自养不产氧细菌, 它们以硫化物为电子受体, 在一些盐碱性湖泊中是主要的初级生产者, 在分层湖泊的化学跃变层以下光线可达的水体中, 在小范围内能够积累很大的生物量, 并在湖泊中形成肉眼可见的紫色水层(Rogozin et al ., 2012 ).在化学跃变层的厌氧性水体以及蓝藻暴发后形成的厌氧环境下, 都能够检测到较高丰度的硫酸盐还原细菌(每毫升水体104 -106 个细胞).湖泊水体细菌进行的硫氧化还原过程中常常会利用重金属作为中间电子供体或受体, 从而也被认为在重金属甲基化等过程中发挥重要作用(Teske et al ., 1998 ; Chirwa & Wang, 2000 ; Sorokin et al ., 2011 ). ...
Alkaline phosphatase activity in two planktonic desmid species and the possible role of an extracellular envelope.
1
1998
... 碳和磷是构成细菌的最基本元素.在湖泊生态系统中几乎所有细菌均参与碳、磷元素的生物地球化学过程.但是目前相关研究匮乏.在湖泊水体中, 与C循环有关的一类细菌为甲基营养细菌, 它是一类以单甲基分子为营养的自养细菌, 并最终将甲基分子氧化为CO2 .甲基营养细菌在淡水水体中广泛分布, 在一些湖泊中能够达到每升水0.6-5.5×109 个细胞, 约占浮游细菌总数的10-46%(Ross et al ., 1997 ).另一类与C循环有关的是产甲烷微生物, 主要存在于湖泊沉积物等厌氧环境下, 且以产甲烷古菌为主, 本文不再详述.在湖泊生态系统中多种形态的水体浮游细菌都可以在胞内或胞外分泌磷酸酶, 进而利用各种复杂形态的有机磷, 形成不同类型的适应状态和生态位分化(Spijkerman & Coesel, 1998 ). ...
Sulfate-reducing bacteria and their activities in cyanobacterial mats of Solar Lake (Sinai, Egypt).
1
1998
... 湖泊水体中与硫循环有关的细菌主要包括硫氧化细菌和硫酸盐还原细菌.其中紫硫细菌是一类化能自养不产氧细菌, 它们以硫化物为电子受体, 在一些盐碱性湖泊中是主要的初级生产者, 在分层湖泊的化学跃变层以下光线可达的水体中, 在小范围内能够积累很大的生物量, 并在湖泊中形成肉眼可见的紫色水层(Rogozin et al ., 2012 ).在化学跃变层的厌氧性水体以及蓝藻暴发后形成的厌氧环境下, 都能够检测到较高丰度的硫酸盐还原细菌(每毫升水体104 -106 个细胞).湖泊水体细菌进行的硫氧化还原过程中常常会利用重金属作为中间电子供体或受体, 从而也被认为在重金属甲基化等过程中发挥重要作用(Teske et al ., 1998 ; Chirwa & Wang, 2000 ; Sorokin et al ., 2011 ). ...
Nitrogen fixation in Lake Mendota, Madison, Wisconsin.
1
1975
... (1)湖泊中的固氮细菌发现的主要是蓝细菌的某些种类, 如念珠藻、水华束丝藻等.这些种类的蓝藻在夏季表层水体无机氮被消耗匮乏时有着很强的生存优势(Torrey & Lee, 1975 ).不过也有学者认为蓝细菌的固氮作用并不显著, 因其可直接利用来自于沉积物中的铵离子和硝酸根离子生存, 并不完全依赖于固氮作用(Ferber et al ., 2004 ).另外一些梭菌、硫螺菌属的细菌在5-10 m深的水柱可以通过异养代谢来进行固氮作用(Keirn & Brezonik, 1971 ). ...
Prokaryotic diversity: magnitude, dynamics, and controlling factors.
1
2002
... 在微生物生态的研究领域长期存在着这样一个观点, 即微生物具有全球性的分布特征, 但只有环境条件合适, 微生物才能够在新生境中定居成功和大量繁衍. 这个观点由Beijerinck (1913)提出, 并且经过Baas-Becking(1934)简要概述为: 微生物可以分布于任何生境中, 但环境在其中起选择作用.细菌具有区别于多细胞生物的3个典型特征: (1)更加丰富的物种多样性; (2)具有远距离被动传播的能力; (3)更快的繁殖速率(Logares et al ., 2009 ).另外, 研究还发现细菌具有极高的基因多样性、表型可塑性和潜在的快速进化能力(Jürgens & Güde, 1994 ).基于细菌的这些特性, 研究者提出这样一个假设: 细菌比多细胞生物更能跨越地理与环境障碍, 成功入侵并定居在不同的生境中.该假设与Beijerinck (1913)提出的细菌全球性分布的观点一致, 即“生物可以分布于任何生境中”.最常被引用来支持这一观点的论据是细菌具有巨大的丰富度和多样性(Fenchel & Finlay, 2004 ).一方面, 细菌个体非常小, 促进了其全球性散布; 另一方面, 细菌通过休眠来最小化灭绝的可能性(Curtis et al ., 2002 , Torsvik et al ., 2002 ).同时当地的环境特征也是调节细菌聚集和分布的关键因子(Martiny et al ., 2006 ).例如: 在湖泊水体中, 细菌多样性会受多种环境资源的影响, 包括生物(例如: 捕食、寄生等)和非生物环境因子(例如: 水体营养盐水平、温度、紫外辐射等).那么, 我们面临着这样一个问题, 即水体细菌的聚集和分布到底是否受生物地理条件的限制? 如果是, 它们与大型有机体分布的生物地理格局是否表现出一致的特点? 水体细菌的生物地理分布特征的生态意义又是什么? 这些尚未解答的问题, 给未来的研究者们带来了新的机遇和挑战. ...
A close link between bacterial community composition and environmental heterogeneity in maritime Antarctic lakes.
1
2010
... 湖泊生态系统, 特别是浅水湖泊生态系统, 一般按初级生产者的组成分成两种类型: 一种是当营养负荷较低时, 以大型沉水植物为主要初级生产者的清水湖泊; 另一种是当营养负荷较高时, 以浮游植物(藻类)为主要初级生产者的浊水湖泊.湖泊从清水草型向浊水藻型转变时(湖泊富营养化), 生物多样性下降(Hanson & Butler, 1994 ; Riemann & Winding, 2001 ; Villaescusa et al ., 2010 ).Hanson和Butler(1994)认为湖泊从清水状态转变为浊水状态后, 虽然没有改变湖泊生态系统中的经典食物链的传递(藻类→浮游动物→鱼类), 但是却导致组成微生物食物网的所有生物的生物量显著增加(Mathes & Arndt, 1994 ). ...
Nitrate accumulation in aerobic hypolimnia: relative importance of benthic and planktonic nitrifiers in an oligotrophic lake.
1
1981
... (2)湖泊中的硝化细菌主要存在于湖泊氧化性水体和表层沉积物之中.但是在湖泊水体分层时期(一般在夏季), 变温层以下的水体由于获得了来自于表层水的氧气, 及来自于湖泊沉积物的还原性氮, 也能观察到明显的硝化作用, 产生硝酸盐的能力大概在0.1-23.2 mmol NO3 - -N/m-3 (Vincent & Downes, 1981 ).硝化螺旋菌、硝化单胞菌是分布较为广泛的水体硝化细菌, 富集试验发现不同来源的硝化细菌对铵离子的亲和力有所不同: 来自于深水层的硝化细菌只能利用中低浓度的氨离子(<5 μM), 而来自于沉积物的硝化细菌能够利用浓度高达12.5 μM的铵离子(Whitby et al ., 2001 ). ...
Do patterns of bacterial diversity along salinity gradients differ from those observed for macroorganisms?
1
2011
... 盐度是内陆水体另一重要环境因子, 但目前相关研究较少.Wu等(2006)通过研究位于青藏高原青海-西藏段上16个高山湖泊(盐度从0.02%的淡水到22.3%的超盐水), 发现在淡水系统和超盐水系统中只有很少一部分细菌组成发生重叠.与大型生物不同的是, 湖泊水体细菌多样性随着水体盐度增加并未呈现减少趋势.细菌在高盐度水体中的高度分化是其多样性显著增加的重要机制(Wang et al ., 2011 ). ...
Abundances, identity, and growth state of Actinobacteria in mountain lakes of different UV transparency.
1
2005
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity.
1
2006
... 原生动物与细菌间的捕食和被捕食关系构成了湖泊微生物网的基础.在捕食细菌的原生动物中, 异养鞭毛虫(HNFs)被认为是影响细菌群落结构和组成的最主要的因子之一(Šimek & Chrzanowski, 1992 ; Young, 2006 ).研究表明HNFs对细菌的捕食作用同样显著改变了细菌群落的形态分布特征, 在高强度的捕食压力下, 中等大小的细菌更容易被捕食, 使得细菌群落形态呈极小或丝状分布(Hahn & Höfle, 2001 ).除大小特征外, Wu等(2004)认为极小或丝状细菌难以被异养鞭毛虫消化也是导致其不被捕食的重要原因.尽管研究者们认识到水体病毒对细菌群落结构的形成存在重要影响(Weinbauer et al ., 2006 ), 但目前, 对病毒如何引起细菌的死亡及影响细菌组成间的竞争的研究仍非常之少. ...
A comparison of ammonia-oxidiser populations in eutrophic and oligotrophic basins of a large freshwater lake.
1
2001
... (2)湖泊中的硝化细菌主要存在于湖泊氧化性水体和表层沉积物之中.但是在湖泊水体分层时期(一般在夏季), 变温层以下的水体由于获得了来自于表层水的氧气, 及来自于湖泊沉积物的还原性氮, 也能观察到明显的硝化作用, 产生硝酸盐的能力大概在0.1-23.2 mmol NO3 - -N/m-3 (Vincent & Downes, 1981 ).硝化螺旋菌、硝化单胞菌是分布较为广泛的水体硝化细菌, 富集试验发现不同来源的硝化细菌对铵离子的亲和力有所不同: 来自于深水层的硝化细菌只能利用中低浓度的氨离子(<5 μM), 而来自于沉积物的硝化细菌能够利用浓度高达12.5 μM的铵离子(Whitby et al ., 2001 ). ...
Vegetation of the Siskiyou mountains, Oregon and California.
1
1960
... 湖泊水体中细菌数量众多, 约105 -108 个/mL, 且种类丰富, 将每一种细菌从湖泊中都采集到几乎不可能, 因此研究者们只能采集有限的样本量, 并通过外推的方法, 估算出湖泊细菌的物种丰度和多样性.目前常用于描述物种多样性的参数有3个, 分别是: α多样性, β多样性和γ多样性(Whittaker, 1960 ). ...
Evolution and measurement of species diversity.
1972
Successful predation of filamentous bacteria by a nanoflagellate challenges current models of flagellate bacterivory.
2004
a) High predictability of the seasonal dynamics of a species-like Polynucleobacter population in a freshwater lake.
1
2006
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
b) Differences in structure and dynamics of Polynucleobacter communities in a temperate and a subtropical lake, revealed at three phylogenetic levels.
1
2006
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
Bacterioplankton community composition along a salinity gradient of sixteen high-mountain lakes located on the Tibetan Plateau, China.
2006
Submersed macrophytes play a key role in structuring bacterioplankton community composition in the large, shallow, subtropical Taihu Lake, China.
1
2007
... 研究表明, 大型沉水植物的生物量和组成是影响太湖中水体细菌群落结构和多样性的关键因子, 同时也是调控太湖水体细菌群落组成季节动态的主要驱动因子(Wu et al ., 2007 ; Zeng et al ., 2012 ).当湖泊水体发生富营养化, 蓝藻爆发有助于具有溶菌作用的病毒和噬细胞菌属(Cytophaga )细菌的生长, 同样显著影响着水体细菌群落结构和多样性(Rashidan & Bird, 2001 ). ...
Geographic and environmental sources of variation in lake bacterial community composition.
2
2005
... pH是决定湖泊细菌群落结构的另一关键因子, 野外原位试验研究表明, 湖泊水体细菌群落结构和多样性随着水体pH变化而发生显著变化(如: Lindström et al ., 2005 ; Yannarell & Triplett, 2005 ).pH可以通过影响不同种类细菌的生长状况(包括: 绝灭、繁衍、种的形成等; Langenheder et al ., 2006 ), 直接影响多样性; 也可通过影响湖泊生态系统中的其他环境因子(如: 有机物质的分子结构; Yannarell & Triplett, 2005 )来间接影响水体细菌群落的结构和多样性. ...
... ), 直接影响多样性; 也可通过影响湖泊生态系统中的其他环境因子(如: 有机物质的分子结构; Yannarell & Triplett, 2005 )来间接影响水体细菌群落的结构和多样性. ...
The selective value of bacterial shape.
1
2006
... 原生动物与细菌间的捕食和被捕食关系构成了湖泊微生物网的基础.在捕食细菌的原生动物中, 异养鞭毛虫(HNFs)被认为是影响细菌群落结构和组成的最主要的因子之一(Šimek & Chrzanowski, 1992 ; Young, 2006 ).研究表明HNFs对细菌的捕食作用同样显著改变了细菌群落的形态分布特征, 在高强度的捕食压力下, 中等大小的细菌更容易被捕食, 使得细菌群落形态呈极小或丝状分布(Hahn & Höfle, 2001 ).除大小特征外, Wu等(2004)认为极小或丝状细菌难以被异养鞭毛虫消化也是导致其不被捕食的重要原因.尽管研究者们认识到水体病毒对细菌群落结构的形成存在重要影响(Weinbauer et al ., 2006 ), 但目前, 对病毒如何引起细菌的死亡及影响细菌组成间的竞争的研究仍非常之少. ...
Macrophyte species drive the variation of bacterioplankton community composition in a shallow freshwater lake.
1
2012
... 研究表明, 大型沉水植物的生物量和组成是影响太湖中水体细菌群落结构和多样性的关键因子, 同时也是调控太湖水体细菌群落组成季节动态的主要驱动因子(Wu et al ., 2007 ; Zeng et al ., 2012 ).当湖泊水体发生富营养化, 蓝藻爆发有助于具有溶菌作用的病毒和噬细胞菌属(Cytophaga )细菌的生长, 同样显著影响着水体细菌群落结构和多样性(Rashidan & Bird, 2001 ). ...
Typical freshwater bacteria: an analysis of available 16S rRNA gene sequences from plankton of lakes and rivers.
2002
Rapid screening for freshwater bacterial groups by using reverse line blot hybridization.
2
2003
... 在较低分类单元上湖泊生态系统中也存在着一些典型的细菌群系, 主要包括: (1)放线菌门中的acI群系, 该群系约占湖泊放线菌总数量的90%(Warnecke et al ., 2005 ), 是放线菌门中数量最多的细菌类群; (2)α-变形菌亚门的LD12群系, 是海洋SAR11细菌的淡水姐妹群系(Logares et al ., 2009 ), 同时也是α-变形菌亚门中分布最为广泛的细菌类群, 广泛分布于全球不同类型的湖泊中(Zwart et al ., 2003 ; Eiler & Bertilsson, 2004 ); (3)β-变形菌亚门中的betI(Limnohabitans )和betII(Polynucleobacter )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...
... )群系, 目前是全球分布最为广泛, 数量最为丰富的细菌类型(Zwart et al ., 2003 ; Hahn, 2003 ; Wu & Hahn, 2006a ,b ; Jezberová et al ., 2010 ; Hahn et al ., 2011 ). ...



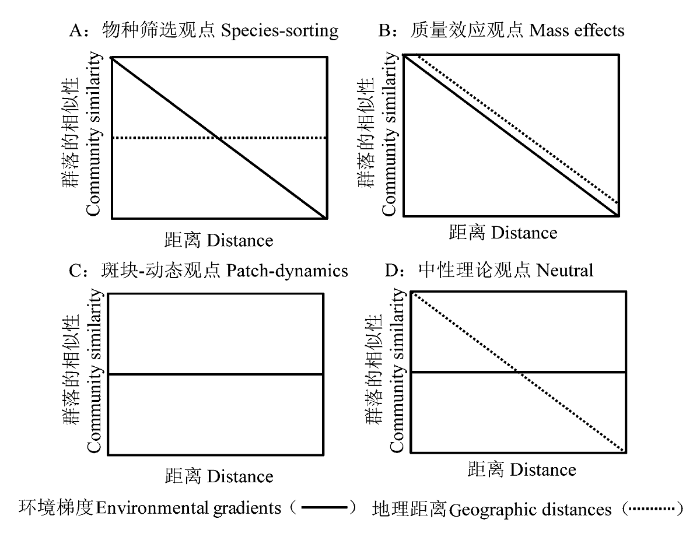
{kind=link}
{kind=link}