0 引言
特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 )。植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 )。分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容。
特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 )。特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 )。植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 )。
近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 )。特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强。
特有类群在不同尺度的生物多样性保护研究中均备受关注。在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 )。在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 )。许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等。
特有现象是一种生物地理学现象, 对特有现象的深入认识体现在生物分类单元和分布区两个概念的理解。其中, 生物分类单元包括等级(科、属、种等)、数量、系统发育关系等。在过去一个世纪中, 系统学研究日新月异, 尤其是分支分类学(cladistic taxonomy)的方法论和DNA测序技术的快速发展, 为系统发育研究提供了较好的基础, 使人们对植物之间的系统发育关系有了更清楚的认识。另外, 化石记录和分子钟技术为进一步估测谱系分支的时间格局提供了保障。这为特有现象的深入理解提供了重要的基础。分布区的概念包括自然区域、行政区域或规范的栅格, 分布数据趋向于规范化和精细化。研究分布区的基本单元数据包括地理坐标点数据和多边形面数据两种形式, 地理坐标点通常以经纬度坐标点来表示; 而面数据关乎尺度大小的问题, 通常称之为空间分析单元(operative geographic unit, OGU)。从当前诸多分析研究来看, OGU基本可分为两大类: 一类为自然地理区或行政区等不规则的区域, 在全球或洲际等大尺度上以及早期的研究中应用较多; 另一类为规则的栅格或网格, 这种规则的操作单元便于采用比较成熟的数量分析方法进行分析, 而且可以直接排除面积效应造成的影响, 因此随着数据精度的不断提高而受到关注(Morrone, 1994 ; Szumik & Goloboff, 2004 )。随着数据采集方式的逐步改进以及现代计算机技术的不断更新, 基于规则栅格或网格的数据收集方法受到更为广泛的关注。
特有现象及其重要性的评价需要借助一定的方法。近年来, 随着不同尺度生物多样性研究工作, 尤其是系统学研究不断深入和地理分布数据进一步规范化, 有关特有植物分布格局的测度方法也有了较大的改进, 尤其在保护生物地理学研究领域(Ladle & Whittaker, 2011 )。本文总结了相关的研究进展, 并将主要的测度方法汇总于表1 , 对比了各种方法的特点、应用现状与前景。
1 物种丰富度的空间分布
1.1 特有物种丰富度
特有物种丰富度(endemic species richness)是指局限分布于某一区域的特有物种的数量。它是大尺度生物多样性保护研究中最常用的算法之一, 特点是简单、直观。特有维管植物丰富度是确定全球生物多样性热点地区的两个重要参考指标之一(Myers et al ., 2000 ; Mittermeier et al ., 2005 )。基于物种丰富度的统计也是当前生物多样性保护策略制定中最主要的参照指标。但由于特有现象本身是尺度依赖的, 且不同的生物地理区域和自然地理区域的差异迥异, 因此, 基于该指数确定的生物多样性热点地区只能在宏观尺度上发挥一定的指导作用。
1.2 狭域特有物种丰富度
为了能更好地对特有中心进行定义, 且更符合一致的统计标准, 一些生物地理学家们主观界定了某一区域范围(区域大小), 将局限分布于该范围内的分类群定义为特有类群, Gaston(1994)将其命名为“边界特有(threshold endemism)”。顾名思义, 狭域特有物种丰富度(narrow-range endemic species richness)就是指分布区狭窄(人为指定某一范围)的特有物种的丰富度。在多数研究中, 人们习惯称此类物种为狭域特有种。例如, 以经纬度网格为例, 把分布区范围局限于一个或两个经纬网格定为特有, 依次类推。该种算法对于狭域特有类群的研究具有较好的效果, 从一定程度上统一了特有物种的分布边界。然而, 由于此种方法是人为界定特有区域范围, 很多研究者质疑其客观性; 而且, 如果定义的范围偏小, 则仅能关注分布区非常狭窄的物种(Crisp et al ., 2001 ; Linder, 2001b ), 而将分布范围稍大的狭域分布物种都排除在外, 因此应用范围非常有限。
1.3 加权特有性
特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法。该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异。物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 )。为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 )。Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1):
(1)
其中, n 是所关注的空间分析单元分布的物种数, W i 是该空间单元中第i 个物种的权重。该算法应用时多是以规则的栅格为空间分布单元, 根据分布区的大小(分布区所覆盖的栅格数量)赋予物种不同的权重, 即其分布区面积(所覆盖栅格数量)的倒数。因此, 物种分布区越小, 被赋予的权重值越高。将每一个栅格中出现的所有物种的权重值加和, 得到相应栅格的加权特有性值。加权特有性值高的地区一般就是狭域特有物种分布较为集中的地区; 反过来, 物种越丰富的地区, 由于物种基数较大, 其加权特有性值也会越高。该方法也被用到不规则空间分析单元的分析中, 此时, 每个目标物种的分布区大小改用所覆盖空间单元的面积之和来表示(Thorne et al. , 2009 ; Huang et al ., 2012 )。
1.4 校正加权特有性
尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 )。可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度。为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE):
(2)
其中, n 是所关注的空间分析单元包含的研究区域的特有物种数, W i 是第i 个空间分析单元中每个特有物种分布的权重, S 是所关注空间分析单元中的总物种数。虽然早有研究提到这种算法(Dony & Denholm, 1985 ), 但由于使用的是特有种的人为边界, 故缺乏客观性(Crisp et al ., 2001 )。
此外, Laffan和Crisp(2003) 在校正加权特有性指数的基础上, 考虑了邻域空间相关性的作用, 发展了空间校正加权特有性(spatial corrected weighted endemism, SCWE)的计算方法:
(3)
其中, N 是与所关注栅格的相邻栅格数, n 是所关注栅格中出现的特有物种数量, R 是第i 个物种分布的栅格数, O 是第i 个物种在相邻栅格中出现的次数。虽然该算法考虑了空间相关作用, 但很难界定邻域空间作用的尺度, 而且在很大程度上模糊了相邻栅格的差异, 因此目前并未得到很好的推广和应用。
2 特有性简约分析及网格式特有性简约分析
特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 )。所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域。尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 )。对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 )。确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的。
特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法。这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域。这一方法的空间分析单元为自然地理单元。然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE)。该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 )。目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 )。
特有性简约分析及相关方法的使用范围非常有限, 往往仅适用于某个类群或相近类群, 所涉及的物种数量通常不超过几百种。该方法的原理是对给定分类单元所有可能的系统发生树进行比较, 选择其中长度最短、代价最小的树作为最终的系统发生树, 即最大简约树, 进而建构出一棵反映分类单元之间最小变化的系统发生树。在实际研究中, 得出的结果很大程度上需要结合研究者的经验进行判断。因此, 该方法在分析某个类群或相近类群的系统演化关系时虽然具有明显的优势, 但在多样性热点地区或特有中心确定的研究中优势并不明显。
3 整合系统发育信息的空间分布格局
3.1 谱系多样性
虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 )。Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性。PD的计算公式见方程(4):
(4)
其中, C 是研究区域所有分类群构成的系统进化树, c 是C 上的一个分支(即两个节点间的片段), Lc 是c 的支长。
系统进化树的构建主要采用化石记录和分子系统学研究相结合的方法。首先通过若干基因测序、比对、拼接, 继而利用最大似然法、最大简约法、贝叶斯法等推断物种之间的进化关系。即先对已知某些类群的共同祖先的年代进行标定, 再通过分子钟等技术, 对进化树内部各节点的分化时间进行推断。这种方法已经成为分子生物地理学及系统发育研究的主要手段之一。但由于当前还很难获得某一地区的所有物种的序列信息, 因此也有人基于被子植物系统树(angiosperm phylogeny group, APG) (Bremer et al ., 2003 )对所研究区域内出现的所有被子植物物种的系统发育关系进行拟合(Webb et al ., 2008 )。
由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 )。但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件。为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE)。该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度。计算公式如下:
(5)
其中, C 为研究区域所有分类群构成的谱系进化树, c 是C 上的一个分支(即两个节点间的片段), Lc 是c 的支长, Rc 是c 分支的分布范围(等于经c 分支分化而来的所有C 集合中物种的分布区面积的并集, 即所有重叠区域的面积仅被计算一次)。
虽然谱系特有性指数将空间尺度和谱系信息进行了很好的整合, 但在应用过程中, 必须对分支与通常的分类概念等级加以区分。此外, 该指数在计算中以分支作为谱系多样性测度的基本单位, 而分支的空间分布范围完全依赖于现存实际分类群的空间分布范围。
3.2 物种进化特异性与空间加权进化特异性
进化特异性(evolutionary distinctiveness, ED)是基于精确的系统进化树结构和明确的类群演化时间信息而建立的另一种反映谱系多样性的指数(Isaac et al ., 2007 )。该指数着重强调了某一分类群(通常指物种)所拥有的特异遗传信息的多少。与Faith的谱系多样性指数的不同在于系统发育树中的每个物种都有对应的进化特异性数值, 从而将特有遗传信息与物种一一对应。Cadotte和Davies (2010)利用进化特异性指数, 并结合物种的空间分布范围, 将物种的空间分布与进化特异性相结合, 提出了空间加权进化特异性(biogeographically weighted evolutionary distinctiveness, BED)指数, 即方程(6):
(6)
其中, n是物种数量, m是分支数量, Lj 是第j分支的支长, Nj 是从第j分支衍生出的终端类群的数量, Ri 是第i个种的分布范围。
该指数兼顾了物种空间分布范围、物种遗传特异性和谱系多样性, 与谱系特有性相比, 具有更强的可操作性。因此, 如果有比较全面和精确的物种遗传序列信息, 那么该方法必将在今后生物多样性优先保护区确定中发挥更大的作用。
4 各个指数的应用现状及前景
参考Web of Science文献索引库, 我们整理了近20年(1993-2012年)各个相关指数的应用情况 (图1 )(检索时间2012年11月)。总体看来, 在涉及特有类群的生物多样性保护研究中, 特有物种丰富度指数的使用最为频繁, 其次为谱系多样性指数(图1 A)。这两个指数都是多样性指数的直接借用。然而从阶段性研究数量占20年研究总量的百分比来看, 涉及系统进化树的指数的研究近年增幅更为明显(图1 B)。
图1 特有类群多样性分布格局不同测度方法的研究数量及随时间的变化(数据源于Web of Science, 检索时间2012年11月)。测度方法名称同表1 。
Fig. 1 The number and percentage of studies measuring patterns of endemic taxa employing different metrics (referring to Web of Science and consulting at November 2012). Codes of different metrics are the same as in Table 1 .
总之, 特有类群格局分析的测度方法已经从简单的数量统计上升到数量生态学测度方法的运用, 从单纯的物种丰富度到谱系多样性, 进而过渡到地理空间分布范围与谱系多样性的结合。这些测度方法的改进和完善, 在很大程度上得益于当前数据信息的收集更加精细以及计算机技术的迅猛发展。数据信息广度和精度以及分析工具的改善, 使有关特有现象本身的研究逐步突破传统的认识和分析手段。随着系统进化树的完善和分析方法的改进, 特有现象的研究难以量化的观念(Good, 1974 )也逐步被改变。
从图1 还可以看出, 基于物种层面特有类群格局分析测度方法的研究仍然是格局分析的主流。原因有二: 一是物种信息是相对容易获取的; 二是对于物种分布格局的认识相对清晰, 有利于进一步探究其成因。虽然关于特有物种分布格局形成机理的研究很多, 但莫衷一是(Cox & Moore, 2005 ; Lomolino et al ., 2006 )。特有性简约分析虽然考虑了系统发育结构关系特征, 但这类方法本身限定了其量化程度, 并且不适用于大类群或多物种的分析。相比之下, 空间地理分布与谱系多样性结合的测度方法, 将会随着数据的不断完善, 尤其是遗传多样性信息的精确测度以及计算机软件的成熟运用和进一步发展, 而呈现更广阔的应用前景。而且由于这类方法具有时空兼顾的特点, 可能为认识动态的生态位保守性理论提供新的途径, 从而建立更加合理的物种分布模型, 为未来各种可能情景下物种多样性分布格局变化提供科学的预测。
5 我国的研究状况及展望
我国是世界上物种多样特别丰富的国家之一, 也是国际上特有种异常丰富的国家之一(McNeely et al ., 1990 ; WCMC, 1992 ), 种子植物中特有种占52.1%(Huang et al ., 2011 )。我国的特有类群为认识生物多样性格局及其形成机理提供了重要契机, 但我国在涉及特有植物格局研究方面与国际研究水平仍相去甚远, 特别是在研究方法的探讨方面还非常薄弱。
首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多。从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 )。虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导。
其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善。相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 )。但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作。
我国相关研究相对滞后的原因主要是由于物种基数大, 在名录和分布信息的整理方面涉及诸多问题。目前我国在物种名录及其分布信息的整理和规范化方面已经有了很好的工作积累(Huang et al. , 2011 ; The Biodiversity Committee of the Chinese Academy of Sciences, 2012 ), 加上计算机技术的发展, 为深入分析中国特有植物多样性分布格局奠定了基础。我们认为具体可以围绕以下四个方面开展工作:
(1)深入物种水平的特有现象格局分析。今后仍需加强基于精细空间分析单元(县或点)的特有物种分布格局的深入研究, 至少在物种分布范围方面还有待深入研究。这不但可以为中国生物多样性保护研究提供客观科学的参考依据, 也有望揭示中国植物区系发生、发展和演化中存在的若干问题。具体可以探讨以下问题:
(i)物种分布区范围的地理空间分布格局及其成因。Brown和Maurer(1989)针对北美和欧洲的物种分布范围进行了研究, 认为自然物理地貌决定了物种分布范围的空间走向。但我国作为在北半球的主要陆地区域, 尚缺乏对这一论断的研究验证。我国自然地貌复杂, 为研究此类问题提供了极为有利的天然试验场。
(ii)探讨现有分布区大小与当前环境梯度的变化关系, 以验证(Qian和Richlefs, 2004) 提出的地史成因较环境因素更有利于特有物种形成的假说。
(iii)探讨物种分布区的破碎化程度是否与物种的系统发育年龄间存在一定的相关性。人们常认为残遗分布区处于持续收缩的状态, 因为残遗种与现在生境条件是不和谐的。也就是说残遗种更容易出现间断分布区, 而新近形成种则更倾向于形成连续分布区。这些问题的验证一方面有助于我们更清晰认识生物地理学理论, 另一方面也为宏观尺度物种多样性分布格局形成机制提供理论支持。
(2)继续开展物种的基因测序工作, 完善谱系多样性格局分析。对于生物多样性保护而言, 所有特有物种的谱系多样性与其物种丰富度空间分布格局是否一致, 还没有明确的结论。基于个别类群的线粒体或叶绿体基因测序构建的进化树表明(Forest et al ., 2007 ), 物种丰富度与谱系多样性的空间分布格局并不完全一致。我们也利用不完善的谱系树进行了相应的研究(Huang et al. , 2012 ), 但由于谱系树精度粗糙, 并未得出物种丰富度和谱系多样性的空间分布格局存在显著差异的结果。上述结果的不同, 很可能是由于谱系树不完全。因此, 要想获得客观的结果, 还需要更多物种的基因序列数据的支持。
(3)开展特有植物区系β多样性格局分析。β多样性度量的是时空尺度上物种组成的变化。直接反映了物种组成的相似性程度, 因而也与植物区系区划和植被区划的关系密切(陈圣宾等, 2010 )。特有植物的分布是植物区系分区的重要参考依据(Takhtajan, 1986 ; 王荷生, 1992 )。吴征镒(1979)在整合大量中国植物特有科、属分布信息的基础上, 确定了中国植物区系分区方案。此外, 中国拥有世界上唯一的包括从寒带到热带各种过渡森林植被类型连续带(吴征镒, 1980 ), 而构成这些森林植被类群主体的优势种及建群种多是中国特有植物, 因此对中国特有植物物种β多样性以及基于精细数据信息的谱系β多样性空间分布格局的研究将会为中国植被区划方案的完善以及中国植被发展历史和演替动态的认识提供新的途径。
(4)开展特有物种分布区变化的模拟与预测。生物类群的过去、现在和将来在地理空间上的分布格局的变化是当前生物地理学家和保护生物学家最关心的问题之一。在当前可获取的信息和保护资源有限的情况下, 针对特有类群的多样性分布格局的研究, 对于探明生物多样性的格局和过程具有重要的启发和借鉴意义。近些年来, 人们对特有现象分布格局已经开展了大量的数据收集和野外调查工作, 并在此基础上初步明确了现存特有类群的空间分布格局。但这些类群为什么会特有?它们是如何产生或形成的?它们的分布在未来有何变化?模型模拟分析是重建过去和预测未来最有力的手段(王娟和倪健, 2006 ), 为回答上述问题提供了可能的途径。现有的研究表明, 特有植物多样性分布格局与一些因素存在明确的相关性, 通过这些关联, 一方面可以借助于古地质学、古生物学提供的历史片段信息, 进行类群的祖先分布区重建(Williams et al. , 2006 ); 另一方面可以利用现在类群分布格局及其形成机理, 建立机理模型, 进行不同情景下分布区变化的模拟(Pearman et al ., 2008 ), 进而为物种 尤其是特有或濒危物种的未来分布格局变化提供可能的预测。
致谢
感谢中央民族大学黄建华老师在本文撰写及修改过程中积极支持和鼓励, 感谢中国科学院植物研究所阳文静博士提供修改建议, 感谢中国科学院沈阳应用生态研究所吕晓涛博士修改英文摘要。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Anderson S 1994 ) Area and endemism.
The Quarterly Review of Biology , 69 , 451 -172 .
[本文引用: 1]
[2]
Beard JS Chapman AR Gioia P 2000 ) Species richness and endemism in the western Australian flora.
Journal of Biogeography , 27 , 1257 -1268 .
[本文引用: 2]
[3]
Borchsenius F 1997 ) Patterns of plant species endemism in Ecuador.
Biodiversity and Conservation , 6 , 379 -399 .
[本文引用: 1]
[4]
Bremer B Bremer K Chase MW Reveal JL Soltis DE Soltis PS Stevens PF Anderberg AA Fay MF Goldblatt P Judd WS Kallersjo M Karehed J Kron KA Lundberg J Nickrent DL Olmstead RG Oxelman B Pires JC Rodman JE Rudall PJ Savolainen V Sytsma KJ van der Bank M Wurdack K Xiang JQY Zmarzty S 2003 ) An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG II.
Botanical Journal of the Linnean Society , 141 , 399 -436 .
[本文引用: 1]
[5]
Brooks TM Mittermeier RA da Fonseca GAB Gerlach J Hoffmann M Lamoreux JF Mittermeier CG Pilgrim JD Rodrigues ASL 2006 ) Global biodiversity conservation priorities.
Science , 313 , 58 -61 .
[本文引用: 3]
[6]
Brown JH Lomolino MV 1998 ) Biogeography, 2nd edn
. Sinauer Associates, Sunderland.
[本文引用: 1]
[7]
Brown JH Maurer BA 1989 ) Macroecology: the division of food and space among species on continents.
Science , 243 , 1145 -1150 .
[8]
Burke A 2004 ) A preliminary account of patterns of endemism in Namibia's Sperrgebiet—the succulent karoo.
Journal of Biogeography , 31 , 1613 -1622 .
[本文引用: 1]
[9]
Burke A 2007 ) Plant endemism in the central Namib Desert.
Evolutionary Ecology Research , 9 , 283 -297 .
[本文引用: 1]
[10]
Cadotte MW Davies TJ 2010 ) Rarest of the rare: advances in combining evolutionary distinctiveness and scarcity to inform conservation at biogeographical scales.
Diversity and Distributions , 16 , 376 -385 .
[11]
Cadotte MW Davies TJ Regetz J Kembel SW Cleland E Oakley TH 2010 ) Phylogenetic diversity metrics for ecological communities: integrating species richness, abundance and evolutionary history.
Ecology Letters , 13 , 96 -105 .
[12]
Cain SA 1944 ) Foundations of Plant Geography
. Harper, New York.
[本文引用: 3]
[13]
Carine MA Humphries CJ Guma IR Reyes-Betancort JA Guerra AS 2009 ) Areas and algorithms: evaluating numerical approaches for the delimitation of areas of endemism in the Canary Islands archipelago.
Journal of Biogeography , 36 , 593 -611 .
[本文引用: 2]
[14]
Chen SB 陈圣宾 ), Ouyang ZY 欧阳志云 ), Xu WH 徐卫华 ), Xiao Y 肖燚 ) (2010 ) A review of beta diversity studies.
Biodiversity Science (生物多样性 ), 18 , 323 -335 . (in Chinese with English abstract)
[本文引用: 1]
[15]
Contreras-Medina R Luna-Vega I 2007 ) Species richness, endemism and conservation of Mexican gymnosperms.
Biodiversity and Conservation , 16 , 1803 -1821 .
[本文引用: 1]
[16]
Cox CB Moore PD 2005 ) Biogeography: An Ecological and Evolutionary Approach, 7th edn . Bath Press , London .
[本文引用: 1]
[17]
Craven P Vorster P 2006 ) Patterns of plant diversity and endemism in Namibia.
Bothalia , 36 , 175 -189 .
[本文引用: 1]
[18]
Crisp MD Laffan S Linder HP Monro A 2001 ) Endemism in the Australian flora.
Journal of Biogeography , 28 , 183 -198 .
[本文引用: 4]
[19]
Dhar U 2002 ) Conservation implications of plant endemism in high-altitude Himalaya.
Current Science , 82 , 141 -148 .
[本文引用: 1]
[20]
Dony JG Denholm I 1985 ) Some quantitative methods of assessing the conservation value of ecologically similar sites.
Journal of Applied Ecology , 22 , 229 -238 .
[本文引用: 1]
[21]
Editorial Committee of Flora Reipublicae Popularis Sinicae (中国植物志编辑委员会 ) (2004 ) Flora Reipublicae Popularis Sinicae, Vol. 1 (中国植物志第一卷 ). Science Press , Beijing . (in Chinese)
[22]
Faith DP 1992 ) Conservation evaluation and phylogenetic diversity.
Biological Conservation , 61 , 1 -10 .
[本文引用: 1]
[23]
Forest F Grenyer R Rouget M Davies TJ Cowling RM Faith DP Balmford A Manning JC Proches S van der Bank M Reeves G Hedderson TAJ Savolainen V 2007 ) Preserving the evolutionary potential of floras in biodiversity hotspots.
Nature , 445 , 757 -760 .
[本文引用: 4]
[24]
Gaston KJ 1994 ) Rarity
. Chapman & Hall, London.
[25]
Gaston KJ Blackburn TM 1996 ) The spatial distribution of threatened species: macroscales and New World birds.
Proceedings of the Royal Society B: Biological Sciences , 263 , 235 -240 .
[本文引用: 2]
[26]
Good R 1974 ) The Geography of the Flowering Plants, 4th edn
. Longman, London.
[本文引用: 4]
[27]
Grill A Casula P Lecis R Menken S 2007 ) Endemism in Sardinia. In: Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity (eds Weiss S, Ferrand N) , pp. 273 -296 . Springer, Berlin.
[本文引用: 1]
[28]
Huang JH Chen B Liu CR Lai JS Zhang JL Ma KP 2012 ) Identifying hotspots of endemic woody seed plant diversity in China.
Diversity and Distributions , 18 , 673 -688 .
[本文引用: 3]
[29]
Huang JH Chen JH Ying JS Ma KP 2011 ) Features and distribution patterns of Chinese endemic seed plant species.
Journal of Systematics and Evolution , 49 , 81 -94 .
[本文引用: 3]
[30]
Huang XL Lei FM Qiao GX 2008 ) Areas of endemism and patterns of diversity for aphids of the Qinghai-Tibetan Plateau and the Himalayas.
Journal of Biogeography , 35 , 230 -240 .
[本文引用: 1]
[31]
Hugo R Exequiel E 2005 ) Plant endemism and natural protected areas in the peninsula of Baja California, Mexico.
Biological Conservation , 122 , 141 -150 .
[本文引用: 2]
[32]
Huston MA 1994 ) Biological Diversity, the Coexistence of Species on Changing Landscapes . Cambridge University Press , Cambridge .
[本文引用: 1]
[33]
Isaac NJB Turvey ST Collen B Waterman C Baillie JEM 2007 ) Mammals on the EDGE: conservation priorities based on threat and phylogeny.
PLoS ONE , 2 , 1 -7 .
[本文引用: 2]
[34]
Kessler M Herzog SK Fjeldså J Back K 2001 ) Species richness and endemism of plant and bird communities along two gradients of elevation, humidity, and land use in the Bolivian Andes.
Diversity and Distributions , 7 , 61 -77 .
[本文引用: 1]
[35]
Ladle RJ Whittaker RJ 2011 ) Conservation Biogeography
. John Wiley & Sons Ltd. Oxford.
[本文引用: 1]
[36]
Laffan SW Crisp MD 2003 ) Assessing endemism at multiple spatial scales, with an example from the Australian vascular flora.
Journal of Biogeography , 30 , 511 -520 .
[本文引用: 2]
[37]
Lamoreux JF Morrison JC Ricketts TH Olson DM Dinerstein E McKnight MW Shugart HH 2006 ) Global tests of biodiversity concordance and the importance of endemism.
Nature , 440 , 212 -214 .
[本文引用: 5]
[38]
Linder HP 1995 ) Setting conservation priorities—the importance of endemism and phylogeny in the southern African orchid genus Herschelia.
Conservation Biology , 9 , 585 -595 .
[本文引用: 2]
[39]
Linder HP 2001 a) On areas of endemism, with an example from the African Restionaceae.
Systematic Biology , 50 , 892 -912 .
[40]
Linder HP 2001 b) Plant diversity and endemism in sub-Saharan tropical Africa.
Journal of Biogeography , 28 , 169 -182 .
[本文引用: 4]
[41]
Lomolino MV Riddle BR Brown JH 2006 ) Biogeography, 3rd edn
. Sinauer Associates, Inc., Sunderland, Massachusetts.
[本文引用: 8]
[42]
Lopez-Pujol J Zhang FM Sun HQ Ying TS Ge S 2011 ) Centres of plant endemism in China: places for survival or for speciation ?
Journal of Biogeography , 38 , 1267 -1280 .
[本文引用: 1]
[43]
Ma KP 马克平 ), Qian YQ 钱迎倩 ), Wang C 王晨 ) (1994 ) Status and future trends of biological diversity research. In: Principles and Methodologies of Biodiversity Studies (生物多样性研究的原理与方法) (eds Qian YQ (钱迎倩), Ma KP (马克平)) , pp. 1 -12 . Chinese Science and Technology Press , Beijing . (in Chinese)
[本文引用: 1]
[44]
Mace GM Gittleman JL Purvis A 2003 ) Preserving the tree of life.
Science , 300 , 1707 -1709 .
[本文引用: 1]
[45]
Malcolm JR Liu CR Neilson RP Hansen L Hannah L 2006 ) Global warming and extinctions of endemic species from biodiversity hotspots.
Conservation Biology , 20 , 538 -548 .
[本文引用: 1]
[46]
May RM 1990 ) Taxonomy as destiny.
Nature , 347 , 129 -130 .
[本文引用: 1]
[47]
McNeely JA Miller KR Reid WV Mittermeier RA Werner TB 1990 ) Conserving the World’s Biological Diversity
. IUCN, WRI, WWF, World Bank, Washington DC.
[本文引用: 1]
[48]
Mittermeier RA Gil PR Hoffman M Pilgrim J Brooks T Mittermeier CG Lamoreux J da Fonseca GAB 2005 ) Hotspots Revisited . The University of Chicago Press , Chicago .
[本文引用: 1]
[49]
Mooers AO Atkins RA 2003 ) Indonesia's threatened birds: over 500 million years of evolutionary heritage at risk.
Animal Conservation , 6 , 183 -188 .
[本文引用: 1]
[50]
Morrone JJ 1994 ) On the identification of areas of endemism.
Systematic Biology , 43 , 438 -441 .
[本文引用: 4]
[51]
Myers N 1990 ) The biodiversity challenge expanded hotspots analysis.
The Environmentalist , 10 , 243 -256 .
[本文引用: 1]
[52]
Myers N Mittermeier RA Mittermeier CG da Fonseca GAB Kent J 2000 ) Biodiversity hotspots for conservation priorities.
Nature , 403 , 853 -858 .
[本文引用: 4]
[53]
Nelson GJ Platnick N 1981 ) Systematics and Biogeography: Cladistics and Vicariance . Columbia University Press , New York .
[本文引用: 1]
[54]
Orme CDL Davies RG Burgess M Eigenbrod F Pickup N Olson VA Webster AJ Ding TS Rasmussen PC Ridgely RS Stattersfield AJ Bennett PM Blackburn TM Gaston KJ Owens IPF 2005 ) Global hotspots of species richness are not congruent with endemism or threat.
Nature , 436 , 1016 -1019 .
[本文引用: 2]
[55]
Pearman PB Guisan A Broennimann O Randin CF 2008 ) Niche dynamics in space and time.
Trends in Ecology and Evolution , 23 , 149 -158 .
[本文引用: 1]
[56]
Polasky S Csuti B Vossler CA Meyers SM 2001 ) A comparison of taxonomic distinctness versus richness as criteria for setting conservation priorities for North American birds.
Biological Conservation , 97 , 99 -105 .
[本文引用: 1]
[57]
Prendergast JR Quinn RM Lawton JH Eversham BC Gibbons DW 1993 ) Rare species, the coincidence of diversity hotspots and conservation strategies.
Nature , 365 , 335 -337 .
[本文引用: 1]
[58]
Qian H Ricklefs RE 2004 ) Geographical distribution and ecological conservatism of disjunct genera of vascular plants in eastern Asia and eastern North America.
Journal of Ecology , 92 , 253 -265 .
[本文引用: 1]
[59]
Raven PH Axelrod DI 1974 ) Angiosperm biogeography and past continental movements.
Annals of the Missouri Botanical Garden , 61 , 539 -673 .
[本文引用: 1]
[60]
Redding DW Mooers AO 2006 ) Incorporating evolutionary measures into conservation prioritization.
Conservation Biology , 20 , 1670 -1678 .
[本文引用: 1]
[61]
Reyjol Y Hugueny B Pont D Bianco PG Beier U Caiola N Casals F Cowx I Economou A Ferreira T Haidvogl G Noble R de Sostoa A Vigneron T Virbickas T 2007 ) Patterns in species richness and endemism of European freshwater fish.
Global Ecology and Biogeography , 16 , 65 -75 .
[本文引用: 1]
[62]
Reyjol Y Hugueny B Bianco PG Pont D 2008 ) Congruence between riverine fish species richness and endemism at the biogeographical scale: the role of history.
Aquatic Conservation: Marine and Freshwater Ecosystems , 18 , 1311 -1314 .
[本文引用: 2]
[63]
Richardson IBK 1978 ) Endemic taxa and the taxonomist. In: Essays in Plant Taxonomy (ed. Steet HE) , pp. 245 -262 . Academic Press , London; New York .
[本文引用: 1]
[64]
Riemann H Ezcurra E 2005 ) Plant endemism and natural protected areas in the peninsula of Baja California, Mexico.
Biological Conservation , 122 , 141 -150 .
[本文引用: 1]
[65]
Rodrigues ASL Gaston KJ 2002 ) Maximising phylogenetic diversity in the selection of networks of conservation areas.
Biological Conservation , 105 , 103 -111 .
[本文引用: 2]
[66]
Rosauer D Laffan SW Crisp MD Donnellan SC Cook LG 2009 ) Phylogenetic endemism: a new approach for identifying geographical concentrations of evolutionary history.
Molecular Ecology , 18 , 4061 -4072 .
[本文引用: 1]
[67]
Rosen BR 1992 ) Empiricism and the biogeographical black-box—concepts and methods in marine paleobiogeography
. Palaeogeography, Palaeoclimatology, Palaeoecology , 92 , 171 -205 .
[本文引用: 1]
[68]
Rosen BR Smith AB 1988 ) Tectonics from fossils? Analysis of reef coral and sea urchin distributions from late Cretaceous to recent, using a new method. In: Gondwana and Tethys (eds Audley-Charles MG, Hallam A) , pp. 275 -306 . Geological Society of London, Oxford.
[本文引用: 1]
[69]
Sechrest W Brooks TM da Fonseca GAB Konstant WR Mittermeier RA Purvis A Rylands AB Gittleman JL 2002 ) Hotspots and the conservation of evolutionary history.
Proceedings of the National Academy of Sciences, USA , 99 , 2067 -2071 .
[本文引用: 1]
[70]
Szumik CA Goloboff PA 2004 ) Areas of endemism: an improved optimality criterion.
Systematic Biology , 53 , 968 -977 .
[本文引用: 1]
[71]
Takhtajan AL 1986 ) Floristic Regions of the World . University of California Press , Berkeley .
[本文引用: 2]
[72]
Temu RPC Andrew SM 2008 ) Endemism of plants in the Uluguru Mountains, Morogoro, Tanzania.
Forest Ecology and Management , 255 , 2858 -2869 .
[本文引用: 1]
[73]
The Biodiversity Committee of the Chinese Academy of Sciences (2012 ) Catalogue of Life China: 2012 Annual Checklist . Science Press , Beijing .
[本文引用: 1]
[74]
Thorne JH Viers JH Price J Storns DM 2009 ) Spatial patterns of endemic plants in California.
Natural Areas Journal , 29 , 344 -366 .
[本文引用: 1]
[75]
Torres NM Diniz JAF 2004 ) Phylogenetic autocorrelation and evolutionary diversity of Carnivora (Mammalia) in conservation units of the New World.
Genetics and Molecular Biology , 27 , 511 -516 .
[本文引用: 1]
[76]
Tribsch A Schonswetter P 2003 ) Patterns of endemism and comparative phylogeography confirm palaeoenvironmental evidence for Pleistocene refugia in the eastern Alps.
Taxon , 52 , 477 -497 .
[本文引用: 1]
[77]
Trinder-Smith TH Cowling RM Linder HP 1996 ) Profiling a besieged flora: endemic and threatened plants of the Cape Peninsula, South Africa.
Biodiversity and Conservation , 5 , 575 -589 .
[本文引用: 1]
[78]
Vane-Wright RI Humphries CJ Williams PH 1991 ) What to protect? Systematics and the agony of choice.
Biological Conservation , 55 , 235 -254 .
[本文引用: 1]
[79]
Xing FW 邢福武 ),Wu DL 吴德邻 ), Li ZX 李泽贤 ), Ye HG 叶华谷 ), Chen BH 陈炳辉 ) (1995 ) Endemic plants of Hainan Island.
Journal of Tropical and Subtropical Botany (热带亚热带植物学报 ), 3 , 1 -12 . (in Chinese with English abstract)
[本文引用: 1]
[80]
Wang HS 王荷生 ) (1992 ) Floristic Plant Geography (植物区系地理 ). Science Press , Beijing . (in Chinese)
[本文引用: 2]
[81]
Wang HS 王荷生 ), Zhang YL 张镱锂 ) (1994 a) The distribution patterns of spermatophytic families and genera endemic to China.
Acta Geographica Sinica (地理学报 ), 49 , 403 -417 . (in Chinese with English abstract)
[82]
Wang HS 王荷生 ), Zhang YL 张镱锂 ) (1994 b) The biodiversity and characters of spermatophytic genera endemic to China.
Acta Botanica Yunnanica (云南植物研究 ), 16 , 209 -220 . (in Chinese with English abstract)
[本文引用: 1]
[83]
Wang J 王娟 ), Ni J 倪健 ) (2006 ) Review of modelling the distribution of plant species.
Journal of Plant Ecology , 30 , 1040 -1053 . (in Chinese with English abstract)
[本文引用: 1]
[84]
Wang S 汪松 ) (1998 ) China Red Data Book of Endangered Animals (中国濒危动物红皮书 ). Science Press , Beijing . (in Chinese)
[本文引用: 1]
[85]
WCMC (1992 ) Global Biodiversity: Status of the Earth’s Living Resources
. Chapman and Hall, London.
[本文引用: 1]
[86]
Webb CO Ackerly DD Kembel SW 2008 ) Phylocom: software for the analysis of phylogenetic community structure and trait evolution.
Bioinformatics , 24 , 2098 -2100 .
[本文引用: 1]
[87]
Whittaker RJ Araujo MB Paul J Ladle RJ Watson JEM Willis KJ 2005 ) Conservation biogeography: assessment and prospect.
Diversity and Distributions , 11 , 3 -23 .
[88]
Williams P 2000 ) Some properties of rarity scores used in site quality assessment.
British Journal of Entomology and Natural History , 13 , 73 -86 .
[本文引用: 1]
[89]
Williams PD Pollock DD Blackburne BP Goldstein RA 2006 ) Assessing the accuracy of ancestral protein reconstruction methods.
PLOS Computational Biology , 2 , 598 -605 .
[本文引用: 1]
[90]
Williams PH Prance GT Humphries CJ Edwards KS 1996 ) Promise and problems in applying quantitative complementary areas for representing the diversity of some neotropical plants (families Dichapetalaceae, Lecythidaceae, Caryocaraceae, Chrysobalanaceae and Proteaceae).
Biological Journal of the Linnean Society , 58 , 125 -157 .
[本文引用: 1]
[91]
Woinarski JCZ Hempel C Cowie I Brennan K Kerrigan R Leach G Russell-Smith J 2006 ) Distributional pattern of plant species endemic to the Northern Territory, Australia.
Australian Journal of Botany , 54 , 627 -640 .
[本文引用: 1]
[92]
Wood JRI 2006 ) Inter-Andean dry valleys of Bolivia—floristic affinities and patterns of endemism: insights from Acanthaceae, Asclepiadaceae and Labiatae. In: Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography and Conservation (eds Pennington RT, Lewis GP, Ratter JA) , pp. 235 -256 . CRC Press , Boca Raton .
[本文引用: 2]
[93]
Wu ZY 吴征镒 ) (1979 ) The regionalization of Chinese flora.
Acta Botanica Yunnanica (云南植物研究 ), 1 , 1 -19 . (in Chinese with English abstract)
[94]
Wu ZY 吴征镒 ) (1980 ) Vegetation of China (中国植被 ). Science Press , Beijing . (in Chinese)
[本文引用: 2]
[95]
Wu ZY 吴征镒 ), Wang HS 王荷生 ) (1983 ) Physical Geography of China: Phytogeography, Vol. 1 (中国植物地理: 植物地理, 上册 ). Science Press , Beijing . (in Chinese)
[96]
Wulff EV 1960 ) (translated by Zhong CX (仲崇信), Zhang MZ (张梦庄)) An Introduction to Historical Plant Geography (历史植物地理学引论 ). Science Press , Beijing . (in Chinese)
[97]
Yang QR 杨亲二 ), Zuo JP 左家哺 ) (1998 ) Endemism of higher plants in China. In: Research and Conservation of Species Diversity (eds Song YL (宋延龄), Yang QR (杨亲二), Huang YQ (黄永青)) , pp. 79 -87 . Zhejiang Science and Technology Press, Hangzhou. (in Chinese with English abstract)
[本文引用: 1]
[98]
Ying JS 应俊生 ) (1996 ) Areography of the endemic genera of seed plants in China.
Acta Phytotaxonomica Sinica (植物分类学报 ), 34 , 479 -485 . (in Chinese with English abstract)
[本文引用: 1]
[99]
Ying JS 应俊生 ), Zhang YL 张玉龙 ) (1994 ) The Endemic Genera of Seed Plants of China (中国种子植物特有属 ). Science Press , Beijing . (in Chinese)
[本文引用: 1]
[100]
Yu XL 喻勋林 ), Xiao YT 肖育檀 ) (1999 ) Primary study of limestone endemic plants in Hunan Province.
Journal of Central South Forestry University (中南林学院学报 ), 19 , 34 -38 . (in Chinese with English abstract)
[本文引用: 1]
[101]
Zang DK 臧德奎 ), Fan JH 樊金会 ), Zhao LY 赵兰勇 ), Fan BM 樊宝敏 ) (1994 ) Study on the endemic plants from Shandong Province.
Bulletin of Botanical Research (植物研究 ), 14 , 48 -58 . (in Chinese with English abstract)
[本文引用: 1]
[102]
Zhang W 张伟 ), Zhao SL 赵善伦 ) (2000 ) Studies on the biodiversity and protection of Chinese endemic of tracheophyta in Shandong Province.
Journal of Shandong Normal University (山东师范大学学报 ), 15 , 409 -412 . (in Chinese with English abstract)
[本文引用: 1]
[103]
Zhao YZ 赵一之 ), Cao R 曹瑞 ) (1996 ) The endemic plants in Nei Monggol.
Acta Scientiarum Naturalium Universitatis NeiMonggol (内蒙古大学学报 ), 27 , 208 -213 . (in Chinese with English abstract)
[本文引用: 1]
Area and endemism.
1
1994
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
Species richness and endemism in the western Australian flora.
2
2000
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Patterns of plant species endemism in Ecuador.
1
1997
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG II.
1
2003
... 系统进化树的构建主要采用化石记录和分子系统学研究相结合的方法.首先通过若干基因测序、比对、拼接, 继而利用最大似然法、最大简约法、贝叶斯法等推断物种之间的进化关系.即先对已知某些类群的共同祖先的年代进行标定, 再通过分子钟等技术, 对进化树内部各节点的分化时间进行推断.这种方法已经成为分子生物地理学及系统发育研究的主要手段之一.但由于当前还很难获得某一地区的所有物种的序列信息, 因此也有人基于被子植物系统树(angiosperm phylogeny group, APG) (Bremer et al ., 2003 )对所研究区域内出现的所有被子植物物种的系统发育关系进行拟合(Webb et al ., 2008 ). ...
Global biodiversity conservation priorities.
3
2006
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
Biogeography, 2nd edn
1
1998
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
Macroecology: the division of food and space among species on continents.
1989
A preliminary account of patterns of endemism in Namibia's Sperrgebiet—the succulent karoo.
1
2004
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Plant endemism in the central Namib Desert.
1
2007
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
Rarest of the rare: advances in combining evolutionary distinctiveness and scarcity to inform conservation at biogeographical scales.
2010
Phylogenetic diversity metrics for ecological communities: integrating species richness, abundance and evolutionary history.
2010
Foundations of Plant Geography
3
1944
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
... ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
Areas and algorithms: evaluating numerical approaches for the delimitation of areas of endemism in the Canary Islands archipelago.
2
2009
... 特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 ).所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域.尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
A review of beta diversity studies.
1
2010
... (3)开展特有植物区系β多样性格局分析.β多样性度量的是时空尺度上物种组成的变化.直接反映了物种组成的相似性程度, 因而也与植物区系区划和植被区划的关系密切(陈圣宾等, 2010 ).特有植物的分布是植物区系分区的重要参考依据(Takhtajan, 1986 ; 王荷生, 1992 ).吴征镒(1979)在整合大量中国植物特有科、属分布信息的基础上, 确定了中国植物区系分区方案.此外, 中国拥有世界上唯一的包括从寒带到热带各种过渡森林植被类型连续带(吴征镒, 1980 ), 而构成这些森林植被类群主体的优势种及建群种多是中国特有植物, 因此对中国特有植物物种β多样性以及基于精细数据信息的谱系β多样性空间分布格局的研究将会为中国植被区划方案的完善以及中国植被发展历史和演替动态的认识提供新的途径. ...
Species richness, endemism and conservation of Mexican gymnosperms.
1
2007
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
1
2005
... 从图1 还可以看出, 基于物种层面特有类群格局分析测度方法的研究仍然是格局分析的主流.原因有二: 一是物种信息是相对容易获取的; 二是对于物种分布格局的认识相对清晰, 有利于进一步探究其成因.虽然关于特有物种分布格局形成机理的研究很多, 但莫衷一是(Cox & Moore, 2005 ; Lomolino et al ., 2006 ).特有性简约分析虽然考虑了系统发育结构关系特征, 但这类方法本身限定了其量化程度, 并且不适用于大类群或多物种的分析.相比之下, 空间地理分布与谱系多样性结合的测度方法, 将会随着数据的不断完善, 尤其是遗传多样性信息的精确测度以及计算机软件的成熟运用和进一步发展, 而呈现更广阔的应用前景.而且由于这类方法具有时空兼顾的特点, 可能为认识动态的生态位保守性理论提供新的途径, 从而建立更加合理的物种分布模型, 为未来各种可能情景下物种多样性分布格局变化提供科学的预测. ...
Patterns of plant diversity and endemism in Namibia.
1
2006
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Endemism in the Australian flora.
4
2001
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 为了能更好地对特有中心进行定义, 且更符合一致的统计标准, 一些生物地理学家们主观界定了某一区域范围(区域大小), 将局限分布于该范围内的分类群定义为特有类群, Gaston(1994)将其命名为“边界特有(threshold endemism)”.顾名思义, 狭域特有物种丰富度(narrow-range endemic species richness)就是指分布区狭窄(人为指定某一范围)的特有物种的丰富度.在多数研究中, 人们习惯称此类物种为狭域特有种.例如, 以经纬度网格为例, 把分布区范围局限于一个或两个经纬网格定为特有, 依次类推.该种算法对于狭域特有类群的研究具有较好的效果, 从一定程度上统一了特有物种的分布边界.然而, 由于此种方法是人为界定特有区域范围, 很多研究者质疑其客观性; 而且, 如果定义的范围偏小, 则仅能关注分布区非常狭窄的物种(Crisp et al ., 2001 ; Linder, 2001b ), 而将分布范围稍大的狭域分布物种都排除在外, 因此应用范围非常有限. ...
... 其中, n 是所关注的空间分析单元包含的研究区域的特有物种数, W i 是第i 个空间分析单元中每个特有物种分布的权重, S 是所关注空间分析单元中的总物种数.虽然早有研究提到这种算法(Dony & Denholm, 1985 ), 但由于使用的是特有种的人为边界, 故缺乏客观性(Crisp et al ., 2001 ). ...
Conservation implications of plant endemism in high-altitude Himalaya.
1
2002
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Some quantitative methods of assessing the conservation value of ecologically similar sites.
1
1985
... 其中, n 是所关注的空间分析单元包含的研究区域的特有物种数, W i 是第i 个空间分析单元中每个特有物种分布的权重, S 是所关注空间分析单元中的总物种数.虽然早有研究提到这种算法(Dony & Denholm, 1985 ), 但由于使用的是特有种的人为边界, 故缺乏客观性(Crisp et al ., 2001 ). ...
Conservation evaluation and phylogenetic diversity.
1
1992
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
Preserving the evolutionary potential of floras in biodiversity hotspots.
4
2007
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
... ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
... (2)继续开展物种的基因测序工作, 完善谱系多样性格局分析.对于生物多样性保护而言, 所有特有物种的谱系多样性与其物种丰富度空间分布格局是否一致, 还没有明确的结论.基于个别类群的线粒体或叶绿体基因测序构建的进化树表明(Forest et al ., 2007 ), 物种丰富度与谱系多样性的空间分布格局并不完全一致.我们也利用不完善的谱系树进行了相应的研究(Huang et al. , 2012 ), 但由于谱系树精度粗糙, 并未得出物种丰富度和谱系多样性的空间分布格局存在显著差异的结果.上述结果的不同, 很可能是由于谱系树不完全.因此, 要想获得客观的结果, 还需要更多物种的基因序列数据的支持. ...
The spatial distribution of threatened species: macroscales and New World birds.
2
1996
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
The Geography of the Flowering Plants, 4th edn
4
1974
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
... ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
... 总之, 特有类群格局分析的测度方法已经从简单的数量统计上升到数量生态学测度方法的运用, 从单纯的物种丰富度到谱系多样性, 进而过渡到地理空间分布范围与谱系多样性的结合.这些测度方法的改进和完善, 在很大程度上得益于当前数据信息的收集更加精细以及计算机技术的迅猛发展.数据信息广度和精度以及分析工具的改善, 使有关特有现象本身的研究逐步突破传统的认识和分析手段.随着系统进化树的完善和分析方法的改进, 特有现象的研究难以量化的观念(Good, 1974 )也逐步被改变. ...
1
2007
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Identifying hotspots of endemic woody seed plant diversity in China.
3
2012
... 其中, n 是所关注的空间分析单元分布的物种数, W i 是该空间单元中第i 个物种的权重.该算法应用时多是以规则的栅格为空间分布单元, 根据分布区的大小(分布区所覆盖的栅格数量)赋予物种不同的权重, 即其分布区面积(所覆盖栅格数量)的倒数.因此, 物种分布区越小, 被赋予的权重值越高.将每一个栅格中出现的所有物种的权重值加和, 得到相应栅格的加权特有性值.加权特有性值高的地区一般就是狭域特有物种分布较为集中的地区; 反过来, 物种越丰富的地区, 由于物种基数较大, 其加权特有性值也会越高.该方法也被用到不规则空间分析单元的分析中, 此时, 每个目标物种的分布区大小改用所覆盖空间单元的面积之和来表示(Thorne et al. , 2009 ; Huang et al ., 2012 ). ...
... 其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善.相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 ).但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作. ...
... (2)继续开展物种的基因测序工作, 完善谱系多样性格局分析.对于生物多样性保护而言, 所有特有物种的谱系多样性与其物种丰富度空间分布格局是否一致, 还没有明确的结论.基于个别类群的线粒体或叶绿体基因测序构建的进化树表明(Forest et al ., 2007 ), 物种丰富度与谱系多样性的空间分布格局并不完全一致.我们也利用不完善的谱系树进行了相应的研究(Huang et al. , 2012 ), 但由于谱系树精度粗糙, 并未得出物种丰富度和谱系多样性的空间分布格局存在显著差异的结果.上述结果的不同, 很可能是由于谱系树不完全.因此, 要想获得客观的结果, 还需要更多物种的基因序列数据的支持. ...
Features and distribution patterns of Chinese endemic seed plant species.
3
2011
... 我国是世界上物种多样特别丰富的国家之一, 也是国际上特有种异常丰富的国家之一(McNeely et al ., 1990 ; WCMC, 1992 ), 种子植物中特有种占52.1%(Huang et al ., 2011 ).我国的特有类群为认识生物多样性格局及其形成机理提供了重要契机, 但我国在涉及特有植物格局研究方面与国际研究水平仍相去甚远, 特别是在研究方法的探讨方面还非常薄弱. ...
... 其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善.相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 ).但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作. ...
... 我国相关研究相对滞后的原因主要是由于物种基数大, 在名录和分布信息的整理方面涉及诸多问题.目前我国在物种名录及其分布信息的整理和规范化方面已经有了很好的工作积累(Huang et al. , 2011 ; The Biodiversity Committee of the Chinese Academy of Sciences, 2012 ), 加上计算机技术的发展, 为深入分析中国特有植物多样性分布格局奠定了基础.我们认为具体可以围绕以下四个方面开展工作: ...
Areas of endemism and patterns of diversity for aphids of the Qinghai-Tibetan Plateau and the Himalayas.
1
2008
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
Plant endemism and natural protected areas in the peninsula of Baja California, Mexico.
2
2005
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
1
1994
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
Mammals on the EDGE: conservation priorities based on threat and phylogeny.
2
2007
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
... 进化特异性(evolutionary distinctiveness, ED)是基于精确的系统进化树结构和明确的类群演化时间信息而建立的另一种反映谱系多样性的指数(Isaac et al ., 2007 ).该指数着重强调了某一分类群(通常指物种)所拥有的特异遗传信息的多少.与Faith的谱系多样性指数的不同在于系统发育树中的每个物种都有对应的进化特异性数值, 从而将特有遗传信息与物种一一对应.Cadotte和Davies (2010)利用进化特异性指数, 并结合物种的空间分布范围, 将物种的空间分布与进化特异性相结合, 提出了空间加权进化特异性(biogeographically weighted evolutionary distinctiveness, BED)指数, 即方程(6): ...
Species richness and endemism of plant and bird communities along two gradients of elevation, humidity, and land use in the Bolivian Andes.
1
2001
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Conservation Biogeography
1
2011
... 特有现象及其重要性的评价需要借助一定的方法.近年来, 随着不同尺度生物多样性研究工作, 尤其是系统学研究不断深入和地理分布数据进一步规范化, 有关特有植物分布格局的测度方法也有了较大的改进, 尤其在保护生物地理学研究领域(Ladle & Whittaker, 2011 ).本文总结了相关的研究进展, 并将主要的测度方法汇总于表1 , 对比了各种方法的特点、应用现状与前景. ...
Assessing endemism at multiple spatial scales, with an example from the Australian vascular flora.
2
2003
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 此外, Laffan和Crisp(2003) 在校正加权特有性指数的基础上, 考虑了邻域空间相关性的作用, 发展了空间校正加权特有性(spatial corrected weighted endemism, SCWE)的计算方法: ...
Global tests of biodiversity concordance and the importance of endemism.
5
2006
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Setting conservation priorities—the importance of endemism and phylogeny in the southern African orchid genus Herschelia.
2
1995
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
a) On areas of endemism, with an example from the African Restionaceae.
2001
Plant diversity and endemism in sub-Saharan tropical Africa.
4
2001
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 为了能更好地对特有中心进行定义, 且更符合一致的统计标准, 一些生物地理学家们主观界定了某一区域范围(区域大小), 将局限分布于该范围内的分类群定义为特有类群, Gaston(1994)将其命名为“边界特有(threshold endemism)”.顾名思义, 狭域特有物种丰富度(narrow-range endemic species richness)就是指分布区狭窄(人为指定某一范围)的特有物种的丰富度.在多数研究中, 人们习惯称此类物种为狭域特有种.例如, 以经纬度网格为例, 把分布区范围局限于一个或两个经纬网格定为特有, 依次类推.该种算法对于狭域特有类群的研究具有较好的效果, 从一定程度上统一了特有物种的分布边界.然而, 由于此种方法是人为界定特有区域范围, 很多研究者质疑其客观性; 而且, 如果定义的范围偏小, 则仅能关注分布区非常狭窄的物种(Crisp et al ., 2001 ; Linder, 2001b ), 而将分布范围稍大的狭域分布物种都排除在外, 因此应用范围非常有限. ...
... 特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 ).所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域.尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
Biogeography, 3rd edn
8
2006
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
... 特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 ).所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域.尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
... ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
... 从图1 还可以看出, 基于物种层面特有类群格局分析测度方法的研究仍然是格局分析的主流.原因有二: 一是物种信息是相对容易获取的; 二是对于物种分布格局的认识相对清晰, 有利于进一步探究其成因.虽然关于特有物种分布格局形成机理的研究很多, 但莫衷一是(Cox & Moore, 2005 ; Lomolino et al ., 2006 ).特有性简约分析虽然考虑了系统发育结构关系特征, 但这类方法本身限定了其量化程度, 并且不适用于大类群或多物种的分析.相比之下, 空间地理分布与谱系多样性结合的测度方法, 将会随着数据的不断完善, 尤其是遗传多样性信息的精确测度以及计算机软件的成熟运用和进一步发展, 而呈现更广阔的应用前景.而且由于这类方法具有时空兼顾的特点, 可能为认识动态的生态位保守性理论提供新的途径, 从而建立更加合理的物种分布模型, 为未来各种可能情景下物种多样性分布格局变化提供科学的预测. ...
Centres of plant endemism in China: places for survival or for speciation ?
1
2011
... 其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善.相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 ).但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作. ...
1
1994
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
Preserving the tree of life.
1
2003
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
Global warming and extinctions of endemic species from biodiversity hotspots.
1
2006
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Taxonomy as destiny.
1
1990
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
Conserving the World’s Biological Diversity
1
1990
... 我国是世界上物种多样特别丰富的国家之一, 也是国际上特有种异常丰富的国家之一(McNeely et al ., 1990 ; WCMC, 1992 ), 种子植物中特有种占52.1%(Huang et al ., 2011 ).我国的特有类群为认识生物多样性格局及其形成机理提供了重要契机, 但我国在涉及特有植物格局研究方面与国际研究水平仍相去甚远, 特别是在研究方法的探讨方面还非常薄弱. ...
1
2005
... 特有物种丰富度(endemic species richness)是指局限分布于某一区域的特有物种的数量.它是大尺度生物多样性保护研究中最常用的算法之一, 特点是简单、直观.特有维管植物丰富度是确定全球生物多样性热点地区的两个重要参考指标之一(Myers et al ., 2000 ; Mittermeier et al ., 2005 ).基于物种丰富度的统计也是当前生物多样性保护策略制定中最主要的参照指标.但由于特有现象本身是尺度依赖的, 且不同的生物地理区域和自然地理区域的差异迥异, 因此, 基于该指数确定的生物多样性热点地区只能在宏观尺度上发挥一定的指导作用. ...
Indonesia's threatened birds: over 500 million years of evolutionary heritage at risk.
1
2003
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
On the identification of areas of endemism.
4
1994
... 特有现象是一种生物地理学现象, 对特有现象的深入认识体现在生物分类单元和分布区两个概念的理解.其中, 生物分类单元包括等级(科、属、种等)、数量、系统发育关系等.在过去一个世纪中, 系统学研究日新月异, 尤其是分支分类学(cladistic taxonomy)的方法论和DNA测序技术的快速发展, 为系统发育研究提供了较好的基础, 使人们对植物之间的系统发育关系有了更清楚的认识.另外, 化石记录和分子钟技术为进一步估测谱系分支的时间格局提供了保障.这为特有现象的深入理解提供了重要的基础.分布区的概念包括自然区域、行政区域或规范的栅格, 分布数据趋向于规范化和精细化.研究分布区的基本单元数据包括地理坐标点数据和多边形面数据两种形式, 地理坐标点通常以经纬度坐标点来表示; 而面数据关乎尺度大小的问题, 通常称之为空间分析单元(operative geographic unit, OGU).从当前诸多分析研究来看, OGU基本可分为两大类: 一类为自然地理区或行政区等不规则的区域, 在全球或洲际等大尺度上以及早期的研究中应用较多; 另一类为规则的栅格或网格, 这种规则的操作单元便于采用比较成熟的数量分析方法进行分析, 而且可以直接排除面积效应造成的影响, 因此随着数据精度的不断提高而受到关注(Morrone, 1994 ; Szumik & Goloboff, 2004 ).随着数据采集方式的逐步改进以及现代计算机技术的不断更新, 基于规则栅格或网格的数据收集方法受到更为广泛的关注. ...
... 特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 ).所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域.尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
... ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
The biodiversity challenge expanded hotspots analysis.
1
1990
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Biodiversity hotspots for conservation priorities.
4
2000
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
... 特有物种丰富度(endemic species richness)是指局限分布于某一区域的特有物种的数量.它是大尺度生物多样性保护研究中最常用的算法之一, 特点是简单、直观.特有维管植物丰富度是确定全球生物多样性热点地区的两个重要参考指标之一(Myers et al ., 2000 ; Mittermeier et al ., 2005 ).基于物种丰富度的统计也是当前生物多样性保护策略制定中最主要的参照指标.但由于特有现象本身是尺度依赖的, 且不同的生物地理区域和自然地理区域的差异迥异, 因此, 基于该指数确定的生物多样性热点地区只能在宏观尺度上发挥一定的指导作用. ...
1
1981
... 特有区域(areas of endemism, AOE)是分支生物地理学(cladistic biogeography)分析的基本单元, 是历史生物地理学研究的基本问题(Nelson & Platnick, 1981 ; Morrone, 1994 ).所谓特有区域是指有两个或两个以上特有类群重叠或分布一致的区域.尽管特有区域的定义看起来非常简单, 但在实际操作中却困难重重(Lomolino et al ., 2006 ), 其定义仍存在一定争议(Linder, 2001 ; Lomolino et al ., 2006 ; Carine et al ., 2009 ).对于特有区域的界定主要强调以下三个标准: (1)物种分布区重叠或物种并存的程度, (2)存在引起物种隔离分化的地理屏障, (3)地理屏障引起的同宗异域进化(Lomolino et al ., 2006 ).确定特有区域的研究方法也有所不同(Lomolino et al ., 2006 ), 总体上包括基于简约分析(parsimony analysis, PA)和聚类分析两大类, 其他算法都是由这两种算法衍生出来的. ...
Global hotspots of species richness are not congruent with endemism or threat.
2
2005
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Niche dynamics in space and time.
1
2008
... (4)开展特有物种分布区变化的模拟与预测.生物类群的过去、现在和将来在地理空间上的分布格局的变化是当前生物地理学家和保护生物学家最关心的问题之一.在当前可获取的信息和保护资源有限的情况下, 针对特有类群的多样性分布格局的研究, 对于探明生物多样性的格局和过程具有重要的启发和借鉴意义.近些年来, 人们对特有现象分布格局已经开展了大量的数据收集和野外调查工作, 并在此基础上初步明确了现存特有类群的空间分布格局.但这些类群为什么会特有?它们是如何产生或形成的?它们的分布在未来有何变化?模型模拟分析是重建过去和预测未来最有力的手段(王娟和倪健, 2006 ), 为回答上述问题提供了可能的途径.现有的研究表明, 特有植物多样性分布格局与一些因素存在明确的相关性, 通过这些关联, 一方面可以借助于古地质学、古生物学提供的历史片段信息, 进行类群的祖先分布区重建(Williams et al. , 2006 ); 另一方面可以利用现在类群分布格局及其形成机理, 建立机理模型, 进行不同情景下分布区变化的模拟(Pearman et al ., 2008 ), 进而为物种 尤其是特有或濒危物种的未来分布格局变化提供可能的预测. ...
A comparison of taxonomic distinctness versus richness as criteria for setting conservation priorities for North American birds.
1
2001
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
Rare species, the coincidence of diversity hotspots and conservation strategies.
1
1993
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Geographical distribution and ecological conservatism of disjunct genera of vascular plants in eastern Asia and eastern North America.
1
2004
... (ii)探讨现有分布区大小与当前环境梯度的变化关系, 以验证(Qian和Richlefs, 2004) 提出的地史成因较环境因素更有利于特有物种形成的假说. ...
Angiosperm biogeography and past continental movements.
1
1974
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
Incorporating evolutionary measures into conservation prioritization.
1
2006
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
Patterns in species richness and endemism of European freshwater fish.
1
2007
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
Congruence between riverine fish species richness and endemism at the biogeographical scale: the role of history.
2
2008
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
... 尽管一些研究表明在某些群落或类群中, 总物种丰富度和特有物种丰富度的分布格局并不一致(Prendergast et al ., 1993 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 但也有很多研究表明特有现象中心与物种丰富度中心重叠(Beard et al ., 2000 ; Kessler et al ., 2001 ; Contreras-Medina & Luna-Vega, 2007 ; Reyjol et al ., 2007 , 2008 ).可见加权特有性值的空间分布格局很大程度上依赖于物种丰富程度.为充分体现特有物种的空间分布特征, 同时避免物种丰富度本身对特有植物分布格局的影响, Crisp等(2001)利用加权特有性值与总物种丰富度的比值来体现某一地区的特有性, 即校正加权特有性指数(corrected weighted endemism, CWE): ...
1
1978
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
Plant endemism and natural protected areas in the peninsula of Baja California, Mexico.
1
2005
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
Maximising phylogenetic diversity in the selection of networks of conservation areas.
2
2002
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
Phylogenetic endemism: a new approach for identifying geographical concentrations of evolutionary history.
1
2009
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
Empiricism and the biogeographical black-box—concepts and methods in marine paleobiogeography
1
1992
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
1
1988
... 特有现象对种系发生研究具有重要意义, 而简约分析是系统发生学中的重要分析方法, 因此, 特有性简约分析(parsimony analysis of endemicity, PAE)是分支生物地理学中用于分析特有现象分布格局的常用方法.这一方法最早由Rosen等提出(Rosen & Smith, 1988 ; Rosen, 1992 ), 他们基于物种的自然地理分布和系统发育树来确定物种分布的特有区域.这一方法的空间分析单元为自然地理单元.然而, 很多自然地理单元的边界很难界定, 因此, Juan J. Morrone在此基础上, 引入规则的网格作为空间分析单元(Morrone, 1994 ; Lomolino et al ., 2006 ), 并将其定义为样方式特有性简约分析(Quadrat-PAE).该方法的计算可分为以下5步: (1)将整个研究区域划分为规则的网格; (2)确定研究区域内所有物种的地理分布; (3)创建物种分布位点矩阵, 即物种对应栅格的0/1矩阵; (4)基于(3)的物种分布矩阵执行最大简约分支分析, 构建最大简约树; (5)基于(4)中分支系统树的结果选定特有区域(Morrone, 1994 ; Lomolino et al ., 2006 ).目前该方法仍然是特有区域确定的主要分析方法(Huang et al ., 2008 ; Carine et al ., 2009 ). ...
Hotspots and the conservation of evolutionary history.
1
2002
... 由于谱系多样性指数反映了系统发育信息的多样性特征, 而且与物种多样性的分布格局并不完全一致(Rodrigues & Gaston, 2002 ; Mace et al ., 2003 ; Forest et al ., 2007 ), 因此, 该测度方法引起了保护生物学家更多的关注, 并被引入生物多样性格局分析及优先保护区评定的研究之中(Sechrest et al ., 2002 ; Mooers & Atkins, 2003 ; Redding & Mooers, 2006 ; Forest et al ., 2007 ; Isaac et al ., 2007 ; Rosauer et al ., 2009 ).但是由于该指数仅考虑了分类群在系统进化树中的信息, 并没有兼顾分类群的空间分布范围, 而分类群的空间分布范围是当前优先保护区确定时考虑的首要条件.为了让谱系多样性特征在生物多样性优先保护区确定中发挥更大的作用, Rosauer等(2009)将分类群的空间分布范围与系统发育信息结合, 创建了谱系特有性指数(phylogenetic endemism, PE).该指数不但测度了空间分布范围的相对变化, 同时也反映了分类群在系统进化树上的分散程度.计算公式如下: ...
Areas of endemism: an improved optimality criterion.
1
2004
... 特有现象是一种生物地理学现象, 对特有现象的深入认识体现在生物分类单元和分布区两个概念的理解.其中, 生物分类单元包括等级(科、属、种等)、数量、系统发育关系等.在过去一个世纪中, 系统学研究日新月异, 尤其是分支分类学(cladistic taxonomy)的方法论和DNA测序技术的快速发展, 为系统发育研究提供了较好的基础, 使人们对植物之间的系统发育关系有了更清楚的认识.另外, 化石记录和分子钟技术为进一步估测谱系分支的时间格局提供了保障.这为特有现象的深入理解提供了重要的基础.分布区的概念包括自然区域、行政区域或规范的栅格, 分布数据趋向于规范化和精细化.研究分布区的基本单元数据包括地理坐标点数据和多边形面数据两种形式, 地理坐标点通常以经纬度坐标点来表示; 而面数据关乎尺度大小的问题, 通常称之为空间分析单元(operative geographic unit, OGU).从当前诸多分析研究来看, OGU基本可分为两大类: 一类为自然地理区或行政区等不规则的区域, 在全球或洲际等大尺度上以及早期的研究中应用较多; 另一类为规则的栅格或网格, 这种规则的操作单元便于采用比较成熟的数量分析方法进行分析, 而且可以直接排除面积效应造成的影响, 因此随着数据精度的不断提高而受到关注(Morrone, 1994 ; Szumik & Goloboff, 2004 ).随着数据采集方式的逐步改进以及现代计算机技术的不断更新, 基于规则栅格或网格的数据收集方法受到更为广泛的关注. ...
2
1986
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
... (3)开展特有植物区系β多样性格局分析.β多样性度量的是时空尺度上物种组成的变化.直接反映了物种组成的相似性程度, 因而也与植物区系区划和植被区划的关系密切(陈圣宾等, 2010 ).特有植物的分布是植物区系分区的重要参考依据(Takhtajan, 1986 ; 王荷生, 1992 ).吴征镒(1979)在整合大量中国植物特有科、属分布信息的基础上, 确定了中国植物区系分区方案.此外, 中国拥有世界上唯一的包括从寒带到热带各种过渡森林植被类型连续带(吴征镒, 1980 ), 而构成这些森林植被类群主体的优势种及建群种多是中国特有植物, 因此对中国特有植物物种β多样性以及基于精细数据信息的谱系β多样性空间分布格局的研究将会为中国植被区划方案的完善以及中国植被发展历史和演替动态的认识提供新的途径. ...
Endemism of plants in the Uluguru Mountains, Morogoro, Tanzania.
1
2008
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
1
2012
... 我国相关研究相对滞后的原因主要是由于物种基数大, 在名录和分布信息的整理方面涉及诸多问题.目前我国在物种名录及其分布信息的整理和规范化方面已经有了很好的工作积累(Huang et al. , 2011 ; The Biodiversity Committee of the Chinese Academy of Sciences, 2012 ), 加上计算机技术的发展, 为深入分析中国特有植物多样性分布格局奠定了基础.我们认为具体可以围绕以下四个方面开展工作: ...
Spatial patterns of endemic plants in California.
1
2009
... 其中, n 是所关注的空间分析单元分布的物种数, W i 是该空间单元中第i 个物种的权重.该算法应用时多是以规则的栅格为空间分布单元, 根据分布区的大小(分布区所覆盖的栅格数量)赋予物种不同的权重, 即其分布区面积(所覆盖栅格数量)的倒数.因此, 物种分布区越小, 被赋予的权重值越高.将每一个栅格中出现的所有物种的权重值加和, 得到相应栅格的加权特有性值.加权特有性值高的地区一般就是狭域特有物种分布较为集中的地区; 反过来, 物种越丰富的地区, 由于物种基数较大, 其加权特有性值也会越高.该方法也被用到不规则空间分析单元的分析中, 此时, 每个目标物种的分布区大小改用所覆盖空间单元的面积之和来表示(Thorne et al. , 2009 ; Huang et al ., 2012 ). ...
Phylogenetic autocorrelation and evolutionary diversity of Carnivora (Mammalia) in conservation units of the New World.
1
2004
... 虽然已有研究表明物种丰富度能有效地替代谱系多样性(Polasky et al ., 2001 ; Rodrigues & Gaston, 2002 ; Torres & Diniz, 2004 ; Brooks et al ., 2006 ), 但也有研究表明物种多样性和谱系多样性格局明显不一致(Faith, 1992 ; Forest et al ., 2007 ).Faith(1992)首先定义谱系多样性指数(phylogenetic diversity, PD)为某区域内出现的所有分类群对应的最小生成路径支长的和, 反映了某一地区物种组成的系统进化特征多样性.PD的计算公式见方程(4): ...
Patterns of endemism and comparative phylogeography confirm palaeoenvironmental evidence for Pleistocene refugia in the eastern Alps.
1
2003
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
Profiling a besieged flora: endemic and threatened plants of the Cape Peninsula, South Africa.
1
1996
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
What to protect? Systematics and the agony of choice.
1
1991
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
Endemic plants of Hainan Island.
1
1995
... 首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多.从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 ).虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导. ...
2
1992
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
... (3)开展特有植物区系β多样性格局分析.β多样性度量的是时空尺度上物种组成的变化.直接反映了物种组成的相似性程度, 因而也与植物区系区划和植被区划的关系密切(陈圣宾等, 2010 ).特有植物的分布是植物区系分区的重要参考依据(Takhtajan, 1986 ; 王荷生, 1992 ).吴征镒(1979)在整合大量中国植物特有科、属分布信息的基础上, 确定了中国植物区系分区方案.此外, 中国拥有世界上唯一的包括从寒带到热带各种过渡森林植被类型连续带(吴征镒, 1980 ), 而构成这些森林植被类群主体的优势种及建群种多是中国特有植物, 因此对中国特有植物物种β多样性以及基于精细数据信息的谱系β多样性空间分布格局的研究将会为中国植被区划方案的完善以及中国植被发展历史和演替动态的认识提供新的途径. ...
The distribution patterns of spermatophytic families and genera endemic to China.
1994
The biodiversity and characters of spermatophytic genera endemic to China.
1
1994
... 其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善.相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 ).但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作. ...
Review of modelling the distribution of plant species.
1
2006
... (4)开展特有物种分布区变化的模拟与预测.生物类群的过去、现在和将来在地理空间上的分布格局的变化是当前生物地理学家和保护生物学家最关心的问题之一.在当前可获取的信息和保护资源有限的情况下, 针对特有类群的多样性分布格局的研究, 对于探明生物多样性的格局和过程具有重要的启发和借鉴意义.近些年来, 人们对特有现象分布格局已经开展了大量的数据收集和野外调查工作, 并在此基础上初步明确了现存特有类群的空间分布格局.但这些类群为什么会特有?它们是如何产生或形成的?它们的分布在未来有何变化?模型模拟分析是重建过去和预测未来最有力的手段(王娟和倪健, 2006 ), 为回答上述问题提供了可能的途径.现有的研究表明, 特有植物多样性分布格局与一些因素存在明确的相关性, 通过这些关联, 一方面可以借助于古地质学、古生物学提供的历史片段信息, 进行类群的祖先分布区重建(Williams et al. , 2006 ); 另一方面可以利用现在类群分布格局及其形成机理, 建立机理模型, 进行不同情景下分布区变化的模拟(Pearman et al ., 2008 ), 进而为物种 尤其是特有或濒危物种的未来分布格局变化提供可能的预测. ...
1
1998
... 近几十年来, 随着全球生物多样性保护研究的深入, 特有现象及其分布格局的研究受到生态学家和保护生物学家的广泛关注(马克平等, 1994 ; Myers et al ., 2000 ; Hugo & Exequiel, 2005 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).特有物种已成为确定生物多样性优先保护区的重要标准(Myers et al ., 2000 ; Orme et al ., 2005 ; Lamoreux et al ., 2006 ), 主要原因如下: 第一, 由于分布区域狭窄, 特有物种的潜在受威胁风险更大(Linder, 1995 ; Gaston & Blackburn, 1996 ); 第二, 在大尺度上, 特有现象明显的地区通常是生物多样性十分丰富的地区(Huston, 1994 ), 特有种分布中心往往也是物种多样性聚集的中心(Beard et al ., 2000 ; Reyjol et al ., 2008 ); 第三, 濒危物种有很多是分布区狭窄的物种(Wang, 1998 ), 保护特有物种分布区, 通常能更好地保护濒危物种; 第四, 在资源有限的情况下, 人类目前尚没有能力去调查清楚所有的物种, 特有物种边界明确, 可操作性强. ...
Global Biodiversity: Status of the Earth’s Living Resources
1
1992
... 我国是世界上物种多样特别丰富的国家之一, 也是国际上特有种异常丰富的国家之一(McNeely et al ., 1990 ; WCMC, 1992 ), 种子植物中特有种占52.1%(Huang et al ., 2011 ).我国的特有类群为认识生物多样性格局及其形成机理提供了重要契机, 但我国在涉及特有植物格局研究方面与国际研究水平仍相去甚远, 特别是在研究方法的探讨方面还非常薄弱. ...
Phylocom: software for the analysis of phylogenetic community structure and trait evolution.
1
2008
... 系统进化树的构建主要采用化石记录和分子系统学研究相结合的方法.首先通过若干基因测序、比对、拼接, 继而利用最大似然法、最大简约法、贝叶斯法等推断物种之间的进化关系.即先对已知某些类群的共同祖先的年代进行标定, 再通过分子钟等技术, 对进化树内部各节点的分化时间进行推断.这种方法已经成为分子生物地理学及系统发育研究的主要手段之一.但由于当前还很难获得某一地区的所有物种的序列信息, 因此也有人基于被子植物系统树(angiosperm phylogeny group, APG) (Bremer et al ., 2003 )对所研究区域内出现的所有被子植物物种的系统发育关系进行拟合(Webb et al ., 2008 ). ...
Conservation biogeography: assessment and prospect.
2005
Some properties of rarity scores used in site quality assessment.
1
2000
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
Assessing the accuracy of ancestral protein reconstruction methods.
1
2006
... (4)开展特有物种分布区变化的模拟与预测.生物类群的过去、现在和将来在地理空间上的分布格局的变化是当前生物地理学家和保护生物学家最关心的问题之一.在当前可获取的信息和保护资源有限的情况下, 针对特有类群的多样性分布格局的研究, 对于探明生物多样性的格局和过程具有重要的启发和借鉴意义.近些年来, 人们对特有现象分布格局已经开展了大量的数据收集和野外调查工作, 并在此基础上初步明确了现存特有类群的空间分布格局.但这些类群为什么会特有?它们是如何产生或形成的?它们的分布在未来有何变化?模型模拟分析是重建过去和预测未来最有力的手段(王娟和倪健, 2006 ), 为回答上述问题提供了可能的途径.现有的研究表明, 特有植物多样性分布格局与一些因素存在明确的相关性, 通过这些关联, 一方面可以借助于古地质学、古生物学提供的历史片段信息, 进行类群的祖先分布区重建(Williams et al. , 2006 ); 另一方面可以利用现在类群分布格局及其形成机理, 建立机理模型, 进行不同情景下分布区变化的模拟(Pearman et al ., 2008 ), 进而为物种 尤其是特有或濒危物种的未来分布格局变化提供可能的预测. ...
Promise and problems in applying quantitative complementary areas for representing the diversity of some neotropical plants (families Dichapetalaceae, Lecythidaceae, Caryocaraceae, Chrysobalanaceae and Proteaceae).
1
1996
... 特有物种丰富度和边界特有物种丰富度都是统计特有物种的绝对数量, 因此也称为绝对特有值算法.该类方法仅考虑了分类群的丰富度, 而没有考虑到物种分布区大小的差异.物种的分布区域越狭窄, 其潜在受威胁风险就越高(Linder, 1995 ; Gaston & Blackburn, 1996 ), 因此, 在保护中赋予所有的分类群相同的重要程度的方法是不合理的(May, 1990 ; Vane-Wright et al ., 1991 ).为此, Williams等基于优先保护分布区狭窄的分类群的原则, 提出了分布区加权的算法(Williams et al ., 1996 ; Williams, 2000 ).Crisp(2001)等将其应用到植物特有现象的格局分析中, 并命名为加权特有性(weighted endemism, WE), 具体计算如公式(1): ...
Distributional pattern of plant species endemic to the Northern Territory, Australia.
1
2006
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
2
2006
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
... 特有类群在不同尺度的生物多样性保护研究中均备受关注.在全球尺度上, 特有物种数量已经成为确定全球生物多样性“热点地区”的重要参考指标之一(Myers, 1990 ; Myers et al ., 2000 ; Brooks et al ., 2006 ; Lamoreux et al ., 2006 ).在区域尺度上, 特有物种分布格局的研究不仅被用于确定优先保护区(Crisp et al ., 2001 ; Linder, 2001b ; Laffan & Crisp, 2003 )和评估生物多样性保护区的保护效果(Hugo & Exequiel, 2005 ; Riemann & Ezcurra, 2005 ; Lamoreux et al ., 2006 ), 而且也成为开展生物多样性分布中心预测的重要证据(Malcolm et al ., 2006 ).许多国家和地区都利用特有维管植物分布格局的相关信息来确定植物多样性分布的热点地区和优先保护区, 例如非洲的坦桑尼亚(Temu & Andrew, 2008 )、纳米比亚(Burke, 2004 ; Craven & Vorster, 2006 )、撒哈拉地区(Linder, 2001b )、开普半岛(Trinder-Smith et al ., 1996 ), 南美洲的厄瓜多尔(Borchsenius, 1997 )、波利维亚(Wood, 2006 ), 欧亚大陆的意大利撒丁岛(Grill et al ., 2007 )、喜马拉雅地区(Dhar, 2002 )和澳大利亚(Crisp et al ., 2001 ; Woinarski et al ., 2006 )等. ...
The regionalization of Chinese flora.
1979
2
1980
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
... (3)开展特有植物区系β多样性格局分析.β多样性度量的是时空尺度上物种组成的变化.直接反映了物种组成的相似性程度, 因而也与植物区系区划和植被区划的关系密切(陈圣宾等, 2010 ).特有植物的分布是植物区系分区的重要参考依据(Takhtajan, 1986 ; 王荷生, 1992 ).吴征镒(1979)在整合大量中国植物特有科、属分布信息的基础上, 确定了中国植物区系分区方案.此外, 中国拥有世界上唯一的包括从寒带到热带各种过渡森林植被类型连续带(吴征镒, 1980 ), 而构成这些森林植被类群主体的优势种及建群种多是中国特有植物, 因此对中国特有植物物种β多样性以及基于精细数据信息的谱系β多样性空间分布格局的研究将会为中国植被区划方案的完善以及中国植被发展历史和演替动态的认识提供新的途径. ...
1
1998
... 特有现象(endemism)是指某一生物类群单元(如种、属或科)局限分布于某一地理区域内的现象(Cain, 1944 ; Good, 1974 ; Anderson, 1994 ; Brown & Lomolino, 1998 ).植物分类单元经历了起源、分化、迁移和灭绝等事件, 这一进化历史过程在不同地区表现出不均匀的属性, 使得不同植物区系中特有现象的显著程度、性质、组成以及起源和演化过程都有所不同, 形成了世界不同地区植物区系的多样性和复杂性(杨亲二和左家哺, 1998 ).分布区学(Areography)是生物地理学的重要分支之一(Lomolino et al. , 2006 ), 特有现象是分布区学研究的核心内容(Cain, 1944 ; Good, 1974 ), 因此, 特有现象是生物地理学研究的重要内容. ...
Areography of the endemic genera of seed plants in China.
1
1996
... 特有现象是一个地区最重要的区系特征之一, 对于认识一个地区植物区系的性质、发生和演变具有十分重要的意义(Cain, 1944 ; Good, 1974 ; Wood, 2006 ; Burke, 2007 ).特有植物的分布格局是植物区系区划和植被区划的重要依据(吴征镒, 1980 ; Takhtajan, 1986 ; 王荷生, 1992 ; 应俊生, 1996 ).植物种系发生(phylogenesis)的历史与其分布区的地质历史密切相关, 因此特有现象的研究也能为解决系统发育问题提供有价值的线索(Raven & Axelrod, 1974 ; Richardson, 1978 ), 例如, 分布格局与分子系统学结合为维管植物冰期孑遗区的研究提供了有效途径(Tribsch & Schonswetter, 2003 ). ...
1
1994
... 其次, 我国虽然已经在全国层面上初步开展了中国特有种子植物的多样性分布格局研究(Huang et al. , 2011 ), 同时也以特有木本种子植物为例探讨了确定特有植物多样性热点地区的分析方法(Huang et al. , 2012 ), 但这些研究从数据精度和研究的系统性上都有待进一步完善.相对而言, 针对全国特有植物属的多样性分布格局的研究开展较早(王荷生和张镱锂, 1994b ; 应俊生和张玉龙, 1994 ),不但揭示了多样性的空间分布格局, 同时也探讨了其成因, 尤其是古、新特有属的研究为揭示我国植物多样性保存和孑遗中心提供了重要证据(Lopez-Pujol et al. , 2011 ).但由于属水平的研究主要揭示的是地质历史时间尺度的特征和规律, 并不能充分体现物种水平的空间分布格局, 尤其是在生态时间尺度上的变化特征; 要想实现对特有植物多样性分布格局的全面认识及其机理的揭示, 还需要在物种水平上深入开展研究工作. ...
Primary study of limestone endemic plants in Hunan Province.
1
1999
... 首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多.从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 ).虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导. ...
Study on the endemic plants from Shandong Province.
1
1994
... 首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多.从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 ).虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导. ...
Studies on the biodiversity and protection of Chinese endemic of tracheophyta in Shandong Province.
1
2000
... 首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多.从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 ).虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导. ...
The endemic plants in Nei Monggol.
1
1996
... 首先, 我国对特有类群的研究尚处在起步阶段, 涉及其分布格局的研究为数不多.从物种水平看, 已有的研究主要侧重区域性的植物区系特征分析和资源类群调查等描述性的工作(臧得奎等, 1994 ; 邢福武等, 1995 ; 喻勋林和肖育檀, 1996 ; 赵一之和曹瑞, 1996 ; 张伟和赵善伦, 2000 ).虽然这些工作对我国区域性植物多样性保护研究提供了重要的数据基础, 但从全国范围来看, 这些区域性的研究对全国特有植物多样性资源的保护尚不能提供宏观层面的指导. ...



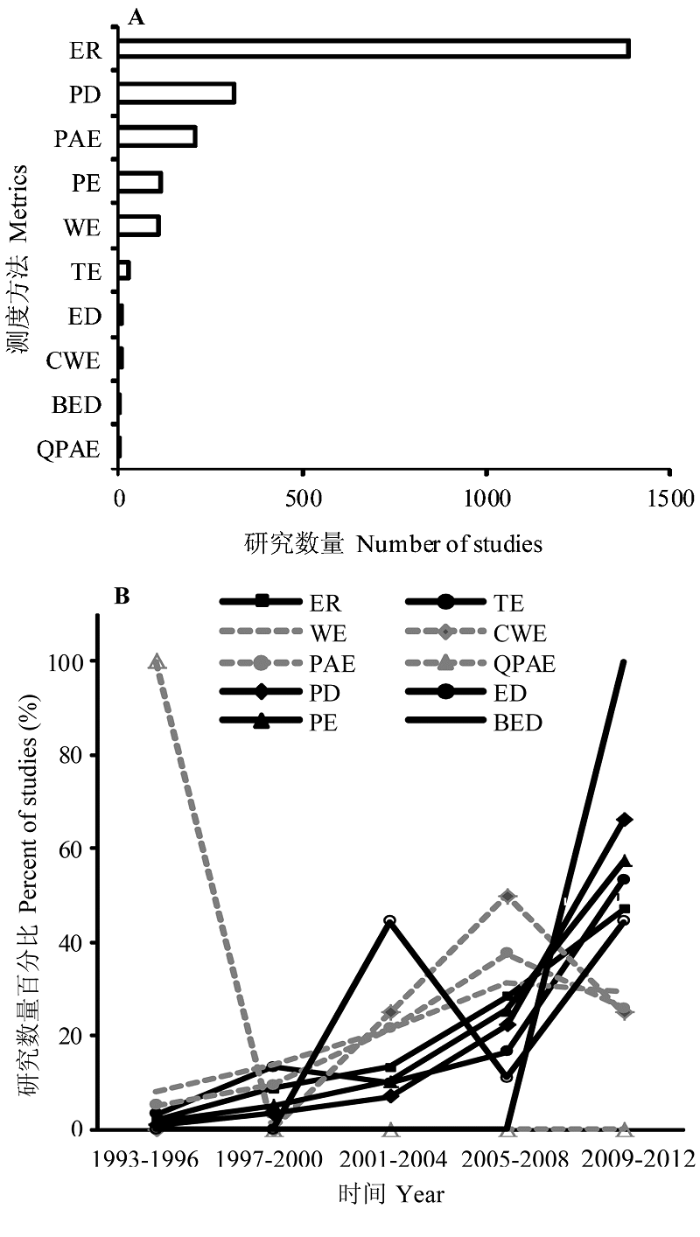
{kind=link}
{kind=link}