0 引言
生态位模型是利用物种已知的分布数据和相关环境变量, 根据一定的算法运算来构建模型, 判断物种的生态需求, 并将运算结果投射(project)至不同的时间和空间中以预测物种的实际分布(actual distribution)和潜在分布(potential distribution)。设想在一个多维的自然空间环境中存在一个点, 这个点关联着多个环境变量参数, 那么就可以将一个物种所有分布点所关联的环境参数抽提出来, 基于一定的算法来推算这个物种的生态需求, 然后将其投射至不同时间和空间的地理空间中, 以推测该物种在特定条件下潜在的分布范围。国际上对生态位模型的说法不一致, 其名称在我国的使用也不一致, 如物种分布模型、物种预测模型、生境预测模型、生态位模型等。
近年来, 生态位模型在生物多样性保护的多个领域得到开拓和应用, 被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布的影响及传染病空间传播的研究中。但是, 在模型运用时却得出了一些不符合实际的结论, 如入侵物种的生态位迁移等(Broennimann et al ., 2007 ; Medley, 2010 ), 故一些学者对生态位模型提出了质疑。这些不当之处或多或少是由于对模型构建的目的认识模糊, 对生态位的概念和生态位模型的原理, 或者模型软件算法理解不透彻造成的(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 )。为了让读者更系统地理解和正确地应用生态位模型, 我们梳理了生态位与物种分布的关系, 结合生态位概念介绍了生态位模型的基本原理, 从物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘种生态位保守性及基于生态位分化的物种界定等方面阐述了它在生物多样性保护中的应用。
1 物种分布与生态位
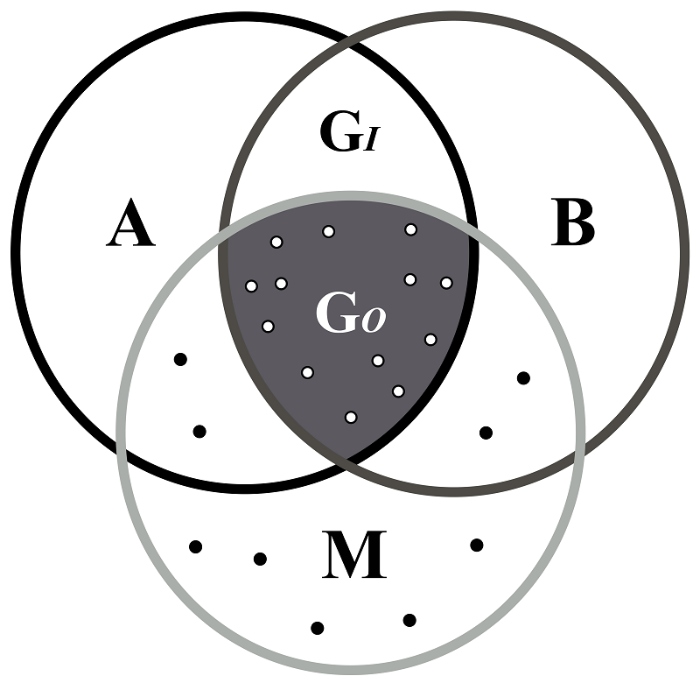
物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 )。在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义。在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 )。Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力。通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 )。前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 )。Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 )。
图1 影响物种地理分布的因素修改自(Soberón & Peterson, 2005 ; Soberón, 2010 )。A环表示对物种分布有利的非生物因素(基础生态位), B环表示物种相互作用中对该物种有利的区域, M环表示物种迁移能力范围内的区域, Go I
Fig. 1 Diagram showing a simplified framework for understanding species distribution adopted from (Soberón & Peterson, 2005 ; Soberón, 2010 ). Overlap of the three circles (GO I
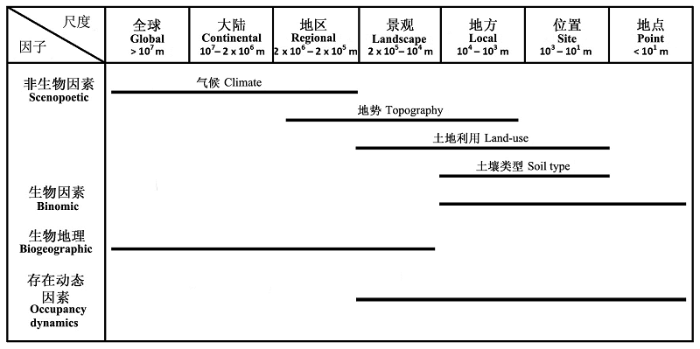
一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等。不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 )。生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 )。这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 )。Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类。前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 )。
图2 不同影响因素在不同的空间尺度下作用于物种的分布修改自(Pearson & Dawson, 2003 ; Soberón, 2007 , 2010 ; Hortal et al ., 2010 )
Fig. 2 Relative importance of factors affecting species distributions across spatial scales adopted from (Pearson & Dawson, 2003 ; Soberón 2007 , 2010 ; Hortal et al ., 2010 )
正如物种是生物学的核心概念, 生态位是生态学中最基本的概念。给生态位下定义的人较多, 最具代表性的是Joseph Grinnell, Charles Sutherland Elton和George Evelyn Hutchinson三人, 后人将他们所给的定义分别称为“空间生态位(spatial niche)”、“功能生态位(functional niche)”和“多维超体积生态位(n-dimentional hypervolume niche)”。Grinnell (1917)的表述为“The response of species to a given set of variables”, 可以认为是维持物种生存的“非生物因素”, 强调的是宏观尺度的气候生态空间; Elton(1927)的表述为“The impact of species in the environment”, 可以认为是物种在群落中的机能和地位, 强调微观尺度物种间的营养关系; Hutchinson(1957)在Grinnell生态位基础上, 提出生态位是指一个超维的空间, 在这个空间中种群可以不断延续, 形成影响较大的Hutchinson生态位概念, 即“A hyper-volume in the multidimensional space of ecological variables, within which a species can maintain a viable population”, 这里也就引出了基础生态位(fundamental niche)的概念。简而言之, 基础生态位就是维持物种生存所必须的所有非生物条件的总和(相当于图1 中的A环), 在这些条件下物种种群表现为正增长, 它反映了一个物种的生理需求, 具有生物学意义。然而由于不同物种间的相互作用, 一个物种的基础生态位通常不能完全表现或反映在特定的地理区域, 因此Hutchinson使用现实生态位(realized niche)来表述基础生态位在这些地理区域的反映(Hutchinson, 1957 )。
2 生态位模型构建的基本原理
2.1 地理空间和生态空间的结合
广义生态位模型的构建包含两种截然不同的思路: 一种为直接的机理性方法(mechanistic approach), 另一种为间接的相关性方法(correlative approach), 前者基于物种对环境耐受性的生理参数来判断物种的潜在分布, 后者基于物种分布点所关联的环境参数来推算物种的分布(Pearson, 2007 )。在实际操作中, 第一种方法要求的物种生理参数的测定是一项繁重的任务, 自然界中多数物种的生理参数是未知的。而随着全球性物种分布数据库的共享以及地理信息系统技术的快速发展, 间接方法的生态位模型应运而生(McCormack et al. , 2010 )。对于多数物种分布的预测来说, 间接方法提供了一种简单易用的途径。本文如未加说明, 所指生态位模型均为间接方法的生态位模型。然而, 生态位的概念是生态位模型的理论基础, 对各种生态位概念的理解将会影响生态位模型构建的合理性, 模型构建的各个环节需要谨慎, 不恰当的方法容易得出错误的结论(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 )。
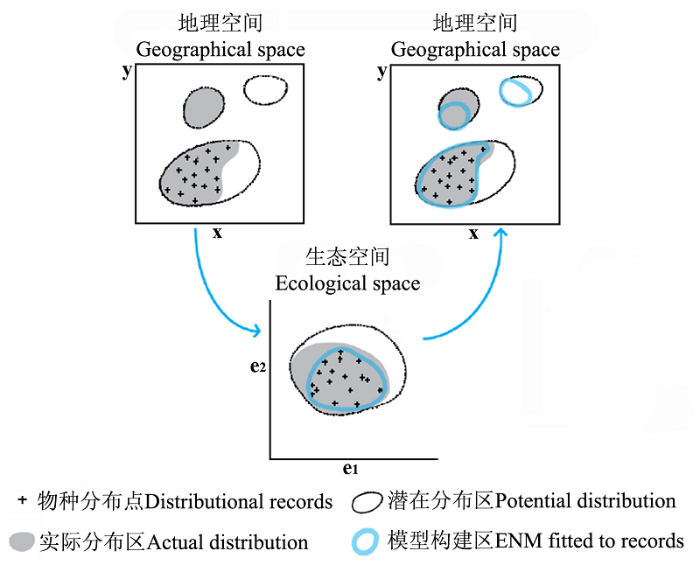
生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间。如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 )。在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 )。另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 )。后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 )。参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法。
图3 示生态位模型将地理空间和生态空间二者结合引自(Pearson, 2007 )
Fig. 3 Ecological niche modeling (ENM) building up the fridge of geographical space and ecological space adopted from (Pearson, 2007 )
2.2 模型构建的前提
生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态。是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的。这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用。在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的。这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的。只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性。近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b )。
在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 )。而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题。文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化。在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性。
3 生态位模型与生态位
3.1 Grinnell生态位和Elton生态位
在实际操作中, Grinnell生态位和Elton生态位均已反映在生态位模型中。该两种生态位与模型构建的环节密切相关, 主要取决于环境变量类型的选择、所采用环境变量的分辨率以及物种本身的迁移能力三个方面(Soberón, 2007 , 2010 )。
在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 )。在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 )。物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 )。理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 )。然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 )。
3.2 现实生态位、潜在生态位和基础生态位
Hutchinson(1957)使用现实生态位来描述基础生态位在地理空间的反映, 这里的限制因素主要指物种间的相互作用。通常来讲, 基础生态位大于现实生态位, 这是由于物种间的相互作用和物种的迁移能力一定程度上影响了基础生态位在地理空间的反映(图1 ), 因此一些物种并未在其生态适宜地区出现(Holt, 2003 ); 另一方面, 由于汇种群的影响, 一些物种会暂时迁移到不适宜生存的区域, 如迁徙的鸟类, 这是现实生态位大于基础生态位的情况(Pulliam, 2000 )。因此, 在构建生态位模型时需要注意是否包含了汇种群的分布点(图1 )。
在某一历史时期, 可获取的生态空间是有限的。Jackson和Overpeck(2000)使用潜在生态位(potential niche)来描述基础生态位在有限生态空间中的反映, 即基础生态位和可获取生态空间的交集(Colwell & Rangel, 2009 )。潜在生态位体现了特定历史时期内基础生态位在生态空间中的可获取性, 物种间的相互作用及物种的迁移能力限制了其在特定地理空间的反映, 所以说现实生态位是潜在生态位的一部分(Sillero, 2011 )。
在实际操作中, 生态位模型所模拟的是现实生态位, 而不是基础生态位。然而, 由于各模型算法所采用“不存在”数据(absence records)不同, 所模拟的现实生态位亦是有差别的(Soberón, 2007 ; Sillero, 2011 )。例如, 有的生态位模型同时使用存在数据(presence records)和不存在数据来模拟, 有的使用假设不存在(pseudo-absence)来替代物种的不存在数据, 所得到的模拟结果是不同的。另外, 机理性生态位模型反映的是特定生态空间中的基础生态位, 即潜在生态位, 并不是基础生态位(Sillero, 2011 )。
4 生态位模型在生物多样性保护中的应用
4.1 物种生态需求的分析
地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性。例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到。生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具。特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要。近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 )。
4.2 发现未知物种和种群
一直以来, 地球上物种的确切数目及其分布困扰着生物学家。前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 )。生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足。另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足。这方面的应用值得分类学者尝试。
4.3 自然保护区的选择和设计
在自然保护区选择和设计时, 可应用生态位模型对物种实际分布范围进行模拟, 从而使自然保护区充分发挥功能, 减少不必要的开支。为了得到较为真实的物种分布, 在模型的构建上通常采取较为保守的策略, 需充分考虑影响物种分布的限制因素, 对模型的运算设定较多的限制(Araújo & Peterson, 2012 )。生态位模型的输出结果是一个对分布可能性大小的估测, 在后期校正时, 还应考虑物种间的相互作用和物种的迁移能力等因素(Pearson, 2007 )。
4.4 物种入侵风险的评价
生物入侵是生物多样性丧失的主要原因之一, 生态位模型可用于对入侵物种潜在分布的模拟, 这一领域的应用较为广泛。通常是基于物种原产地的分布数据来构建模型, 然后用于判断该物种在入侵地的潜在分布(图3 ), 为该物种的防范和监控提供参考。因此, 模型构建所选用的物种分布点必须能够反映该物种的分布范围, 所选环境变量也必须对该物种的分布具有较大的限制作用, 这样才能提高模型的可转移性(Araújo & Peterson, 2012 ), 从而准确模拟物种在入侵地的分布范围。这一应用的理论基础是生态位的保守性, 只有在此前提下进行模型的转移和预测才具有合理性。如果某物种在原产地的分布数据比较难获取, 那么也可以基于其在入侵地的分布数据来构建模型, 进行潜在分布范围的模拟(Zhu et al ., 2007 )。
4.5 气候变化对生物分布的影响
生态位模型可模拟未来气候条件下物种的潜在分布。基本思路是: 基于已有的物种分布数据和当前的气候参数构建模型, 将其投射至将来的气候条件下, 来模拟未来物种潜在分布范围。目前, 基于联合国政府间气候变化委员会(IPCC)的评估报告, 加拿大、英国和澳大利亚等国的研究机构相继模拟出了未来不同时期的气候参数, 为未来物种潜在分布的模拟提供了基础(Hijmans et al ., 2005 )。在这类应用中, 强调了气候因素对物种分布的限制作用, 忽略了大尺度下物种间的相互作用和物种的迁移能力(Peterson et al ., 2002 )。这一应用的理论基础仍是生态位的保守性, 只有在此前提下, 模拟得到的未来气候条件下物种的分布才有价值(Petitpierre et al ., 2012 )。在大尺度范围下, 全球气候变化对北美和欧洲的鸟类和植物被广泛研究(e.g. Thuiller et al . 2005 ), 我国这方面的研究需要加强。
4.6 近缘物种生态位保守性
生态位的保守性对于物种的界定、入侵的模拟、气候变化对物种分布的影响以及生物多样性等方面的研究具有重要意义。生态位的保守性是一个敏感的话题, 近年引起了激烈的讨论。引发这场讨论的是Peterson等(1999)发表在Science 上的文章, 截至2013年1月1日该篇文章的引用已经超过了680次。Peterson等(1999)采用GARP模型对分布于墨西哥湾特万特佩克地峡(Isthmus of Tehuantepec)两侧的姊妹种(包括哺乳类、鸟类和蝴蝶)进行了模拟, 发现异域种化的姊妹种之间能够很好地相互模拟, 例如基于Atthis heloisa (Lesson & Delattre, 1839)的分布数据构建的模型能够很好地模拟其姊妹种A. ellioti (Ridgway, 1878)的分布; 同样, 基于A. ellioti 的分布记录的模型也能够很好地模拟A. heloisa 的种群分布记录。但是, 高级阶元间的保守性相对较差。目前学者们普遍认可的结论是: 生态位是进化的, 但是在较短的进化历史内, 是倾向保守的, 生态位的保守性与系统发育关系并不具有一致性(Wiens & Graham, 2005 ; Peterson, 2011 )。如果我们试图从进化上来验证物种间生态位的保守性, 那么首先要确认物种间的近缘关系, 它们是姊妹种, 或者是同一属, 或者是系统发育树上的近缘分支, 基于此来探讨生态位的保守与否才有意义。
4.7 生态位分化与物种界定
物种的界定是进化生物学的核心争论问题, 分歧的核心是对物种定义的不同。近年来, 生态位的分化被越来越多地用于物种的界定上(e.g. Leaché et al ., 2009 )。物种分化和生态位分化的步调可能是一致的(即生态位的分化性), 但也可能是不完全一致的(即生态位的保守性)。在进化的历史过程中, 新物种的产生总是倾向于保留其祖先的生态位, 然而, 新物种在适应新的环境时, 会发生一些适应性的基因突变, 这些突变可能导致生态位的分化, 反过来又促进物种的分化。总之, 生态位分化的依据为物种的界定提供了一条途径, 越来越多的学者呼吁在物种界定时考虑生态因素(Rissler & Apodaca, 2007 )。基于生态位模型的生态位分化研究也越来越细, 从物种间的生态空间分析, 到物种间的生态位测试, 如Warren等(2008, 2010)等开发的生态位等值性测试(niche equivalency or identity test)和相似性测试(similarity or background test)已经被越来越多的学者采用, 并且与DNA条形码结合用于新物种的描述(Hawlitschek et al ., 2011 )。
生态位模型用于物种的界定似乎对一些特殊类群更有利, 特别是一些迁移能力较弱、在不为人知的地区分布的特有种(Raxworthy et al ., 2007 ; Zhu et al. , 2013)。我国学者将生态位等值性测试运用在中国林蛙复合体的物种界定上(Zhou et al ., 2012 )。在进行生态位等值性和相似性测试时, 需要将二者结合起来, 并对物种的分布范围给出一个合理的解释, 否则单一的生态位等值性测试不足以说明物种基础生态位的分化(Warren et al ., 2010 ; Peterson & Soberón, 2012 )。
5 小结
影响物种分布的因素主要有非生物的气候因素、物种间的相互作用以及物种的迁移能力3种, 不同的因素在不同的空间尺度下起主要作用。生态位模型利用物种分布点所关联的环境变量去推算物种的生态需求, 模拟物种的分布。在实际操作中, Grinnell生态位和Elton生态位均反映在了模型构建中, 这取决于环境变量类型的选择、所采用环境变量的分辨率以及物种本身的迁移能力。生态位模型所模拟的是现实生态位或潜在生态位, 并不是基础生态位。基础生态位反映了一个物种的生理需求, 具有生物学意义; 潜在生态位体现了物种基础生态位在特定历史时期生态空间的可获取性, 物种间的相互作用及物种的迁移能力限制了其在特定地理空间的反映。在生物多样性的保护中, 生态位模型已得到了广泛的应用。只有对模型理论有了深入的理解, 才能够促进生态位模型在这些生物多样性保护领域得到更为合理的运用和发展。
致谢
感谢美国Kansas大学A. Townsend Peterson教授在生态位模型学习中的指导和帮助。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Araújo MB Peterson AT 2012 ) Uses and misuses of bioclimatic envelope modelling.
Ecology , 93 , 1527 -1539 .
[本文引用: 5]
[2]
Bourg NA McShea WJ Gill DE 2005 ) Putting a CART before the search: successful habitat prediction for a rare forest herb.
Ecology , 86 , 2793 -2804 .
[本文引用: 1]
[3]
Broennimann O Treier UA Müller-Scharer H Thuiller W Peterson AT Guisan A 2007 ) Evidence of climatic niche shift during biological invasion.
Ecology Letters , 10 , 701 -709 .
[本文引用: 3]
[4]
Brown JH 1995 ) Macroecology . University of Chicago Press , Chicago .
[本文引用: 1]
[5]
Colwell RK Rangel TF 2009 ) Hutchinson’s duality: the once and future niche
. Proceedings of the National Academy of Sciences,USA , 106 , 19651 -19658 .
[本文引用: 1]
[6]
Dorji S Vernes K Rajaratnam R 2011 ) Habitat correlates of the red panda in the temperate forests of Bhutan.
PLOS ONE , 6 , e26483 .
[本文引用: 1]
[7]
Elton CS 1927 ) Animal Ecology
. Sidgwick and Jackson, London.
[8]
Etterson JR Shaw RG 2001 ) Constraint to adaptive evolution in response to global warming.
Science , 294 , 151 -154 .
[本文引用: 1]
[9]
Gaston KJ 2003 ) The Structure and Dynamics of Geographic Ranges . Oxford University Press , Oxford .
[本文引用: 1]
[10]
Grinnell J 1917 ) The niche-relationships of the California Thrasher.
The Auk , 34 , 427 -433 .
[11]
Guisan A Thuiller W 2005 ) Predicting species distribution: offering more than simple habitat models.
Ecology Letters , 8 , 993 -1009 .
[本文引用: 1]
[12]
Hawlitschek O Porch N Hendrich L Balke M 2011 ) Ecological niche modelling and nDNA sequencing support a new, morphologically cryptic beetle species unveiled by DNA Barcoding.
PLOS ONE , 6 , e16662 .
[本文引用: 1]
[13]
Hijmans RJ Cameron SE Parra JL Jones PG Jarvis A 2005 ) Very high resolution interpolated climate surfaces for global land areas.
International Journal of Climatology , 25 , 1965 -1978 .
[本文引用: 2]
[14]
Hirzel AH Hausser J Chessel D Perrin N 2002 ) Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data ?
Ecology , 83 , 2027 -2036 .
[本文引用: 1]
[15]
Holt RD 2003 ) On the evolutionary ecology of species’ ranges.
Evolutionary Ecology Research , 5 , 159 -178 .
[本文引用: 1]
[16]
Hortal J Roura-Pascual N Sanders NJ Rahbek C 2010 ) Understanding (insect) species distributions across spatial scales.
Ecography , 33 , 51 -53 .
[本文引用: 4]
[17]
Hutchinson GE 1957 ) Concluding remarks.
Cold Spring Harbor Symposium on Quantitative Biology , 22 , 415 -427 .
[本文引用: 1]
[18]
Irfan-Ullah M Amarnath G Murthy MSR Peterson AT 2007 ) Mapping the geographic distribution of Aglaia bourdillonii Gamble (Meliaceae), an endemic and threatened plant, using ecological niche modeling.
Plant Conservation and Biodiversity , 16 , 1917 -1925 .
[本文引用: 1]
[19]
Jackson ST Overpeck JT 2000 ) Responses of plant populations and communities to environmental changes of the late Quaternary.
Paleobiology , 26 , 194 -220 .
[20]
Leaché AD Koo MS Spencer CL Papenfuss TJ Fisher RN McGuire JA 2009 ) Quantifying ecological, morphological, and genetic variation to delimit species in the coast horned lizard species complex (Phrynosoma)
. Proceedings of the National Academy of Sciences, USA , 106 , 12418 -12423 .
[本文引用: 1]
[21]
Luoto M Heikkinen RK Pöyry J Saarinen K 2006 ) Determinants of the biogeographical distribution of butterflies in boreal regions.
Journal of Biogeography , 33 , 1764 -1778 .
[本文引用: 1]
[22]
Mackey BG Lindenmayer DB 2001 ) Towards a hierarchical framework for modelling the spatial distribution of animals.
Journal of Biogeography , 28 , 1147 -1166 .
[本文引用: 1]
[23]
McCormack JE Zellmer AJ Knowles LL 2010 ) Does niche divergence accompany allopatric divergence in Aphelocoma jays as predicted under ecological speciation? Insights from tests with niche models.
Evolution , 64 , 1231 -1244 .
[本文引用: 2]
[24]
Medley KA 2010 ) Niche shifts during the global invasion of the Asian tiger mosquito, Aedes albopictus Skuse (Culicidae), revealed by reciprocal distribution models.
Global Ecology and Biogeography , 19 , 122 -133 .
[本文引用: 3]
[25]
Menon S Choudhury BI Khan ML Peterson AT 2010 ) Ecological niche modeling and local knowledge predict new populations of Gymnocladus assamicus, a critically endangered tree species.
Endangered Species Research , 11 , 175 -181 .
[本文引用: 1]
[26]
Owens HL Bentley AC Peterson AT 2011 ) Predicting suitable environments and potential occurrences for coelacanths (Latimeria spp.).
Biodiversity and Conservation , 21 , 577 -587 .
[本文引用: 1]
[27]
Pearson RG 2007 ) Species’ distribution modeling for conservation educators and practitioners. Synthesis. American Museum of Natural History.
.
URL
[本文引用: 4]
[28]
Pearson RG Dawson TP 2003 ) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful ?
Global Ecology and Biogeography , 12 , 361 -371 .
[本文引用: 5]
[29]
Peterson AT 2011 ) Ecological niche conservatism: a time-structured review of evidence.
Journal of Biogeography , 38 , 817 -827 .
[30]
Peterson AT Nakazawa Y 2008 ) Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri.
Global Ecology and Biogeography , 17 , 135 -144 .
[本文引用: 1]
[31]
Peterson AT Navarro-Sigüenza AG 2009 ) Making biodiversity discovery more efficient: an exploratory test using Mexican birds.
Zootaxa , 2246 , 58 -66 .
[本文引用: 1]
[32]
Peterson AT Ortega-Huerta MA Bartley J Sánchez-Cordero V Soberón J Buddemeier RH Stockwell DRB 2002 ) Future projections for Mexican faunas under global climate change scenarios
. Nature , 416 , 626 -629 .
[本文引用: 1]
[33]
Peterson AT Soberón J 2012 ) Integrating fundamental concepts of ecology, biogeography, and sampling into effective ecological niche modeling and species distribution modeling.
Plant Biosystems , 146 , 789 -796 .
[本文引用: 3]
[34]
Peterson AT Soberón J Pearson RG Anderson RP Nakamura M Martínez-Meyer E Araújo MB 2011 ) Ecological Niches and Geographical Distributions . Princeton University Press, New Jersey .
[本文引用: 3]
[35]
Peterson AT Soberón J Sánchez-Cordero V 1999 ) Conservatism of ecological niches in evolutionary time.
Science , 285 , 1265 -1267 .
[36]
Petitpierre B Kueffer C Broennimann O Randin C Daehler C Guisan A 2012 ) Climatic niche shifts are rare among terrestrial plant invaders.
Science , 335 , 1344 -1348 .
[本文引用: 1]
[37]
Pulliam HR 2000 ) On the relationship between niche and distribution.
Ecology Letters , 3 , 349 -361 .
[本文引用: 3]
[38]
Raxworthy CJ Ingram CM Rabibisoa N Pearson RG 2007 ) Applications of ecological niche modeling for species delimitation: a review and empirical evaluation using day geckos (Phelsuma) from Madagascar.
Systematic Biology , 56 , 907 -923 .
[本文引用: 2]
[39]
Raxworthy CJ Martínez-Meyer E Horning N Nussbaum RA Schneider GE Ortega-Huerta MA Peterson AT 2003 ) Predicting distributions of known and unknown reptile species in Madagascar.
Nature , 426 , 837 -841 .
[本文引用: 1]
[40]
Rissler LJ Apodaca JJ 2007 ) Adding more ecology into species delimitation: ecological niche models and phylogeography help define cryptic species in the Black Salamander (Aneides flavipunctatus).
Systematic Biology , 56 , 924 -942 .
[本文引用: 1]
[41]
Robinson LM Elith J Hobday AJ Pearson RG Kendall BE Possingham HP Richardson AJ 2011 ) Pushing the limits in marine species distribution modelling: lessons from the land present challenges and opportunities.
Global Ecology and Biogeography , 20 , 789 -802 .
[本文引用: 1]
[42]
Rödder D Lötters S 2009 ) Niche shift versus niche conservatism? Climatic characteristics of the native and invasive ranges of the Mediterranean house gecko (Hemidactylus turcicus).
Global Ecology and Biogeography , 18 , 674 -687 .
[本文引用: 2]
[43]
Saupe E Barve V Myers C Soberón J Barve N Hensz C Peterson AT Owens HL Lira-Noriega A 2012 ) Variation in niche and distribution model performance: the need for a priori assessment of key causal factors.
Ecological Modelling , 237 , 11 -22 .
[本文引用: 1]
[44]
Sillero N 2011 ) What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods.
Ecological Modelling , 222 , 1343 -1346 .
[本文引用: 5]
[45]
Soberón J 2007 ) Grinnellian and Eltonian niches and geographic distributions of species.
Ecology Letters , 10 , 1115 -1123 .
[本文引用: 8]
[46]
Soberón J Peterson AT 2005 ) Interpretation of models of fundamental ecological niches and species' distributional areas.
Biodiversity Informatics , 2 , 1 -10 .
[本文引用: 3]
[47]
Soberón JM 2010 ) Niche and area of distribution modeling: a population ecology perspective.
Ecography , 33 , 159 -167 .
[本文引用: 6]
[48]
Svenning JC Skov F 2004 ) Limited filling of the potential range in European tree species.
Ecology Letters , 7 , 565 -573 .
[本文引用: 2]
[49]
Thuiller W Lavorel S Araújo MB Sykes MT Prentice IC 2005 ) Climate change threats to plant diversity in Europe.
Proceedings of the National Academy of Sciences, USA , 102 , 8245 -8250 .
[本文引用: 1]
[50]
Warren DL 2012 ) In defense of niche modeling.
Trends in Ecology and Evolution , 27 , 497 -500 .
[本文引用: 3]
[51]
Warren DL Glor RE Turelli M 2008 ) Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution.
Evolution , 62 , 2868 -2883 .
[52]
Warren DL Glor RE Turelli M 2010 ) ENMTools: a toolbox for comparative studies of environmental niche models.
Ecography , 33 , 607 -611 .
[本文引用: 1]
[53]
Whittaker RJ Araújo MB Jepson P Ladle RJ Watson JEM Willis KJ 2005 ) Conservation biogeography: assessment and prospect.
Diversity and Distributions , 11 , 3 -23 .
[本文引用: 2]
[54]
Willis KJ Whittaker RJ 2002 ) Species diversity―scale matters.
Science , 295 , 1245 -1248 .
[本文引用: 1]
[55]
Wiens JJ Graham CH 2005 ) Niche conservatism: integrating evolution, ecology, and conservation biology.
Annual Review of Ecology, Evolution, and Systematics , 36 , 519 -539 .
[本文引用: 1]
[56]
Zhou WW Wen Y Fu JZ Xu YB Jin JQ Ding L Min MS Che J Zhang YP 2012 ) Speciation in the Rana chensinensis species complex and its relationship to the uplift of the Qinghai-Tibetan Plateau.
Molecular Ecology , 21 , 960 -973 .
[本文引用: 1]
[57]
Zhu G Bu W Gao Y Liu G 2012 a) Potential geographic distribution of Brown Marmorated Stink Bug invasion (Halyomorpha halys).
PLOS ONE , 7 , e31246 .
[本文引用: 2]
[58]
[59]
Zhu GP Petersen MJ Bu WJ 2012 b) Selecting biological meaningful environmental dimensions of low discrepancy among ranges to predict potential distribution of bean plataspid invasion.
PLOS ONE , 7 , e46247 .
[本文引用: 3]
[60]
Zhu L Sun OJ Sang WG Li ZY Ma KP 2007 ) Predicting the spatial distribution of an invasive plant species (Eupatorium adenophorum) in China.
Landscape Ecology , 22 , 1143 -1154 .
Uses and misuses of bioclimatic envelope modelling.
5
2012
... 近年来, 生态位模型在生物多样性保护的多个领域得到开拓和应用, 被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布的影响及传染病空间传播的研究中.但是, 在模型运用时却得出了一些不符合实际的结论, 如入侵物种的生态位迁移等(Broennimann et al ., 2007 ; Medley, 2010 ), 故一些学者对生态位模型提出了质疑.这些不当之处或多或少是由于对模型构建的目的认识模糊, 对生态位的概念和生态位模型的原理, 或者模型软件算法理解不透彻造成的(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ).为了让读者更系统地理解和正确地应用生态位模型, 我们梳理了生态位与物种分布的关系, 结合生态位概念介绍了生态位模型的基本原理, 从物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘种生态位保守性及基于生态位分化的物种界定等方面阐述了它在生物多样性保护中的应用. ...
... 广义生态位模型的构建包含两种截然不同的思路: 一种为直接的机理性方法(mechanistic approach), 另一种为间接的相关性方法(correlative approach), 前者基于物种对环境耐受性的生理参数来判断物种的潜在分布, 后者基于物种分布点所关联的环境参数来推算物种的分布(Pearson, 2007 ).在实际操作中, 第一种方法要求的物种生理参数的测定是一项繁重的任务, 自然界中多数物种的生理参数是未知的.而随着全球性物种分布数据库的共享以及地理信息系统技术的快速发展, 间接方法的生态位模型应运而生(McCormack et al. , 2010 ).对于多数物种分布的预测来说, 间接方法提供了一种简单易用的途径.本文如未加说明, 所指生态位模型均为间接方法的生态位模型.然而, 生态位的概念是生态位模型的理论基础, 对各种生态位概念的理解将会影响生态位模型构建的合理性, 模型构建的各个环节需要谨慎, 不恰当的方法容易得出错误的结论(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ). ...
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
... 在自然保护区选择和设计时, 可应用生态位模型对物种实际分布范围进行模拟, 从而使自然保护区充分发挥功能, 减少不必要的开支.为了得到较为真实的物种分布, 在模型的构建上通常采取较为保守的策略, 需充分考虑影响物种分布的限制因素, 对模型的运算设定较多的限制(Araújo & Peterson, 2012 ).生态位模型的输出结果是一个对分布可能性大小的估测, 在后期校正时, 还应考虑物种间的相互作用和物种的迁移能力等因素(Pearson, 2007 ). ...
... 生物入侵是生物多样性丧失的主要原因之一, 生态位模型可用于对入侵物种潜在分布的模拟, 这一领域的应用较为广泛.通常是基于物种原产地的分布数据来构建模型, 然后用于判断该物种在入侵地的潜在分布(图3 ), 为该物种的防范和监控提供参考.因此, 模型构建所选用的物种分布点必须能够反映该物种的分布范围, 所选环境变量也必须对该物种的分布具有较大的限制作用, 这样才能提高模型的可转移性(Araújo & Peterson, 2012 ), 从而准确模拟物种在入侵地的分布范围.这一应用的理论基础是生态位的保守性, 只有在此前提下进行模型的转移和预测才具有合理性.如果某物种在原产地的分布数据比较难获取, 那么也可以基于其在入侵地的分布数据来构建模型, 进行潜在分布范围的模拟(Zhu et al ., 2007 ). ...
Putting a CART before the search: successful habitat prediction for a rare forest herb.
1
2005
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
Evidence of climatic niche shift during biological invasion.
3
2007
... 近年来, 生态位模型在生物多样性保护的多个领域得到开拓和应用, 被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布的影响及传染病空间传播的研究中.但是, 在模型运用时却得出了一些不符合实际的结论, 如入侵物种的生态位迁移等(Broennimann et al ., 2007 ; Medley, 2010 ), 故一些学者对生态位模型提出了质疑.这些不当之处或多或少是由于对模型构建的目的认识模糊, 对生态位的概念和生态位模型的原理, 或者模型软件算法理解不透彻造成的(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ).为了让读者更系统地理解和正确地应用生态位模型, 我们梳理了生态位与物种分布的关系, 结合生态位概念介绍了生态位模型的基本原理, 从物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘种生态位保守性及基于生态位分化的物种界定等方面阐述了它在生物多样性保护中的应用. ...
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
... 在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 ).而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题.文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化.在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性. ...
1
1995
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
Hutchinson’s duality: the once and future niche
1
2009
... 在某一历史时期, 可获取的生态空间是有限的.Jackson和Overpeck(2000)使用潜在生态位(potential niche)来描述基础生态位在有限生态空间中的反映, 即基础生态位和可获取生态空间的交集(Colwell & Rangel, 2009 ).潜在生态位体现了特定历史时期内基础生态位在生态空间中的可获取性, 物种间的相互作用及物种的迁移能力限制了其在特定地理空间的反映, 所以说现实生态位是潜在生态位的一部分(Sillero, 2011 ). ...
Habitat correlates of the red panda in the temperate forests of Bhutan.
1
2011
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
Constraint to adaptive evolution in response to global warming.
1
2001
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
1
2003
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
The niche-relationships of the California Thrasher.
1917
Predicting species distribution: offering more than simple habitat models.
1
2005
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
Ecological niche modelling and nDNA sequencing support a new, morphologically cryptic beetle species unveiled by DNA Barcoding.
1
2011
... 物种的界定是进化生物学的核心争论问题, 分歧的核心是对物种定义的不同.近年来, 生态位的分化被越来越多地用于物种的界定上(e.g. Leaché et al ., 2009 ).物种分化和生态位分化的步调可能是一致的(即生态位的分化性), 但也可能是不完全一致的(即生态位的保守性).在进化的历史过程中, 新物种的产生总是倾向于保留其祖先的生态位, 然而, 新物种在适应新的环境时, 会发生一些适应性的基因突变, 这些突变可能导致生态位的分化, 反过来又促进物种的分化.总之, 生态位分化的依据为物种的界定提供了一条途径, 越来越多的学者呼吁在物种界定时考虑生态因素(Rissler & Apodaca, 2007 ).基于生态位模型的生态位分化研究也越来越细, 从物种间的生态空间分析, 到物种间的生态位测试, 如Warren等(2008, 2010)等开发的生态位等值性测试(niche equivalency or identity test)和相似性测试(similarity or background test)已经被越来越多的学者采用, 并且与DNA条形码结合用于新物种的描述(Hawlitschek et al ., 2011 ). ...
Very high resolution interpolated climate surfaces for global land areas.
2
2005
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... 生态位模型可模拟未来气候条件下物种的潜在分布.基本思路是: 基于已有的物种分布数据和当前的气候参数构建模型, 将其投射至将来的气候条件下, 来模拟未来物种潜在分布范围.目前, 基于联合国政府间气候变化委员会(IPCC)的评估报告, 加拿大、英国和澳大利亚等国的研究机构相继模拟出了未来不同时期的气候参数, 为未来物种潜在分布的模拟提供了基础(Hijmans et al ., 2005 ).在这类应用中, 强调了气候因素对物种分布的限制作用, 忽略了大尺度下物种间的相互作用和物种的迁移能力(Peterson et al ., 2002 ).这一应用的理论基础仍是生态位的保守性, 只有在此前提下, 模拟得到的未来气候条件下物种的分布才有价值(Petitpierre et al ., 2012 ).在大尺度范围下, 全球气候变化对北美和欧洲的鸟类和植物被广泛研究(e.g. Thuiller et al . 2005 ), 我国这方面的研究需要加强. ...
Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data ?
1
2002
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
On the evolutionary ecology of species’ ranges.
1
2003
... Hutchinson(1957)使用现实生态位来描述基础生态位在地理空间的反映, 这里的限制因素主要指物种间的相互作用.通常来讲, 基础生态位大于现实生态位, 这是由于物种间的相互作用和物种的迁移能力一定程度上影响了基础生态位在地理空间的反映(图1 ), 因此一些物种并未在其生态适宜地区出现(Holt, 2003 ); 另一方面, 由于汇种群的影响, 一些物种会暂时迁移到不适宜生存的区域, 如迁徙的鸟类, 这是现实生态位大于基础生态位的情况(Pulliam, 2000 ).因此, 在构建生态位模型时需要注意是否包含了汇种群的分布点(图1 ). ...
Understanding (insect) species distributions across spatial scales.
4
2010
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
... 不同影响因素在不同的空间尺度下作用于物种的分布修改自(Pearson & Dawson, 2003 ; Soberón, 2007 , 2010 ; Hortal et al ., 2010 ) ...
... Relative importance of factors affecting species distributions across spatial scales adopted from (Pearson & Dawson, 2003 ; Soberón 2007 , 2010 ; Hortal et al ., 2010 ) ...
Concluding remarks.
1
1957
... 正如物种是生物学的核心概念, 生态位是生态学中最基本的概念.给生态位下定义的人较多, 最具代表性的是Joseph Grinnell, Charles Sutherland Elton和George Evelyn Hutchinson三人, 后人将他们所给的定义分别称为“空间生态位(spatial niche)”、“功能生态位(functional niche)”和“多维超体积生态位(n-dimentional hypervolume niche)”.Grinnell (1917)的表述为“The response of species to a given set of variables”, 可以认为是维持物种生存的“非生物因素”, 强调的是宏观尺度的气候生态空间; Elton(1927)的表述为“The impact of species in the environment”, 可以认为是物种在群落中的机能和地位, 强调微观尺度物种间的营养关系; Hutchinson(1957)在Grinnell生态位基础上, 提出生态位是指一个超维的空间, 在这个空间中种群可以不断延续, 形成影响较大的Hutchinson生态位概念, 即“A hyper-volume in the multidimensional space of ecological variables, within which a species can maintain a viable population”, 这里也就引出了基础生态位(fundamental niche)的概念.简而言之, 基础生态位就是维持物种生存所必须的所有非生物条件的总和(相当于图1 中的A环), 在这些条件下物种种群表现为正增长, 它反映了一个物种的生理需求, 具有生物学意义.然而由于不同物种间的相互作用, 一个物种的基础生态位通常不能完全表现或反映在特定的地理区域, 因此Hutchinson使用现实生态位(realized niche)来表述基础生态位在这些地理区域的反映(Hutchinson, 1957 ). ...
Mapping the geographic distribution of Aglaia bourdillonii Gamble (Meliaceae), an endemic and threatened plant, using ecological niche modeling.
1
2007
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
Responses of plant populations and communities to environmental changes of the late Quaternary.
2000
Quantifying ecological, morphological, and genetic variation to delimit species in the coast horned lizard species complex (Phrynosoma)
1
2009
... 物种的界定是进化生物学的核心争论问题, 分歧的核心是对物种定义的不同.近年来, 生态位的分化被越来越多地用于物种的界定上(e.g. Leaché et al ., 2009 ).物种分化和生态位分化的步调可能是一致的(即生态位的分化性), 但也可能是不完全一致的(即生态位的保守性).在进化的历史过程中, 新物种的产生总是倾向于保留其祖先的生态位, 然而, 新物种在适应新的环境时, 会发生一些适应性的基因突变, 这些突变可能导致生态位的分化, 反过来又促进物种的分化.总之, 生态位分化的依据为物种的界定提供了一条途径, 越来越多的学者呼吁在物种界定时考虑生态因素(Rissler & Apodaca, 2007 ).基于生态位模型的生态位分化研究也越来越细, 从物种间的生态空间分析, 到物种间的生态位测试, 如Warren等(2008, 2010)等开发的生态位等值性测试(niche equivalency or identity test)和相似性测试(similarity or background test)已经被越来越多的学者采用, 并且与DNA条形码结合用于新物种的描述(Hawlitschek et al ., 2011 ). ...
Determinants of the biogeographical distribution of butterflies in boreal regions.
1
2006
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
Towards a hierarchical framework for modelling the spatial distribution of animals.
1
2001
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
Does niche divergence accompany allopatric divergence in Aphelocoma jays as predicted under ecological speciation? Insights from tests with niche models.
2
2010
... 广义生态位模型的构建包含两种截然不同的思路: 一种为直接的机理性方法(mechanistic approach), 另一种为间接的相关性方法(correlative approach), 前者基于物种对环境耐受性的生理参数来判断物种的潜在分布, 后者基于物种分布点所关联的环境参数来推算物种的分布(Pearson, 2007 ).在实际操作中, 第一种方法要求的物种生理参数的测定是一项繁重的任务, 自然界中多数物种的生理参数是未知的.而随着全球性物种分布数据库的共享以及地理信息系统技术的快速发展, 间接方法的生态位模型应运而生(McCormack et al. , 2010 ).对于多数物种分布的预测来说, 间接方法提供了一种简单易用的途径.本文如未加说明, 所指生态位模型均为间接方法的生态位模型.然而, 生态位的概念是生态位模型的理论基础, 对各种生态位概念的理解将会影响生态位模型构建的合理性, 模型构建的各个环节需要谨慎, 不恰当的方法容易得出错误的结论(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ). ...
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
Niche shifts during the global invasion of the Asian tiger mosquito, Aedes albopictus Skuse (Culicidae), revealed by reciprocal distribution models.
3
2010
... 近年来, 生态位模型在生物多样性保护的多个领域得到开拓和应用, 被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布的影响及传染病空间传播的研究中.但是, 在模型运用时却得出了一些不符合实际的结论, 如入侵物种的生态位迁移等(Broennimann et al ., 2007 ; Medley, 2010 ), 故一些学者对生态位模型提出了质疑.这些不当之处或多或少是由于对模型构建的目的认识模糊, 对生态位的概念和生态位模型的原理, 或者模型软件算法理解不透彻造成的(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ).为了让读者更系统地理解和正确地应用生态位模型, 我们梳理了生态位与物种分布的关系, 结合生态位概念介绍了生态位模型的基本原理, 从物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘种生态位保守性及基于生态位分化的物种界定等方面阐述了它在生物多样性保护中的应用. ...
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
... 在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 ).而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题.文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化.在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性. ...
Ecological niche modeling and local knowledge predict new populations of Gymnocladus assamicus, a critically endangered tree species.
1
2010
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
Predicting suitable environments and potential occurrences for coelacanths (Latimeria spp.).
1
2011
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
Species’ distribution modeling for conservation educators and practitioners. Synthesis. American Museum of Natural History.
4
2007
... 广义生态位模型的构建包含两种截然不同的思路: 一种为直接的机理性方法(mechanistic approach), 另一种为间接的相关性方法(correlative approach), 前者基于物种对环境耐受性的生理参数来判断物种的潜在分布, 后者基于物种分布点所关联的环境参数来推算物种的分布(Pearson, 2007 ).在实际操作中, 第一种方法要求的物种生理参数的测定是一项繁重的任务, 自然界中多数物种的生理参数是未知的.而随着全球性物种分布数据库的共享以及地理信息系统技术的快速发展, 间接方法的生态位模型应运而生(McCormack et al. , 2010 ).对于多数物种分布的预测来说, 间接方法提供了一种简单易用的途径.本文如未加说明, 所指生态位模型均为间接方法的生态位模型.然而, 生态位的概念是生态位模型的理论基础, 对各种生态位概念的理解将会影响生态位模型构建的合理性, 模型构建的各个环节需要谨慎, 不恰当的方法容易得出错误的结论(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ). ...
... 示生态位模型将地理空间和生态空间二者结合引自(Pearson, 2007 ) ...
... Ecological niche modeling (ENM) building up the fridge of geographical space and ecological space adopted from (Pearson, 2007 ) ...
... 在自然保护区选择和设计时, 可应用生态位模型对物种实际分布范围进行模拟, 从而使自然保护区充分发挥功能, 减少不必要的开支.为了得到较为真实的物种分布, 在模型的构建上通常采取较为保守的策略, 需充分考虑影响物种分布的限制因素, 对模型的运算设定较多的限制(Araújo & Peterson, 2012 ).生态位模型的输出结果是一个对分布可能性大小的估测, 在后期校正时, 还应考虑物种间的相互作用和物种的迁移能力等因素(Pearson, 2007 ). ...
Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful ?
5
2003
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
... ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
... 不同影响因素在不同的空间尺度下作用于物种的分布修改自(Pearson & Dawson, 2003 ; Soberón, 2007 , 2010 ; Hortal et al ., 2010 ) ...
... Relative importance of factors affecting species distributions across spatial scales adopted from (Pearson & Dawson, 2003 ; Soberón 2007 , 2010 ; Hortal et al ., 2010 ) ...
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
Ecological niche conservatism: a time-structured review of evidence.
2011
Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri.
1
2008
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
Making biodiversity discovery more efficient: an exploratory test using Mexican birds.
1
2009
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
Future projections for Mexican faunas under global climate change scenarios
1
2002
... 生态位模型可模拟未来气候条件下物种的潜在分布.基本思路是: 基于已有的物种分布数据和当前的气候参数构建模型, 将其投射至将来的气候条件下, 来模拟未来物种潜在分布范围.目前, 基于联合国政府间气候变化委员会(IPCC)的评估报告, 加拿大、英国和澳大利亚等国的研究机构相继模拟出了未来不同时期的气候参数, 为未来物种潜在分布的模拟提供了基础(Hijmans et al ., 2005 ).在这类应用中, 强调了气候因素对物种分布的限制作用, 忽略了大尺度下物种间的相互作用和物种的迁移能力(Peterson et al ., 2002 ).这一应用的理论基础仍是生态位的保守性, 只有在此前提下, 模拟得到的未来气候条件下物种的分布才有价值(Petitpierre et al ., 2012 ).在大尺度范围下, 全球气候变化对北美和欧洲的鸟类和植物被广泛研究(e.g. Thuiller et al . 2005 ), 我国这方面的研究需要加强. ...
Integrating fundamental concepts of ecology, biogeography, and sampling into effective ecological niche modeling and species distribution modeling.
3
2012
... 近年来, 生态位模型在生物多样性保护的多个领域得到开拓和应用, 被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布的影响及传染病空间传播的研究中.但是, 在模型运用时却得出了一些不符合实际的结论, 如入侵物种的生态位迁移等(Broennimann et al ., 2007 ; Medley, 2010 ), 故一些学者对生态位模型提出了质疑.这些不当之处或多或少是由于对模型构建的目的认识模糊, 对生态位的概念和生态位模型的原理, 或者模型软件算法理解不透彻造成的(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ).为了让读者更系统地理解和正确地应用生态位模型, 我们梳理了生态位与物种分布的关系, 结合生态位概念介绍了生态位模型的基本原理, 从物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘种生态位保守性及基于生态位分化的物种界定等方面阐述了它在生物多样性保护中的应用. ...
... 广义生态位模型的构建包含两种截然不同的思路: 一种为直接的机理性方法(mechanistic approach), 另一种为间接的相关性方法(correlative approach), 前者基于物种对环境耐受性的生理参数来判断物种的潜在分布, 后者基于物种分布点所关联的环境参数来推算物种的分布(Pearson, 2007 ).在实际操作中, 第一种方法要求的物种生理参数的测定是一项繁重的任务, 自然界中多数物种的生理参数是未知的.而随着全球性物种分布数据库的共享以及地理信息系统技术的快速发展, 间接方法的生态位模型应运而生(McCormack et al. , 2010 ).对于多数物种分布的预测来说, 间接方法提供了一种简单易用的途径.本文如未加说明, 所指生态位模型均为间接方法的生态位模型.然而, 生态位的概念是生态位模型的理论基础, 对各种生态位概念的理解将会影响生态位模型构建的合理性, 模型构建的各个环节需要谨慎, 不恰当的方法容易得出错误的结论(Araújo & Peterson, 2012 ; Peterson & Soberón, 2012 ). ...
... 生态位模型用于物种的界定似乎对一些特殊类群更有利, 特别是一些迁移能力较弱、在不为人知的地区分布的特有种(Raxworthy et al ., 2007 ; Zhu et al. , 2013).我国学者将生态位等值性测试运用在中国林蛙复合体的物种界定上(Zhou et al ., 2012 ).在进行生态位等值性和相似性测试时, 需要将二者结合起来, 并对物种的分布范围给出一个合理的解释, 否则单一的生态位等值性测试不足以说明物种基础生态位的分化(Warren et al ., 2010 ; Peterson & Soberón, 2012 ). ...
3
2011
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
... 生态位的保守性对于物种的界定、入侵的模拟、气候变化对物种分布的影响以及生物多样性等方面的研究具有重要意义.生态位的保守性是一个敏感的话题, 近年引起了激烈的讨论.引发这场讨论的是Peterson等(1999)发表在Science 上的文章, 截至2013年1月1日该篇文章的引用已经超过了680次.Peterson等(1999)采用GARP模型对分布于墨西哥湾特万特佩克地峡(Isthmus of Tehuantepec)两侧的姊妹种(包括哺乳类、鸟类和蝴蝶)进行了模拟, 发现异域种化的姊妹种之间能够很好地相互模拟, 例如基于Atthis heloisa (Lesson & Delattre, 1839)的分布数据构建的模型能够很好地模拟其姊妹种A. ellioti (Ridgway, 1878)的分布; 同样, 基于A. ellioti 的分布记录的模型也能够很好地模拟A. heloisa 的种群分布记录.但是, 高级阶元间的保守性相对较差.目前学者们普遍认可的结论是: 生态位是进化的, 但是在较短的进化历史内, 是倾向保守的, 生态位的保守性与系统发育关系并不具有一致性(Wiens & Graham, 2005 ; Peterson, 2011 ).如果我们试图从进化上来验证物种间生态位的保守性, 那么首先要确认物种间的近缘关系, 它们是姊妹种, 或者是同一属, 或者是系统发育树上的近缘分支, 基于此来探讨生态位的保守与否才有意义. ...
Conservatism of ecological niches in evolutionary time.
1999
Climatic niche shifts are rare among terrestrial plant invaders.
1
2012
... 生态位模型可模拟未来气候条件下物种的潜在分布.基本思路是: 基于已有的物种分布数据和当前的气候参数构建模型, 将其投射至将来的气候条件下, 来模拟未来物种潜在分布范围.目前, 基于联合国政府间气候变化委员会(IPCC)的评估报告, 加拿大、英国和澳大利亚等国的研究机构相继模拟出了未来不同时期的气候参数, 为未来物种潜在分布的模拟提供了基础(Hijmans et al ., 2005 ).在这类应用中, 强调了气候因素对物种分布的限制作用, 忽略了大尺度下物种间的相互作用和物种的迁移能力(Peterson et al ., 2002 ).这一应用的理论基础仍是生态位的保守性, 只有在此前提下, 模拟得到的未来气候条件下物种的分布才有价值(Petitpierre et al ., 2012 ).在大尺度范围下, 全球气候变化对北美和欧洲的鸟类和植物被广泛研究(e.g. Thuiller et al . 2005 ), 我国这方面的研究需要加强. ...
On the relationship between niche and distribution.
3
2000
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... Hutchinson(1957)使用现实生态位来描述基础生态位在地理空间的反映, 这里的限制因素主要指物种间的相互作用.通常来讲, 基础生态位大于现实生态位, 这是由于物种间的相互作用和物种的迁移能力一定程度上影响了基础生态位在地理空间的反映(图1 ), 因此一些物种并未在其生态适宜地区出现(Holt, 2003 ); 另一方面, 由于汇种群的影响, 一些物种会暂时迁移到不适宜生存的区域, 如迁徙的鸟类, 这是现实生态位大于基础生态位的情况(Pulliam, 2000 ).因此, 在构建生态位模型时需要注意是否包含了汇种群的分布点(图1 ). ...
Applications of ecological niche modeling for species delimitation: a review and empirical evaluation using day geckos (Phelsuma) from Madagascar.
2
2007
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
... 生态位模型用于物种的界定似乎对一些特殊类群更有利, 特别是一些迁移能力较弱、在不为人知的地区分布的特有种(Raxworthy et al ., 2007 ; Zhu et al. , 2013).我国学者将生态位等值性测试运用在中国林蛙复合体的物种界定上(Zhou et al ., 2012 ).在进行生态位等值性和相似性测试时, 需要将二者结合起来, 并对物种的分布范围给出一个合理的解释, 否则单一的生态位等值性测试不足以说明物种基础生态位的分化(Warren et al ., 2010 ; Peterson & Soberón, 2012 ). ...
Predicting distributions of known and unknown reptile species in Madagascar.
1
2003
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
Adding more ecology into species delimitation: ecological niche models and phylogeography help define cryptic species in the Black Salamander (Aneides flavipunctatus).
1
2007
... 物种的界定是进化生物学的核心争论问题, 分歧的核心是对物种定义的不同.近年来, 生态位的分化被越来越多地用于物种的界定上(e.g. Leaché et al ., 2009 ).物种分化和生态位分化的步调可能是一致的(即生态位的分化性), 但也可能是不完全一致的(即生态位的保守性).在进化的历史过程中, 新物种的产生总是倾向于保留其祖先的生态位, 然而, 新物种在适应新的环境时, 会发生一些适应性的基因突变, 这些突变可能导致生态位的分化, 反过来又促进物种的分化.总之, 生态位分化的依据为物种的界定提供了一条途径, 越来越多的学者呼吁在物种界定时考虑生态因素(Rissler & Apodaca, 2007 ).基于生态位模型的生态位分化研究也越来越细, 从物种间的生态空间分析, 到物种间的生态位测试, 如Warren等(2008, 2010)等开发的生态位等值性测试(niche equivalency or identity test)和相似性测试(similarity or background test)已经被越来越多的学者采用, 并且与DNA条形码结合用于新物种的描述(Hawlitschek et al ., 2011 ). ...
Pushing the limits in marine species distribution modelling: lessons from the land present challenges and opportunities.
1
2011
... 地球上的物种数目庞大, 用测定生理参数的方法来分析所有物种的生态空间需求显然不现实, 加之一些物种本身的地理分布特性, 使得生态位模型显示出优越性.例如有些物种生境一般具有隐蔽性, 局限于某一地区分布, 很难采集到.生态位模型基于可获取的有限的物种分布点及其所关联的环境参数, 为揭开这些物种的神秘“生态面纱”提供了一种有力的工具.特别是对于一些需要保护的物种, 例如世界自然保护联盟(IUCN)红色名录上的物种, 其应用更为重要.近年来, 这一工具不仅被广泛应用到陆地植物和动物(Hirzel et al ., 2002 ; Irfan-Ullah et al ., 2007 ; Luoto et al ., 2006 ; Dorji et al ., 2011 ), 并且已经拓展到了海洋物种(Owens et al ., 2011 ; Robinson et al ., 2011 ). ...
Niche shift versus niche conservatism? Climatic characteristics of the native and invasive ranges of the Mediterranean house gecko (Hemidactylus turcicus).
2
2009
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 ).而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题.文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化.在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性. ...
Variation in niche and distribution model performance: the need for a priori assessment of key causal factors.
1
2012
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods.
5
2011
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
... 在某一历史时期, 可获取的生态空间是有限的.Jackson和Overpeck(2000)使用潜在生态位(potential niche)来描述基础生态位在有限生态空间中的反映, 即基础生态位和可获取生态空间的交集(Colwell & Rangel, 2009 ).潜在生态位体现了特定历史时期内基础生态位在生态空间中的可获取性, 物种间的相互作用及物种的迁移能力限制了其在特定地理空间的反映, 所以说现实生态位是潜在生态位的一部分(Sillero, 2011 ). ...
... 在实际操作中, 生态位模型所模拟的是现实生态位, 而不是基础生态位.然而, 由于各模型算法所采用“不存在”数据(absence records)不同, 所模拟的现实生态位亦是有差别的(Soberón, 2007 ; Sillero, 2011 ).例如, 有的生态位模型同时使用存在数据(presence records)和不存在数据来模拟, 有的使用假设不存在(pseudo-absence)来替代物种的不存在数据, 所得到的模拟结果是不同的.另外, 机理性生态位模型反映的是特定生态空间中的基础生态位, 即潜在生态位, 并不是基础生态位(Sillero, 2011 ). ...
... ).例如, 有的生态位模型同时使用存在数据(presence records)和不存在数据来模拟, 有的使用假设不存在(pseudo-absence)来替代物种的不存在数据, 所得到的模拟结果是不同的.另外, 机理性生态位模型反映的是特定生态空间中的基础生态位, 即潜在生态位, 并不是基础生态位(Sillero, 2011 ). ...
Grinnellian and Eltonian niches and geographic distributions of species.
8
2007
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 不同影响因素在不同的空间尺度下作用于物种的分布修改自(Pearson & Dawson, 2003 ; Soberón, 2007 , 2010 ; Hortal et al ., 2010 ) ...
... Relative importance of factors affecting species distributions across spatial scales adopted from (Pearson & Dawson, 2003 ; Soberón 2007 , 2010 ; Hortal et al ., 2010 ) ...
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
... 在实际操作中, Grinnell生态位和Elton生态位均已反映在生态位模型中.该两种生态位与模型构建的环节密切相关, 主要取决于环境变量类型的选择、所采用环境变量的分辨率以及物种本身的迁移能力三个方面(Soberón, 2007 , 2010 ). ...
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... 在实际操作中, 生态位模型所模拟的是现实生态位, 而不是基础生态位.然而, 由于各模型算法所采用“不存在”数据(absence records)不同, 所模拟的现实生态位亦是有差别的(Soberón, 2007 ; Sillero, 2011 ).例如, 有的生态位模型同时使用存在数据(presence records)和不存在数据来模拟, 有的使用假设不存在(pseudo-absence)来替代物种的不存在数据, 所得到的模拟结果是不同的.另外, 机理性生态位模型反映的是特定生态空间中的基础生态位, 即潜在生态位, 并不是基础生态位(Sillero, 2011 ). ...
Interpretation of models of fundamental ecological niches and species' distributional areas.
3
2005
... 影响物种地理分布的因素修改自(Soberón & Peterson, 2005 ; Soberón, 2010 ).A环表示对物种分布有利的非生物因素(基础生态位), B环表示物种相互作用中对该物种有利的区域, M环表示物种迁移能力范围内的区域, Go I
... Diagram showing a simplified framework for understanding species distribution adopted from (Soberón & Peterson, 2005 ; Soberón, 2010 ). Overlap of the three circles (GO I
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
Niche and area of distribution modeling: a population ecology perspective.
6
2010
... 物种的分布是静态的又是动态的, 是其生态位在历史过程中复杂表达的结果(Brown, 1995 ).在某一历史时期, 同一物种的生态位在不同环境下的表达亦是不同的, 模拟物种在这些条件下的分布具有重要意义.在不同的环境尺度下, 影响物种分布的各类因素的作用程度是不同的(Gaston, 2003 ).Soberón和Peterson(2005)将影响物种分布的因素总结为以下4类: (1)非生物因素(abiotic or scenopoetic factor), 包括气候、土壤条件等影响物种生理特性的各种因子; (2)物种间的相互作用(biotic or bionomic factor), 可以是对物种有利的(如物种间的互利共生等), 也可以是不利的(如竞争、取食等); (3)该地区位于物种的迁移能力范围之内, 这取决于物种本身的迁移能力和地理区域的特性; (4)物种或种群对新环境的适应能力, 即在新的环境下, 物种改变其生理特性以适应环境的能力.通常情况下, 在较短的历史时期内, 物种的生态位是保守的, 生态位的进化相对较小(Etterson & Shaw, 2001 ; Peterson, 2011 ).前3种因素的划分不是绝对的(Soberón, 2007 ), 它们在不同的空间尺度下以不同的机制作用于物种的分布(Hortal et al ., 2010 ).Soberón和Peterson(2005)使用BAM图来直观地展示前3个因素对物种分布的影响(图1 ), 被学者广泛接受和采用(Rödder & Lötters, 2009 ; Soberón, 2010 ; Sillero, 2011 ). ...
... 影响物种地理分布的因素修改自(Soberón & Peterson, 2005 ; Soberón, 2010 ).A环表示对物种分布有利的非生物因素(基础生态位), B环表示物种相互作用中对该物种有利的区域, M环表示物种迁移能力范围内的区域, Go I
... Diagram showing a simplified framework for understanding species distribution adopted from (Soberón & Peterson, 2005 ; Soberón, 2010 ). Overlap of the three circles (GO I
... 不同影响因素在不同的空间尺度下作用于物种的分布修改自(Pearson & Dawson, 2003 ; Soberón, 2007 , 2010 ; Hortal et al ., 2010 ) ...
... Relative importance of factors affecting species distributions across spatial scales adopted from (Pearson & Dawson, 2003 ; Soberón 2007 , 2010 ; Hortal et al ., 2010 ) ...
... 在实际操作中, Grinnell生态位和Elton生态位均已反映在生态位模型中.该两种生态位与模型构建的环节密切相关, 主要取决于环境变量类型的选择、所采用环境变量的分辨率以及物种本身的迁移能力三个方面(Soberón, 2007 , 2010 ). ...
Limited filling of the potential range in European tree species.
2
2004
... 在环境变量类型的选择上, 如果选取的变量主要为气候变量参数, 例如常见的19种生物气候变量(Hijmans et al ., 2005 ), 那么所模拟的生态位即为Grinnell生态位; 如果环境变量参数又添加了高分辨率的植被参数, 例如归一化植被指数(NDVI)和绿度植被指数(GVI)等, 那么所模拟的生态位便有了Elton生态位的成分(McCormack et al ., 2010 ).在分辨率的选择上, Elton生态位倾向出现在较小尺度的生物类环境变量, Grinnell生态位倾向出现在较大尺度的气候变量(Soberón, 2007 ).物种自身的迁移能力也受到越来越多的重视(Pulliam, 2000 ; Pearson & Dawson, 2003 ; Svenning & Skov, 2004 ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
... ).理论上讲, 在低分辨率的环境变量条件下, 多数地理空间是位于物种的迁移能力范围之内的(Svenning & Skov, 2004 ), 这些环境变量可以是非生物因素也可以是生物因素(Soberón, 2007 ).然而,由于源-汇种群(source-sink population)的影响(Pulliam, 2000 ), 一些物种的迁移能力较强, 便会出现在不适宜其生存的区域(图1 ). ...
Climate change threats to plant diversity in Europe.
1
2005
... 生态位模型可模拟未来气候条件下物种的潜在分布.基本思路是: 基于已有的物种分布数据和当前的气候参数构建模型, 将其投射至将来的气候条件下, 来模拟未来物种潜在分布范围.目前, 基于联合国政府间气候变化委员会(IPCC)的评估报告, 加拿大、英国和澳大利亚等国的研究机构相继模拟出了未来不同时期的气候参数, 为未来物种潜在分布的模拟提供了基础(Hijmans et al ., 2005 ).在这类应用中, 强调了气候因素对物种分布的限制作用, 忽略了大尺度下物种间的相互作用和物种的迁移能力(Peterson et al ., 2002 ).这一应用的理论基础仍是生态位的保守性, 只有在此前提下, 模拟得到的未来气候条件下物种的分布才有价值(Petitpierre et al ., 2012 ).在大尺度范围下, 全球气候变化对北美和欧洲的鸟类和植物被广泛研究(e.g. Thuiller et al . 2005 ), 我国这方面的研究需要加强. ...
In defense of niche modeling.
3
2012
... 生态位模型将物种分布相关联的地理空间和生态空间联系起来, 它基于物种分布的地理空间信息来构建维持物种生存的生态空间.如图3 所示, 生态位模型利用物种分布点相关联的环境参数进行运算来构建模型, 将其投射至另一地理空间判断该物种未知的实际分布地和潜在分布地(Araújo & Peterson, 2012 ).在实际运用中, 生态位模型所基于的物种分布点关联着其他物种的分布, 受其他物种分布的影响(如寄生、竞争、取食等), 所以生态位模型所模拟的是现实生态位, 并不是该物种基础生态位在环境中的完全释放(Soberón & Peterson, 2005 ).另外, 生态位模型所模拟的物种分布图与简单的物种分布图和丰富度图是不同的(Warren, 2012 ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
... ).后两种分布图仅基于物种的分布地点来推算物种的空间分布(Warren, 2012 ), 而生态位模型是基于物种分布点所关联的环境参数来推算物种的生态需求, 然后将其投射至地理空间来估测物种的地理分布, 这其中包含了物种分布点和环境参数, 环境参数在很大程度上决定了所模拟的生态位成分和地理分布(Soberón, 2007 ).参照国外学者建议使用“ecological niche modeling”来描述这一方法(Sillero, 2011 ; Saupe et al ., 2012 ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
... ; Warren, 2012 ), 我们建议在国内使用“生态位模型”这一术语来取代物种分布模型、物种预测模型、生境预测模型等说法. ...
Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution.
2008
ENMTools: a toolbox for comparative studies of environmental niche models.
1
2010
... 生态位模型用于物种的界定似乎对一些特殊类群更有利, 特别是一些迁移能力较弱、在不为人知的地区分布的特有种(Raxworthy et al ., 2007 ; Zhu et al. , 2013).我国学者将生态位等值性测试运用在中国林蛙复合体的物种界定上(Zhou et al ., 2012 ).在进行生态位等值性和相似性测试时, 需要将二者结合起来, 并对物种的分布范围给出一个合理的解释, 否则单一的生态位等值性测试不足以说明物种基础生态位的分化(Warren et al ., 2010 ; Peterson & Soberón, 2012 ). ...
Conservation biogeography: assessment and prospect.
2
2005
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
... 一直以来, 地球上物种的确切数目及其分布困扰着生物学家.前者称为Linnean不足(Linnean Shortfall), 指人们对地球上物种多样性的数目掌握不全面; 后者称为Wallacean不足(Wallacean Shortfall), 指人们对这些物种的分布范围掌握不足(Whittaker et al ., 2005 ).生态位模型可以帮助人们判断未知种群的实际分布地(Bourg et al ., 2005 ; Menon et al ., 2010 ), 从而为野外采集或调查提供参考, 来弥补Wallacean不足.另外, 由于物种间的相互作用以及物种的迁移能力有限, 模拟得到的某物种潜在分布地可能并不为该物种所占据, 反而可能被另外一个未知的物种所占据(Raxworthy et al ., 2003 , 2007 ), 基于此设想而进行野外调查或许可以加快新物种的发现和记述(Peterson & Navarro-Sigüenza, 2009 ), 进而弥补Linnean不足.这方面的应用值得分类学者尝试. ...
Species diversity―scale matters.
1
2002
... 一般来说, 非生物的因素(主要是气候因素)主要在大尺度空间影响物种的分布, 这些因素很大程度上决定了物种的分布范围和格局, 包括生理制约、物种对气候和生境梯度的响应和选择等.不同的非生物因素, 其作用的尺度也有差别(图2 ; Pearson & Dawson, 2003 ).生物的影响因子对物种分布的影响较为复杂, 主要在较小的空间尺度下作用于物种的分布, 而在较大的空间尺度下, 物种间的相互作用被弱化, 变得不明显(Mackey & Lindenmayer, 2001 ; Pearson & Dawson, 2003 ; Guisan & Thuiller, 2005 ; Hortal et al ., 2010 ).这与它们在不同的空间尺度下对物种多样性格局的影响是一致的(Whittaker et al ., 2001 ; Willis & Whittaker, 2002 ).Hortal等(2010)将Soberón(2007, 2010)所指的物种迁移能力分为生物地理因素(biogeographic factors)和物种的存在动态因素(occupancy dynamics)两类.前者指物种的生物地理分布区和地理隔离因素等, 主要在较大的空间尺度下影响物种的分布; 后者指物种的种群动态、短距离的迁移和地区性的干扰因素, 它们主要在相对较小尺度下影响物种的分布(图2 ). ...
Niche conservatism: integrating evolution, ecology, and conservation biology.
1
2005
... 生态位的保守性对于物种的界定、入侵的模拟、气候变化对物种分布的影响以及生物多样性等方面的研究具有重要意义.生态位的保守性是一个敏感的话题, 近年引起了激烈的讨论.引发这场讨论的是Peterson等(1999)发表在Science 上的文章, 截至2013年1月1日该篇文章的引用已经超过了680次.Peterson等(1999)采用GARP模型对分布于墨西哥湾特万特佩克地峡(Isthmus of Tehuantepec)两侧的姊妹种(包括哺乳类、鸟类和蝴蝶)进行了模拟, 发现异域种化的姊妹种之间能够很好地相互模拟, 例如基于Atthis heloisa (Lesson & Delattre, 1839)的分布数据构建的模型能够很好地模拟其姊妹种A. ellioti (Ridgway, 1878)的分布; 同样, 基于A. ellioti 的分布记录的模型也能够很好地模拟A. heloisa 的种群分布记录.但是, 高级阶元间的保守性相对较差.目前学者们普遍认可的结论是: 生态位是进化的, 但是在较短的进化历史内, 是倾向保守的, 生态位的保守性与系统发育关系并不具有一致性(Wiens & Graham, 2005 ; Peterson, 2011 ).如果我们试图从进化上来验证物种间生态位的保守性, 那么首先要确认物种间的近缘关系, 它们是姊妹种, 或者是同一属, 或者是系统发育树上的近缘分支, 基于此来探讨生态位的保守与否才有意义. ...
Speciation in the Rana chensinensis species complex and its relationship to the uplift of the Qinghai-Tibetan Plateau.
1
2012
... 生态位模型用于物种的界定似乎对一些特殊类群更有利, 特别是一些迁移能力较弱、在不为人知的地区分布的特有种(Raxworthy et al ., 2007 ; Zhu et al. , 2013).我国学者将生态位等值性测试运用在中国林蛙复合体的物种界定上(Zhou et al ., 2012 ).在进行生态位等值性和相似性测试时, 需要将二者结合起来, 并对物种的分布范围给出一个合理的解释, 否则单一的生态位等值性测试不足以说明物种基础生态位的分化(Warren et al ., 2010 ; Peterson & Soberón, 2012 ). ...
Potential geographic distribution of Brown Marmorated Stink Bug invasion (Halyomorpha halys).
2
2012
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
... 在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 ).而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题.文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化.在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性. ...
Selecting biological meaningful environmental dimensions of low discrepancy among ranges to predict potential distribution of bean plataspid invasion.
3
2012
... 生态位模型构建有以下3个前提条件(Peterson et al ., 2011 ): (1)物种的生态需求和分布处于平衡的状态.是指在一段历史时期内, 物种的分布处于一种饱和的状态, 即在所有适合某物种分布的地方均有了该物种的分布; (2)物种的迁移能力是无限的.这个前提强调物种的迁移能力, 忽略了物种间的相互作用、物种本身的迁移能力以及地理的阻隔作用.在现实中, 种间作用和阻隔作用有时比较强, 会影响模型的预测; (3)物种的生态位是保守的.这个前提是最为关键的, 生态位保守性指在一段历史时期内物种的生态位是不变的.只有在生态位保守性的前提下, 构建后的模型投射至另一地理空间或时间下才具有合理性.近年, 常有物种入侵过程中伴随生态位迁移的报道(Broennimann et al ., 2007 ; Medley, 2010 ), 然而, 当采用较为合理的环境变量后, 模型的转移能力(transferability)可明显提高(Peterson & Nakazawa, 2008 ; Zhu et al. , 2012a , b ). ...
... 在生态位模型构建中, 所需的物种分布点本身就已经包含物种间的相互作用, 因此模型所模拟的是现实生态位或者潜在生态位, 并不是基础生态位(图1 ).而基础生态位才具有真正进化上的生物学意义, 因此只有明确了现实生态位在多大程度上反映了基础生态位, 才能够更好地说明生态位保守性问题.文献中所报道的生态位迁移(Broennimann et al ., 2007 ; Medley, 2010 ), 只能说是基础生态位在不同地区的反映, 或是现实生态位的不同, 不能说是进化意义上的基础生态位的迁移或分化.在实际中, 不同地理种群的现实生态位在不同的生态空间维度下又可能是不同的(Rödder & Lötters, 2009 ; Zhu et al ., 2012a , b ), 它们可能在温度上是一致的, 而在湿度上是不一致的, 这将会增加空间问题的复 杂性. ...
... 生物入侵是生物多样性丧失的主要原因之一, 生态位模型可用于对入侵物种潜在分布的模拟, 这一领域的应用较为广泛.通常是基于物种原产地的分布数据来构建模型, 然后用于判断该物种在入侵地的潜在分布(图3 ), 为该物种的防范和监控提供参考.因此, 模型构建所选用的物种分布点必须能够反映该物种的分布范围, 所选环境变量也必须对该物种的分布具有较大的限制作用, 这样才能提高模型的可转移性(Araújo & Peterson, 2012 ), 从而准确模拟物种在入侵地的分布范围.这一应用的理论基础是生态位的保守性, 只有在此前提下进行模型的转移和预测才具有合理性.如果某物种在原产地的分布数据比较难获取, 那么也可以基于其在入侵地的分布数据来构建模型, 进行潜在分布范围的模拟(Zhu et al ., 2007 ). ...
Predicting the spatial distribution of an invasive plant species (Eupatorium adenophorum) in China.
2007
 , 刘国卿, 卜文俊, 高玉葆
, 刘国卿, 卜文俊, 高玉葆

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}