


珊瑚礁是地球上已知的海洋生物多样性最高的生态系统, 尤其在东南亚海区, 即中印度—西太平洋区的珊瑚礁, 其珊瑚和礁栖生物种类的多样性最为丰富(McManus, 2010)。浮游动物是珊瑚礁生物群落中, 特别是珊瑚和鱼类的重要食物和营养来源(Alldredge & King, 1977, 2009; Coma et al., 1999; Heidelberg, 2004)。珊瑚虫不能从共生虫黄藻的光合作用中获得氮、磷等营养, 必须捕食浮游动物予以补充(Johannes et al., 1970; Sebens et al., 1996), 因此浮游动物在珊瑚礁生态系统的物质循环和能量流动中具有十分重要的地位。
南海紧邻东南亚海区, 珊瑚礁类型主要为岸礁和环礁, 在岸礁(尹健强等, 2008; Nakajima et al., 2008, 2009)和环礁(陈清潮和尹健强, 1982; 陈清潮等, 1989; 钱宏林等, 1990; 章淑珍和李纯厚, 1997; 尹健强等, 2003)浮游动物群落方面已开展过一些研究。此外, 在浮游动物物种多样性方面, 陈清潮(1983)发现了黄岩岛环礁的桡足类一新种中华歪水蚤(Tortanus sinicus); Chen等(2004)、陈清潮(2008)发现了南沙群岛渚碧礁的桡足类两个新种膨大歪水蚤(T. tumidus)和珍妮纺锤水蚤(Acartia shuzheni); 张谷贤和尹健强(2002)首次在国内报道了在渚碧礁出现毛颚类的锄虫属(Spadella)种类。但总体来说, 南海特别是南海诸岛珊瑚礁的浮游动物多样性和群落结构, 由于受取样困难等因素的制约, 研究还很不充分。本文根据2004年5月在南沙群岛渚碧礁的调查资料, 进一步分析浮游动物的群落结构和多样性, 为深入开展珊瑚礁生态系统研究提供科学资料。
1 材料与方法
1.1 研究区概况及调查站位
渚碧礁位于南沙群岛海区北部, 是一个典型的珊瑚礁, 中心位置大约在10°55′ N、114°05′ E。礁体近似梨形, NE-SW向, 长5.75 km, 宽3.25 km, 面积约16.1 km2。礁坪北、西和西南部较宽, 达500-600 m, 地势稍高; 东部和南部较窄, 仅约300 m, 地势较低矮。中间澙湖面积约9.5 km2, 水深大部分在20 m左右, 最大深度为24 m。属热带和赤道带海洋季风气候, 终年高温高湿, 5月下旬至9月盛行西南季风, 11月至翌年4月中旬盛行东北季风, 4-5月与10月分别为东北、西南与西南、东北季风转换时期(林锡贵和张庆荣, 1990)。该礁为封闭型环礁, 边缘全被礁坪围封, 没有口门通往外海。大潮低潮时, 礁坪基本露出, 澙湖水体与外海水不能交换, 高潮时外海海水才能漫入澙湖; 小潮时, 礁盘可终日被海水淹没。
在渚碧礁共设10个大面站, 其中1、2、3、4、5号站位于澙湖内, 6、7、8、9、10号站位于礁坪, 另在西南礁坪设1个连续站(图1)。礁坪连续站的水深在0.8-1.8 m之间(受潮汐影响), 其他礁坪测站的水深在1.1-2.3 m之间, 澙湖的测站水深在17.0-21.5 m之间。
图1
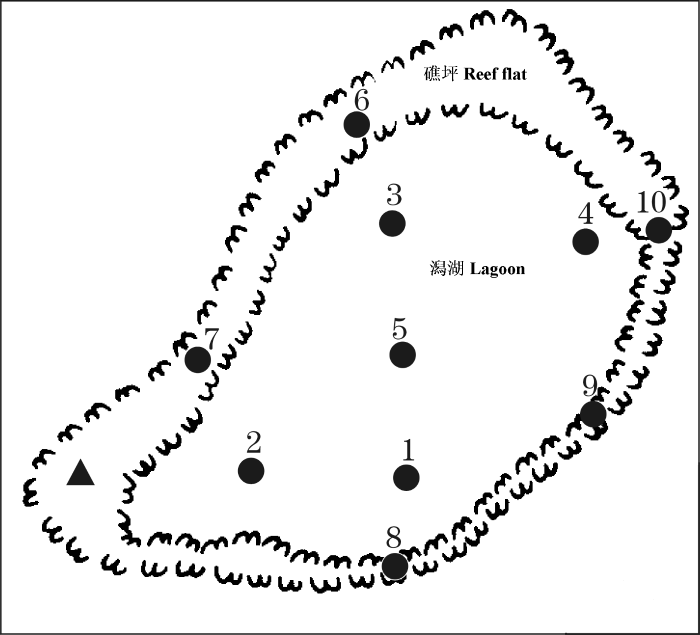
图1
渚碧礁浮游动物调查站位图(●大面站; ▲连续站)
Fig. 1
Maps with the sampling stations of zooplankton in the Zhubi Atoll of the South China Sea (●sampling stations within the lagoon and on the reef flat; ▲one continuous observatory station)
1.2 样品采集与环境因子观测
调查于2004年5月5-15日进行, 大面站分别使用浅水I型浮游生物网(网口内径50 cm, 网长145 cm, 网目孔径505 μm)和修改的浅水II型浮游生物网(网口内径50 cm, 网长145 cm, 网目孔径169 μm)由底至海面垂直拖网采集。采样在上午8:00-11:30进行。连续站在小潮期间进行(2004年5月13-14日), 使用浅水II型浮游生物网由底至海面垂直拖网采集, 每隔2 h进行一次, 共取样13次, 每次采样重复3次。样品均用5%的福尔马林溶液固定保存, 运回实验室在解剖镜下鉴定计数。大多数浮游动物鉴定到种(陈清潮和章淑珍, 1974a, b; 郑重等, 1984)。放射虫、水母类、介形类等个别类群未完全鉴定到种; 幼虫按习惯鉴定到类群或类型。在进行种数统计时, 未鉴定到种的类群按1种统计。浮游动物的丰度、多样性指数、均匀度分布根据浅水II型网的分析结果,并进行了两种网具分析结果的比较。
温度和盐度使用YSI 6600进行现场测定。在连续站进行了叶绿素a的测定, 现场采水后, 经0.45 μm孔径醋酸纤维滤膜过滤、丙酮萃取后使用Turner-Design 10型荧光计测定。
1.3 数据分析
采用Shannon-Wiener多样性指数(H')和Pielou均匀度指数(J)计算浮游动物的群落多样性(Shannon, 1948):
式中, Pi为该站中第i种的个体数与该站总个体数的比值, S为该站的种数。
优势种的确定由优势度(Y)决定, 计算公式如下:
式中, ni为第i种的个体数, N为总个体数, fi为第i种在各站位出现的频率。优势度Y >0.02确定为优势种(徐兆礼和陈亚瞿, 1989)。
应用t检验法进行了两种网具采集效果的差异显著性检验。
依据大面站浅水II型网调查获得浮游动物数据, 为了减少机会种对群落结构分析的影响, 将浮游动物种类按照丰度由高到低排列, 挑选出前50种, 从而得到用于群落结构分析的物种。将这些种类的丰度以log10(x+1)转换, 利用PRIMER6.0软件, 采用Bray-Curtis相似性系数进行等级聚类(hierarchical cluster)分析调查海区浮游动物群落结构特征(Clarke & Gorley, 2006)。
2 结果
2.1 环境因子
2.1.1 大面站
大面站总体的表层和底层温度平均值 (Mean±SD)分别为29.87±0.14℃和29.81 ± 0.17℃; 澙湖区分别为29.82 ± 0.04 ℃和29.70 ± 0.06 ℃; 礁坪区分别为29.91 ± 0.20 ℃和29.91 ± 0.19 ℃。总体来说, 调查区表层和底层的温度差别非常小, 礁坪区的水温略微高于澙湖区。由于澙湖水较深, 而礁坪水浅, 澙湖区所有站位的底层水温稍低于表层, 礁坪区的表层和底层水温几乎没有差别(图2a)。礁坪区各站的温度变化相对较大, 与采样时间差异和天气变化(如降雨, 8号站)有关。
图2
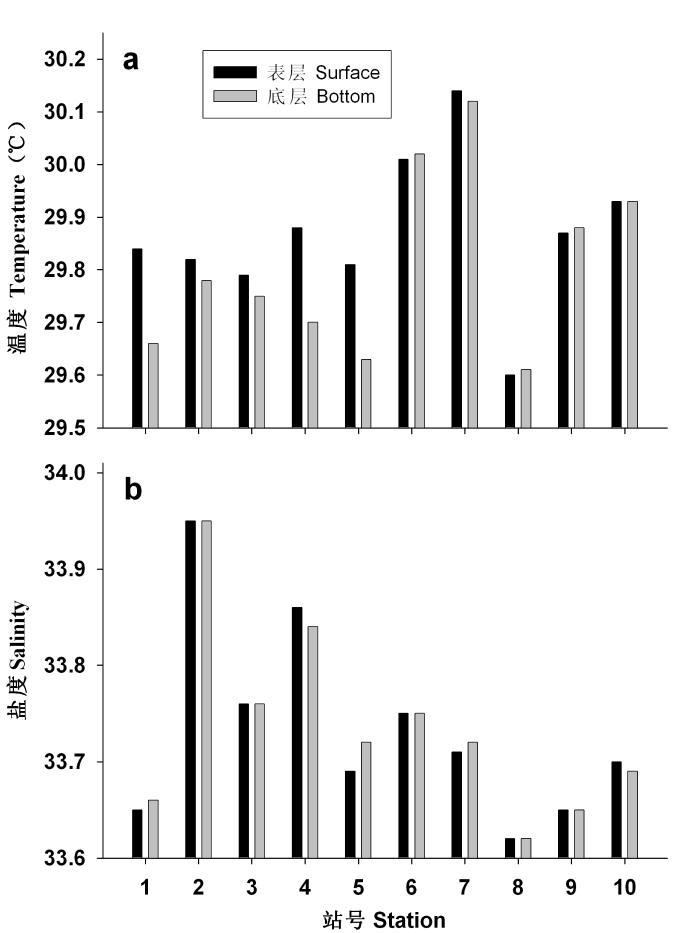
图2
渚碧礁大面站表层和底层的温度(a)与盐度(b)分布
Fig. 2
Distribution of temperature (a) and salinity (b) at the surface and bottom layers in the Zhubi Atoll
大面站总体的表层和底层盐度平均值分别为33.73 ± 0.10和33.74 ± 0.10; 澙湖区分别为33.78 ± 0.12和33.97 ± 0.11; 礁坪区分别为33.69 ± 0.05和33.69 ± 0.05。总体来说, 调查区盐度垂直分布均匀, 澙湖区各站的盐度变化幅度稍大于礁坪区(图2b)。
2.1.2 连续站
连续站的水深变化受潮汐影响, 采样期间的潮汐类型为全日潮, 高潮出现于白天, 16:00达最高潮; 低潮出现于夜晚, 2:00-4:00为最低潮(图3a), 涨潮历时大于落潮历时。在高潮期间水流较急, 礁外海水可明显影响调查位置; 而在低潮期间由于受礁坪 的阻挡, 水流缓慢, 礁外海水对调查位置的影响较小。
图3
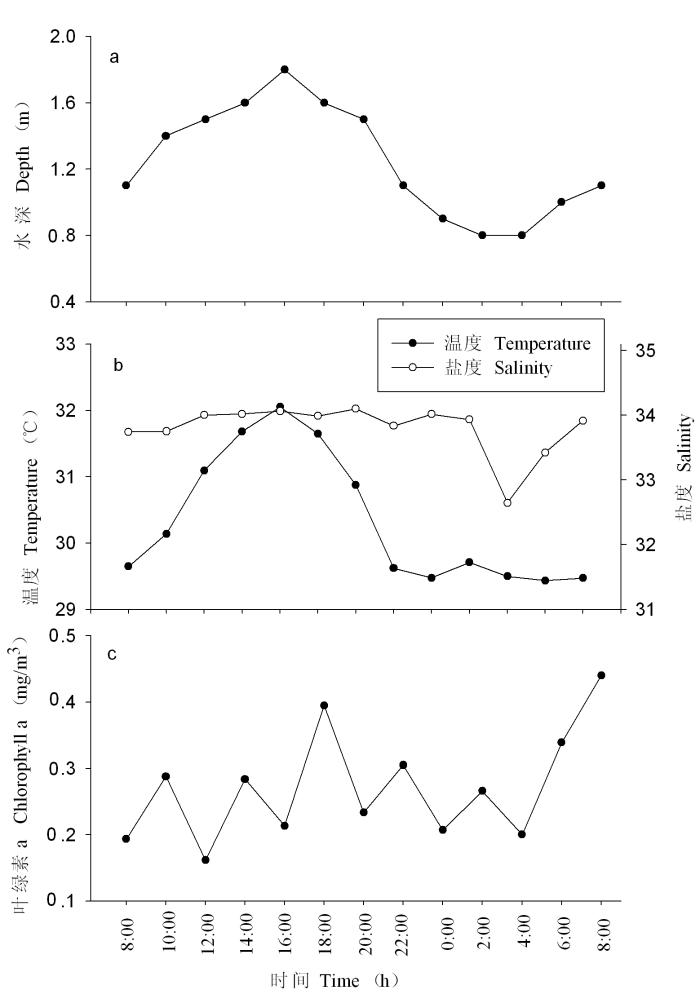
图3
渚碧礁连续站的水深(a)、温度和盐度(b)及叶绿素a含量(c)的昼夜变化
Fig. 3
Diurnal variations of depth (a), temperature (℃), salinity (b) and chlorophyll a (mg/m3) (c) at the continuous observatory station
2.2 种类组成及其种数分布
本次调查鉴定出的浮游动物超过96个种(包括17种以上的未定种)和浮游幼虫17个类群(或类型)(附录I), 包括放射虫1种、水母类3种、软体动物1种、枝角类2种、介形类4种、桡足类65种、涟虫类1种、端足类1种、十足类1种、毛颚类10种、被囊类6种和头索动物1种。其中桡足类的种类最多, 其种数占浮游动物总种数(包括浮游幼虫)的57.5%, 其次为浮游幼虫, 占浮游动物总种数的15.0%。使用浅水II型网采集的浮游动物的种类数显著多于浅水I型网, 除四叶小舌水母(Liriope tetraphylla)1种外, 用浅水I型网能采到的种类都能用浅水II型网采到。在澙湖出现的种类显著多于礁坪, 用浅水I型网和浅水II型网采集的样品, 在澙湖出现的种类分别为礁坪的2.25倍和2.93倍。
浮游动物种数的平面分布见表1。根据浅水II型网采样的分析结果, 澙湖区和礁坪区的浮游动物种数分布差异显著, 前者的变化范围为19-32种, 平均为27.6 ± 5.2种/站, 后者的变化范围为2-14种, 平均为7.8 ± 5/站, 前者平均数为后者的3.5倍。
表1 浮游动物种数、多样性指数和均匀度的分布
Table 1
| 区域 Region | 站号 Station | 浅水I型网 Planktonic net I | 浅水II型网 Planktonic net II | |||||
|---|---|---|---|---|---|---|---|---|
| 种数 Richness (S) | 多样性指数 Diversity index (H ') | 均匀度 Evenness index (J) | 种数 Richness (S) | 多样性指数 Diversity index (H ') | 均匀度 Evenness index (J) | |||
| 澙湖 Lagoon | 1 | 4 | 1.796 | 0.898 | 31 | 2.136 | 0.431 | |
| 2 | 8 | 2.529 | 0.843 | 19 | 1.382 | 0.325 | ||
| 3 | 11 | 2.572 | 0.743 | 29 | 1.892 | 0.390 | ||
| 4 | 8 | 1.619 | 0.468 | 32 | 2.160 | 0.363 | ||
| 5 | 11 | 2.504 | 0.724 | 27 | 2.098 | 0.441 | ||
| 礁坪 Reef flat | 6 | 0 | - | - | 7 | 2.210 | 0.787 | |
| 7 | 2 | 1.000 | 1.000 | 2 | 1.000 | 1.000 | ||
| 8 | 0 | - | - | 12 | 2.960 | 0.826 | ||
| 9 | 6 | 1.787 | 0.691 | 14 | 3.442 | 0.904 | ||
| 10 | 1 | 0.000 | 1.000 | 4 | 2.000 | 1.000 | ||
2.3 丰度分布
浮游动物丰度变化范围为8.4-3,326.5 inds./m3, 平均为926.0 ± 1,155.8 inds./m3。澙湖区的平均丰度高达1,798.1 ± 1,050.2 inds./m3, 礁坪区的仅为53.9 ±38.2 inds./m3, 澙湖区浮游动物丰度明显高于礁坪区(图4), 为礁坪区的33.4倍。
图4
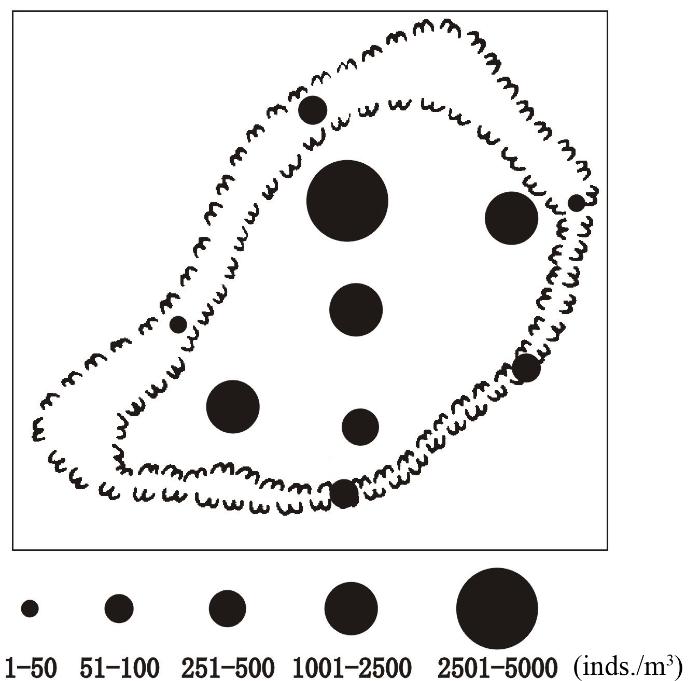
图4
渚碧礁浮游动物总丰度(inds./m3)的分布
Fig. 4
Distribution of zooplankton abundance (inds./m3) in the Zhubi Atoll
桡足类、被囊类和浮游幼虫是渚碧礁浮游动物的三大类群, 它们的丰度分别占浮游动物总丰度的78.56%、13.51%和7.26%。从图5可以看出, 它们均密集于澙湖区, 而在礁坪区的数量非常稀少。桡足类、被囊类在澙湖区的丰度分别是礁坪区的41.54和47.48倍, 而幼虫的差别相对较小, 仅为11.78倍, 可能与底栖生物主要分布于礁坪有一定的关系。
图5
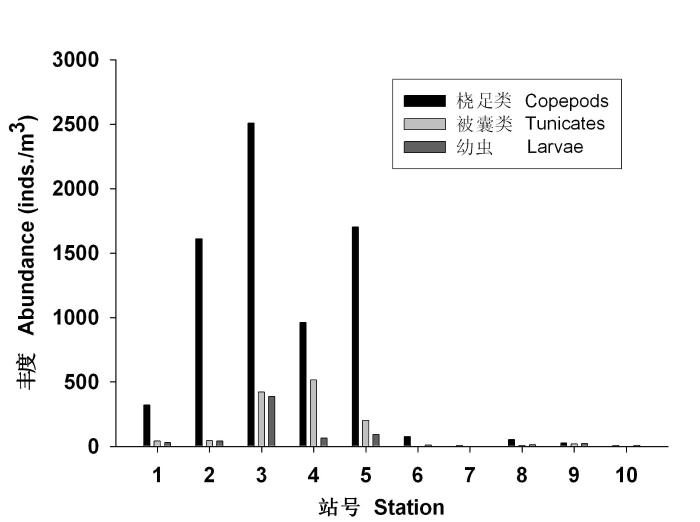
图5
浮游动物主要类群丰度(inds./m3)的分布
Fig. 5
Distribution of abundance (inds./m3) of copepods, tunicates and larvae groups
图6
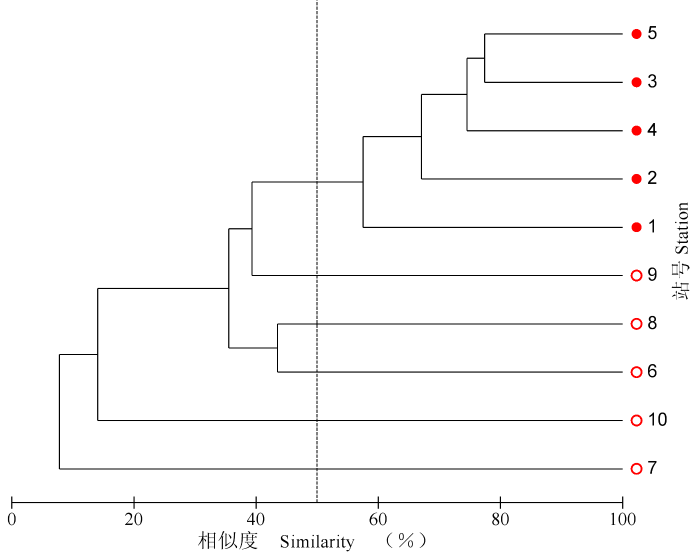
图6
渚碧礁浮游动物群落聚类树枝图(●为潟湖站号; ○为礁坪站号)
Fig. 6
Cluster analysis of the zooplankton community in the Zhubi Atoll
2.4 优势种及其丰度分布特征
从表2可看出两种网具采集的浮游动物优势种既有相似, 也有不同。长尾住囊虫(Oikopleura longicauda)、梭形住囊虫(O. fusifornis)是两种网具都采集到的优势种, 但二者在浅水II型网的丰度均明显大于浅水I型网; 奥氏胸刺水蚤(Centropages orsinii)、珍妮纺锤水蚤属于中小型桡足类, 在浅水II型网的平均丰度很高, 优势度也很明显, 但在浅水I型网样品中的数量非常稀少。优势种在渚碧礁均呈明显的区域分布特征, 在礁坪的出现频率低、数量少, 在澙湖的出现频率高、数量丰富。特别是奥氏胸刺水蚤、珍妮纺锤水蚤在澙湖的平均丰度可达1,082.9 ± 697.3 inds./m3和275.5 ± 222.2 inds./m3, 分别占澙湖区浮游动物总丰度的60.22%和15.32%, 而在礁坪的平均丰度仅为0.8 ± 1.9 inds./m3和15.3 ±21.4 inds./m3。同时, 奥氏胸刺水蚤、珍妮纺锤水蚤在澙湖的数量分布呈群集现象, 最大丰度可分别达2,169 inds./m3和494 inds./m3。
表2 浅水I型和II型网具采集的浮游动物优势种的丰度和优势度
Table 2
| 网具 Planktonic net | 种类 Species | 平均丰度 Abundance (inds./m3) | 占总丰度的百分比 Percentage (%) | 出现频率 Frequency | 优势度 Dominance |
|---|---|---|---|---|---|
| 浅水II型网 Net II | 奥氏胸刺水蚤 Centropages orsinii | 541.86 | 58.52 | 0.6 | 0.351 |
| 珍妮纺锤水蚤 Acartia shuzheni | 145.38 | 15.70 | 0.7 | 0.110 | |
| 长尾住囊虫 Oikopleura longicauda | 76.14 | 8.22 | 0.7 | 0.058 | |
| 梭形住囊虫 O. fusifornis | 45.62 | 4.93 | 0.7 | 0.034 | |
| 腹足类面盘幼虫 Gastropoda veliger | 36.27 | 3.92 | 0.8 | 0.031 | |
| 浅水I型网 Net I | 长尾住囊虫 Oikopleura longicauda | 5.50 | 38.40 | 0.6 | 0.230 |
| 短尾类溞状幼虫 Brachyura zoea larva | 1.28 | 8.93 | 0.6 | 0.054 | |
| 长尾类幼虫 Macrura larva | 1.55 | 10.80 | 0.5 | 0.054 | |
| 红住囊虫 Oikopleura rufescens | 1.96 | 13.70 | 0.3 | 0.041 | |
| 梭形住囊虫 O. fusifornis | 1.13 | 7.88 | 0.3 | 0.024 |
2.5 多样性指数、均匀度和群落结构聚类分析
大面站总体以及澙湖区、礁坪区的多样性指数平均值分别为2.128 ± 0.694、1.934 ± 0.326、2.322 ± 0.939; 均匀度分别为0.647 ± 0.280、0.390 ± 0.048、0.903 ± 0.098, 均低于2002年5月的调查结果(尹健强等, 2003)。从表1可看出, 澙湖区的浮游动物群落由于优势种非常突出, 种间数量分布不均匀, 各站的均匀度值明显低于礁坪区。浅水II型网的均匀度也明显低于浅水I型网。
聚类分析结果表明, 澙湖区的浮游动物明显属于一个单独的群落, 各站的相似度大约在60-80%之间。澙湖区浮游动物群落的特征是种类多、丰度大、优势种突出、均匀度低。礁坪区的浮游动物群落由于既受澙湖, 也受礁外海水的影响, 环境变化大, 各站的相似度低, 大约在10%和50%之间变化。
2.6 连续站浮游动物种类与丰度的昼夜变化
连续站的浮游动物种数和丰度的昼夜差异非常显著, 日间出现的浮游动物种数为16种, 而夜间出现的种数为73种(附录I), 是日间的4.6倍; 日间的浮游动物平均丰度仅为8.79 ± 12.80 inds./m3, 而夜间则高达405.67 ± 239.9 inds./m3, 是日间的46.2倍。白天浮游动物种类和数量都非常贫乏, 正午至午后2:00甚至为零; 在日落后1小时, 即20:00显著上升; 在日落后3小时, 即22:00达到最高峰, 浮游动物的种数和丰度分别高达41种和737.4 inds./m3; 24:00开始逐渐下降, 至次日上午10:00种类和数量变得相当稀少(图7a)。
图7
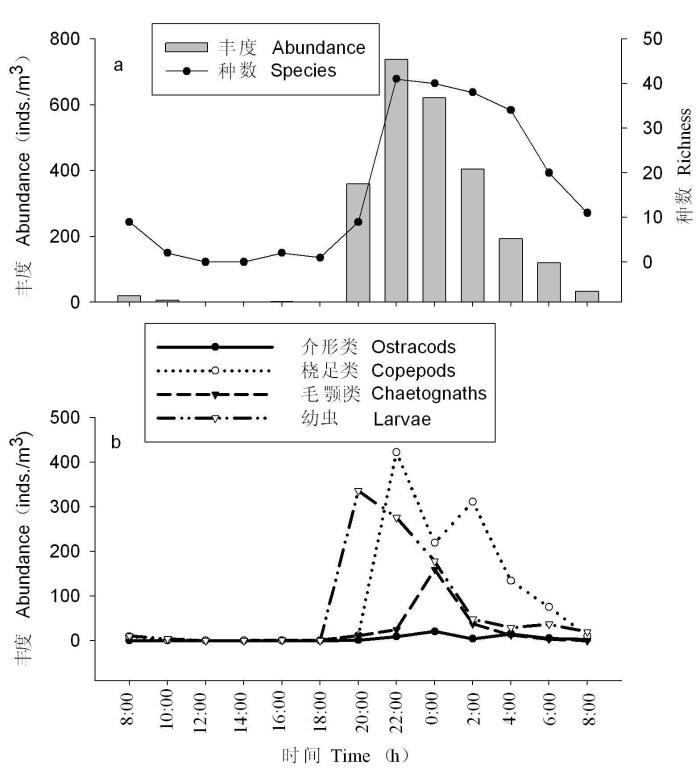
图7
浮游动物种数、总丰度(a)和主要类群丰度(b)的昼夜变化
Fig. 7
Diurnal variations of richness, abundance of zooplankton (a) and main groups (b) at continuous observatory station
浮游动物各主要类群丰度均呈显著的白天下降、夜晚上升的昼夜垂直移动, 但夜晚上升的时间有差异, 底栖动物和游泳动物的幼虫最先上升, 其次是桡足类, 再次是毛颚类和介形类(图7b)。
2.7 两种网具采集样品分析结果的比较
王荣和王克(2003)、尹健强等(2008)在不同海区进行了两种网目孔径大小不同的浮游生物网的捕获性能的对比分析, 发现对于个体较大的种类, 两种网具的结果差异不很显著, 而对于个体较小的种类则差别很大。表3列出两种网具采集的大面站10个站浮游动物总种数、总丰度、多样性指数、均匀度和优势种丰度的差异显著性检验结果。从表中可看出, 浮游动物总种数、总丰度以及奥氏胸刺水蚤、珍妮纺锤水蚤的丰度差异极显著或显著, 而个体较大的种类梭形住囊虫、长尾住囊虫、短尾类溞状幼虫和长尾类幼虫的差异不显著。
表3 两种网具采集样品分析结果的比较(t检验法)
Table 3
| 类别 Variable | 浅水II型网 Net II (169 μm) | 浅水I型网 Net I (505 μm) | 差异性 Significance | |||||
|---|---|---|---|---|---|---|---|---|
| 平均数 Mean | 标准差 SD | 变异系数 CV(%) | 平均数 Mean | 标准差 SD | 变异系数 CV(%) | |||
| 总种数 Richness | 17.7 | 11.5 | 65.0 | 5.1 | 4.0 | 84.4 | ** | |
| 总丰度 Abundance (inds./m3) | 926.0 | 1,155.8 | 124.8 | 14.3 | 15.1 | 105.1 | * | |
| 多样性指数 Diversity index | 2.128 | 0.694 | 32.6 | 1.067 | 1.381 | 77.3 | ns | |
| 均匀度 Evenness index | 0.647 | 0.280 | 43.3 | 0.537 | 0.396 | 73.3 | ns | |
| 优势种丰度(inds./m3) | ||||||||
| 珍妮纺锤水蚤 Acartia shuzheni | 145.4 | 202.4 | 139.2 | 0.2 | 0.3 | 163.1 | * | |
| 奥氏胸刺水蚤 Centropages orsinii | 541.9 | 735.7 | 135.8 | 0.3 | 0.4 | 128.6 | * | |
| 梭形住囊虫 Oikopleura fusifornis | 45.6 | 73.7 | 161.5 | 1.1 | 2.8 | 244.4 | ns | |
| 长尾住囊虫 O. longicauda | 76.1 | 153.5 | 201.6 | 5.5 | 9.8 | 178.4 | ns | |
| 短尾类溞状幼虫 Brachyura zoea larva | 1.0 | 1.6 | 166.0 | 1.3 | 1.4 | 113.1 | ns | |
| 腹足类面盘幼虫 Gastropoda veliger | 36.3 | 83.3 | 229.7 | 0.2 | 0.6 | 316.2 | ns | |
| 长尾类幼虫 Macrura larva | 2.6 | 3.9 | 147.9 | 1.5 | 1.9 | 12.9 | ns | |
3 讨论
3.1 渚碧礁的浮游动物的种类多样性及来源
渚碧礁的浮游动物种类丰富, 本次调查(附录I) 共鉴定浮游动物96种, 加上前人记录过的泡螔螺(Limacina bulimoides)、马蹄螔螺(L. trachiformis)(章淑珍和李纯厚, 1997)、爪室水母(Chelophyes appendiculata)、小哲水蚤(Nannocalanus minor)、小拟哲水蚤(Paracalanus parvus)、太平洋齿箭虫(Serratosagitta pacifica) (尹健强等, 2003)、膨大歪水蚤(Chen et al., 2004), 渚碧礁已经报道的浮游动物种类(除浮游幼虫)已过百种, 这些物种基本上也是南沙群岛及其邻近海区的常见种类(陈清潮, 2003)。 但也许会存在一些珊瑚礁特有种类, 如中华歪水蚤、珍妮纺锤水蚤、膨大歪水蚤等目前仍只见报道出现于珊瑚礁中(陈清潮, 1983, 2008; Chen et al., 2004)。
珊瑚礁的浮游动物主要有两个来源: 一是来自于礁外海水, 即外海类型; 二是来自于珊瑚礁本身, 即礁栖类型(Emery, 1968; Alldredge & King, 1977; Heidelberg et al., 2004, 2010; Nakajima et al., 2009), 但确定浮游动物的来源有些困难, 底栖性浮游动物基本上来自于珊瑚礁, 浮游幼虫等阶段性浮游生物既可来自于礁内, 也可以来自于礁外。终生浮游生物通常认为来自于礁外, 但一些研究(Emery, 1968; Sale et al., 1978; Madhupratap et al., 1991; Heidelberg et al., 2004, 2010)发现纺锤水蚤(Acartia)、胸刺水蚤(Centropages)等传统上归类为外海终生浮游动物的种类也可以改变行为, 适应栖息于珊瑚礁。奥氏胸刺水蚤、珍妮纺锤水蚤和长尾住囊虫(表2)也是2002年5月渚碧礁调查时的优势种(尹健强等, 2003), 表明它们适合栖息于珊瑚礁, 特别是澙湖, 也属于礁栖类型。终生浮游生物在渚碧礁不但种类占多数, 数量上也占优势。
本次调查中, 浮游幼虫无论种类和数量都在群落中占据重要地位, 符合珊瑚礁浮游动物群落特征(Goswami & Goswami, 1990; Carleton & Doherty, 1998)。珊瑚礁生态系统以生物多样性丰富而著称, 复杂多样的生境栖息着种类繁多的营固着、穴居、隐居、爬行、游动以及游泳等各种生活方式的礁栖无脊椎动物和脊椎动物。这些动物的幼虫成为了珊瑚礁浮游动物群落的重要组成部分。李新正等(2007)研究表明渚碧礁的底栖动物的种类和数量都很丰富。
底栖性浮游动物通常被认为是珊瑚礁浮游动物重要的来源之一。它们白天栖息于珊瑚礁基底表面或沉积物中, 而夜晚移动到水柱中, 包括糠虾类、钩虾类、涟虫类、介形类、等足类、猛水蚤类、剑水蚤类、端足类、多毛类等(Alldredge & King, 1977, 1980; Porter & Porter, 1977; Heidelberg et al., 2004)。本次调查中底栖性浮游动物种类和数量都很少, 原因主要是大面站调查均在白天进行, 连续站也只进行了几次的夜晚采集。
3.2 潟湖与礁坪的差异原因分析
尹健强等(2003)对渚碧环礁澙湖与礁坪浮游动物的种类与丰度分布差异作过报道, Alldredge和 King (1977)报道了大堡礁礁坪与澙湖的浮游动物数量差异不显著, 但总体来说这方面的研究报道很少。有关环礁澙湖的浮游动物群落与邻近海区的比较, 在太平洋(Gerber, 1981; Borgne et al., 1989; Carleton & Doherty, 1998)和印度洋(Pratap et al., 1977; Goswami, 1983; Goswami & Goswami, 1990)都有报道。在太平洋面积和水深都较大的环礁, 澙湖内浮游动物的种类组成与邻近开阔的大洋有明显的不同, 数量也较为丰富; 在印度洋面积和水深都较小的环礁, 既有学者认为澙湖与邻近海区有差异(Goswami, 1983; Goswami & Goswami, 1990), 也有学者认为澙湖内不存在特有的浮游动物区系(Pratap et al., 1977), 但都发现澙湖内的浮游动物数量低于邻近海区, 认为是由于浮游动物从邻近海区越过礁坪进入澙湖时被珊瑚礁群落作为食物所消耗。渚碧礁澙湖的浮游动物群落特征与太平洋的环礁更为相似。章淑珍和李纯厚(1997)也报道了南沙群岛的三个环礁: 半月礁、仁爱礁、渚碧礁澙湖的小型浮游动物丰度相当高, 最高可达5,128 inds./m3。
渚碧礁远离大陆, 又位于海洋表层, 温度和盐度空间变化小(图2), 不是影响礁坪和潟湖浮游动物群落空间差异的主要因素。礁坪和潟湖的空间异质性高: 礁坪水浅, 受风浪、海流、潮汐影响大, 水体交换非常快, 而且受太阳的辐射影响也大, 昼夜温差大(图3b); 澙湖水深, 由于被礁坪围封, 受风浪、海流、潮汐影响小, 水体交换相对缓慢, 浮游植物、颗粒有机碳(POC)和颗粒有机氮(PON)也较礁坪丰富(Shen et al., 2010; Yang et al., 2011)。珊瑚黏液、有机碎屑等颗粒有机物同浮游植物一样也是珊瑚礁浮游动物的重要食物来源(Johannes, 1967; Richman et al., 1975; Gerber & Marshall, 1982; Gottfried & Roman, 1983)。因此, 澙湖较为独特的生境形成了与礁坪不同的浮游动物群落。一些学者认为珊瑚礁浮游动物的分布与地形结构有关(Emery, 1968; Heidelberg et al., 2010)。此外, 珊瑚以及鱼类等礁栖动物主要分布于礁坪, 礁坪的浮游动物丰度较低与它们的摄食也有很大的关系。
3.3 两种网具分析结果的比较
中小型浮游动物在渚碧礁具有十分重要的地位, 不但种类多, 而且数量占绝对优势。例如,珍妮纺锤水蚤、隆哲水蚤(Acrocalanus)、丽哲水蚤(Calocalanus)、微刺哲水蚤(Canthocalanus pauper)、奥氏胸刺水蚤、基齿哲水蚤(Clausocalanus)、大眼水蚤(Corycaeus)、双长腹剑水蚤(Dioithona)、尖额谐猛水蚤(Euterpina acutifrons)、羽刺大眼水蚤(Farranula)、瘦长毛猛水蚤(Macrosetella gracilis)、小毛猛水蚤(Microsetella norvegica)、长腹剑水蚤(Oithona)、隆水蚤(Oncaea)、拟哲水蚤(Paraca- lanus)、强额孔雀哲水蚤(Pavocalanus crassirostris)、锥形宽水蚤(Temora turbinata)、三锥水蚤(Triconia)等很多的桡足类的体长基本上在1.50 mm以下(张武昌等, 2010)。此外, 双壳类面盘幼虫、腹足类面盘幼虫、无节幼虫、鸟喙尖头溞(Penilia avirostris)、肥胖三角溞(Pseudevadne tergestina)等种类的个体也非常细小。在浅水II型浮游生物网样品中, 奥氏胸刺水蚤和珍妮纺锤水蚤2个种的丰度即占浮游动物总丰度的74.22%(表2)。
国内早期的珊瑚礁浮游动物研究由于使用网孔较大的浅水I型浮游生物网进行调查, 结果显示珊瑚礁浮游动物种类和数量非常稀少(陈清潮和尹健强, 1982; 陈清潮等, 1989; 钱宏林等, 1990), 低估了浮游动物在珊瑚礁生态系统中的重要性; 后期改用了网孔较小的浅水II型浮游生物网进行调查, 发现珊瑚礁的浮游动物种类和数量都相当丰富(章淑珍和李纯厚, 1997; 尹健强等, 2003, 2008)。因为中小型浮游动物的个体体宽多数小于505 μm, 在使用浅水I型浮游生物网拖网过程中基本上漏掉了。本次调查使用两种网具的采集结果也表明无论浮游动物的总种数、总丰度, 还是主要优势种(奥氏胸刺水蚤和珍妮纺锤水蚤)的丰度都差异显著(表3), 更充分证实了渚碧礁的浮游动物主要由中小型种类所组成。
世界上其他珊瑚礁也有相似的研究结果。在中太平洋的Enewetak环礁的浮游动物群落以小型桡足类以及被囊类的有尾类(Appendiculata, 又称幼虫纲(Larvacea))占优势(Gerber & Marshall, 1982), 同本次调查结果相类似。而在南太平洋的Taiaro环礁由于使用了与浅水I型浮游生物网相类似的网具(网目孔径: 500 μm)进行调查, 浮游动物的种类数和总丰度都很低(Carleton & Doherty, 1998)。在南海的一些岸礁的浮游动物群落也是以中小型的浮游动物占优势(Nakajima et al., 2008, 2009)。Lewis和Boers(1991)报道了在加勒比海的岸礁浮游动物群落中以桡足幼体和小型浮游动物的丰度最大。Hamner和Carleton(1979)观察到了大堡礁澙湖的2种小型桡足类Acartia australis(体长<1.3 mm)和奥氏胸刺水蚤(体长<1.4 mm)的群集现象。珊瑚礁的浮游动物的种类组成和数量不但随时间和空间而改变, 而且研究结果与采样技术有关。因此, 今后在进行珊瑚礁浮游动物调查研究时, 应当选用合适的采样方法。
3.4 珊瑚礁浮游动物的昼夜垂直移动对物种多样性和群落结构的影响
珊瑚礁浮游动物具有昼夜垂直移动行为。本次调查, 渚碧礁礁坪白天的浮游动物种类和数量稀少, 而夜晚明显增加(图7), 与在南海和东南亚(Porter & Porter, 1977; 尹健强等, 2003; Nakajima et al., 2008, 2009)、红海(Yahel et al., 2005a, b)、印度洋(Goswami & Goswami, 1990; Madhupratap et al., 1991)、太平洋(Alldredge & King, 2009)、大堡礁(Alldredge & King, 1977; Hamner & Carleton, 1979)、墨西哥湾和加勒比海(Emery, 1968; Ohlhorst, 1982; Heidelberg et al., 2010)等世界上不同海区的珊瑚礁的研究结果相一致。由此表明, 光照度的昼夜变化是影响珊瑚礁浮游动物垂直移动的主要因素, 浮游动物白天下沉到较深水层, 也可能是躲避强烈的太阳辐射和高温(图3b)。
2002年5月连续站调查(尹健强等, 2003)时的涨落潮时间与本次调查(图3a)相反, 即高潮出现于夜晚, 低潮出现于白天, 但两次调查的浮游动物种类和数量白天减少、夜晚增加的规律没有改变, 表明潮汐变化不会改变浮游动物的昼夜垂直移动节律。
有些研究(Yahel et al., 2005a; Nakajima et al., 2008, 2009)报道, 浮游动物大型种类(>200 μm 或>350 μm)的数量昼夜变化明显, 而小型种类变化不显著。而在渚碧礁浮游动物的昼夜变化与个体大小无关, 不管小型种类如微刺哲水蚤、奥氏胸刺水蚤、弓角基齿哲水蚤(Clausocalanus arcuicornis)、驼背羽刺大眼水蚤(Farranula gibbula)、中隆水蚤(Oncaea media)、矮拟哲水蚤(Paracalanus nanus), 还是大型种类如凶形猛箭虫(Ferosagitta ferox)、正形滨箭虫(Aidanosagitta regularis)、短尾类溞状幼虫、长尾类幼虫、多毛类幼虫等昼夜变化均非常显著。
珊瑚礁浮游动物的昼夜垂直移动对物种多样性和群落结构有相当大的影响: 一是一些终生浮游生物白天下降到较深水层, 夜晚上升到表层可以随水流进入珊瑚礁, 如幼平头水蚤(Candacia catula)、弓角基齿哲水蚤、印度真刺水蚤(Euchaeta indica)、异尾宽水蚤(Temora discaudata)、孟加蛮虫戎(Lestr- igonus bengalensis)、粗壮猛箭虫(Ferosagitta robusta)、太平洋钅毳虫(Krohnitta pacifica)等多种终生浮游生物仅出现于夜晚的样品中, 二是一些底栖性浮游动物白天栖息于底部, 夜晚上升到水柱中, 如星萤、针尾涟虫、文昌鱼等(见附录I)。可见, 珊瑚礁浮游动物群落的种类组成和丰度空间分布格局是随着浮游动物的昼夜垂直移动的变化过程而改变。
致谢:
许战洲、张建林进行温度、盐度观测和叶绿素a水样分析, 谨致谢忱。
附录I 浮游动物种类名录及不同网具、区域以及连续站昼夜出现的种类比较
Appendix I The list of zooplankton species in the Zhubi Atoll and comparison between planktonic net I and II with regions, the appearance of day and night at continuous observatory station
参考文献
Distribution, abundance, and substrate preferences of demersal reef zooplankton at Lizard Island Lagoon, Great Barrier Reef
Effects of moonlight on the vertical migration patterns of demersal zooplankton
Near-surface enrichment of zooplankton over a shallow back reef: implications for coral reef food webs
Zooplankton of Tikehau atoll (Tuamotu archipelago) and its relationship to particulate matter
Tropical zooplankton in the highly-enclosed lagoon of Taiaro Atoll (Tuamotu Archipelago, French Polynesia)
The pelagic copepods of the South China Sea. I
The pelagic copepods of the South China Sea. II
The zooplankton of Huangyan Atoll
The pelagic copepods of the South China Sea. III
A new species Tortanus(Copepoda, Calanoida) from the Nansha Archipelago in the South China Sea
Subclass Copepoda
Prey capture by a benthic coral reef hydrozoan
Preliminary observations on coral reef plankton
Species composition and abundance of lagoon zooplankton at Eniwetak Atoll, Marshall Islands
Characterization of the suspended particulate organic matter and feeding by the lagoon zooplankton at Enewetak Atoll
Production & zooplankton community structure in the lagoon & surrounding sea at Kavaratti Atoll (Lakshadweep)
Diel variation in zooplankton in Minicoy lagoon and Kavaratti atoll (Lakshadweep Islands)
Ingestion and incorporation of coral-mucus detritus by reef zooplankton
Copepod swarms: attributes and role in coral reef ecosystems
Composition and sources of reef zooplankton on a Jamaican forereef along with implications for coral feeding
Vertical distribution and diel patterns of zooplankton abundance and biomass at Conch Reef, Florida Keys (USA)
DOI:10.1093/plankt/fbp101
URL
PMID:20046854
[本文引用: 4]

Zooplankton play an important role in the trophic dynamics of coral reef ecosystems. Detailed vertical and temporal distribution and biomass of zooplankton were evaluated at four heights off the bottom and at six times throughout the diel cycle over a coral reef in the Florida Keys (USA). Zooplankton abundance averaged 4396 +/- 1949 SD individuals m(-3), but temporal and spatial distributions varied for individual zooplankton taxa by time of day and by height off the bottom. Copepods comprised 93-96% of the abundance in the samples. Taxon-based zooplankton CHN values paired with abundance data were used to estimate biomass. Average daily biomass ranged from 3.1 to 21.4 mg C m(-3) and differed by both height off the bottom and by time of day. While copepods were the numerically dominant organisms, their contribution to biomass was only 35% of the total zooplankton biomass. Our findings provide important support for the new emerging paradigm of how zooplankton are distributed over reefs.
Ecology of organic aggregates in the vicinity of a coral reef
The role of zooplankton in the nutrition of some scleractinian corals
Patchiness and composition of coral reef demersal zooplankton
Macrobenthic community characters of Zhubi Reef, Nansha Islands, South China Sea
Weather climatic features of Nansha Islands and neighbouring sections
Zooplankton of the lagoons of the Laccadives: diel patterns and emergence
Diel variation in abundance, biomass and size composition of zooplankton community over a coral-reef in Redang Island, Malaysia
Diel variation of zooplankton in the tropical coral-reef water of Tioman Island, Malaysia
DOI:10.1007/s10452-008-9208-5
URL
[本文引用: 5]
Zooplankton was sampled at 3-h intervals for a 48-h period from a coral reef of Tioman Island, Malaysia. It was size-fractionated into three size classes: 100–200, 200–335, and >335 μm using different sieves with different mesh sizes. Total zooplankton (>100 μm) abundance and biomass in the water column were high later at night (0300 h), not just after sunset as previously described in other studies. Only the largest size-fraction (>335 μm) of zooplankton significantly differed in biomass and abundance between day and night. The increase in the large zooplankton later in the night is suggested to be caused by the advection of pelagic species into the reef. This work has provided a measurement of the variation of zooplankton community over coral reef that can exist on a scale of hours.]]>
Diel migration patterns of demersal reef zooplankton
Quantitative sampling of demersal plankton migrating from different coral reef substrates
Comparative studies on the abundance of zooplankton in the surrounding sea & lagoons in the Lakshadweep
Zooplankton in the Huangyan Atoll and adjacent waters
The rate of mucus production by corals and its assimilation by the coral reef copepod Acartia negligens
Faunal relationships among the near-reef zooplankton at three locations on Heron Reef, Great Barrier Reef, and seasonal changes in this fauna
Zooplankton capture by two scleractinian corals,Madracis mirabilis and Montastrea cavernosa, in a field enclosure
A mathematical theory of communication
Occurrence of brackish water phytoplankton species at a closed coral reef in Nansha Islands, South China Sea
Field test of capture capabilities of two plankton nets
Aggregated inten-sity of dominant species of zooplankton in autumn in the East China Sea and Yellow Sea
Near-bottom depletion of zooplankton over coral reefs. I. Diurnal dynamics and size distribution
Diel pattern with abrupt crepuscular changes of zooplankton over a coral reef
Carbon and nitrogen cycling in the Zhubi coral reef lagoon of the South China Sea as revealed by 210Po and 210Pb
Zooplanktonic community characteristics in waters of Zhubi coral reef (Nansha Islands) in spring
Summer zooplankton in coral reef area of Leizhou Peninsula, China
Diurnal vertical migration of chaetognath around Nansha Islands waters, southern South China Sea
The nutrition role of microzooplankton in the lagoon ecosystem from coral reefs of the Nansha Islands

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}